
Landscape of Noncoding RNA in the Hypoxic Tumor Microenvironment

Abstract

1. Introduction
2. LncRNAs and circRNAs Act as Novel Players in the Hypoxic Tumor Microenvironment
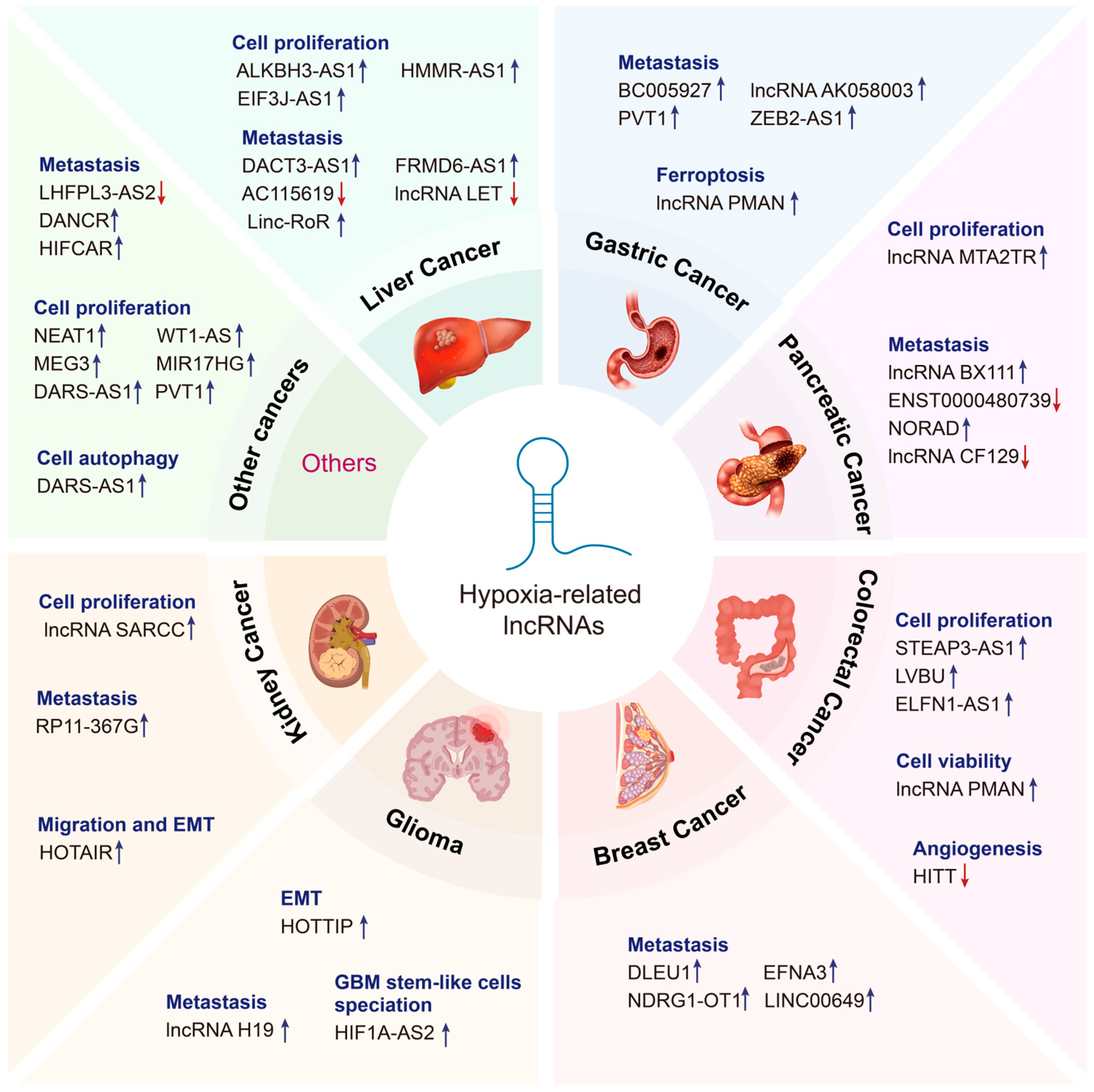
3. Hypoxia-Related lncRNAs in Cancers
3.1. Gastrointestinal Cancers
3.1.1. Liver Cancer
3.1.2. Gastric Cancer
3.1.3. Pancreatic Cancer
3.1.4. Colorectal Cancer
3.2. Breast Cancer
3.3. Glioma
3.4. Renal Cell Carcinoma
3.5. Other Cancers
4. Hypoxia-Related circRNAs in Cancers
4.1. Gastrointestinal Cancers
4.1.1. Liver Cancer
4.1.2. Colorectal Cancer
4.1.3. Pancreatic Cancer
4.1.4. Gastric Cancer
4.1.5. Other Gastrointestinal Cancers
4.2. Breast Cancer
4.3. Lung Cancer
4.4. Cervical Cancer
4.5. Glioma
4.6. Other Cancers
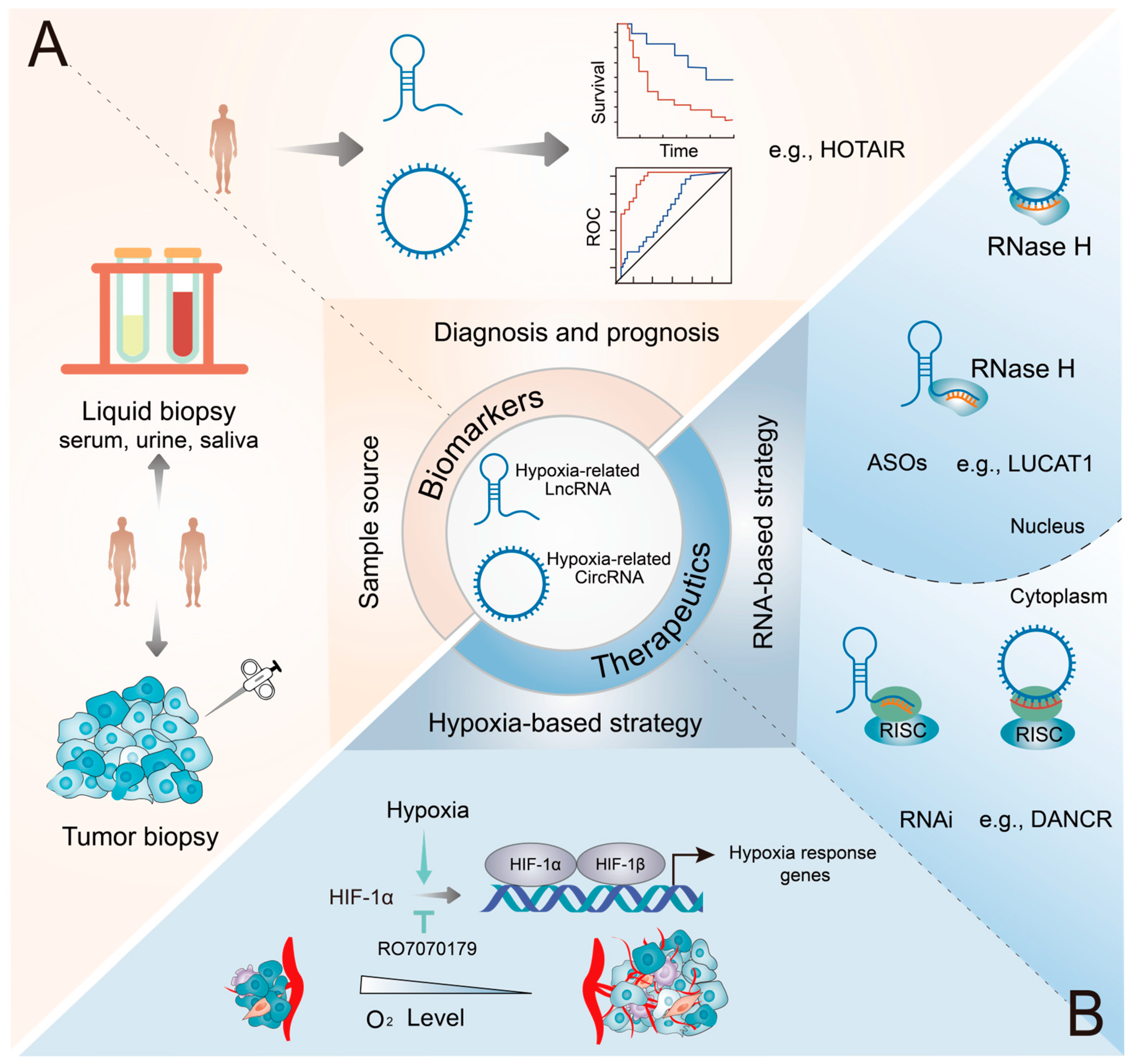
5. Clinical Relevance of Hypoxia-Related lncRNAs and circRNAs in Cancers
5.1. Biomarker Potentials
5.2. Therapeutic Potential
6. Concluding Remarks and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Casey, S.C.; Amedei, A.; Aquilano, K.; Azmi, A.S.; Benencia, F.; Bhakta, D.; Bilsland, A.E.; Boosani, C.S.; Chen, S.; Ciriolo, M.R.; et al. Cancer prevention and therapy through the modulation of the tumor microenvironment. Semin. Cancer Biol. 2015, 35, S199–S223. [Google Scholar] [CrossRef] [PubMed]
- Rankin, E.B.; Giaccia, A.J. Hypoxic control of metastasis. Science 2016, 352, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Choudhry, H.; Harris, A.L.; McIntyre, A. The tumour hypoxia induced non-coding transcriptome. Mol. Asp. Med. 2016, 47–48, 35–53. [Google Scholar] [CrossRef]
- Shih, J.W.; Kung, H.J. Long non-coding RNA and tumor hypoxia: New players ushered toward an old arena. J. Biomed. Sci. 2017, 24, 53. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, K.; Bayraktar, R.; Ferracin, M.; Calin, G.A. Non-coding RNAs in disease: From mechanisms to therapeutics. Nat. Rev. Genet. 2024, 25, 211–232. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, J.; Li, Y.; Song, T.; Wu, Y.; Fang, S.; Bu, D.; Li, H.; Sun, L.; Pei, D.; et al. NONCODEV6: An updated database dedicated to long non-coding RNA annotation in both animals and plants. Nucleic Acids Res. 2021, 49, D165–D171. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhao, F.; Zhang, J. circAtlas 3.0: A gateway to 3 million curated vertebrate circular RNAs based on a standardized nomenclature scheme. Nucleic Acids Res. 2024, 52, D52–D60. [Google Scholar] [CrossRef] [PubMed]
- Bouchard-Bourelle, P.; Desjardins-Henri, C.; Mathurin-St-Pierre, D.; Deschamps-Francoeur, G.; Fafard-Couture, É.; Garant, J.M.; Elela, S.A.; Scott, M.S. snoDB: An interactive database of human snoRNA sequences, abundance and interactions. Nucleic Acids Res. 2020, 48, D220–D225. [Google Scholar] [CrossRef]
- Chandra Gupta, S.; Nandan Tripathi, Y. Potential of long non-coding RNAs in cancer patients: From biomarkers to therapeutic targets. Int. J. Cancer 2017, 140, 1955–1967. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.M.; Chang, H.Y. Long Noncoding RNAs in Cancer Pathways. Cancer Cell 2016, 29, 452–463. [Google Scholar] [CrossRef] [PubMed]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef]
- Rajappa, A.; Banerjee, S.; Sharma, V.; Khandelia, P. Circular RNAs: Emerging Role in Cancer Diagnostics and Therapeutics. Front. Mol. Biosci. 2020, 7, 577938. [Google Scholar] [CrossRef]
- Shen, G.; Li, X.; Jia, Y.F.; Piazza, G.A.; Xi, Y. Hypoxia-regulated microRNAs in human cancer. Acta Pharmacol. Sin. 2013, 34, 336–341. [Google Scholar] [CrossRef]
- Xiong, Q.; Zhang, Y.; Li, J.; Zhu, Q. Small Non-Coding RNAs in Human Cancer. Genes 2022, 13, 2072. [Google Scholar] [CrossRef] [PubMed]
- Chakrabortty, A.; Patton, D.J.; Smith, B.F.; Agarwal, P. miRNAs: Potential as Biomarkers and Therapeutic Targets for Cancer. Genes 2023, 14, 1375. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Bao, Z.; Yang, Z.; Huang, Z.; Zhou, Y.; Cui, Q.; Dong, D. LncRNADisease 2.0: An updated database of long non-coding RNA-associated diseases. Nucleic Acids Res. 2019, 47, D1034–D1037. [Google Scholar] [CrossRef] [PubMed]
- Coan, M.; Haefliger, S.; Ounzain, S.; Johnson, R. Targeting and engineering long non-coding RNAs for cancer therapy. Nat. Rev. Genet. 2024, 25, 578–595. [Google Scholar] [CrossRef] [PubMed]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef]
- Pandya, G.; Kirtonia, A.; Sethi, G.; Pandey, A.K.; Garg, M. The implication of long non-coding RNAs in the diagnosis, pathogenesis and drug resistance of pancreatic ductal adenocarcinoma and their possible therapeutic potential. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188423. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Jakobsen, T.; Hager, H.; Kjems, J. The emerging roles of circRNAs in cancer and oncology. Nat. Rev. Clin. Oncol. 2022, 19, 188–206. [Google Scholar] [CrossRef]
- Pisignano, G.; Michael, D.C.; Visal, T.H.; Pirlog, R.; Ladomery, M.; Calin, G.A. Going circular: History, present, and future of circRNAs in cancer. Oncogene 2023, 42, 2783–2800. [Google Scholar] [CrossRef]
- Heo, J.B.; Sung, S. Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA. Science 2011, 331, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.L.; Wang, Y.; et al. Extensive translation of circular RNAs driven by N(6)-methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Anderson, K.M.; Chang, C.L.; Makarewich, C.A.; Nelson, B.R.; McAnally, J.R.; Kasaragod, P.; Shelton, J.M.; Liou, J.; Bassel-Duby, R.; et al. A micropeptide encoded by a putative long noncoding RNA regulates muscle performance. Cell 2015, 160, 595–606. [Google Scholar] [CrossRef]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 Is a Circular RNA that Can Be Translated and Functions in Myogenesis. Mol. Cell 2017, 66, 22–37.e29. [Google Scholar] [CrossRef]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Li Santi, A.; Gorrasi, A.; Alfieri, M.; Montuori, N.; Ragno, P. A novel oncogenic role for urokinase receptor in leukemia cells: Molecular sponge for oncosuppressor microRNAs. Oncotarget 2018, 9, 27823–27834. [Google Scholar] [CrossRef] [PubMed]
- Iyer, M.K.; Niknafs, Y.S.; Malik, R.; Singhal, U.; Sahu, A.; Hosono, Y.; Barrette, T.R.; Prensner, J.R.; Evans, J.R.; Zhao, S.; et al. The landscape of long noncoding RNAs in the human transcriptome. Nat. Genet. 2015, 47, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Guarnerio, J.; Bezzi, M.; Jeong, J.C.; Paffenholz, S.V.; Berry, K.; Naldini, M.M.; Lo-Coco, F.; Tay, Y.; Beck, A.H.; Pandolfi, P.P. Oncogenic Role of Fusion-circRNAs Derived from Cancer-Associated Chromosomal Translocations. Cell 2016, 165, 289–302. [Google Scholar] [CrossRef]
- Kanojia, D.; Kirtonia, A.; Srujana, N.S.V.; Jeevanandan, S.P.; Shyamsunder, P.; Sampath, S.S.; Dakle, P.; Mayakonda, A.; Kaur, H.; Yanyi, J.; et al. Transcriptome analysis identifies TODL as a novel lncRNA associated with proliferation, differentiation, and tumorigenesis in liposarcoma through FOXM1. Pharmacol. Res. 2022, 185, 106462. [Google Scholar] [CrossRef]
- Dinescu, S.; Ignat, S.; Lazar, A.D.; Constantin, C.; Neagu, M.; Costache, M. Epitranscriptomic Signatures in lncRNAs and Their Possible Roles in Cancer. Genes 2019, 10, 52. [Google Scholar] [CrossRef]
- Shao, C.; Yang, F.; Miao, S.; Liu, W.; Wang, C.; Shu, Y.; Shen, H. Role of hypoxia-induced exosomes in tumor biology. Mol. Cancer 2018, 17, 120. [Google Scholar] [CrossRef] [PubMed]
- Cowman, S.J.; Koh, M.Y. Revisiting the HIF switch in the tumor and its immune microenvironment. Trends Cancer 2022, 8, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Defining the role of hypoxia-inducible factor 1 in cancer biology and therapeutics. Oncogene 2010, 29, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Moreno Roig, E.; Yaromina, A.; Houben, R.; Groot, A.J.; Dubois, L.; Vooijs, M. Prognostic Role of Hypoxia-Inducible Factor-2α Tumor Cell Expression in Cancer Patients: A Meta-Analysis. Front. Oncol. 2018, 8, 224. [Google Scholar] [CrossRef]
- Choudhry, H.; Harris, A.L. Advances in Hypoxia-Inducible Factor Biology. Cell Metab. 2018, 27, 281–298. [Google Scholar] [CrossRef] [PubMed]
- Kaelin, W.G., Jr.; Ratcliffe, P.J. Oxygen sensing by metazoans: The central role of the HIF hydroxylase pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, W.; Zhou, Q.; Chen, C.; Yuan, W.; Liu, J.; Li, X.; Sun, Z. Roles of circRNAs in the tumour microenvironment. Mol. Cancer 2020, 19, 14. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, D.; Xie, H.; Hu, Y. Interplay of long non-coding RNAs and HIF-1α: A new dimension to understanding hypoxia-regulated tumor growth and metastasis. Cancer Lett. 2021, 499, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Jiao, B.; Liu, S.; Zhao, H.; Zhuang, Y.; Ma, S.; Lin, C.; Hu, J.; Liu, X. Hypoxia-responsive circRNAs: A novel but important participant in non-coding RNAs ushered toward tumor hypoxia. Cell Death Dis. 2022, 13, 666. [Google Scholar] [CrossRef]
- Wang, L.; Li, B.; Bo, X.; Yi, X.; Xiao, X.; Zheng, Q. Hypoxia-induced LncRNA DACT3-AS1 upregulates PKM2 to promote metastasis in hepatocellular carcinoma through the HDAC2/FOXA3 pathway. Exp. Mol. Med. 2022, 54, 848–860. [Google Scholar] [CrossRef]
- Lu, Q.; Wang, H.; Lei, X.; Ma, Q.; Zhao, J.; Sun, W.; Guo, C.; Huang, D.; Xu, Q. LncRNA ALKBH3-AS1 enhances ALKBH3 mRNA stability to promote hepatocellular carcinoma cell proliferation and invasion. J. Cell Mol. Med. 2022, 26, 5292–5302. [Google Scholar] [CrossRef]
- Zhang, Q.; Wei, T.; Yan, L.; Zhu, S.; Jin, W.; Bai, Y.; Zeng, Y.; Zhang, X.; Yin, Z.; Yang, J.; et al. Hypoxia-Responsive lncRNA AC115619 Encodes a Micropeptide That Suppresses m6A Modifications and Hepatocellular Carcinoma Progression. Cancer Res. 2023, 83, 2496–2512. [Google Scholar] [CrossRef]
- Yang, F.; Huo, X.S.; Yuan, S.X.; Zhang, L.; Zhou, W.P.; Wang, F.; Sun, S.H. Repression of the long noncoding RNA-LET by histone deacetylase 3 contributes to hypoxia-mediated metastasis. Mol. Cell 2013, 49, 1083–1096. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Lei, X.; Lu, Q.; Wu, Q.; Ma, Q.; Huang, D.; Zhang, Y. LncRNA FRMD6-AS1 promotes hepatocellular carcinoma cell migration and stemness by regulating SENP1/HIF-1alpha axis. Pathol. Res. Pract. 2023, 243, 154377. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yan, I.K.; Haga, H.; Patel, T. Modulation of hypoxia-signaling pathways by extracellular linc-RoR. J. Cell Sci. 2014, 127, 1585–1594. [Google Scholar] [CrossRef]
- Yang, X.; Yao, B.; Niu, Y.; Chen, T.; Mo, H.; Wang, L.; Guo, C.; Yao, D. Hypoxia-induced lncRNA EIF3J-AS1 accelerates hepatocellular carcinoma progression via targeting miR-122-5p/CTNND2 axis. Biochem. Biophys. Res. Commun. 2019, 518, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, Y.; Dong, K.; Zhang, H.; Gong, J.; Wang, S. Exosomal lncRNA HMMR-AS1 mediates macrophage polarization through miR-147a/ARID3A axis under hypoxia and affects the progression of hepatocellular carcinoma. Environ. Toxicol. 2022, 37, 1357–1372. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Song, J.; Gao, Y.; Huang, S.; Dou, R.; Zhong, P.; Huang, G.; Han, L.; Zheng, J.; Zhang, X.; et al. Hypoxia-induced HIF-1alpha/lncRNA-PMAN inhibits ferroptosis by promoting the cytoplasmic translocation of ELAVL1 in peritoneal dissemination from gastric cancer. Redox Biol. 2022, 52, 102312. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Y.; Sun, L.; Min, J.; Liu, J.; Chen, D.; Zhang, H.; Zhang, H.; Zhang, H.; Zhou, Y.; et al. Long noncoding RNA BC005927 upregulates EPHB4 and promotes gastric cancer metastasis under hypoxia. Cancer Sci. 2018, 109, 988–1000. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.; Zhang, H.; Sun, L.; Zhou, Y.; Jin, H.; Zhang, H.; Zhang, H.; Liu, J.; Guo, H.; et al. Hypoxia-inducible lncRNA-AK058003 promotes gastric cancer metastasis by targeting γ-synuclein. Neoplasia 2014, 16, 1094–1106. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Liu, H.W.; Chen, J.Q.; Wang, S.H.; Hao, L.Q.; Liu, M.; Wang, B. The long noncoding RNA PVT1 functions as a competing endogenous RNA by sponging miR-186 in gastric cancer. Biomed. Pharmacother. 2017, 88, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Gao, H.; Liu, K.; Gao, B.; Ren, H.; Li, Z.; Liu, F. The lncRNA ZEB2-AS1 is upregulated in gastric cancer and affects cell proliferation and invasion via miR-143-5p/HIF-1alpha axis. Onco Targets Ther. 2019, 12, 657–667. [Google Scholar] [CrossRef]
- Liu, M.; Zhong, J.; Zeng, Z.; Huang, K.; Ye, Z.; Deng, S.; Chen, H.; Xu, F.; Li, Q.; Zhao, G. Hypoxia-induced feedback of HIF-1alpha and lncRNA-CF129 contributes to pancreatic cancer progression through stabilization of p53 protein. Theranostics 2019, 9, 4795–4810. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, X.; Wen, C.; Huo, Z.; Wang, W.; Zhan, Q.; Cheng, D.; Chen, H.; Deng, X.; Peng, C.; et al. Long noncoding RNA NORAD, a novel competing endogenous RNA, enhances the hypoxia-induced epithelial-mesenchymal transition to promote metastasis in pancreatic cancer. Mol. Cancer 2017, 16, 169. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Xu, F.Y.; Zheng, H.; Cheng, P.; Chen, Q.Y.; Ye, Z.; Zhong, J.X.; Deng, S.J.; Liu, M.L.; Huang, K.; et al. LncRNA-MTA2TR functions as a promoter in pancreatic cancer via driving deacetylation-dependent accumulation of HIF-1alpha. Theranostics 2019, 9, 5298–5314. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.J.; Chen, H.Y.; Ye, Z.; Deng, S.C.; Zhu, S.; Zeng, Z.; He, C.; Liu, M.L.; Huang, K.; Zhong, J.X.; et al. Hypoxia-induced LncRNA-BX111 promotes metastasis and progression of pancreatic cancer through regulating ZEB1 transcription. Oncogene 2018, 37, 5811–5828. [Google Scholar] [CrossRef]
- Sun, Y.W.; Chen, Y.F.; Li, J.; Huo, Y.M.; Liu, D.J.; Hua, R.; Zhang, J.F.; Liu, W.; Yang, J.Y.; Fu, X.L.; et al. A novel long non-coding RNA ENST00000480739 suppresses tumour cell invasion by regulating OS-9 and HIF-1alpha in pancreatic ductal adenocarcinoma. Br. J. Cancer 2014, 111, 2131–2141. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, L.; Zhao, K.; Lin, Q.; Li, H.; Xue, X.; Ge, W.; He, H.; Liu, D.; Xie, H.; et al. A novel LncRNA HITT forms a regulatory loop with HIF-1alpha to modulate angiogenesis and tumor growth. Cell Death Differ. 2020, 27, 1431–1446. [Google Scholar] [CrossRef] [PubMed]
- Huan, L.; Guo, T.; Wu, Y.; Xu, L.; Huang, S.; Xu, Y.; Liang, L.; He, X. Hypoxia induced LUCAT1/PTBP1 axis modulates cancer cell viability and chemotherapy response. Mol. Cancer 2020, 19, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Jiang, J.; Huang, Z.; Jin, P.; Peng, L.; Luo, M.; Zhang, Z.; Chen, Y.; Xie, N.; Gao, W.; et al. Hypoxia-induced lncRNA STEAP3-AS1 activates Wnt/β-catenin signaling to promote colorectal cancer progression by preventing m(6)A-mediated degradation of STEAP3 mRNA. Mol. Cancer 2022, 21, 168. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Peng, J.; Xie, X.; Yu, F.; Wang, W.; Pan, Q.; Jin, H.; Huang, X.; Yu, H.; Li, S.; et al. Roles of lncRNA LVBU in regulating urea cycle/polyamine synthesis axis to promote colorectal carcinoma progression. Oncogene 2022, 41, 4231–4243. [Google Scholar] [CrossRef]
- Jing, X.; Du, L.; Shi, S.; Niu, A.; Wu, J.; Wang, Y.; Wang, C. Hypoxia-Induced Upregulation of lncRNA ELFN1-AS1 Promotes Colon Cancer Growth and Metastasis Through Targeting TRIM14 via Sponging miR-191-5p. Front. Pharmacol. 2022, 13, 806682. [Google Scholar] [CrossRef]
- Ma, H.N.; Chen, H.J.; Liu, J.Q.; Li, W.T. Long non-coding RNA DLEU1 promotes malignancy of breast cancer by acting as an indispensable coactivator for HIF-1alpha-induced transcription of CKAP2. Cell Death Dis. 2022, 13, 625. [Google Scholar] [CrossRef]
- Zhang, J.; Du, C.; Zhang, L.; Wang, Y.; Zhang, Y.; Li, J. LncRNA LINC00649 promotes the growth and metastasis of triple-negative breast cancer by maintaining the stability of HIF-1alpha through the NF90/NF45 complex. Cell Cycle 2022, 21, 1034–1047. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Maldonado, L.; Tiana, M.; Roche, O.; Prado-Cabrero, A.; Jensen, L.; Fernandez-Barral, A.; Guijarro-Munoz, I.; Favaro, E.; Moreno-Bueno, G.; Sanz, L.; et al. EFNA3 long noncoding RNAs induced by hypoxia promote metastatic dissemination. Oncogene 2015, 34, 2609–2620. [Google Scholar] [CrossRef] [PubMed]
- Chao, H.H.; Luo, J.L.; Hsu, M.H.; Chen, L.H.; Lu, T.P.; Tsai, M.H.; Chuang, E.Y.; Chuang, L.L.; Lai, L.C. Regulatory mechanisms and function of hypoxia-induced long noncoding RNA NDRG1-OT1 in breast cancer cells. Cell Death Dis. 2022, 13, 807. [Google Scholar] [CrossRef]
- Mineo, M.; Ricklefs, F.; Rooj, A.K.; Lyons, S.M.; Ivanov, P.; Ansari, K.I.; Nakano, I.; Chiocca, E.A.; Godlewski, J.; Bronisz, A. The Long Non-coding RNA HIF1A-AS2 Facilitates the Maintenance of Mesenchymal Glioblastoma Stem-like Cells in Hypoxic Niches. Cell Rep. 2016, 15, 2500–2509. [Google Scholar] [CrossRef]
- Wu, W.; Hu, Q.; Nie, E.; Yu, T.; Wu, Y.; Zhi, T.; Jiang, K.; Shen, F.; Wang, Y.; Zhang, J.; et al. Hypoxia induces H19 expression through direct and indirect Hif-1alpha activity, promoting oncogenic effects in glioblastoma. Sci. Rep. 2017, 7, 45029. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, W.; Liu, G.; Xie, S.; Li, Q.; Li, Y.; Lin, Z. Long non-coding RNA HOTTIP promotes hypoxia-induced epithelial-mesenchymal transition of malignant glioma by regulating the miR-101/ZEB1 axis. Biomed. Pharmacother. 2017, 95, 711–720. [Google Scholar] [CrossRef]
- Zhai, W.; Sun, Y.; Jiang, M.; Wang, M.; Gasiewicz, T.A.; Zheng, J.; Chang, C. Differential regulation of LncRNA-SARCC suppresses VHL-mutant RCC cell proliferation yet promotes VHL-normal RCC cell proliferation via modulating androgen receptor/HIF-2alpha/C-MYC axis under hypoxia. Oncogene 2016, 35, 4866–4880. [Google Scholar] [CrossRef] [PubMed]
- Shao, I.H.; Peng, P.H.; Wu, H.H.; Chen, J.L.; Lai, J.C.; Chang, J.S.; Wu, H.T.; Wu, K.J.; Pang, S.T.; Hsu, K.W. RP11-367G18.1 V2 enhances clear cell renal cell carcinoma progression via induction of epithelial-mesenchymal transition. Cancer Med. 2023, 12, 9788–9801. [Google Scholar] [CrossRef] [PubMed]
- Hong, Q.; Li, O.; Zheng, W.; Xiao, W.Z.; Zhang, L.; Wu, D.; Cai, G.Y.; He, J.C.; Chen, X.M. LncRNA HOTAIR regulates HIF-1alpha/AXL signaling through inhibition of miR-217 in renal cell carcinoma. Cell Death Dis. 2017, 8, e2772. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, W.; Lian, J.; Zhang, H.; Yu, B.; Zhang, M.; Wei, F.; Wu, J.; Jiang, J.; Jia, Y.; et al. The lncRNA PVT1 regulates nasopharyngeal carcinoma cell proliferation via activating the KAT2A acetyltransferase and stabilizing HIF-1alpha. Cell Death Differ. 2020, 27, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Liu, X.; Mao, Y.P.; Yang, X.J.; Wang, Y.Q.; Zhang, P.P.; Lei, Y.; Hong, X.H.; He, Q.M.; Ma, J.; et al. Long non-coding RNA DANCR stabilizes HIF-1alpha and promotes metastasis by interacting with NF90/NF45 complex in nasopharyngeal carcinoma. Theranostics 2018, 8, 5676–5689. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Zhao, Y.; Li, X.; Tao, Z.; Hou, M.; Ma, H. Overexpression of HIF-2alpha-Dependent NEAT1 Promotes the Progression of Non-Small Cell Lung Cancer through miR-101-3p/SOX9/Wnt/β-Catenin Signal Pathway. Cell Physiol. Biochem. 2019, 52, 368–381. [Google Scholar] [CrossRef]
- Cheng, Z.; Lu, C.; Wang, H.; Wang, N.; Cui, S.; Yu, C.; Wang, C.; Zuo, Q.; Wang, S.; Lv, Y.; et al. Long noncoding RNA LHFPL3-AS2 suppresses metastasis of non-small cell lung cancer by interacting with SFPQ to regulate TXNIP expression. Cancer Lett. 2022, 531, 1–13. [Google Scholar] [CrossRef]
- Yan, J.; Deng, Y.X.; Cai, Y.L.; Cong, W.D. LncRNA MIR17HG promotes the proliferation, migration, and invasion of retinoblastoma cells by up-regulating HIF-1alpha expression via sponging miR-155-5p. Kaohsiung J. Med. Sci. 2022, 38, 554–564. [Google Scholar] [CrossRef]
- Shih, J.W.; Chiang, W.F.; Wu, A.T.H.; Wu, M.H.; Wang, L.Y.; Yu, Y.L.; Hung, Y.W.; Wang, W.C.; Chu, C.Y.; Hung, C.L.; et al. Long noncoding RNA LncHIFCAR/MIR31HG is a HIF-1alpha co-activator driving oral cancer progression. Nat. Commun. 2017, 8, 15874. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, H.; Mei, Y.; Wu, M. Reciprocal regulation of HIF-1alpha and lincRNA-p21 modulates the Warburg effect. Mol. Cell 2014, 53, 88–100. [Google Scholar] [CrossRef]
- Shen, W.; Zhu, M.; Wang, Q.; Zhou, X.; Wang, J.; Wang, T.; Zhang, J. DARS-AS1 recruits METTL3/METTL14 to bind and enhance DARS mRNA m(6)A modification and translation for cytoprotective autophagy in cervical cancer. RNA Biol. 2022, 19, 751–763. [Google Scholar] [CrossRef]
- Tong, J.; Xu, X.; Zhang, Z.; Ma, C.; Xiang, R.; Liu, J.; Xu, W.; Wu, C.; Li, J.; Zhan, F.; et al. Hypoxia-induced long non-coding RNA DARS-AS1 regulates RBM39 stability to promote myeloma malignancy. Haematologica 2020, 105, 1630–1640. [Google Scholar] [CrossRef]
- McCarty, G.; Loeb, D.M. Hypoxia-sensitive epigenetic regulation of an antisense-oriented lncRNA controls WT1 expression in myeloid leukemia cells. PLoS ONE 2015, 10, e0119837. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.Y.; Lei, B.X.; Lin, Y.Y.; Sui, M.H.; Zhang, M.L.; Zhuang, Z.Q.; Dong, J.T.; Jin, D.M.; Yan, T.B. Long noncoding RNA MEG3 silencing protects against hypoxia-induced pheochromocytoma-12 cell injury through inhibition of TIMP2 promoter methylation. J. Cell Physiol. 2020, 235, 1649–1662. [Google Scholar] [CrossRef]
- Marengo, A.; Rosso, C.; Bugianesi, E. Liver Cancer: Connections with Obesity, Fatty Liver, and Cirrhosis. Annu. Rev. Med. 2016, 67, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Smyth, E.C.; Nilsson, M.; Grabsch, H.I.; van Grieken, N.C.; Lordick, F. Gastric cancer. Lancet 2020, 396, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Hill, H.; Christie, A.; Kim, M.S.; Holloman, E.; Pavia-Jimenez, A.; Homayoun, F.; Ma, Y.; Patel, N.; Yell, P.; et al. Targeting renal cell carcinoma with a HIF-2 antagonist. Nature 2016, 539, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.; Herman, J.; Schulick, R.; Hruban, R.H.; Goggins, M. Pancreatic cancer. Lancet 2011, 378, 607–620. [Google Scholar] [CrossRef]
- Chen, X.; Chen, S. LINC00649 promotes bladder cancer malignant progression by regulating the miR-15a-5p/HMGA1 axis. Oncol. Rep. 2021, 45, 8. [Google Scholar] [CrossRef] [PubMed]
- Si, C.; Yang, L.; Cai, X. LncRNA LINC00649 aggravates the progression of cervical cancer through sponging miR-216a-3p. J. Obstet. Gynaecol. Res. 2022, 48, 2853–2862. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Hakami, M.A.; Hazazi, A.; Abdulaziz, O.; Almasoudi, H.H.; Alhazmi, A.Y.M.; Alkhalil, S.S.; Alharthi, N.S.; Alhuthali, H.M.; Almalki, W.H.; Gupta, G.; et al. HOTAIR: A key regulator of the Wnt/β-catenin signaling cascade in cancer progression and treatment. Pathol. Res. Pract. 2024, 253, 154957. [Google Scholar] [CrossRef] [PubMed]
- Morelli, E.; Fulciniti, M.; Samur, M.K.; Ribeiro, C.F.; Wert-Lamas, L.; Henninger, J.E.; Gullà, A.; Aktas-Samur, A.; Todoerti, K.; Talluri, S.; et al. A MIR17HG-derived long noncoding RNA provides an essential chromatin scaffold for protein interaction and myeloma growth. Blood 2023, 141, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Hansen, T.B.; Veno, M.T.; Kjems, J. Circular RNAs in cancer: Opportunities and challenges in the field. Oncogene 2018, 37, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. HIF-1 mediates metabolic responses to intratumoral hypoxia and oncogenic mutations. J. Clin. Investig. 2013, 123, 3664–3671. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Wang, M.; Dong, Y.; Xu, B.; Chen, J.; Ding, Y.; Qiu, S.; Li, L.; Karamfilova Zaharieva, E.; Zhou, X.; et al. Circular RNA circRNF20 promotes breast cancer tumorigenesis and Warburg effect through miR-487a/HIF-1alpha/HK2. Cell Death Dis. 2020, 11, 145. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, F.; Xiong, Y.; Wang, N.; Gu, Y.; Qiu, X. CircZFR functions as a sponge of miR-578 to promote breast cancer progression by regulating HIF1A expression. Cancer Cell Int. 2020, 20, 400. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, M.; Yu, D.; Hou, G.; Wu, J.; Li, F. CircRBM33 downregulation inhibits hypoxia-induced glycolysis and promotes apoptosis of breast cancer cells via a microRNA-542-3p/HIF-1alpha axis. Cell Death Discov. 2022, 8, 126. [Google Scholar] [CrossRef]
- Liang, G.; Liu, Z.; Tan, L.; Su, A.N.; Jiang, W.G.; Gong, C. HIF1alpha-associated circDENND4C Promotes Proliferation of Breast Cancer Cells in Hypoxic Environment. Anticancer. Res. 2017, 37, 4337–4343. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; He, Y.; Zhang, N.; Zhang, S.; Li, Y.; Wang, X.; Liang, Y.; Chen, X.; Zhao, W.; et al. Hypoxia-Induced FUS-circTBC1D14 Stress Granules Promote Autophagy in TNBC. Adv. Sci. 2023, 10, e2204988. [Google Scholar] [CrossRef]
- Yang, R.; Chen, H.; Xing, L.; Wang, B.; Hu, M.; Ou, X.; Chen, H.; Deng, Y.; Liu, D.; Jiang, R.; et al. Hypoxia-induced circWSB1 promotes breast cancer progression through destabilizing p53 by interacting with USP10. Mol. Cancer 2022, 21, 88. [Google Scholar] [CrossRef]
- Chen, T.; Wang, X.; Li, C.; Zhang, H.; Liu, Y.; Han, D.; Li, Y.; Li, Z.; Luo, D.; Zhang, N.; et al. CircHIF1A regulated by FUS accelerates triple-negative breast cancer progression by modulating NFIB expression and translocation. Oncogene 2021, 40, 2756–2771. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yang, R.; Xing, L.; Wang, B.; Liu, D.; Ou, X.; Deng, Y.; Jiang, R.; Chen, J. Hypoxia-inducible CircPFKFB4 Promotes Breast Cancer Progression by Facilitating the CRL4(DDB2) E3 Ubiquitin Ligase-mediated p27 Degradation. Int. J. Biol. Sci. 2022, 18, 3888–3907. [Google Scholar] [CrossRef]
- Wu, D.; Chen, T.; Zhao, X.; Huang, D.; Huang, J.; Huang, Y.; Huang, Q.; Liang, Z.; Chen, C.; Chen, M.; et al. HIF1alpha-SP1 interaction disrupts the circ-0001875/miR-31-5p/SP1 regulatory loop under a hypoxic microenvironment and promotes non-small cell lung cancer progression. J. Exp. Clin. Cancer Res. 2022, 41, 156. [Google Scholar] [CrossRef]
- Chi, Y.; Luo, Q.; Song, Y.; Yang, F.; Wang, Y.; Jin, M.; Zhang, D. Circular RNA circPIP5K1A promotes non-small cell lung cancer proliferation and metastasis through miR-600/HIF-1alpha regulation. J. Cell Biochem. 2019, 120, 19019–19030. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Xu, Y.; Hu, J.; Zhang, S.; Li, M.; Xu, L. A novel circular RNA, hsa-circ-0000211, promotes lung adenocarcinoma migration and invasion through sponging of hsa-miR-622 and modulating HIF1-α expression. Biochem. Biophys. Res. Commun. 2020, 521, 395–401. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, H.; Xia, L.; Lan, F. Circular RNA circ_0061140 accelerates hypoxia-induced glycolysis, migration, and invasion in lung adenocarcinoma through the microRNA-653/hexokinase 2 (HK2) axis. Bioengineered 2022, 13, 7156–7166. [Google Scholar] [CrossRef]
- Joseph, N.A.; Chiou, S.H.; Lung, Z.; Yang, C.L.; Lin, T.Y.; Chang, H.W.; Sun, H.S.; Gupta, S.K.; Yen, L.; Wang, S.D.; et al. The role of HGF-MET pathway and CCDC66 cirRNA expression in EGFR resistance and epithelial-to-mesenchymal transition of lung adenocarcinoma cells. J. Hematol. Oncol. 2018, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Pan, X.; Zhu, D.; Deng, Z.; Jiang, R.; Wang, X. Circular RNA MAT2B Promotes Glycolysis and Malignancy of Hepatocellular Carcinoma Through the miR-338-3p/PKM2 Axis Under Hypoxic Stress. Hepatology 2019, 70, 1298–1316. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Q.; Zuo, X.L.; Cai, J.; Zhang, Y.; Han, G.Y.; Zhang, L.; Ding, W.Z.; Wu, J.D.; Wang, X.H. Hypoxia-associated circPRDM4 promotes immune escape via HIF-1alpha regulation of PD-L1 in hepatocellular carcinoma. Exp. Hematol. Oncol. 2023, 12, 17. [Google Scholar] [CrossRef]
- Yang, W.; Liu, Y.; Gao, R.; Xiu, Z.; Sun, T. Knockdown of cZNF292 suppressed hypoxic human hepatoma SMMC7721 cell proliferation, vasculogenic mimicry, and radioresistance. Cell Signal 2019, 60, 122–135. [Google Scholar] [CrossRef]
- Wei, Y.; Chen, X.; Liang, C.; Ling, Y.; Yang, X.; Ye, X.; Zhang, H.; Yang, P.; Cui, X.; Ren, Y.; et al. A Noncoding Regulatory RNAs Network Driven by Circ-CDYL Acts Specifically in the Early Stages Hepatocellular Carcinoma. Hepatology 2020, 71, 130–147. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Du, B.; Zhan, Y.; Wang, K.; Wang, X.; Chen, B.; Wei, X.; Xiao, J. Antitumor effects of circ-EPHB4 in hepatocellular carcinoma via inhibition of HIF-1alpha. Mol. Carcinog. 2019, 58, 875–886. [Google Scholar] [CrossRef]
- Fu, Z.; Zhang, P.; Zhang, R.; Zhang, B.; Xiang, S.; Zhang, Y.; Luo, Z.; Huang, C. Novel hypoxia-induced HIF1alpha-circTDRD3-positive feedback loop promotes the growth and metastasis of colorectal cancer. Oncogene 2023, 42, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, Y.; Hao, W.; Zhao, W.; Yang, G.; Jing, C. Hypoxia-induced Circular RNA hsa_circ_0006508 Promotes the Warburg Effect in Colorectal Cancer Cells. Balk. Med. J. 2023, 40, 21–27. [Google Scholar] [CrossRef]
- Chen, L.Y.; Wang, L.; Ren, Y.X.; Pang, Z.; Liu, Y.; Sun, X.D.; Tu, J.; Zhi, Z.; Qin, Y.; Sun, L.N.; et al. The circular RNA circ-ERBIN promotes growth and metastasis of colorectal cancer by miR-125a-5p and miR-138-5p/4EBP-1 mediated cap-independent HIF-1alpha translation. Mol. Cancer 2020, 19, 164. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, J.; Lin, W.; Yuan, Q.; Lu, Y.; Wang, H.; Chen, Y.; Chen, L.; Dai, P.; Long, H.; et al. circEXOC6B interacting with RRAGB, an mTORC1 activator, inhibits the progression of colorectal cancer by antagonizing the HIF1A-RRAGB-mTORC1 positive feedback loop. Mol. Cancer 2022, 21, 135. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, H.; Yang, Y.; Wang, X.; Deng, T.; Liu, R.; Ning, T.; Bai, M.; Li, H.; Zhu, K.; et al. Hypoxia induced exosomal circRNA promotes metastasis of Colorectal Cancer via targeting GEF-H1/RhoA axis. Theranostics 2020, 10, 8211–8226. [Google Scholar] [CrossRef]
- Ou, Z.L.; Luo, Z.; Wei, W.; Liang, S.; Gao, T.L.; Lu, Y.B. Hypoxia-induced shedding of MICA and HIF1A-mediated immune escape of pancreatic cancer cells from NK cells: Role of circ_0000977/miR-153 axis. RNA Biol. 2019, 16, 1592–1603. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhu, Z.; Xiao, W.; Zong, G.; Wang, C.; Jiang, W.; Li, K.; Shen, J.; Guo, X.; Cui, J.; et al. Hypoxia-induced circRNF13 promotes the progression and glycolysis of pancreatic cancer. Exp. Mol. Med. 2022, 54, 1940–1954. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Wang, X.; Zhai, S.; Shi, M.; Peng, C.; Deng, X.; Fu, D.; Wang, J.; Shen, B. Hypoxia-induced exosomal circPDK1 promotes pancreatic cancer glycolysis via c-myc activation by modulating miR-628-3p/BPTF axis and degrading BIN1. J. Hematol. Oncol. 2022, 15, 128. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhao, Y.; Chen, Q.; Zhu, S.; Niu, Y.; Ye, Z.; Hu, P.; Chen, D.; Xu, P.; Chen, J.; et al. Hypoxic exosomal HIF-1alpha-stabilizing circZNF91 promotes chemoresistance of normoxic pancreatic cancer cells via enhancing glycolysis. Oncogene 2021, 40, 5505–5517. [Google Scholar] [CrossRef]
- Du, Y.; Zhang, J.Y.; Gong, L.P.; Feng, Z.Y.; Wang, D.; Pan, Y.H.; Sun, L.P.; Wen, J.Y.; Chen, G.F.; Liang, J.; et al. Hypoxia-induced ebv-circLMP2A promotes angiogenesis in EBV-associated gastric carcinoma through the KHSRP/VHL/HIF1alpha/VEGFA pathway. Cancer Lett. 2022, 526, 259–272. [Google Scholar] [CrossRef]
- Li, H.; Cao, B.; Zhao, R.; Li, T.; Xu, X.; Cui, H.; Deng, H.; Gao, J.; Wei, B. circDNMT1 Promotes Malignant Progression of Gastric Cancer Through Targeting miR-576-3p/Hypoxia Inducible Factor-1 Alpha Axis. Front. Oncol. 2022, 12, 817192. [Google Scholar] [CrossRef]
- Chen, W.; Ji, Y. CircC6orf132 Facilitates Proliferation, Migration, Invasion, and Glycolysis of Gastric Cancer Cells Under Hypoxia by Acting on the miR-873-5p/PRKAA1 Axis. Front. Genet. 2021, 12, 636392. [Google Scholar] [CrossRef]
- Liang, L.; Zhu, Y.; Li, J.; Zeng, J.; Wu, L. ALKBH5-mediated m6A modification of circCCDC134 facilitates cervical cancer metastasis by enhancing HIF1A transcription. J. Exp. Clin. Cancer Res. 2022, 41, 261. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, Y.; Zhang, Y.; Zheng, J.; Wang, S.; Cao, G. Circular RNA hsa_circ_0004543 Aggravates Cervical Cancer Development by Sponging MicroRNA hsa-miR-217 to Upregulate Hypoxia-Inducible Factor. J. Oncol. 2022, 2022, 4031403. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Huang, T.; Feng, W. Circular RNA HIPK3 Promotes EMT of Cervical Cancer Through Sponging miR-338-3p to Up-Regulate HIF-1alpha. Cancer Manag. Res. 2020, 12, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Zou, D.; Sun, Y.; Dai, Y. Hypoxia-associated circDENND2A promotes glioma aggressiveness by sponging miR-625-5p. Cell Mol. Biol. Lett. 2019, 24, 24. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Li, B.; Zhao, R.; Pan, Z.; Zhang, S.; Qiu, W.; Guo, Q.; Qi, Y.; Gao, Z.; Fan, Y.; et al. Hypoxia-induced circADAMTS6 in a TDP43-dependent manner accelerates glioblastoma progression via ANXA2/ NF-kappaB pathway. Oncogene 2023, 42, 138–153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Song, Y.C.; Qiu, F.; Wang, Z.C.; Li, N.; Zhao, F.B. Hypoxic glioma cell-secreted exosomal circ101491 promotes the progression of glioma by regulating miR-125b-5p/EDN1. Brain Res. Bull. 2023, 195, 55–65. [Google Scholar] [CrossRef]
- Xi, Y.; Shen, Y.; Wu, D.; Zhang, J.; Lin, C.; Wang, L.; Yu, C.; Yu, B.; Shen, W. CircBCAR3 accelerates esophageal cancer tumorigenesis and metastasis via sponging miR-27a-3p. Mol. Cancer 2022, 21, 145. [Google Scholar] [CrossRef]
- Gong, W.; Xu, J.; Wang, Y.; Min, Q.; Chen, X.; Zhang, W.; Chen, J.; Zhan, Q. Nuclear genome-derived circular RNA circPUM1 localizes in mitochondria and regulates oxidative phosphorylation in esophageal squamous cell carcinoma. Signal Transduct. Target. Ther. 2022, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Yang, W.; Jiang, N.; Shi, J.; Chen, L.; Zhong, G.; Bi, J.; Dong, W.; Wang, Q.; Wang, C.; et al. Hypoxia-elevated circELP3 contributes to bladder cancer progression and cisplatin resistance. Int. J. Biol. Sci. 2019, 15, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, Z.; Liu, B.; Sun, L. Silencing of circCYP51A1 represses cell progression and glycolysis by regulating miR-490-3p/KLF12 axis in osteosarcoma under hypoxia. J. Bone Oncol. 2022, 37, 100455. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Zhang, Y.; Wu, P.; Li, D.; Lu, Y.; Shen, P.; Yang, T.; Shi, G.; Chen, Q.; Yuan, H.; et al. CircSTX6 promotes pancreatic ductal adenocarcinoma progression by sponging miR-449b-5p and interacting with CUL2. Mol. Cancer 2022, 21, 121. [Google Scholar] [CrossRef]
- Chen, H.; Mao, M.; Jiang, J.; Zhu, D.; Li, P. Circular RNA CDR1as acts as a sponge of miR-135b-5p to suppress ovarian cancer progression. Onco Targets Ther. 2019, 12, 3869–3879. [Google Scholar] [CrossRef]
- Tang, J.; Wang, R.; Tang, R.; Gu, P.; Han, J.; Huang, W. CircRTN4IP1 regulates the malignant progression of intrahepatic cholangiocarcinoma by sponging miR-541-5p to induce HIF1A production. Pathol. Res. Pract. 2022, 230, 153732. [Google Scholar] [CrossRef] [PubMed]
- Vogel, A.; Meyer, T.; Sapisochin, G.; Salem, R.; Saborowski, A. Hepatocellular carcinoma. Lancet 2022, 400, 1345–1362. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Ren, S.; Liu, J.; Feng, Y.; Li, Z.; He, L.; Li, L.; Cao, X.; Wang, Z.; Zhang, Y. Knockdown of circDENND4C inhibits glycolysis, migration and invasion by up-regulating miR-200b/c in breast cancer under hypoxia. J. Exp. Clin. Cancer Res. 2019, 38, 388. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Fillmore, C.M.; Hammerman, P.S.; Kim, C.F.; Wong, K.K. Non-small-cell lung cancers: A heterogeneous set of diseases. Nat. Rev. Cancer 2014, 14, 535–546. [Google Scholar] [CrossRef]
- Noguchi, M.; Morikawa, A.; Kawasaki, M.; Matsuno, Y.; Yamada, T.; Hirohashi, S.; Kondo, H.; Shimosato, Y. Small adenocarcinoma of the lung. Histologic characteristics and prognosis. Cancer 1995, 75, 2844–2852. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, H.; Wang, J.; Wang, X.; Qian, L.; Xu, F.; Song, W.; Wu, D.; Shen, Z.; Feng, D.; et al. DeltaNp63alpha exerts antitumor functions in cervical squamous cell carcinoma. Oncogene 2020, 39, 905–921. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; Wick, W.; Aldape, K.; Brada, M.; Berger, M.; Pfister, S.M.; Nishikawa, R.; Rosenthal, M.; Wen, P.Y.; Stupp, R.; et al. Glioma. Nat. Rev. Dis. Primers 2015, 1, 15017. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Abdelmohsen, K.; Kim, J.; Yang, X.; Martindale, J.L.; Tominaga-Yamanaka, K.; White, E.J.; Orjalo, A.V.; Rinn, J.L.; Kreft, S.G.; et al. Scaffold function of long non-coding RNA HOTAIR in protein ubiquitination. Nat. Commun. 2013, 4, 2939. [Google Scholar] [CrossRef]
- Weng, X.; Liu, H.; Ruan, J.; Du, M.; Wang, L.; Mao, J.; Cai, Y.; Lu, X.; Chen, W.; Huang, Y.; et al. HOTAIR/miR-1277-5p/ZEB1 axis mediates hypoxia-induced oxaliplatin resistance via regulating epithelial-mesenchymal transition in colorectal cancer. Cell Death Discov. 2022, 8, 310. [Google Scholar] [CrossRef]
- Meng, L.; Ward, A.J.; Chun, S.; Bennett, C.F.; Beaudet, A.L.; Rigo, F. Towards a therapy for Angelman syndrome by targeting a long non-coding RNA. Nature 2015, 518, 409–412. [Google Scholar] [CrossRef]
- Tuveson, D.; Clevers, H. Cancer modeling meets human organoid technology. Science 2019, 364, 952–955. [Google Scholar] [CrossRef] [PubMed]
- Barbier, A.J.; Jiang, A.Y.; Zhang, P.; Wooster, R.; Anderson, D.G. The clinical progress of mRNA vaccines and immunotherapies. Nat. Biotechnol. 2022, 40, 840–854. [Google Scholar] [CrossRef]
- Qu, L.; Yi, Z.; Shen, Y.; Lin, L.; Chen, F.; Xu, Y.; Wu, Z.; Tang, H.; Zhang, X.; Tian, F.; et al. Circular RNA vaccines against SARS-CoV-2 and emerging variants. Cell 2022, 185, 1728–1744.e1716. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hypoxia-Related lncRNAs | Expression | Interacting Partner | Target Genes/Pathways | Functions | Cancer Type | Reference |
|---|---|---|---|---|---|---|
| DACT3-AS1 | ↑ | HDAC2/FOXA3 | FOXA3 | Facilitates HCC cell metastasis is achieved by enhancing FOXA3 deacetylation through promoting the interaction between HDAC2 and FOXA3 | HCC | [47] |
| ALKBH3-AS1 | ↑ | ALKBH3 | Facilitates the proliferation and invasion of HCC cells by enhancing the stability of ALKBH3 mRNA | HCC | [48] | |
| AC115619 | ↓ | WTAP | Inhibits HCC cell metastasis by encoding a micropeptide that suppresses m6A modification | HCC | [49] | |
| lncRNA LET | ↓ | NF90 | HIF-1α | Inhibits HCC metastasis by decreasing the mRNA expression of HIF-1α through enhanced NF90 degradation. | HCC | [50] |
| FRMD6-AS1 | ↑ | SENP1 | HIF-1α | Promotes HCC cell metastasis is achieved by enhancing the protease activity of SENP1 to regulate SUMOylation of HIF-1α | HCC | [51] |
| Linc-RoR | ↑ | miR-145 | HIF-1α | Facilitates HCC cell proliferation by the miR-145/HIF-1α axis | HCC | [52] |
| EIF3J-AS1 | ↑ | miR-122-5p | CTNND2 | Promotes HCC cell proliferation and invasion by the miR-122-5p/CTNND2 axis | HCC | [53] |
| HMMR-AS1 | ↑ | miR-147 | ARID3A | Promotes HCC cell proliferation and influences macrophage polarization via the miR-147/ARID3A axis | HCC | [54] |
| lncRNA PMAN | ↑ | ELAVL1 | SLC7A11 | Facilitates GC cell metastasis by promoting the cytoplasmic localization of ELAVL1 to increase the stability of SLC7A11 mRNA | GC | [55] |
| BC005927 | ↑ | EPHB4 | Promotes GC cell metastasis by upregulating EPHB4 | GC | [56] | |
| lncRNA AK058003 | ↑ | SNCG | Promotes GC metastasis by regulating SNGG via DNA demethylation | GC | [57] | |
| PVT1 | ↑ | miR-186 | HIF-1α | Facilitates GC cell invasion by the miR-186/HIF-1α axis | GC | [58] |
| ZEB2-AS1 | ↑ | miR-143-5p | HIF-1α | Promotes GC cell proliferation and invasion by the miR-143-5p/HIF-1α axis | GC | [59] |
| lncRNA CF129 | ↓ | p53 | FOXC2 | Inhibits pancreatic cell proliferation and invasion by inducing the MKRN1-mediated ubiquitin-dependent degradation of p53 | Pancreatic cancer | [60] |
| NORAD | ↑ | hsa-miR-125a-3p | RhoA | Facilitates pancreatic cancer cell metastasis by the miR-125a-3/RhoA axis | Pancreatic cancer | [61] |
| lncRNA MTA2TR | ↑ | ATF3 | MTA2 | Facilitates proliferation and invasion by recruiting ATF3 to the MTA2 promoter region to activate the transcription of MTA2 | Pancreatic cancer | [62] |
| lncRNA BX111 | ↑ | YB1 | ZEB1 | Promotes pancreatic cancer cell metastasis by recruiting YB1 to the ZEB1 promoter region to activate the transcription of ZEB1 | Pancreatic cancer | [63] |
| ENST0000480739 | ↓ | OS-9/HIF-1α | Inhibits pancreatic cancer cell invasion by suppressing HIF-1α expression | Pancreatic cancer | [64] | |
| HITT | ↓ | YBX1 | Suppresses CRC angiogenesis and tumor growth by binding with YB-1 to downregulate HIF-1α | CRC | [65] | |
| LUCAT1 | ↑ | PTBP1 | Facilitates CRC cell viability and chemotherapy response by modifying the alternative splicing of DNA damage-related genes through interaction with PTBP1 | CRC | [66] | |
| STEAP3-AS1 | ↑ | STEAP3 | Wnt/β-catenin signaling | Promotes CRC cell proliferation and metastasis by preventing m6A-mediated degradation of STEAP3 mRNA | CRC | [67] |
| LVBU | ↑ | miR-10a/miR-34c | BCL6 | Facilitates CRC cell proliferation by sponging miR-10a/miR-34c to enhance BCL6 expression | CRC | [68] |
| ELFN1-AS1 | ↑ | miR-191-5p | TRIM14 | Promotes CRC cell proliferation and invasion by sponging miR-191-5p to increase TRIM14 expression | CRC | [69] |
| DLEU1 | ↑ | CKAP2 | ERK/STAT3 | Promotes BC cell metastasis by interacting with CKAP2 to activate the ERK and STAT3 signaling pathways | BC | [70] |
| LINC00649 | ↑ | NF90/NF45 | HIF-1α | Facilitates BC cell metastasis by maintaining the stability of HIF-1α through the NF90/NF45 complex | BC | [71] |
| EFNA3 | ↑ | miRNA-210 | EFNA3 | Facilitates BC cell metastasis by the miR-210/EFNA3 axis | BC | [72] |
| NDRG1-OT1 | ↑ | miR-875-3p | Facilitates BC cell metastasis via sponging miR-875-3p | BC | [73] | |
| HIF1A-AS2 | ↑ | HMGA1 | DHX9/IGF2BP2 | Promotes the expression of HMGA1 by interacting with DHX9 and IGF2BP2 mRNA binding complexes | Glioma | [74] |
| lncRNA H19 | ↑ | miR-181d | Promotes glioma cell migration and invasion by acting as a ceRNA of miR-181d to increase β-catenin expression | Glioma | [75] | |
| HOTTIP | ↑ | miR-101 | ZEB1 | Facilitates epithelial-mesenchymal transition through the miR-101/ZEB1 axis | Glioma | [76] |
| lncRNA SARCC | ↑ | AR | HIF-2α/C-MYC | Inhibits proliferation of VHL-mutant cells yet promotes proliferation of VHL-normal cells via modulating the AR/HIF-2α/C-MYC axis | RCC | [77] |
| RP11-367G18.1 V2 | ↑ | H4K16Ac | Facilitates RCC cell metastasis by promoting acetylation of H4K16Ac | RCC | [78] | |
| HOTAIR | ↑ | miR-217 | HIF-1α/AXL | Facilitates RCC cell metastasis and EMT by regulating miR-217/HIF-1α/AXL signaling pathway | RCC | [79] |
| PVT1 | ↑ | KAT2A | HIF-1α | Promotes nasopharyngeal carcinoma cell proliferation by activating KAT2A to increase the stability of HIF-1α | NPC | [80] |
| DANCR | ↑ | NF90/NF45 | HIF-1α | Improves HIF-1α mRNA stability through interacting with the NF90/NF45 complex | NPC | [81] |
| NEAT1 | ↑ | miR-101-3p | OX9/Wnt/β-Catenin | Promotes NSCLC progression via the miR-101-3p/SOX9/Wnt/β-Catenin signal pathway | NSCLC | [82] |
| LHFPL3-AS2 | ↓ | SFPQ | TXNIP | Suppresses NSCLC cell metastasis by interacting with SFPQ to regulate TXNIP expression | NSCLC | [83] |
| MIR17HG | ↑ | miR-155-5p | HIF-1α | Promotes retinoblastoma cell proliferation and invasion by the miR-155-5p/HIF-1α axis | Retinoblastoma | [84] |
| HIFCAR | ↑ | HIF-1α/p300 | Facilitates the recruitment of HIF-1α and the p300 cofactor to the target gene promoters | Oral carcinoma | [85] | |
| lincRNA-p21 | ↑ | VHL/HIF-1α | Enhances glycolysis by inhibiting the VHL-mediated ubiquitination of HIF-1α | Cervical cancer | [86] | |
| DARS-AS1 | ↑ | METTL3/METTL14 | DARS | Promotes cervical cancer cell autophagy by facilitating the translation of DARS through METTL3- and METTL14-mediated m6A modification | Cervical cancer | [87] |
| DARS-AS1 | ↑ | RNF147 | RBM39 | Facilitates multiple myeloma cell proliferation and apoptosis by regulating RBM39 stability through inhibiting its interaction with RNF147 | Multiple myeloma | [88] |
| WT1-AS | ↑ | H3K4/H3K9 | WT-1 | Promotes myeloid leukemia cell proliferation by modulating the methylation of H3K4 and H3K9 to facilitate WT-1 upregulation | Myeloid leukemia | [89] |
| MEG3 | ↑ | DNMT3a/DNMT3b/MBD1 | TIMP2 | Facilitates pheochromocytoma cell proliferation by recruiting DNMT3a, DNMT3b, and MBD1 to accelerate TIMP2 promoter methylation | Pheochromocytoma | [90] |
| Hypoxia-Related circRNAs | Expression | Interacting Partner | Target Genes/Pathways | Functions | Cancers | Reference |
|---|---|---|---|---|---|---|
| CircRNF20 | ↑ | miR-487a | HIF-1α/HK2 | Contributes to BC cell proliferation and aerobic glycolysis | BC | [103] |
| CircZFR | ↑ | miR-578 | HIF-1α | Facilitates BC progression | BC | [104] |
| CircRBM33 | ↑ | miR-542-3p | HIF-1α | Facilitates hypoxia-induced glycolysis | BC | [105] |
| CircDENND4C | ↑ | miR-200b/miR-200c | Promotes BC cell proliferation under hypoxic conditions | BC | [106] | |
| CircTBC1D14 | ↑ | FUS | PRMT1 | Maintains cellular homeostasis and promotes tumor progression | BC | [107] |
| CircWSB1 | ↑ | USP10 | Promotes the proliferation of BC cells | BC | [108] | |
| CircHIF-1α | ↑ | NFIB/FUS | Facilitates the development and metastasis of BC | BC | [109] | |
| CircPFKFB4 | ↑ | DDB1/DDB2 | CRL4DDB2/p27 | Facilitates BC progression | BC | [110] |
| circ-0001875 | ↑ | miR-31-5p | SP1 | Facilitates the progression of NSCLC | NSCLC | [111] |
| CircPIP5K1A | ↑ | miR-600 | HIF-1α | Facilitates tumor proliferation and metastasis | NSCLC | [112] |
| hsa-circ-0000211 | ↑ | miR-622 | HIF-1α | Promotes cancer cell migration and invasion | LUAD | [113] |
| Circ0061140 | ↑ | miR-653 | HK2 | Accelerates hypoxia-induced glycolysis, migration, and invasion in LUAD | LUAD | [114] |
| CircRNA CCDC66 | ↑ | Advances drug resistance and EMT | LUAD | [115] | ||
| CircMAT2B | ↑ | miR-338-3p | PKM2 | Facilitates HCC progression by enhancing glycolysis | HCC | [116] |
| CircPRDM4 | ↑ | HIF-1α | Facilitates immune escape of HCC cells | HCC | [117] | |
| cZNF292 | ↑ | miR-23b-3p | Wnt/β-catenin | Promotes the hypoxia-induced proliferation of human hepatoma SMMC7721 cells, radioresistance, and vasculogenic mimicry | HCC | [118] |
| CircCDYL | ↑ | miR-328-3p | HIF-1AN | Promotes stem-like characteristics and tumor development | HCC | [119] |
| Circ-EPHB4 | ↓ | HIF-1α/PI3K-AKT | Inhibits the development, growth, and metastasis of HCC | HCC | [120] | |
| CircTDRD3 | ↑ | miR-1231 | HIF-1α | Facilitates the growth, migration, and metastasis of CRC cells | CRC | [121] |
| Circ_0006508 | ↑ | miR-1272 | HIF-1α | Promotes the viability and Warburg effect of CRC in vitro | CRC | [122] |
| Circ-Erbin | ↑ | miR-125a-5p/miR-138-5p | 4EBP-1 | Promotes CRC cell proliferation and metastasis | CRC | [123] |
| CircEXOC6B | ↓ | RRAGB | Inhibits the development of CRC cells and increases 5-fluorouracil-induced apoptosis | CRC | [124] | |
| Circ-133 | ↑ | miR-133a | GEF-H1/RhoA | Facilitates metastasis | CRC | [125] |
| Circ_0000977 | ↑ | miR-153 | HIF1/ADAM10 | Modulates the HIF-1α-mediated immune evasion of pancreatic cancer cells in vitro | Pancreatic cancer | [126] |
| CircRNF13 | ↑ | miR-654-3p | PDK3 | Promotes the malignant progression of pancreatic cancer | Pancreatic cancer | [127] |
| CircPDK1 | ↑ | miR-628-3p/BPTF | c-myc | Facilitates the proliferation, migration, and glycolysis of pancreatic cancer cells | Pancreatic cancer | [128] |
| CircZNF91 | ↑ | SIRT1 and HIF-1α | Promotes the resistance of pancreatic cancer cells to GEM | Pancreatic cancer | [129] | |
| ebv-circLMP2A | ↑ | KHSRP | HIF-1α | Promotes angiogenesis in EBV-associated GC | GC | [130] |
| CircDNMT1 | ↑ | miR-576-3p | HIF-1α | Promotes the malignant progression of GC | GC | [131] |
| CircC6orf132 | ↑ | miR-873-5p | PRKAA1 | Promotes GC cell proliferation, migration, invasion, and glycolysis in a hypoxic environment in vitro and in vivo | GC | [132] |
| CircCCDC134 | ↑ | miR-503-5p | HIF-1α | Stimulates HIF-1α transcription and facilitates the growth and metastasis of cervical cancer | Cervical cancer | [133] |
| Circ_0004543 | ↑ | miR-217 | HIF-1α | Aggravates cervical cancer development | Cervical cancer | [134] |
| Circ-HIPK3 | ↑ | miR-338-3p | HIF-1α | Facilitates the EMT of cervical cancer cells | Cervical cancer | [135] |
| CircDENND2A | ↑ | miR-625-5p | Facilitates the aggressiveness of glioma | Glioma | [136] | |
| CircADAMTS6 | ↑ | ANXA2 | Promotes cell proliferation and inhibits apoptosis, accelerating the development of glioblastoma | Glioma | [137] | |
| Circ101491 | ↑ | miR-125b-5p | EDN1 | Promotes the viability, invasiveness, and migration of glioma cells | Glioma | [138] |
| CircBCAR3 | ↑ | miR-27A-3p | TnPO1 | Facilitates the migration and ferroptosis of ESCC | ESCC | [139] |
| CircPUM1 | ↑ | UQCRC2 | Inhibits ESCC cell pyroptosis, thereby promoting the proliferation of ESCC cells both in vivo and in vitro | ESCC | [140] | |
| CircELP3 | ↑ | Promotes the development of bladder cancer and cisplatin resistance | Bladder cancer | [141] | ||
| CircCYP51A1 | ↑ | miR-490-3p | KLF12 | Mediates cancer cell proliferation, migration, invasion, and glycolysis | Osteosarcoma | [142] |
| CircSTX6 | ↑ | miR-449b-5p | HIF-1α | Promotes tumor proliferation and metastasis both in vitro and in vivo | PDAC | [143] |
| CDR1as | ↓ | miR-135b-5p | HIF-1AN | Inhibits the development of ovarian cancer in vitro | Ovarian cancer | [144] |
| CircRTN4IP1 | ↑ | miR-541-5p | HIF-1α | Facilitates tumor formation in vivo | ICC | [145] |
| NcRNAs | Expression | Cancer Type | Source | Sample Size | Sample Grouping | Clinical Significance | Reference |
|---|---|---|---|---|---|---|---|
| lncRNA HOTTIP | ↑ | Glioma | Tissues | 60|56 | Metastatic glioma patients/non-metastatic glioma patients | Associated with metastatic progression (p < 0.0001) | [76] |
| lncRNA PVT1 | ↑ | NPC | Tissues | 10|10 | NPC patients/healthy donors | Associated with diagnosis and prognosis (p < 0.001) | [80] |
| lncRNA HITT | ↓ | CRC | Tissues | 46|46 | CRC patient/matched adjacent controls | Associated with diagnosis and prognosis (p < 0.01) | [65] |
| lncRNA BX111 | ↑ | Pancreatic cancer | Tissues | 17|31 | Resected/non-resected | Associated with metastatic progression (p < 0.01) | [63] |
| circTDRD3 | ↑ | CRC | Tissues | 106|106/55|55 | CRC patients/matched adjacent controls | Associated with tumor size, TNM stage, and lymph node invasion (p < 0.05) | [121] |
| circDENND4C | ↑ | BC | Tissues | 30|30 | BC patients/matched adjacent controls | Associated with tumor size (p < 0.0001) | [106] |
| circ101491 | ↑ | Glioma | Blood sample | 20|20 | Cancer patients/healthy donors | Associated with the degree of tumor differentiation and tumor TNM staging (p = 0.037 and p = 0.009, respectively) | [138] |
| circ-133 | ↑ | CRC | Blood sample | 10|10 | Cancer patients/healthy donors | Associated with TNM staging (p < 0.01) | [125] |
| circHIF1A | ↑ | BC | Blood sample | 101|84 | CRC patients/matched adjacent controls | Associated with overall survival (OS) (p = 0.025) | [109] |
| lncRNA HOTAIR | ↑ | Thyroid Cancer | Blood sample | 90 participants | Cancer patients/normal healthy subjects | Associated with diagnosis | NCT03469544 |
| lncRNA HOTTIP | ↑ | CRC | Blood sample | 60 participants | Cancer patients/normal healthy subjects | Associated with diagnosis | NCT04729855 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, L.; Zou, C.; Zhang, H.; Yang, F.; Qi, G.; Ma, Z. Landscape of Noncoding RNA in the Hypoxic Tumor Microenvironment. Genes 2025, 16, 140. https://doi.org/10.3390/genes16020140
Gong L, Zou C, Zhang H, Yang F, Qi G, Ma Z. Landscape of Noncoding RNA in the Hypoxic Tumor Microenvironment. Genes. 2025; 16(2):140. https://doi.org/10.3390/genes16020140
Chicago/Turabian StyleGong, Lianfeng, Chuanxin Zou, Haixia Zhang, Fei Yang, Gui Qi, and Zhaowu Ma. 2025. "Landscape of Noncoding RNA in the Hypoxic Tumor Microenvironment" Genes 16, no. 2: 140. https://doi.org/10.3390/genes16020140
APA StyleGong, L., Zou, C., Zhang, H., Yang, F., Qi, G., & Ma, Z. (2025). Landscape of Noncoding RNA in the Hypoxic Tumor Microenvironment. Genes, 16(2), 140. https://doi.org/10.3390/genes16020140