
Comparison of Brain Gene Expression Profiles Associated with Auto-Grooming Behavior between Apis cerana and Apis mellifera Infested by Varroa destructor


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Honeybee and Mites Collection
2.2. Auto-Grooming Behavior Experiment
2.3. RNA Extraction, Library Construction and Data Quality Control
2.4. Differential Expression Genes Analysis
2.5. Homologous Gene Analysis
2.6. Real-Time Fluorescence Quantitative PCR Verification
2.7. Statistical Analyses
3. Results
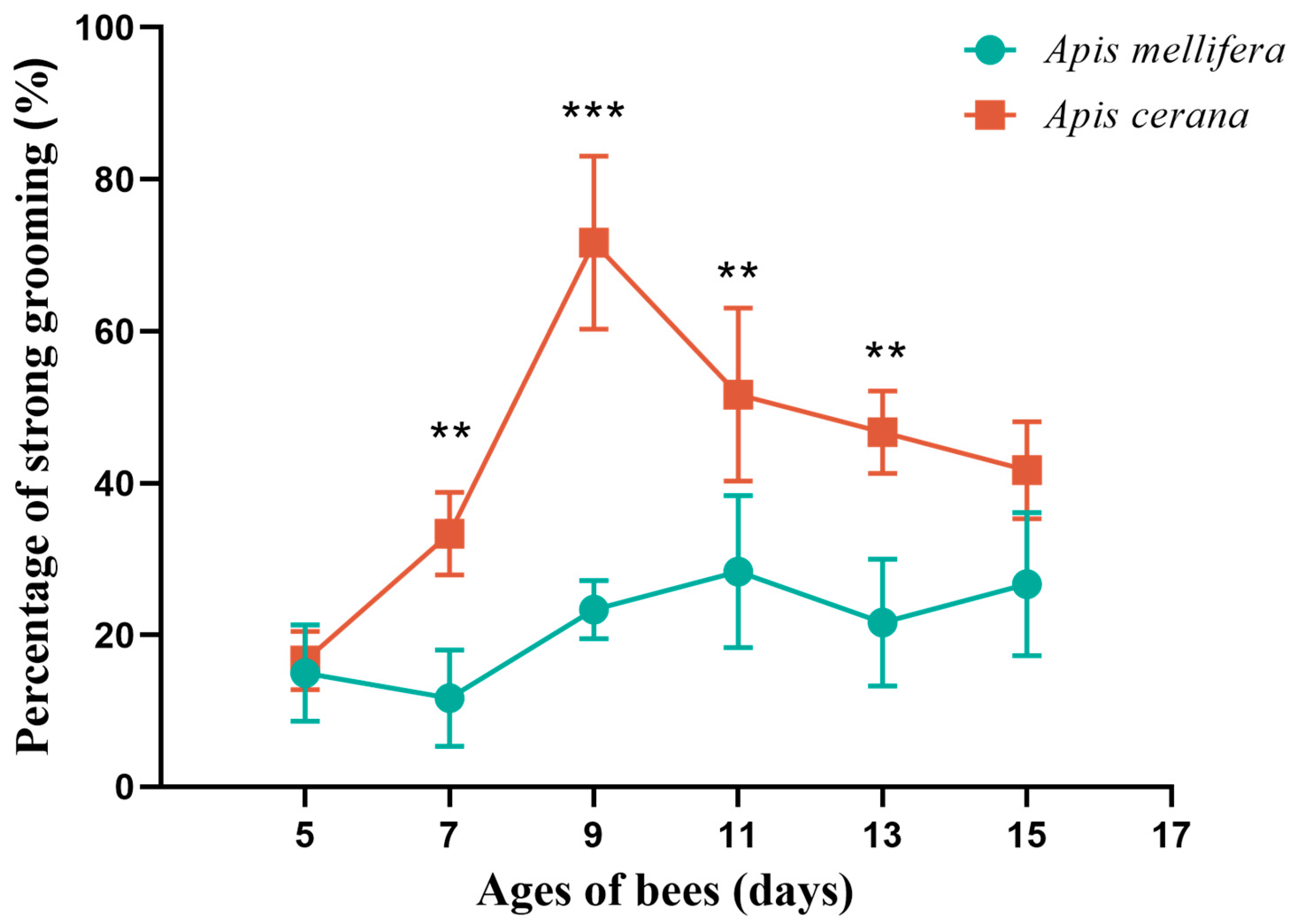
3.1. Different Days Analysis of Auto-Grooming Behavior
3.2. Overview of RNA-Seq Data
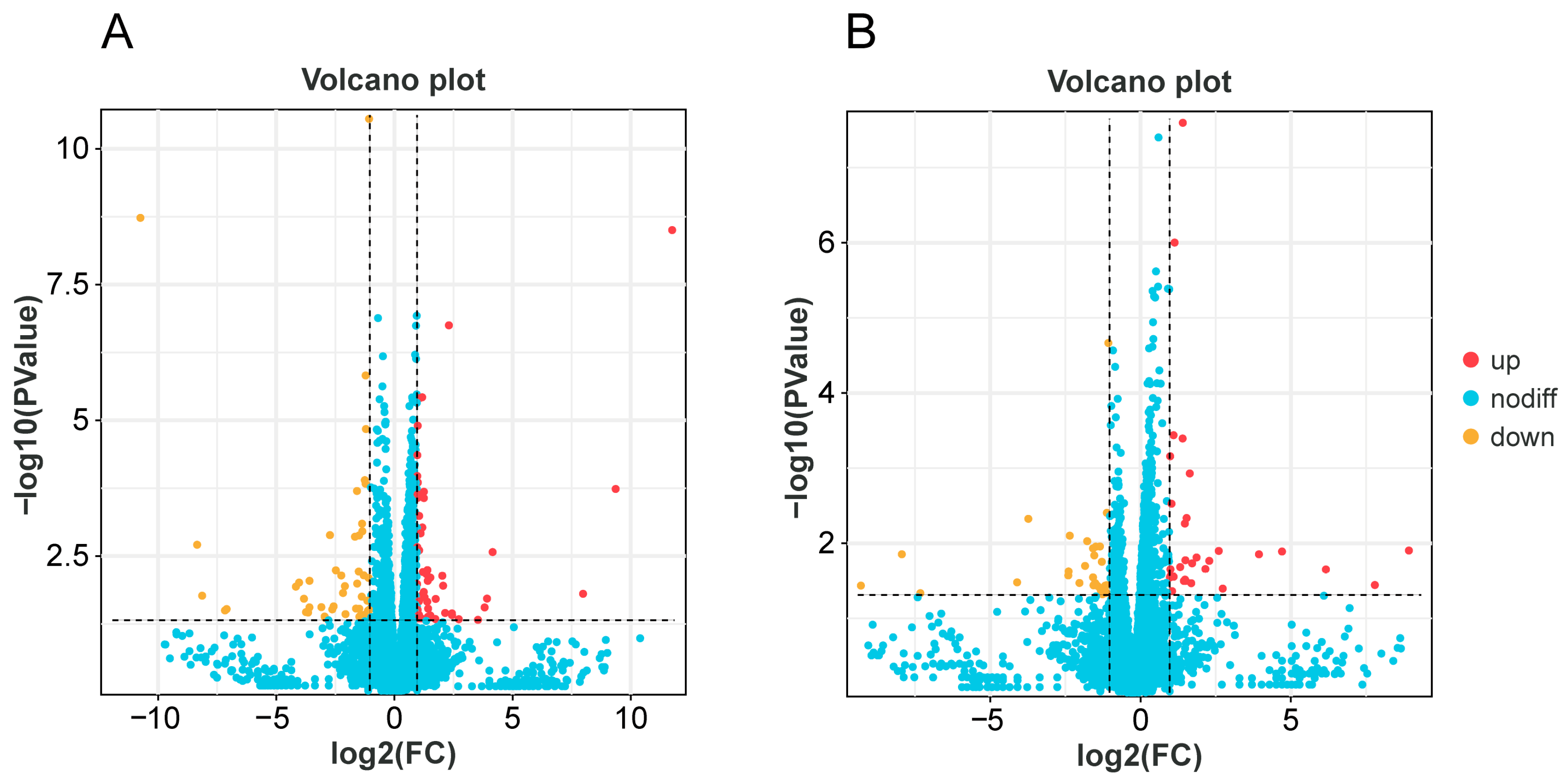
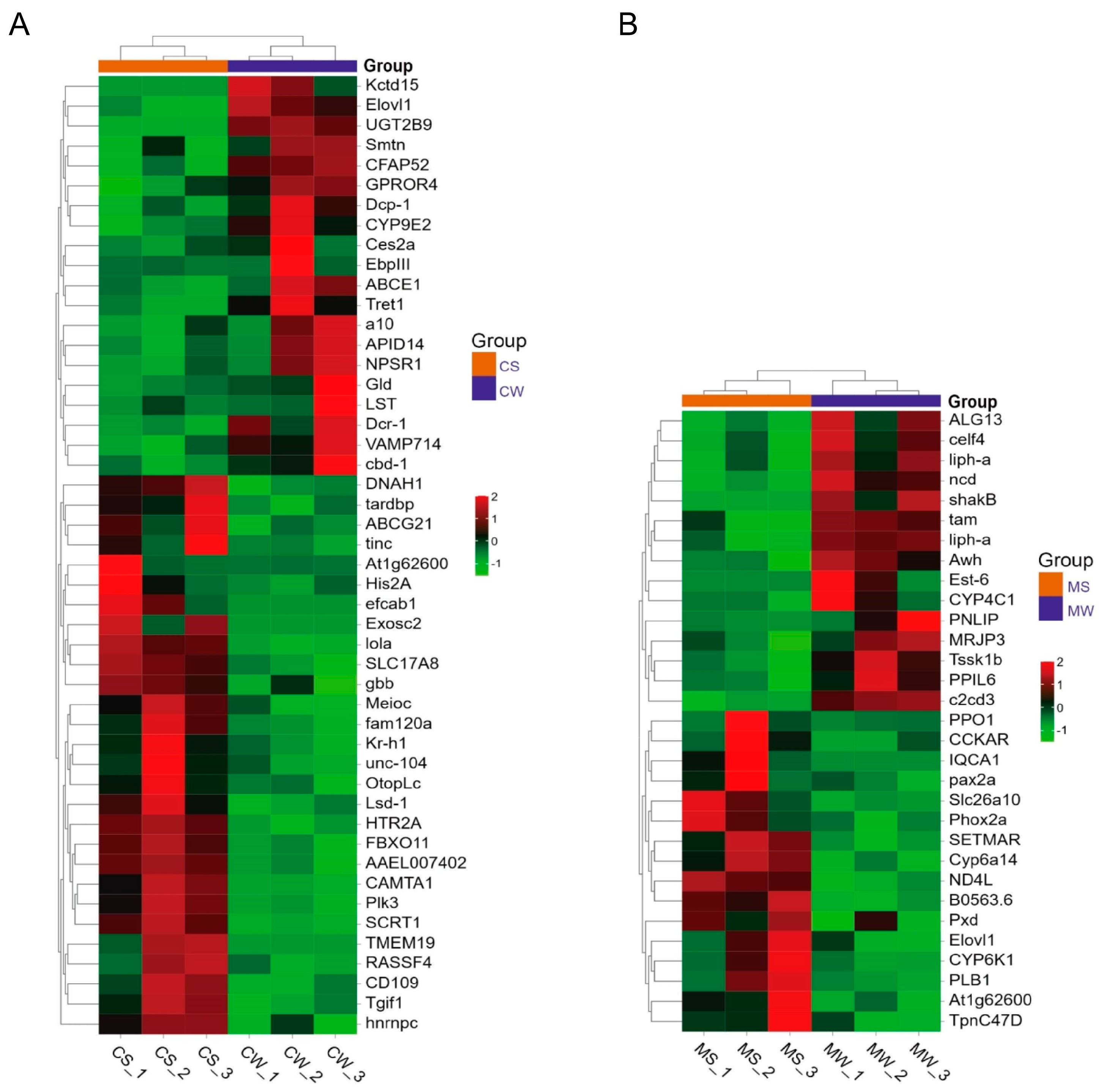
3.3. DEGs Analysis
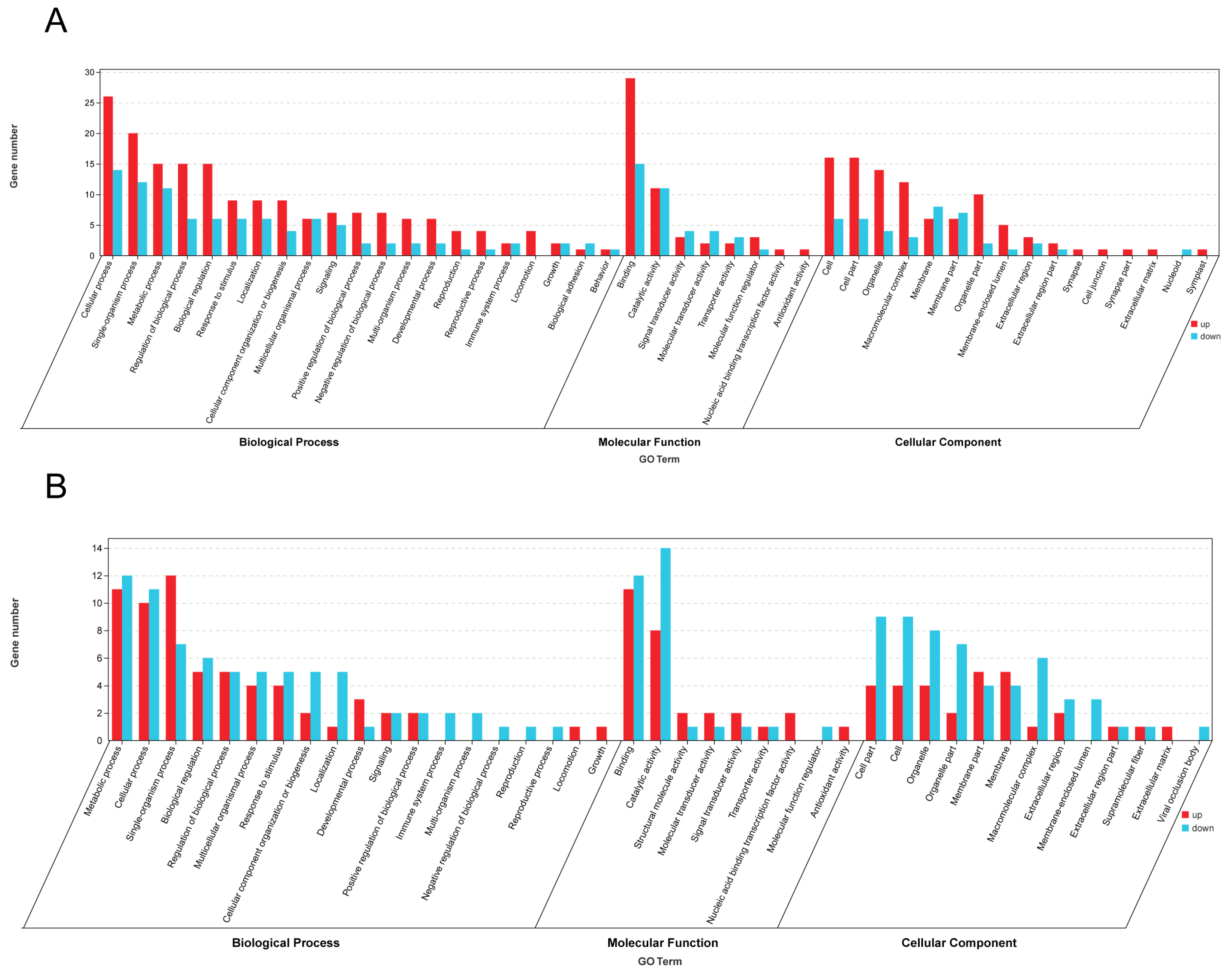
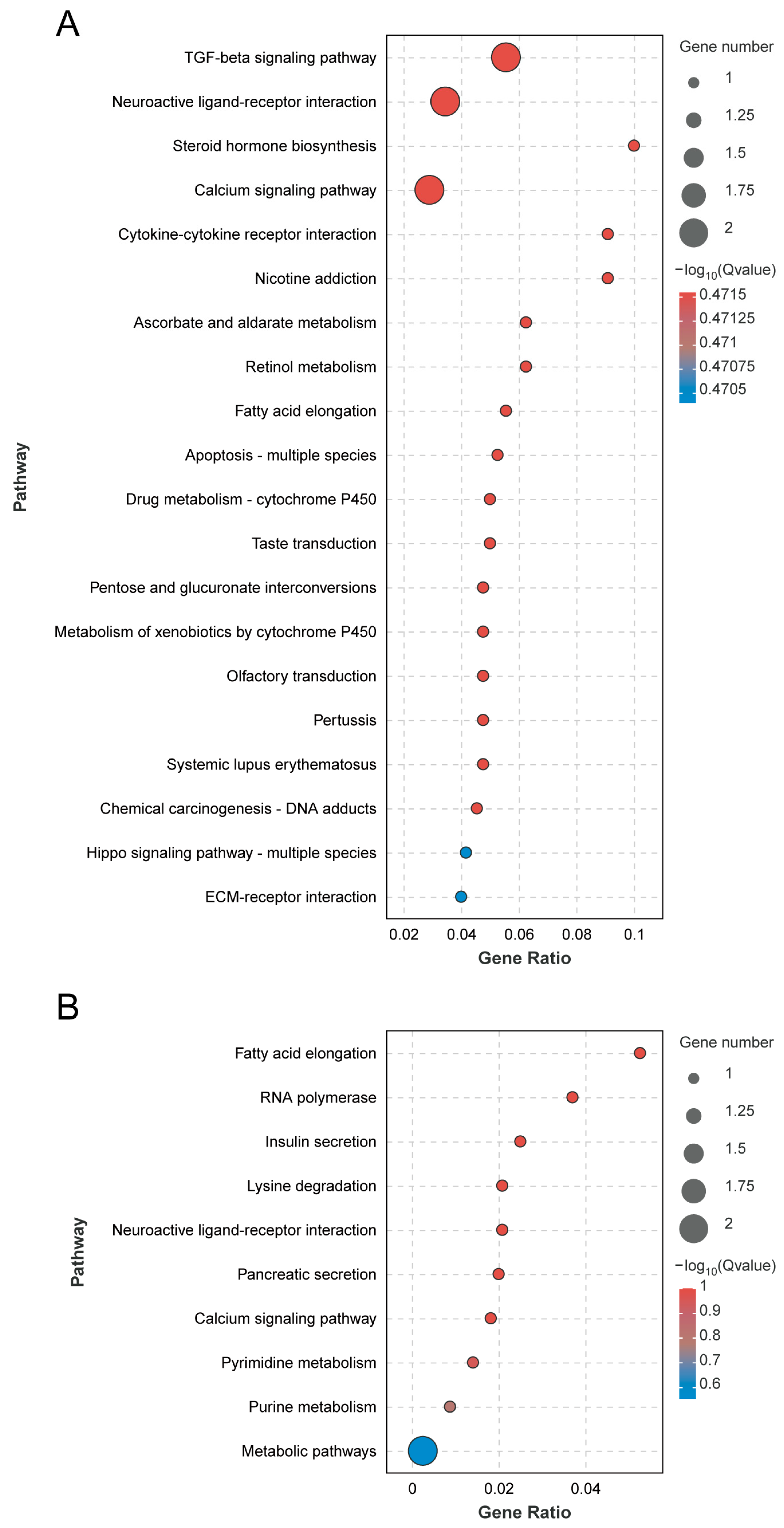
3.4. GO Functional Enrichment and KEGG Pathway Enrichment Analyses of A. cerana and A. mellifera
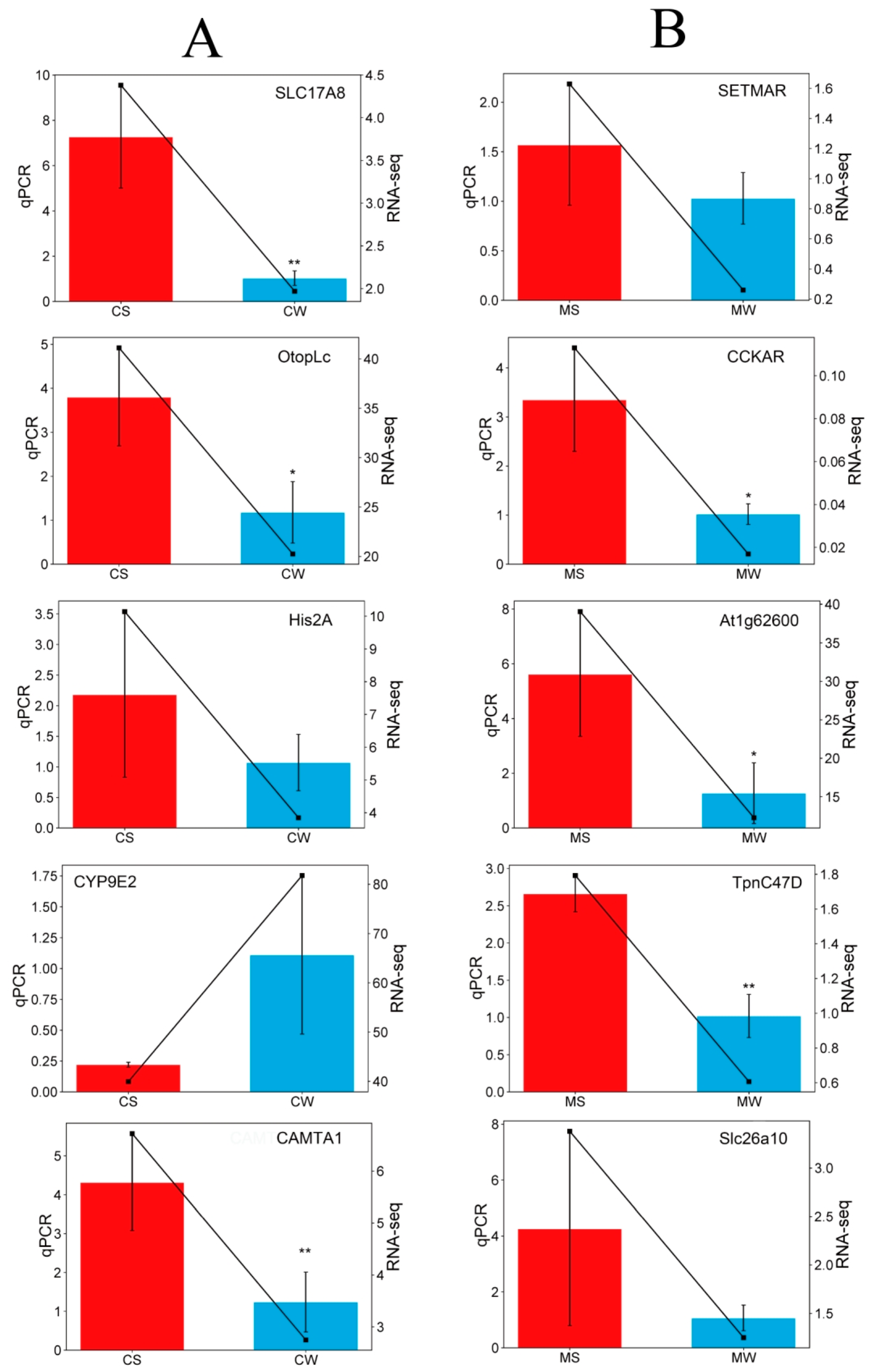
3.5. qPCR Analysis
3.6. Homologous Gene Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Traynor, K.S.; Mondet, F.; de Miranda, J.R.; Techer, M.; Kowallik, V.; Oddie, M.A.Y.; Chantawannakul, P.; McAfee, A. Varroa Destructor: A Complex Parasite, Crippling Honey Bees Worldwide. Trends Parasitol. 2020, 36, P592–P606. [Google Scholar] [CrossRef] [PubMed]
- Warner, S.; Pokhrel, L.R.; Akula, S.M.; Ubah, C.S.; Richards, S.L.; Jensen, H.; Kearney, G.D. A scoping review on the effects of Varroa Mite (Varroa Destructor) on global honey bee decline. Sci. Total Environ. 2024, 906, 167492. [Google Scholar] [CrossRef] [PubMed]
- Techer, M.A.; Roberts, J.M.K.; Cartwright, R.A.; Mikheyev, A.S. The first steps toward a global pandemic: Reconstructing the demographic history of parasite host switches in its native range. Mol. Ecol. 2022, 31, 1358–1374. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa Destructor. J. Invertebr. Pathol. 2010, 103 (Suppl. S1), S96–S119. [Google Scholar] [CrossRef] [PubMed]
- Vilarem, C.; Piou, V.; Vogelweith, F.; Vétillard, A. Varroa Destructor from the Laboratory to the Field: Control, Biocontrol and IPM Perspectives—A Review. Insects 2021, 12, 800. [Google Scholar] [CrossRef] [PubMed]
- Morfin, N.; Goodwin, P.H.; Guzman-Novoa, E. Varroa Destructor and its impacts on honey bee biology. Front. Bee Sci. 2023, 1, 1272937. [Google Scholar] [CrossRef]
- Le Conte, Y.; Meixner, M.; Brandt, A.; Carreck, N.; Costa, C.; Mondet, F.; Ralph, B. Geographical distribution and selection of european honey bees resistant to Varroa Destructor. Insects 2020, 11, 873. [Google Scholar] [CrossRef] [PubMed]
- Luis, A.R.; Grindrod, I.; Webb, G.; Piñeiro, A.P.; Martin, S.J. Recapping and mite removal behaviour in Cuba: Home to the world’s largest population of Varroa-resistant European honeybees. Sci. Rep. 2022, 12, 15597. [Google Scholar] [CrossRef] [PubMed]
- Grindrod, I.; Martin, S.J. Varroa resistance in Apis Cerana: A review. Apidologie 2023, 54, 14. [Google Scholar] [CrossRef]
- Guichard, M.; Dietemann, V.; Neuditschko, M.; Dainat, B. Advances and perspectives in selecting resistance traits against the parasitic mite Varroa Destructor in honey bees. Genet. Sel. Evol. 2020, 52, 71. [Google Scholar] [CrossRef]
- Invernizzi, C.; Zefferino, I.; Santos, E.; Sánchez, L.; Mendoza, Y. Multilevel assessment of grooming behavior against Varroa Destructor in Italian and Africanized honey bees. J. Apic. Res. 2015, 54, 321–327. [Google Scholar] [CrossRef]
- van Alphen, J.J.M.; Fernhout, B.J. Natural selection, selective breeding, and the evolution of resistance of honeybees (Apis Mellifera) against Varroa. Zool. Lett. 2020, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, D.J. Grooming by honey bees as a component of varroa resistant behavior. J. Apic. Res. 2016, 55, 38–48. [Google Scholar] [CrossRef]
- Nganso, B.T.; Fombong, A.T.; Yusuf, A.A.; Pirk, C.W.W.; Stuhl, C.; Torto, B. Hygienic and grooming behaviors in African and European honeybees-New damage categories in Varroa Destructor. PLoS ONE 2017, 12, e0179329. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Novoa, E.; Emsen, B.; Unger, P.; Espinosa-Montaño, L.G.; Petukhova, T. Genotypic variability and relationships between mite infestation levels, mite damage, grooming intensity, and removal of Varroa Destructor mites in selected strains of worker honey bees (Apis Mellifera L.). J. Invertebr. Pathol. 2012, 110, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Arechavaleta-Velasco, M.E.; Alcala-Escamilla, K.; Robles-Rios, C.; Tsuruda, J.M.; Hunt, G.J. Fine-scale linkage mapping reveals a small set of candidate genes influencing honey bee grooming behavior in response to Varroa Mites. PLoS ONE 2012, 7, e47269. [Google Scholar] [CrossRef] [PubMed]
- Hamiduzzaman, M.M.; Emsen, B.; Hunt, G.J.; Subramanyam, S.; Williams, C.E.; Tsuruda, J.M.; Guzman-Novoa, E. Differential Gene Expression Associated with Honey Bee Grooming Behavior in Response to Varroa Mites. Behav. Genet. 2017, 47, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.-S.; Fang, Y.; Xu, S.; Ge, L. The resistance mechanism of the Asian honey bee, Apis Cerana Fabr., to an ectoparasitic mite, Varroa Jacobsoni Oudemans. J. Invertebr. Pathol. 1987, 49, 54–60. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-Seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Panasiuk, B.; Skowronek, W.; Bieńkowska, M.; Gerula, D.; Węgrzynowicz, P. Age of worker bees performing hygienic behaviour in a honeybee colony. J. Apic. Sci. 2010, 54, 109–115. [Google Scholar]
- Pettis, J.S.; Pankiw, T. Grooming behavior by Apis Mellifera L. in the presence of Acarapis Woodi (Rennie) (Acari: Tarsonemidae). Apidologie 1998, 29, 241–253. [Google Scholar] [CrossRef]
- Dadoun, N.; Nait-Mouloud, M.; Mohammedi, A.; Sadeddine Zennouche, O. Differences in grooming behavior between susceptible and resistant honey bee colonies after 13 Years of natural selection. Apidologie 2020, 51, 793–801. [Google Scholar] [CrossRef]
- Weiger, W.A. Serotonergic modulation of behaviour: A phylogenetic overview. Biol. Rev. Camb. Philos. Soc. 1997, 72, 61–95. [Google Scholar] [CrossRef] [PubMed]
- Dag, U.; Nwabudike, I.; Kang, D.; Gomes, M.A.; Kim, J.; Atanas, A.A.; Bueno, E.; Estrem, C.; Pugliese, S.; Wang, Z.; et al. Dissecting the functional organization of the C. elegans serotonergic system at Wwhole-brain scale. bioRxiv 2023, 186, 2574–2592. [Google Scholar] [CrossRef] [PubMed]
- Walz, B.; Baumann, O.; Krach, C.; Baumann, A.; Blenau, W. The aminergic control of cockroach salivary glands. Arch. Insect Biochem. Physiol. 2006, 62, 141–152. [Google Scholar] [CrossRef]
- Colas, J.F.; Launay, J.M.; Kellermann, O.; Rosay, P.; Maroteaux, L. Drosophila 5-HT2 serotonin receptor: Coexpression with fushi-tarazu during segmentation. Proc. Natl. Acad. Sci. USA 1995, 92, 5441–5445. [Google Scholar] [CrossRef]
- Yuan, Q.; Lin, F.; Zheng, X.; Sehgal, A. Serotonin Modulates Circadian Entrainment in Drosophila. Neuron 2005, 47, 115–127. [Google Scholar] [CrossRef]
- Sitaraman, D.; Zars, M.; LaFerriere, H.; Chen, Y.-C.; Sable-Smith, A.; Kitamoto, T.; Rottinghaus, G.E.; Zars, T. Serotonin is necessary for place memory in Drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 5579–5584. [Google Scholar] [CrossRef]
- Coray, R.; Quednow, B.B. The role of serotonin in declarative memory: A systematic review of animal and human research. Neurosci. Biobehav. Rev. 2022, 139, 104729. [Google Scholar] [CrossRef]
- Schürmann, F.W.; Klemm, N. Serotonin-immunoreactive neurons in the brain of the honeybee. J. Comp. Neurol. 1984, 225, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Seidel, C.; Bicker, G. The developmental expression of serotonin-immunoreactivity in the brain of the pupal honeybee. Tissue Cell 1996, 28, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Schulz, D.J.; Robinson, G.E. Biogenic amines and division of labor in honey bee colonies: Behaviorally related changes in the antennal lobes and age-related changes in the mushroom bodies. J. Comp. Physiol. A 1999, 184, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Thamm, M.; Balfanz, S.; Scheiner, R.; Baumann, A.; Blenau, W. Characterization of the 5-HT1A receptor of the honeybee (Apis Mellifera) and involvement of serotonin in phototactic behavior. Cell Mol. Life Sci. 2010, 67, 2467–2479. [Google Scholar] [CrossRef] [PubMed]
- Perrot-Minnot, M.-J.; Dion, E.; Cézilly, F. Modulatory effects of the serotonergic and histaminergic systems on Light in the crustacean Gammarus Pulex. Neuropharmacology 2013, 75, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Chou, V.T.; Johnson, S.A.; Van Vactor, D. Synapse development and maturation at the drosophila neuromuscular junction. Neural Dev. 2020, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Davanger, S.; Manahan-Vaughan, D.; Mulle, C.; Storm-Mathisen, J.; Ottersen, O.P. Protein trafficking, targeting, and interaction at the glutamate synapse. Neuroscience 2009, 158, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Xie, W. Cell adhesion molecules in Drosophila synapse development and function. Sci. China Life Sci. 2012, 55, 20–26. [Google Scholar] [CrossRef]
- Wu, S.; Guo, C.; Zhao, H.; Sun, M.; Chen, J.; Han, C.; Peng, Q.; Qiao, H.; Peng, P.; Liu, Y.; et al. Drosulfakinin signaling in fruitless circuitry antagonizes P1 neurons to regulate sexual arousal in Drosophila. Nat. Commun. 2019, 10, 4770. [Google Scholar] [CrossRef]
- Dockray, G.J. Cholecystokinin and gut-brain signalling. Regul. Pept. 2009, 155, 6–10. [Google Scholar] [CrossRef]
- Rehfeld, J.F.; Friis-Hansen, L.; Goetze, J.P.; Hansen, T.V.O. The biology of cholecystokinin and gastrin peptides. Curr. Top. Med. Chem. 2007, 7, 1154–1165. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Deng, X.; Singh, P. Significant Increase in the aggressive behavior of transgenic mice overexpressing peripheral progastrin peptides: Sssociated changes in CCK2 and serotonin receptors in the CNS. Neuropsychopharmacology 2007, 32, 1813–1821. [Google Scholar] [CrossRef] [PubMed]
- Nässel, D.R.; Williams, M.J. Cholecystokinin-like peptide (DSK) in Drosophila, not only for satiety signaling. Front. Endocrinol. 2014, 5, 219. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Peterson, J.; Nachman, R.J.; Ganetzky, B. Drosulfakinin activates CCKLR-17D1 and promotes larval locomotion and escape response in Drosophila. Fly 2012, 6, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Farah, C.S.; Reinach, F.C. The Troponin complex and regulation of muscle contraction. FASEB J. 1995, 9, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Filatov, V.L.; Katrukha, A.G.; Bulargina, T.V.; Gusev, N.B. Troponin: Structure, properties, and mechanism of functioning. Biochemistry 1999, 64, 969–985. [Google Scholar] [PubMed]
- Lehman, W.; Bullard, B.; Hammond, K. Calcium-dependent myosin from insect flight muscles. J. Gen. Physiol. 1974, 63, 553–563. [Google Scholar] [CrossRef]
- Vibert, P.; Craig, R.; Lehman, W. Steric-model for activation of muscle thin filaments. J. Mol. Biol. 1997, 266, 8–14. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, J.; Wan, K.; Lü, Y.; Ouyang, W.; Huang, J.; Zheng, L.; Miao, L.; Su, S.; Li, Z. Comparison of Brain Gene Expression Profiles Associated with Auto-Grooming Behavior between Apis cerana and Apis mellifera Infested by Varroa destructor. Genes 2024, 15, 763. https://doi.org/10.3390/genes15060763
Liao J, Wan K, Lü Y, Ouyang W, Huang J, Zheng L, Miao L, Su S, Li Z. Comparison of Brain Gene Expression Profiles Associated with Auto-Grooming Behavior between Apis cerana and Apis mellifera Infested by Varroa destructor. Genes. 2024; 15(6):763. https://doi.org/10.3390/genes15060763
Chicago/Turabian StyleLiao, Jiali, Kunlin Wan, Yang Lü, Wenyao Ouyang, Jingnan Huang, Liyuan Zheng, Liuchang Miao, Songkun Su, and Zhiguo Li. 2024. "Comparison of Brain Gene Expression Profiles Associated with Auto-Grooming Behavior between Apis cerana and Apis mellifera Infested by Varroa destructor" Genes 15, no. 6: 763. https://doi.org/10.3390/genes15060763
APA StyleLiao, J., Wan, K., Lü, Y., Ouyang, W., Huang, J., Zheng, L., Miao, L., Su, S., & Li, Z. (2024). Comparison of Brain Gene Expression Profiles Associated with Auto-Grooming Behavior between Apis cerana and Apis mellifera Infested by Varroa destructor. Genes, 15(6), 763. https://doi.org/10.3390/genes15060763