
Orthologs at the Base of the Olfactores Clade

Abstract

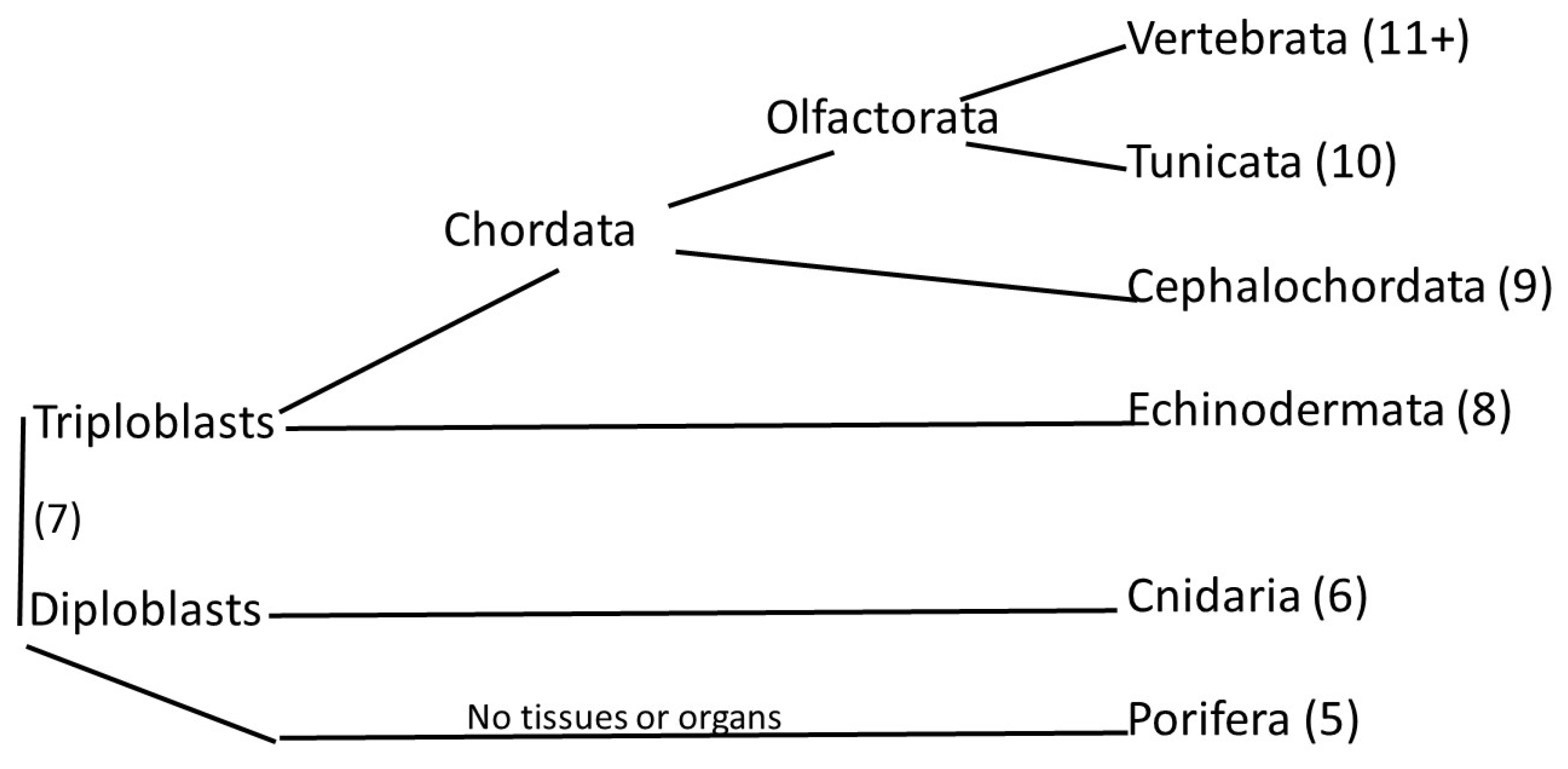
1. Introduction
2. Methods
3. Results and Discussion
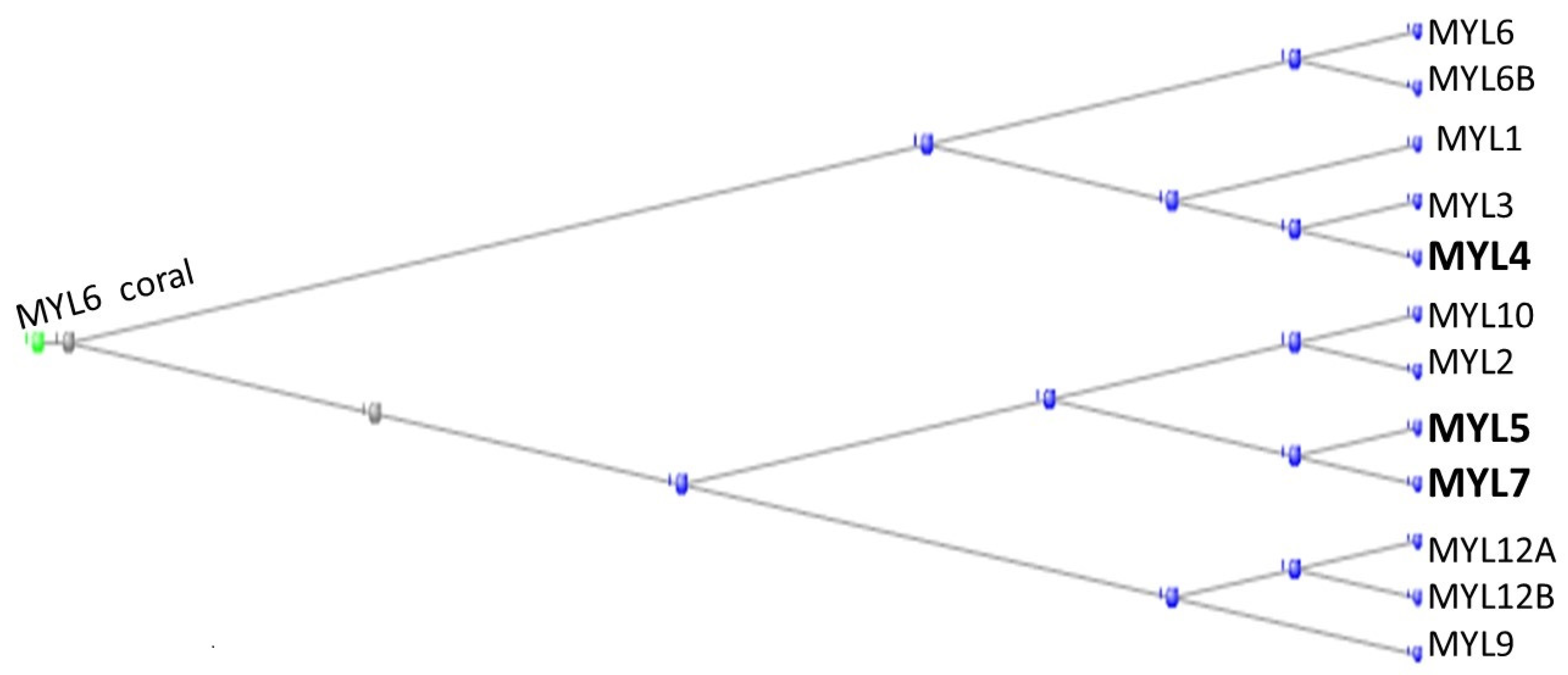
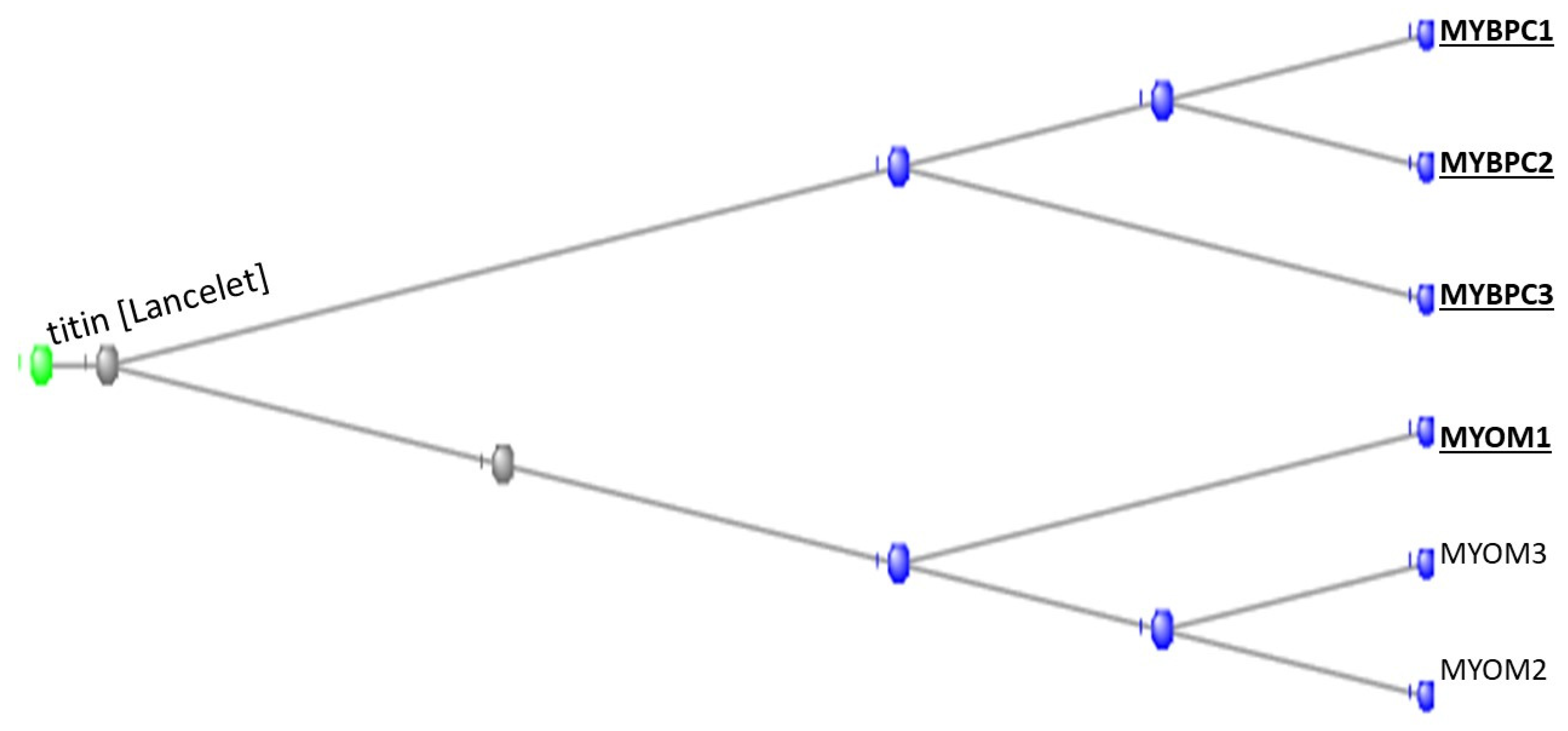
3.1. The Muscle Protein Genes
The Proteins of Muscle
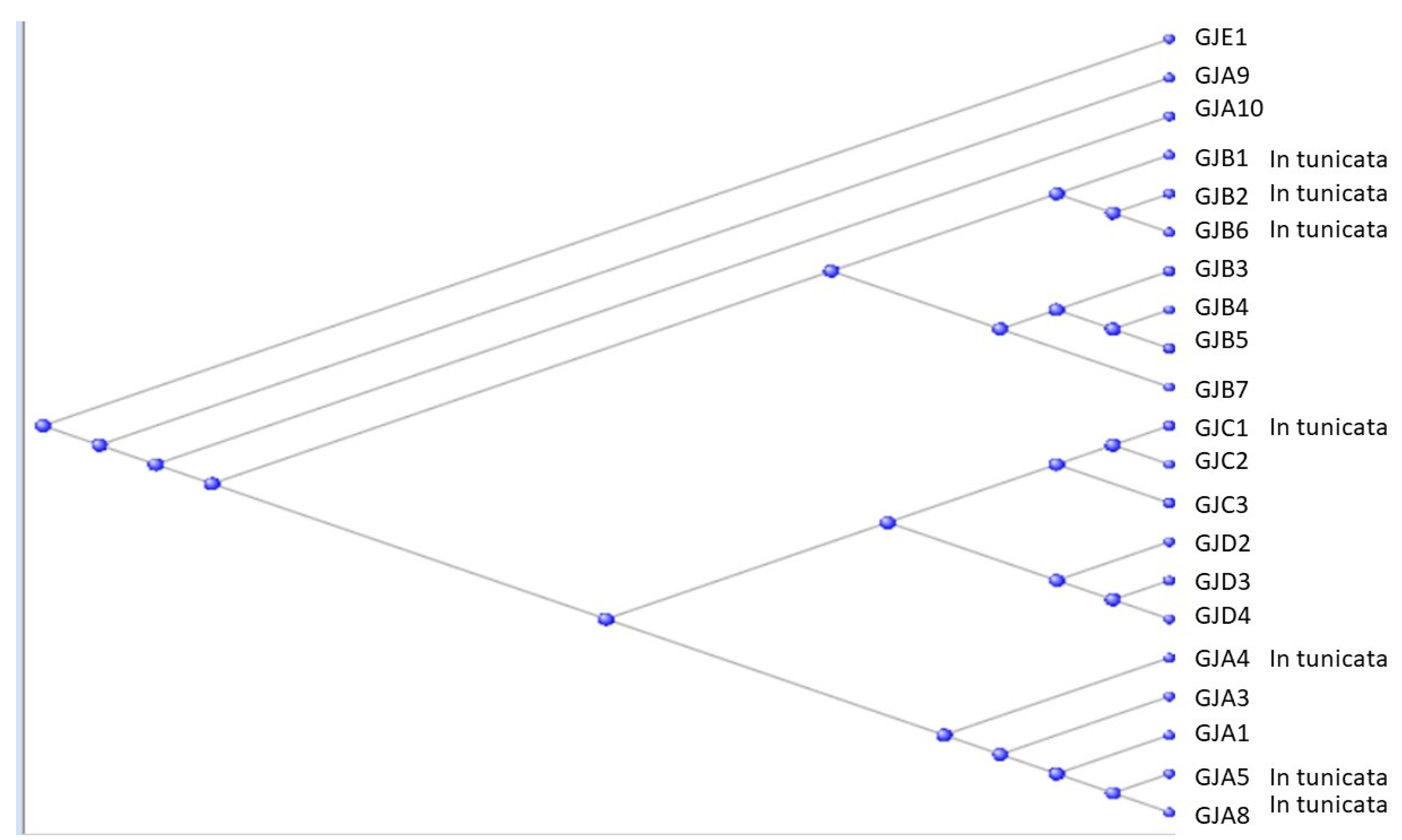
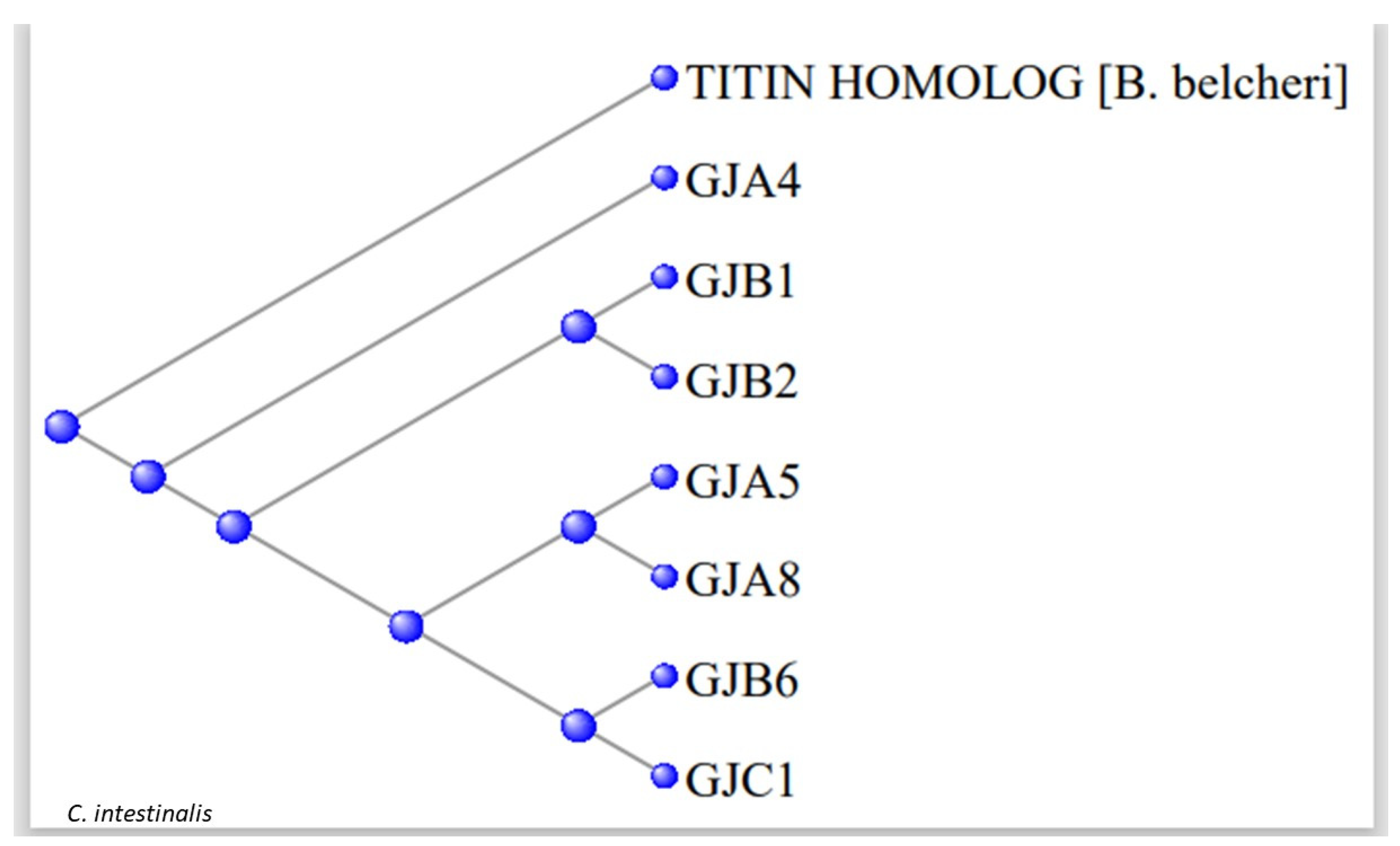
3.2. The Gap Junction Proteins
3.3. The Cadherins
3.4. The Claudins
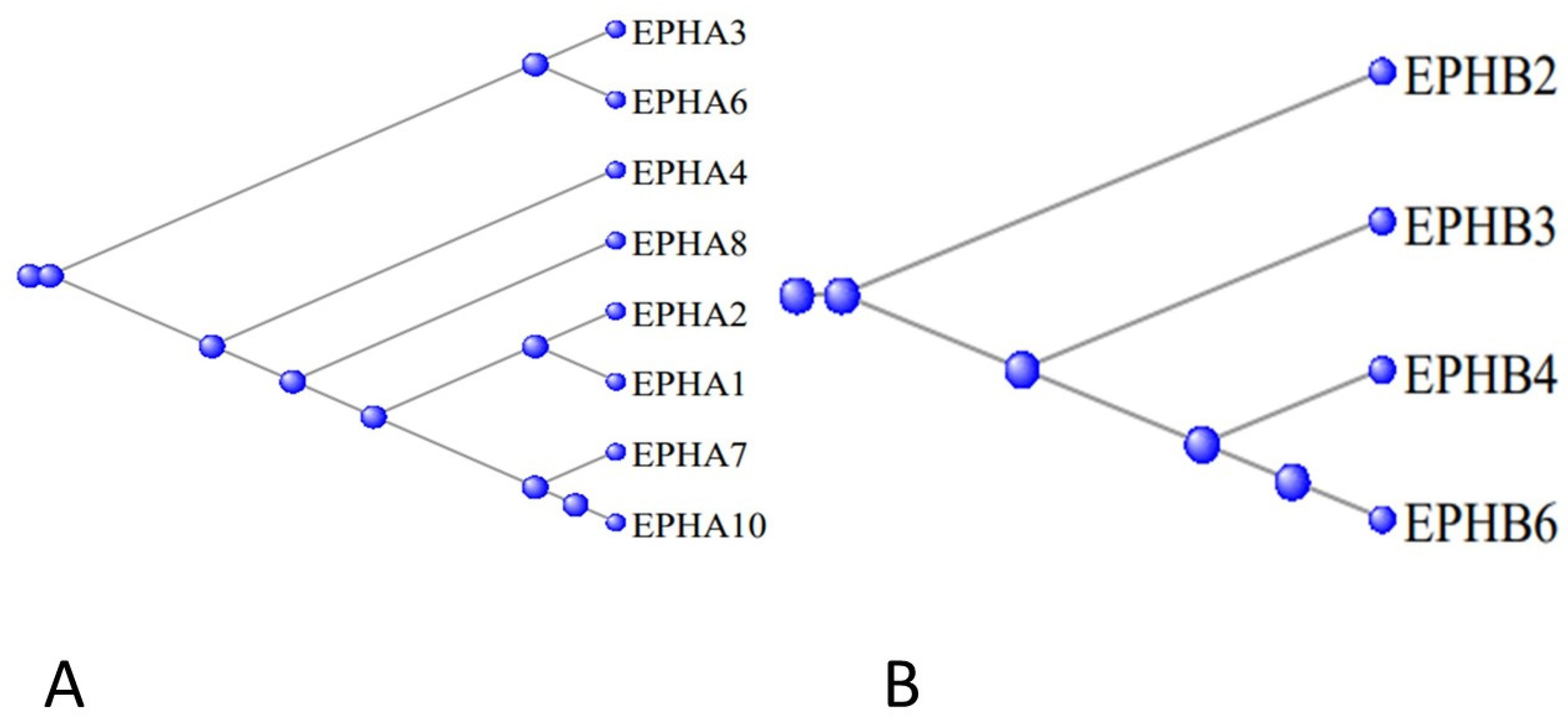
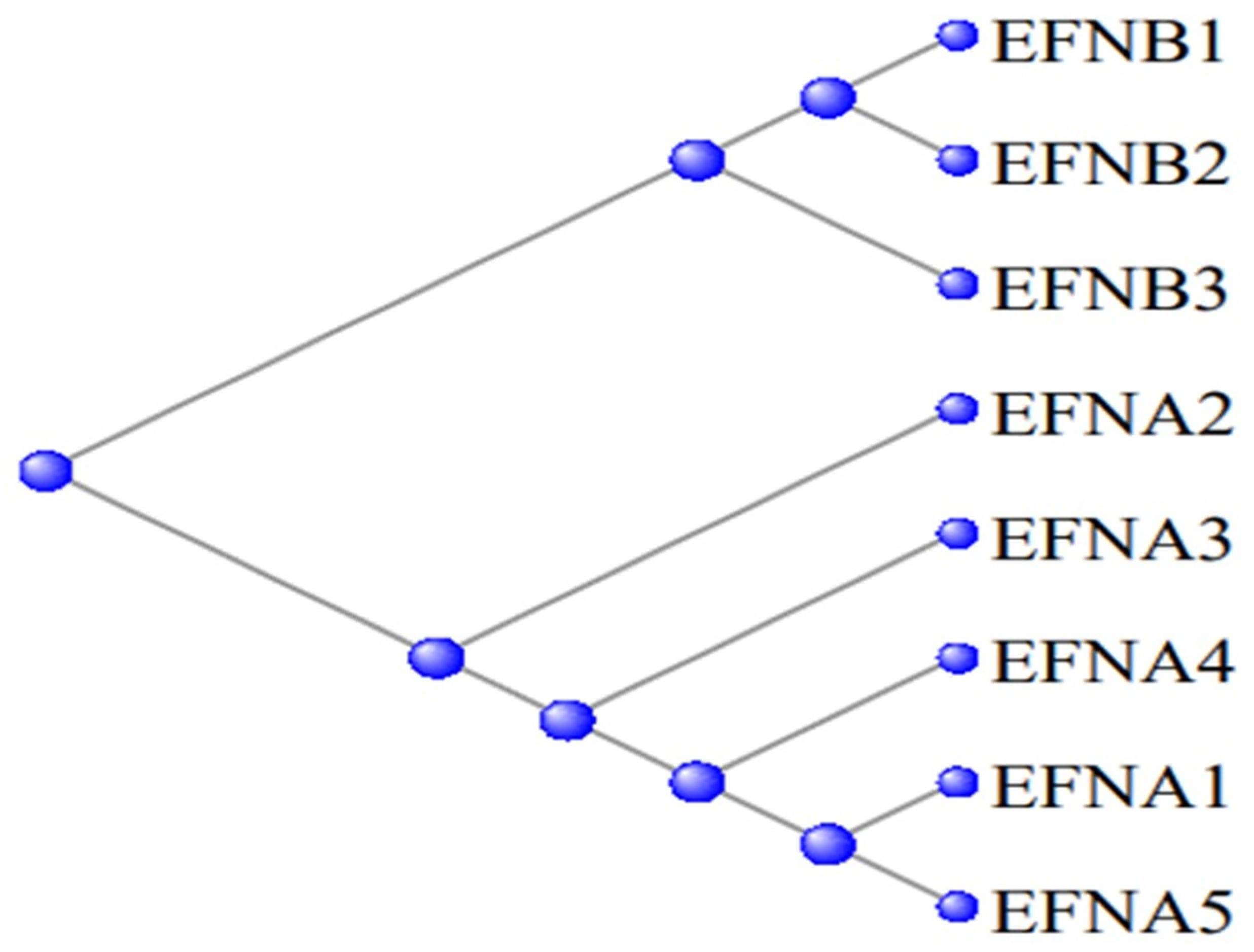
3.5. The Ephrons and Ephrins
3.6. The MAGE Genes
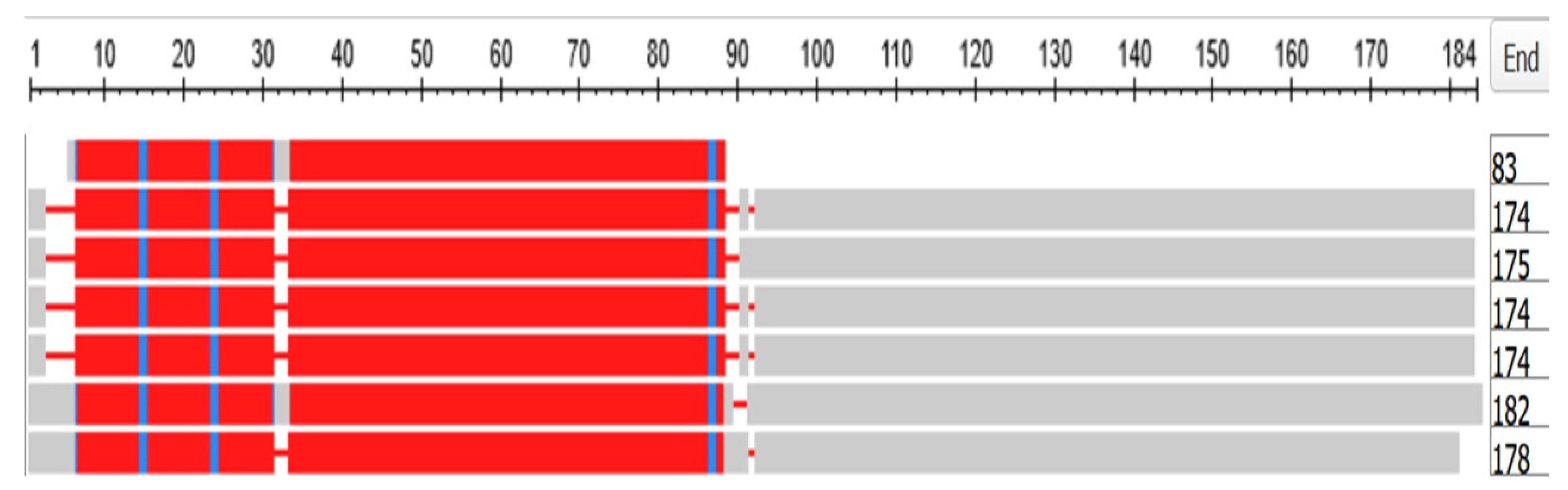
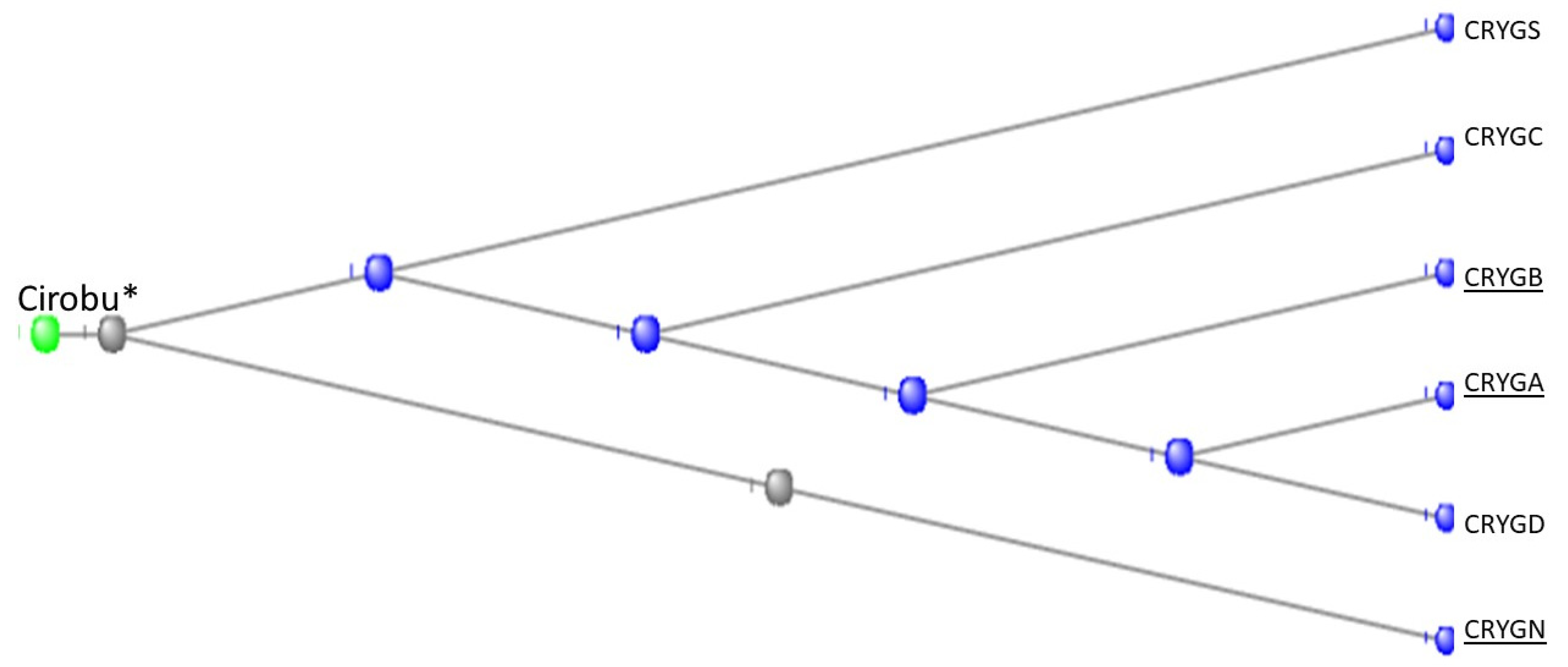
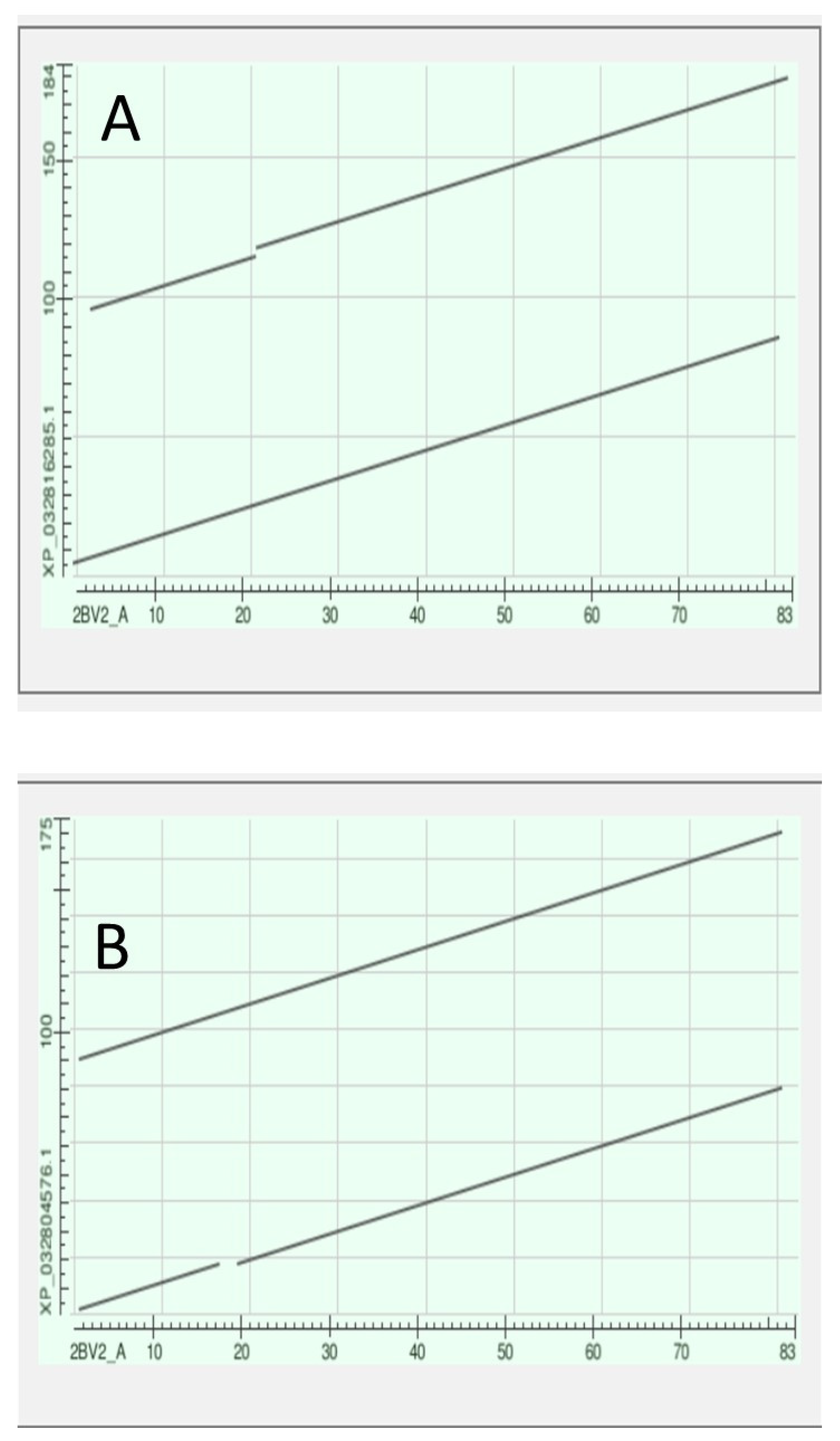
3.7. The Crystallins
The Gamma Crystallins
3.8. The Distal-Less Genes
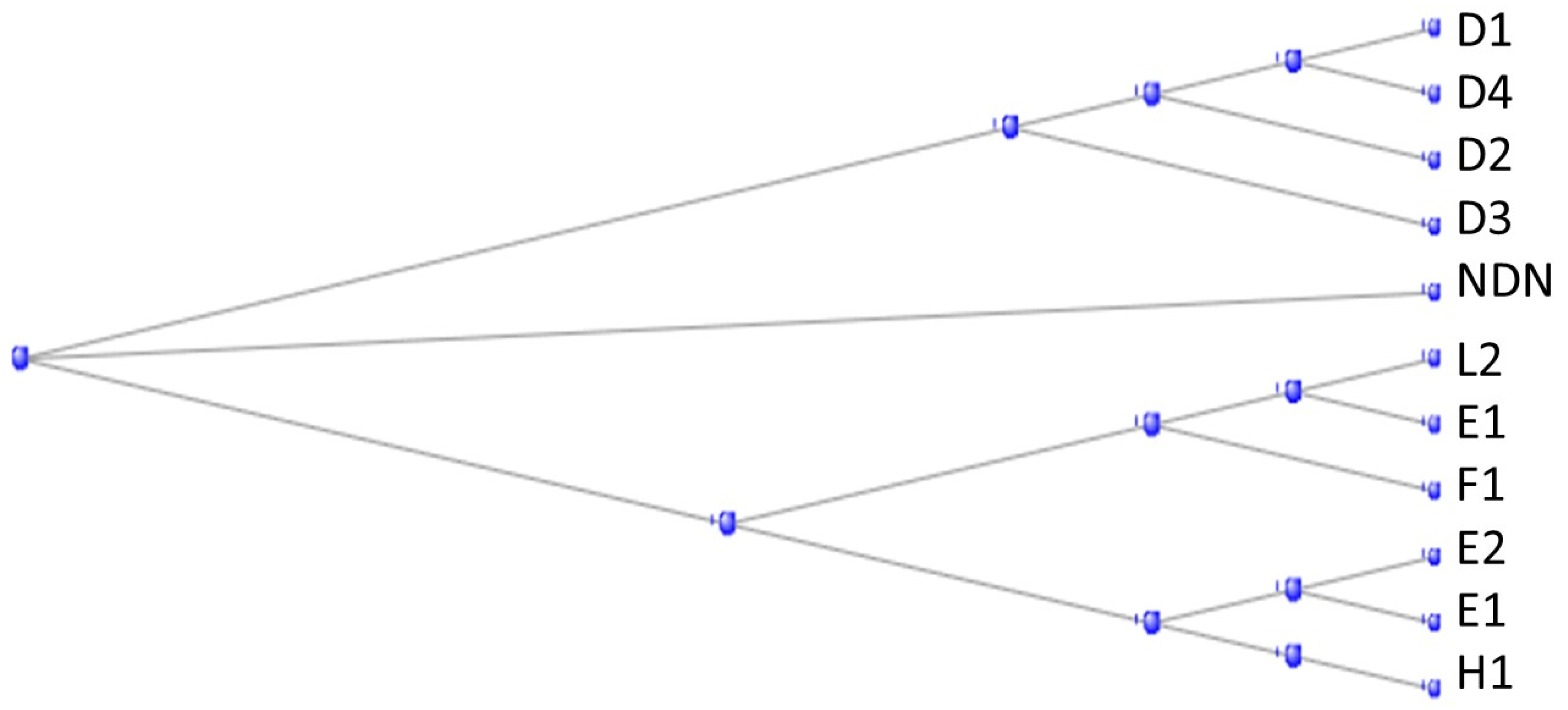
3.9. DAVID Analysis of Individual Orthologs Not Discussed in Previous Subsections
3.10. Tunicate Orthologs That Are Non-Coding Genes
4. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Domazet-Lošo, T.; Tautz, D. Phylostratigraphic tracking of cancer genes suggests a link to the emergence of multicellularity in metazoa. BMC Biol. 2010, 8, 66. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalo-chordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Liebeskind, B.J.; McWhite, D.; Marcotte, E.M. Towards consensus gene ages. Genome Biol. Evol. 2016, 8, 1812–1823. [Google Scholar] [CrossRef] [PubMed]
- Litman, T.; Stein, W.D. Obtaining estimates for the ages of all the protein-coding genes and most of the ontology-identified noncoding genes of the human genome, assigned to 19 phylostrata. Sem. Oncol. 2019, 46, 3–9. [Google Scholar] [CrossRef]
- Colgren, J.; Nichols, S.A. MRTF specifies a muscle-like contractile module in Porifera. Nat. Commun. 2022, 13, 4134. [Google Scholar] [CrossRef] [PubMed]
- Zullo, L.; Bozzo, M.; Daya, A.; Di Clemente, A.; Mancini, F.P.; Megighian, A.; Nesher, N.; Röttinger, E.; Shomrat, T.; Tiozzo, S.; et al. The diversity of muscle s and their regenerative potential across animals. Cells 2020, 9, 1925. [Google Scholar] [CrossRef]
- Jahnel, S.M.; Walz, M.; Technau, U. Development and epithelial organisation of muscle cells in the sea anemone Nematostella vectensis. Front. Zoolog. 2014, 11, 44. [Google Scholar] [CrossRef]
- Wilkie, I.C.; Candia Carnevali, M.D.; Andrietti, F. Mechanical properties of sea-urchin lantern muscles: A comparative investigation of intact muscle groups in Paracentrotus lividus (Lam.) and Stylocidaris affinis (Phil.) (Echinodermata, Echinoidea). J. Comp. Physiol. B 1998, 168, 204–212. [Google Scholar] [CrossRef]
- Hiebert, T.C.; Gemmell, B.J.; von Dassow, G.; Conley, K.R.; Sutherland, K.R. The hydrodynamics and kinematics of the appendicularian tail underpin peristaltic pumping. J. R. Soc. Interface 2023, 20, 20230404. [Google Scholar] [CrossRef]
- Lindskog, C.; Linné, J.; Fagerberg, L.; Hallström, B.M.; Sundberg, C.J.; Lindholm, M.; Huss, M.; Kampf, C.; Choi, H.; Liem, D.A.; et al. The human cardiac and skeletal muscle proteomes defined by transcriptomics and antibody-based profiling. BMC Genom. 2015, 16, 475. [Google Scholar] [CrossRef]
- Schoenauer, R.; Bertoncini, P.; Machaidze, G.; Aebi, U.; Perriard, J.C.; Hegner, M.; Agarkova, I. Myomesin is a molecular spring with adaptable elasticity. J. Mol. Biol. 2005, 349, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Abascal, F.; Zardoya, R. Evolutionary analyses of gap junction protein families. Biochim. Biophys. Acta 2013, 1828, 4–14. [Google Scholar] [CrossRef]
- Lucaciu, S.A.; Leighton, S.E.; Hauser, A.; Yee, R.; Laird, D.W. Diversity in connexin biology. J. Biol. Chem. 2023, 299, 105263. [Google Scholar] [CrossRef] [PubMed]
- Purves, D.; Augustine, G.J.; Fitzpatrick, D.; Katz, L.C.; LaMantia, A.-S.; McNamara, J.O.; Williams, S.M. (Eds.) Neuroscience, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2001. [Google Scholar]
- More, H.L.; OʼConnor, S.M.; Brøndum, E.; Wang, T.; Bertelsen, M.F.; Grøndahl, C.; Kastberg, K.; Hørlyck, A.; Funder, J.; Maxwell Donelan, J. Sensorimotor responsiveness and resolution in the giraffe. J. Exp. Biol. 2013, 216, 1003–1011. [Google Scholar] [CrossRef]
- Weil, M.T.; Heibeck, S.; Topperwien, M.; tom Dieck, S.; Ruhwedel, T.; Salditt, T.; Rodicio, M.C.; Morgan, J.R.; Nave, K.A.; Mobius, W.; et al. Axonal ensheathment in the nervous system of lamprey: Implications for the evolution of myelinating glia. J. Neurosci. 2018, 38, 6586–6596. [Google Scholar] [CrossRef]
- Gould, R.M.; Morrison, H.G.; Gilland, E.; Campbell, R.K. Myelin tetraspan family proteins but no non-tetraspan family proteins are present in the ascidian (C. intestinalis) genome. Biol. Bull. 2005, 209, 49–66. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.D.; Ciatto, C.; Chen, C.P.; Bahna, F.; Rajebhosale, M.; Arkus, N.; Schieren, I.; Jessell, T.M.; Honig, B.; Price, S.R.; et al. Type II cadherin ectodomain structures: Implications for classical cadherin specificity. Cell 2006, 124, 1255–1268. [Google Scholar] [CrossRef]
- Taneyhill, L.A. To adhere or not to adhere: The role of cadherins in neural crest development. Cell Adhes. Migr. 2008, 2, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Oda, H.; Akiyama-Oda, Y.; Zhang, S. Two classic cadherin-related molecules with no cadherin extracellular repeats in the cephalochordate amphioxus: Distinct adhesive specificities and possible involvement in the development of multicell-layered structures. J. Cell Sci. 2003, 117, 2757–2767. [Google Scholar] [CrossRef]
- York, J.R.; McCauley, D.W. The origin and evolution of vertebrate neural crest cells. Open Biol. 2020, 10, 190285. [Google Scholar] [CrossRef]
- Noda, T.; Satoh, N. A comprehensive survey of cadherin superfamily gene expression patterns in C. intestinalis. Gene Expr. Patterns 2008, 8, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, W.R.; Chiba, T.; Krajka, F.R.; Deyts, C.; Satoh, N.; Joly, J.S. Trunk lateral cells are neural crest-like cells in the ascidian C. intestinalis: Insights into the ancestry and evolution of the neural crest. Dev. Biol. 2008, 324, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Niessen, C.M.; Leckband, D.; Yap, A.S. Tissue organization by cadherin adhesion molecules: Dynamic molecular and cellular mechanisms of morphogenetic regulation. Physiol. Rev. 2011, 91, 691–731. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.C.; Nazarko, O.V.; Sando, I.I.I.R.; Salzman, G.S.; Li, N.S.; Südhof, T.C.; Araç, D. Structural basis of latrophilin-FLRT-UNC5 interaction in cell adhesion. Structure 2015, 23, 1678–1691. [Google Scholar] [CrossRef] [PubMed]
- del Toro, D.; Carrasquero-Ordaz, M.A.; Chu, A.; Ruff, T.; Shahin, M.; Jackson, V.A.; Chavent, M.; Berbeira-Santana, M.; Seyit-Bremer, G.; Brignani, S.; et al. Structural basis of teneurin-latrophilin interaction in repulsive guidance of migrating neurons. Cell 2020, 180, 323–339. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M.; Fujita, K.; Hiiragi, T.; Fujimoto, K.; Tsukita, S. Claudin-1 and -2: Novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin. J. Cell Biol. 1998, 141, 1539–1550. [Google Scholar] [CrossRef] [PubMed]
- Günzel, D.; Yu, A.S.L. Claudins and the modulation of tight junction permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef]
- Kollmar, R.; Nakamura, S.K.; Kappler, J.A.; Hudspeth, A.J. Expression and phylogeny of claudins in vertebrate primordia. Proc. Natl. Acad. Sci. USA 2001, 98, 10196–10201. [Google Scholar] [CrossRef] [PubMed]
- Mellott, D.O.; Burke, R.D. The molecular phylogeny of eph receptors and ephrin ligands. BMC Cell Biol. 2008, 9, 1–8. [Google Scholar] [CrossRef]
- Taylor, H.; Campbell, J.; Nobes, C.D. Ephs and ephrins. Curr. Biol. 2017, 27, R83–R102. [Google Scholar] [CrossRef]
- Williaume, G.; de Buyl, S.; Sirour, C.; Haupaix, N.; Bettoni, R.; Imai, K.S.; Satou, Y.; Dupont, G.; Hudson, C.; Yasuo, H. Cell geometry, signal dampening, and a bimodal transcriptional response underlie the spatial precision of an ERK-mediated embryonic induction. Dev. Cell 2021, 56, 2966–2979. [Google Scholar] [CrossRef] [PubMed]
- Fiuza, U.M.; Negishi, T.; Rouan, A.; Yasuo, H.; Lemaire, P. A Nodal/Eph signalling relay drives the transition from apical constriction to apico-basal shortening in ascidian endoderm invagination. Development 2020, 147, dev186965. [Google Scholar] [CrossRef]
- Negishi, T.; Nishida, H. Asymmetric and Unequal Cell Divisions in Ascidian Embryos. In Asymmetric Cell Division in Development, Differentiation and Cancer. Results and Problems in Cell Differentiation; Tassan, J.P., Kubiak, J., Eds.; Springer: Cham, Switzerland, 2017; Volume 61. [Google Scholar] [CrossRef]
- Haupaix, N.; Abitua, P.B.; Sirour, C.; Yasuo, H.; Levine, M.; Hudson, C. Ephrin-mediated restriction of ERK1/2 activity delimits the number of pigment cells in the C. CNS. Dev. Biol. 2014, 394, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Stolfi, A.; Wagner, E.; Taliaferro, J.M.; Chou, S.; Levine, M. Neural tube patterning by Ephrin, FGF and Notch signaling relays. Development 2011, 138, 5429–5439. [Google Scholar] [CrossRef]
- Gee, R.R.F.; Chen, H.; Lee, A.K.; Daly, C.A.; Wilander, B.A.; Fon Tacer, K.; Potts, P.R. Emerging roles of the MAGE protein family in stress response pathways. J. Biol. Chem. 2020, 295, 16121–16155. [Google Scholar]
- Ray, M.E.; Wistow, G.; Su, Y.A.; Meltzer, P.S.; Trent, J.M. AIM1, a novel non-lens member of the bg-crystallin superfamily, is associated with the control of tumorigenicity in human malignant melanoma. Proc. Natl. Acad. Sci. USA 1997, 94, 3229–3234. [Google Scholar] [CrossRef]
- Shimeld Sebastian, M.; Purkiss, A.G.; Dirks, R.P.H.; Bateman, O.A.; Slingsby, C.; Lubsen, N.H. Urochordate-crystallin and the evolutionary origin of the vertebrate eye lens. Curr. Biol. 2005, 15, 1684–1689. [Google Scholar] [CrossRef]
- Riyahi, K.; Shimeld, S.M. Chordate βγ-crystallins and the evolutionary developmental biology of the vertebrate lens. Comp. Biochem. Physiol. Part B 2007, 147, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Kappe, G.; Purkiss, A.G.; van Genesen, S.T.; Slingsby, C.; Lubsen, N.H. Explosive expansion of bc-crystallin genes in the ancestral vertebrate. J. Mol. Evol. 2010, 71, 219–230. [Google Scholar] [CrossRef]
- Slingsby, C.; Wistow, G.J.; Clark, A.R. Evolution of crystallins for a role in the vertebrate eye lens. Prot. Sci. 2013, 22, 367–380. [Google Scholar] [CrossRef]
- Cvekl, A.; Zhao, Y.; Mcgreal Rebecca Xie, Q.; Gu, X.; Zheng, D. Evolutionary origins of Pax6 control of crystallin genes. Genome Biol. Evol. 2017, 9, 2075–2092. [Google Scholar] [CrossRef] [PubMed]
- Zenga, F.; Wunderera, J.; Salvenmosera, W.; Hessb, M.W.; Ladurnera, P.; Rothbächera, U. Papillae revisited and the nature of the adhesive secreting collocytes. Dev. Biol. 2019, 448, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Sakurai, D.; Ohtsuki, H.; Terakita, A.; Shichida, Y.; Usukura, J.; Kusakabe, T.; Tsuda, M. Pigmented and nonpigmented ocelli in the brain vesicle of the ascidian larva. J. Comp. Neur. 2008, 509, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Kozlyuk, N.; Sengupta, S.; Bierma, J.C.; Martin, R.W. Calcium binding dramatically stabilizes an ancestral crystallin fold in tunicate βγ-crystallin. Biochemistry 2016, 55, 6961–6968. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M.; Bronner, G.; Kuttner, F.; Jurgens, G.; Jackie, H. Distal-less encodes a homoeodomain protein required for limb development in Drosophila. Nature 1989, 338, 432–434. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.A.; Nulsen, C.; Nagy, L.M. A complex role for distal-less in crustacean appendage development. Dev. Biol. 2002, 241, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Hammid, A.; Umair, M.; Ahmad, W. A novel heterozygous intragenic sequence variant in DLX6 probably underlies first case of autosomal dominant split-hand/foot malformation type 1. Mol. Syndromol. 2017, 8, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Arendt, D. Many ways to build a polyp. Trends Gen. 2019, 35, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Lowe, C.J.; Wray, G.A. Radical alterations in the roles of homeobox genes during echinoderm evolution. Nature 1997, 389, 718–721. [Google Scholar] [CrossRef]
- Prpic, N.M.; Tautz, D. The expression of the proximodistal axis patterning genes Distal-less and dachshund in the appendages of Glomeris marginata (Myriapoda: Diplopoda) suggests a special role of these genes in patterning the head appendages. Dev. Biol. 2003, 260, 97–112. [Google Scholar] [CrossRef]
- Caracciolo, A.; Di Gregorio, A.; Aniello, F.; Di Lauro, R.; Branno, M. Identification and developmental expression of three Distal-less homeobox containing genes in the ascidian C. intestinalis. Mech. Dev. 2000, 99, 173–176. [Google Scholar] [CrossRef]
- Holland, N.D.; Panganiban, G.; Henyey, E.L.; Holland, L.Z. Sequence and developmental expression of amphidll, an amphioxus Distal-less gene transcribed in the ectoderm, epidermis and nervous system: Insights into evolution of craniate forebrain and neural crest. Development 1996, 122, 2911–2920. [Google Scholar] [CrossRef]
- Shyamala, K.; Yanduri, S.; Girish, H.C.; Murgod, S. Neural crest: The fourth germ layer. J. Oral. Maxillofac. Pathol. 2015, 19, 221–229. [Google Scholar] [CrossRef]
- Wang, S.; DeLeon, C.; Sun, W.; Quae, S.R.; Roth, B.L.; Südhof, T.C. Alternative splicing of latrophilin-3 controls synapse formation. Nature 2024, 626, 128–135. [Google Scholar] [CrossRef]
- Huber, P.A.J. Caldesmon. Int. J. Biochem. Cell Biol. 1997, 29, 1047–1051. [Google Scholar] [CrossRef] [PubMed]
- Polanco, J.; Reyes-Vigil, F.; Weisberg, S.D.; Dhimitruka, I.; Brusés, J.L. differential spatiotemporal expression of type i and type ii cadherins associated with the segmentation of the central nervous system and formation of brain nuclei in the developing mouse. Front. Mol. Neurosci. 2021, 14, 2021. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Munro, E. Differential expression of a classic cadherin directs tissue-level contractile asymmetry during neural tube closure. Dev. Cell 2019, 51, 158–172. [Google Scholar] [CrossRef] [PubMed]
- Paulson, A.F.; Prasad, M.S.; Thuringer, A.H.; Manzerra, P. Regulation of cadherin expression in nervous system development. Cell Adhes. Migr. 2014, 8, 19–28. [Google Scholar] [CrossRef]
- Matsuoka, H.; Yamaoka, A.; Hamashima, T.; Shima, A.; Kosako, M.; Tahara, Y.; Kamishikiryo, J.; Michihara, A. EGF-dependent activation of ELK1 contributes to the induction of CLDND1 expression involved in tight junction formation. Biomedicines 2022, 10, 1792. [Google Scholar] [CrossRef]
- Exposito, J.Y.; Cluzel, C.; Garrone, R.; Lethias, C. Evolution of collagens. Anatom. Rec. 2002, 268, 302–316. [Google Scholar] [CrossRef]
- Panganiban, G.; Rubenstein, J.L.R. Developmental functions of the Distal-less/Dlx homeobox genes. Development 2002, 129, 4371–4386. [Google Scholar] [CrossRef] [PubMed]
- Stolfi, A.; Gainous, T.B.; Young, J.J.; Mori, A.; Levine, M.; Christiaen, L. Early chordate origins of the vertebrate second heart field. Science 2010, 329, 565–568. [Google Scholar] [CrossRef]
- Moser, C.; Gosselé, K.A.; Balaz, M.; Balazova, L.; Horvath, C.; Künzle, P.; Okreglicka, K.M.; Li, F.; Blüher, M.; Stierstorferf, B.; et al. FAM3D: A gut secreted protein and its potential in the regulation of glucose metabolism. Peptides 2023, 167, 171047. [Google Scholar] [CrossRef]
- Chen, W.; Gao, D.; Xie, L.; Wang, A.; Zhao, H.; Guo, C.; Sun, Y.; Nie, Y.; Hong, A.; Xiong, S. SCF-FBXO24 regulates cell proliferation by mediating ubiquitination and degradation of PRMT6. Biochem. Biophys. Res. Comm. 2020, 530, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Satou, Y.; Tokuoka, M.; Oda-Ishii, I.; Tokuhiro, S.; Ishida, T.; Liu, B.; Iwamura, Y. A manually curated gene model set for an ascidian, C. robusta (C. intestinalis type A). Zool. Sci. 2022, 39, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.M.; Khairallah, S.M.; Nguyen, A.H.; Newman-Smith, E.; Smith, W.C. Misregulation of cell adhesion molecules in the C. neural tube closure mutant bugeye. Dev. Biol. 2021, 480, 14–24. [Google Scholar] [CrossRef]
- Abitua, P.B.; Gainous, T.B.; Kaczmarczyk, A.N.; Winchell, C.J.; Hudson, C.; Kamata, K.; Nakagawa, M.; Tsuda, M.; Kusakabe, T.G.; Levine, M. The pre-vertebrate origins of neurogenic placodes. Nature 2015, 524, 462–465. [Google Scholar] [CrossRef]
- Edvardsen, R.B.; Seo, H.C.; Jensen, M.F.; Mialon, A.; Mikhaleva, J.; Bjordal, M.; Cartry, J.; Reinhardt, R.; Weissenbach, J.; Wincker, P.; et al. Remodelling of the homeobox gene complement in the tunicate Oikopleura dioica. Curr. Biol. 2005, 15, R12–R13. [Google Scholar] [CrossRef]
- Labat-de-Hoz, L.; Rubio-Ramos, A.; Correas, I.; Alonso, M.A. The MAL family of proteins: Normal function, expression in cancer, and potential use as cancer biomarkers. Cancers 2023, 15, 2801. [Google Scholar] [CrossRef]
- Chen, Y.H.; Pai, C.W.; Huang, S.W.; Chang, S.N.; Lin, L.Y.; Chiang, F.T.; Lin, J.L.; Hwang, J.J.; Tsai, C.T. Inactivation of myosin binding protein C homolog in zebrafish as a model for human cardiac hypertrophy and diastolic dysfunction. J. Am. Heart Assoc. 2013, 2, e000231. [Google Scholar] [CrossRef]
- Razy-Krajka, F.; Stolf, A. Regulation and evolution of muscle development in tunicates. EvoDevo 2019, 10, 13. [Google Scholar] [CrossRef]
- Heissler, S.M.; Sellers, J.R. Myosin light chains: Teaching old dogs new tricks. Bioarchitecture 2014, 4, 169–188. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, I.; Gholkar, A.A.; Velasquez, E.F.; Guo, X.; Tofig, B.; Damoiseaux, R.; Torres, J.Z. The myosin regulatory light chain Myl5 localizes to mitotic spindle poles and is required for proper cell division. Cytoskeleton 2021, 78, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, J.F.; Gillis, T.E. Evolution of the regulatory control of vertebrate striated muscle: The roles of troponin I and myosin binding protein-C. Physiol. Genom. 2010, 42, 406–419. [Google Scholar] [CrossRef]
- Langea, S.; Pinotsisc, N.; Agarkovad, I.; Ehlere, E. The M-band: The underestimated part of the sarcomere. BBA-Mol. Cell Res. 2020, 1867, 118440. [Google Scholar] [CrossRef] [PubMed]
- Du Pasquier, L. Speculations on the origin of the vertebrate immune system. Immunol. Lett. 2004, 92, 3–9. [Google Scholar] [CrossRef]
- Juriloff, D.M.; Harris, M.J. Insights into the etiology of mammalian neural tube closure defects from developmental, genetic and evolutionary studies. J. Dev. Biol. 2018, 6, 22. [Google Scholar] [CrossRef]
- Medina-Martinez, O.; Shah, R.; Jamrich, M. Pitx3 controls multiple aspects of lens development. Dev. Dyn. 2009, 238, 2193–2201. [Google Scholar] [CrossRef]
- Scales, S.J.; Hesser, B.A.; Masuda, E.S.; Scheller, R.H. Amisyn, a novel syntaxin-binding protein that may regulate snare complex assembly. J. Biol. Chem. 2002, 277, 28271–28279. [Google Scholar] [CrossRef]
- Wei, B.; Jin, J.P. TNNT1, TNNT2, and TNNT3: Isoform genes, regulation, and structure-function relationships. Gene 2016, 582, 1–13. [Google Scholar] [CrossRef]
- Satou, Y.; Imai, K.S. Ascidian Zic Genes. In Zic Family. Advances in Experimental Medicine and Biology; Aruga, J., Ed.; Springer: Singapore, 2018; Volume 1046. [Google Scholar] [CrossRef]
- Chen, J.S.; Pedro, M.S.; Zeller, R.W. miR-124 function during C. intestinalis neuronal development includes extensive interaction with the Notch signaling pathway. Development 2011, 138, 4943–4953. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, S.; Cauteruccio, S.; Manenti, R.; Candiani, S.; Scarì, G.; Licandro, E.; Pennati, R. miR-7 Knockdown by Peptide Nucleic Acids in the Ascidian C. intestinalis. Int. J. Mol. Sci. 2019, 20, 5127. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HGNC | Steps | Equal to Mode | HGNC | Steps | Equal to Mode |
|---|---|---|---|---|---|
| ADGRL3 | 3.15 | 4 | MGAT4D | 4.08 | 4 |
| ANKRD16 | 4.77 | 4 | MSMP | 1.15 | 6 |
| ANKRD42 | 2.38 | 8 | MUM1 | 2.10 | 5 |
| ARHGEF38 | 2.67 | 4 | MYBPC3 | 2.08 | 6 |
| ASB2 | 3.92 | 5 | MYL5 | 5.01 | 3 |
| ATRAID | 0.46 | 11 | NEBL | 2.16 | 6 |
| BCL6B | 2.95 | 4 | NECTIN3 | 1.85 | 4 |
| C18ORF21 | 0.62 | 9 | NEXN | 2.46 | 5 |
| CALML6 | 2.47 | 8 | NPC1L1 | 4.85 | 5 |
| CASR | 1.92 | 7 | OLFML2A | 1.54 | 5 |
| CATIP | 3.18 | 4 | OTULIN | 1.55 | 7 |
| CBLN1 | 1.38 | 4 | PDX1 | 2.77 | 4 |
| CCDC78 | 2.62 | 4 | PHF21A | 2.46 | 4 |
| CEP63 | 3.40 | 5 | PODN | 3.46 | 4 |
| CEP83 | 2.46 | 7 | PRR14 | 2.27 | 6 |
| CKAP2 | 2.54 | 5 | RBM14 | 3.61 | 3 |
| CLDN1 | 0.83 | 7 | RHBDL2 | 4.54 | 4 |
| CLDN18 | 1.25 | 5 | RNF4 | 4.85 | 3 |
| CLDN19 | 0.92 | 7 | SCLT1 | 3.18 | 6 |
| CLDN7 | 1.23 | 6 | SERINC5 | 4.38 | 5 |
| EFNA1 | 1.92 | 4 | SLC35G2 | 3.31 | 4 |
| FABP2 | 1.00 | 8 | SLC35G3 | 4.86 | 3 |
| FAM155B | 1.54 | 5 | SLC35G4 | 5.13 | 3 |
| FAM187A | 1.23 | 6 | SLC35G5 | 5.50 | 3 |
| FAM3D | 2.38 | 4 | SLC35G6 | 4.25 | 4 |
| FBXO24 | 2.23 | 6 | SLC6A14 | 3.54 | 3 |
| FOXH1 | 2.95 | 5 | SLC6A6 | 3.46 | 4 |
| GJA10 | 2.02 | 5 | SNRNP48 | 2.31 | 8 |
| GJA3 | 1.16 | 7 | SPATA21 | 4.50 | 3 |
| GJC2 | 2.38 | 6 | STXBP6 | 1.77 | 8 |
| GNA15 | 4.00 | 3 | TGFB1 | 2.15 | 4 |
| GTF2IRD2 | 3.00 | 3 | TGFB2 | 1.54 | 6 |
| GTF2IRD2B | 4.30 | 3 | TLCD2 | 3.62 | 3 |
| GVQW1 | 4.05 | 2 | TMEM218 | 0.62 | 9 |
| H1FOO | 3.23 | 3 | TNNC1 | 4.08 | 6 |
| HMMR | 2.92 | 8 | TNNC2 | 4.08 | 4 |
| HNF1A | 1.15 | 7 | TSPAN12 | 2.69 | 5 |
| HSPB1 | 3.23 | 3 | WSB1 | 2.31 | 4 |
| IKZF5 | 2.85 | 5 | ZBED5 | 2.46 | 4 |
| IL17C | 1.78 | 4 | ZBED8 | 2.83 | 4 |
| KIZ | 1.23 | 6 | ZC3H7B | 1.38 | 8 |
| LRRC29 | 3.09 | 4 | ZNF91 | 4.88 | 2 |
| Familiar Name | Tissue Distribution | HGNC | Type | Has Tunicate Ortholog? |
|---|---|---|---|---|
| Cadherin E | Epithelial | CDH1 | I | Yes |
| Cadherin N | Neural | CDH2 | I | Yes |
| Cadherin P | Placental | CDH3 | I | |
| Cadherin R | Retinal | CDH4 | I | |
| Cadherin VE | Epithelial | CDH5 | II | |
| Cadherin K | Brain; kidney | CDH6 | II | |
| Cadherin OB | Osteoblast | CDH11 | II | Yes |
| Cadherin BR | Brain | CDH12 | II | |
| Cadherin T and H | Heart | CDH13 | I | |
| Cadherin M | Muscle | CDH15 | I |
| Ephron Genes | Ephrin Genes | ||
|---|---|---|---|
| HGNC Symbol | Latest Ortholog in: | HGNC Symbol | Latest Ortholog in: |
| EPHA2 | Tunicata | EFNA1 | Tunicata |
| EPHA3 | Elasmobranchii | EFNA2 | Tunicata |
| EPHA4 | Tunicata | EFNA3 | Tunicata |
| EPHA5 | Branchiostomata | EFNA4 | Tunicata |
| EPHA6 | Elasmobranchii | EFNA5 | Tunicata |
| EPHA7 | Elasmobranchii | EFNB1 | Tunicata |
| EPHA8 | Elasmobranchii | EFNB2 | Branchiostomato |
| EPHA10 | Osteichthyes | EFNB3 | Elasmobranchii |
| EPHB2 | Branchiostomata | ||
| EPHB3 | Elasmobranchii | ||
| EPHB4 | Cnidaria | ||
| Term | Genes | FDR * | |
|---|---|---|---|
| Cluster 2 | |||
| heterophilic cell-cell adhesion via plasma membrane cell adhesion molecules | NECTIN3, CBLN1, NECTIN1 | 0.358759 | |
| identical protein binding | TFAP2E, COL23A1, HSPB1, NECTIN3, CBLN1, NECTIN1 | 0.671885 | |
| Synapse | NECTIN3, CBLN1, NECTIN1 | 0.935728 | |
| Cluster 4 | |||
| homophilic cell adhesion via plasma membrane adhesion molecules | PALLD, NECTIN3, NECTIN1 | 0.739434 | |
| DOMAIN:Ig-like C2-type 1 | PALLD, NECTIN3, NECTIN1 | 0.656496 | |
| DOMAIN:Ig-like C2-type 2 | PALLD, NECTIN3, NECTIN1 | 0.656496 | |
| Immunoglobulin domain | PALLD, NECTIN3, NECTIN1 | 0.877976 | |
| Immunoglobulin-like fold | PALLD, NECTIN3, NECTIN1 | 0.984375 | |
| Cluster 6 | |||
| Z disc | PALLD, SYNPO2, HSPB1 | 0.481232 | |
| actin cytoskeleton | CALD1, PALLD, SYNPO2 | 0.564483 | |
| actin binding | CALD1, PALLD, SYNPO2 | 0.614614 | |
| focal adhesion | PALLD, SYNPO2, HSPB1 | 0.679141 | |
| Cluster 7 | |||
| Signaling pathways regulating pluripotency of stem cells | FZD3, ZIC3, FZD6 | 0.180791 | |
| Developmental protein | FZD3, ZIC3, FZD6, PITX3 | 0.819281 | |
| Cluster 8 | |||
| Zinc | ZMAT1, ZIC3, ASXL3 | 1 | |
| Zinc-finger | ZMAT1, ZIC3, ASXL3 | 0.988184 | |
| Metal-binding | ZMAT1, ZIC3, ASXL3 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stein, W.D. Orthologs at the Base of the Olfactores Clade. Genes 2024, 15, 657. https://doi.org/10.3390/genes15060657
Stein WD. Orthologs at the Base of the Olfactores Clade. Genes. 2024; 15(6):657. https://doi.org/10.3390/genes15060657
Chicago/Turabian StyleStein, Wilfred D. 2024. "Orthologs at the Base of the Olfactores Clade" Genes 15, no. 6: 657. https://doi.org/10.3390/genes15060657
APA StyleStein, W. D. (2024). Orthologs at the Base of the Olfactores Clade. Genes, 15(6), 657. https://doi.org/10.3390/genes15060657