
Beyond the Anticodon: tRNA Core Modifications and Their Impact on Structure, Translation and Stress Adaptation

Abstract
1. Introduction

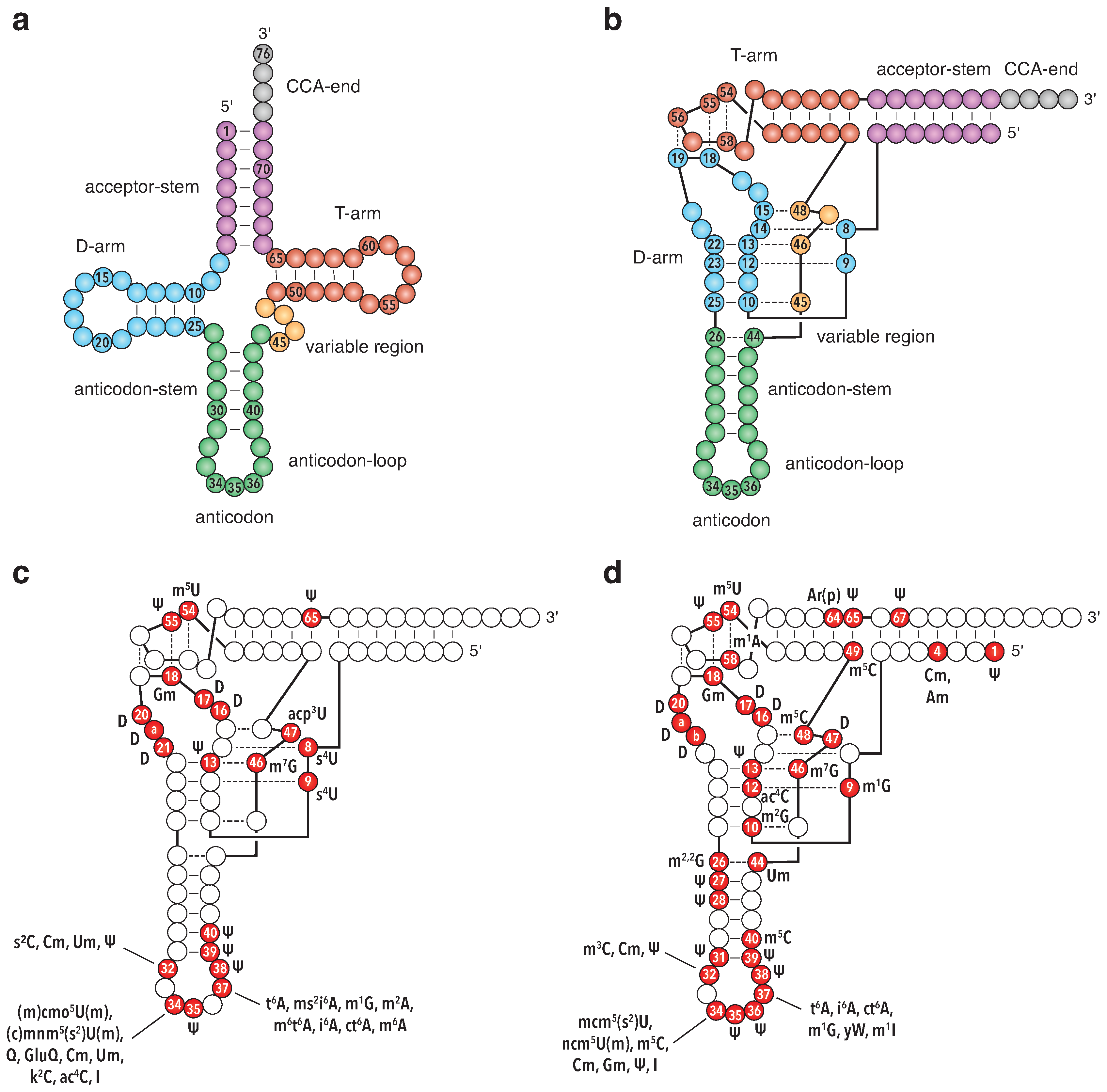
2. Influence of tRNA Core Modification on tRNA Structure and Stability
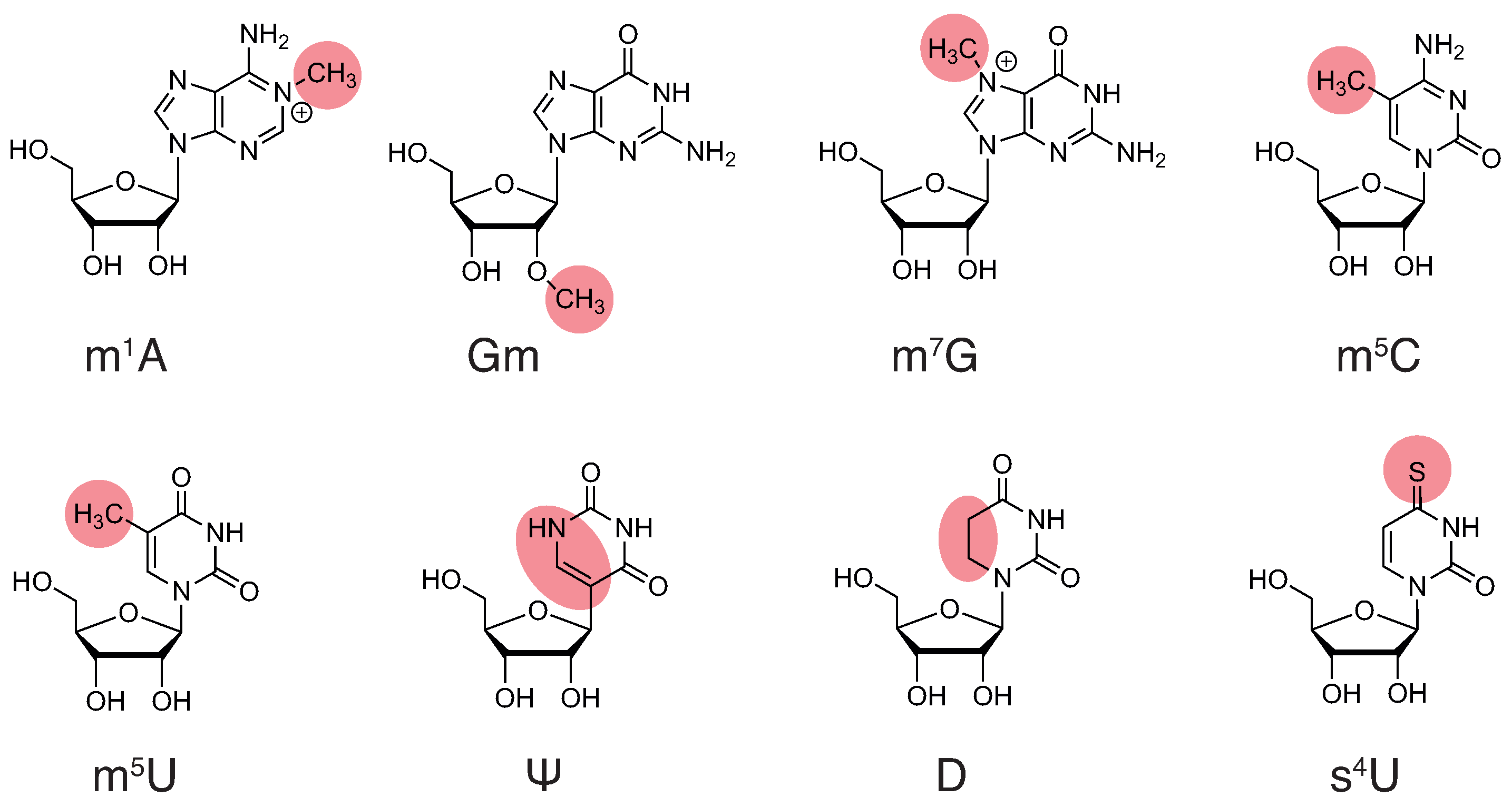
2.1. Modifications and Their Effect on the Physicochemical Properties of the Nucleotides
2.2. Modifications in the tRNA Core and Their Impact on tRNA Structure and Dynamics
2.3. Influence of tRNA Core Modification Enzymes on tRNA Structure: More Than Just Their Catalytic Activity
2.4. tRNA Core Modifications and Their Impact on tRNA Degradation and Cellular Stability

{kind=link}
{kind=link}
{kind=link}
| Organism | tRNA | Lacking Modifications | Conditions | Degradation Pathway | References |
|---|---|---|---|---|---|
| S. cerevisiae | pre- | A58 | 36 °C | Nuclear surveillance | [19,90] |
| S. cerevisiae | A58 | 27 °C, 34 °C | RTD | [91] | |
| S. pombe | A58 | 30 °C, 38.5 °C | RTD | [91] | |
| S. cerevisiae | G9 | 30 °C, late log phase, and w/ or w/o 5FU | Met22-dependent but Xrn1, Rat1, Dxo1, Trf4 and Rrp6-independent degradation | [93] | |
| S. cerevisiae | G46 and (C or 13 or D47) * | 37 °C | RTD | [18,96] | |
| S. cerevisiae | Um44 and C12 * | 36.5 °C | RTD | [96,97] | |
| S. cerevisiae | G26 and C * | 37 °C | RTD | [98] | |
| S. cerevisiae | G46 | 37 °C after thiolutin treatment | RTD | [98] | |
| S. cerevisiae | C12 | 37 °C after thiolutin treatment | RTD | [98] | |
| S. cerevisiae | G26 | 37 °C | RTD | [98] | |
| S. pombe | and | G46 | 36.5 °C, 37.5 °C, 38.5 °C | RTD | [99] |
| T. thermophilus | and | G46 | 70–80 °C | - | [95] |
| V. cholerae | , , , and others | U8 | stationary phase at 37 °C | RNA degradosome | [20] |
| V. cholerae | U8 | log phase at 37 °C | RNA degradosome | [20] | |
| V. cholerae | U8 and (55 or U54) * | log phase at 37 °C and w/o arabinose | RNA degradosome | [20] |
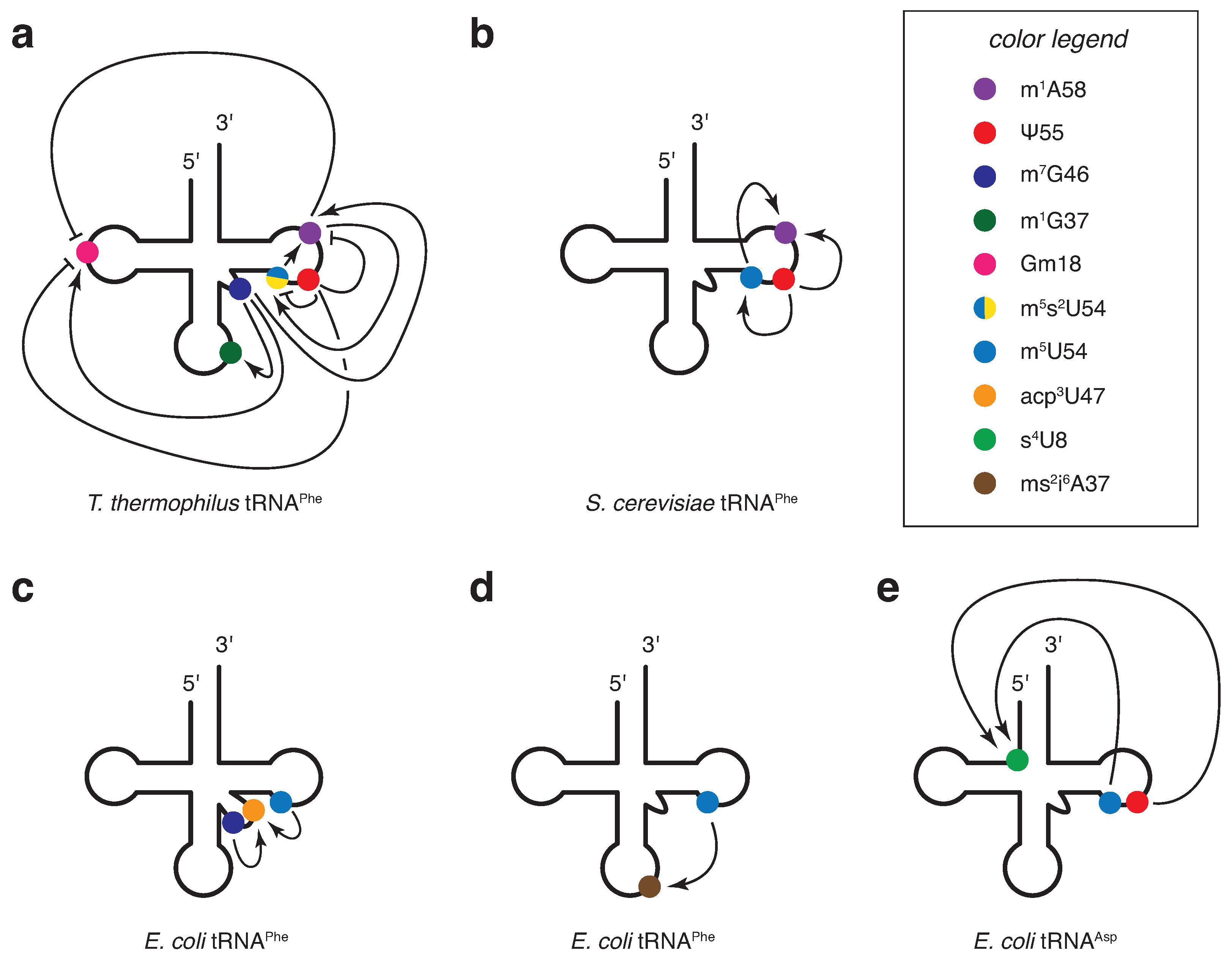
2.5. Interdependence Involving tRNA Core Modifications

3. tRNA Core Modifications and Their Impact on Translation and Stress Adaptation
3.1. tRNA Modification Levels Depend on Growth Conditions
3.2. tRNA Core Modifications and Their Impact on Translation and Fitness
3.3. tRNA Core Modifications and Their Relation with Stress Adaptation Mechanisms
3.3.1. s4U8 Is Implicated in the Response to UV Stress
3.3.2. Gm18 Regulates the Immune-Stimulatory Effect of tRNAs
3.3.3. m7G46 Is Implicated in the Response to Oxidative Stress
| Organism | Modification | Type of Stress | References |
|---|---|---|---|
| S. cerevisiae | Am4 | MMS, NaOCl | [121] |
| E. coli, S. typhimurium | U8 | UV | [147,149,150] |
| E. coli | U8/U9 | heat | [125] |
| S. cerevisiae | G10 | , NaOCl, NaAs | [121] |
| T. thermophilus, E. coli | Gm18 | cold/heat | [95,107,125,163] |
| E. coli, V. cholerae | Gm18 | antibiotics | [123,155,164] |
| S. cerevisiae | Gm18 | NaOCl | [121] |
| S. cerevisiae | G26 | , MMS, NaAs | [121,122] |
| S. cerevisiae | Um44 | , NaAs, NaOCl | [121,165] |
| C. lagenarium, P. aeruginosa, A. baumannii, E. coli | G46 | [158,160,161,162] | |
| T. thermophilus, E. coli | G46 | heat | [95,125] |
| V. cholerae | G46 | antibiotics | [164] |
| S. cerevisiae | G46 | MMS, NaAs | [121,165] |
| E. coli | U47 | heat | [125] |
| S. cerevisiae | C48/C50 | amino acid, glucose and uracil starvation | [124] |
| T. thermophilus | U54 | cold | [107,108] |
| P. furiosus, T. thermophilus | U54 | heat | [95,104,107,166] |
| T. thermophilus | 55 | cold/heat | [107] |
| V. cholerae | 55 | antibiotics | [164] |
| T. thermophilus | A58 | heat | [95,105,107] |
| S. cerevisiae | A58 | [121] | |
| V. cholerae, E. coli | D | antibiotics | [164] |
| S. cerevisiae | C | , MMS, NaOCl, NaAs | [121] |
| S. cerevisiae | Cm | , MMS, NaAs | [121] |
3.3.4. tRNA Core Modifications in the Response to Antibiotic Stress
4. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Phizicky, E.M.; Hopper, A.K. The life and times of a tRNA. RNA 2023, 29, 898–957. [Google Scholar] [CrossRef]
- Berg, M.D.; Brandl, C.J. Transfer RNAs: Diversity in form and function. RNA Biol. 2021, 18, 316–339. [Google Scholar] [CrossRef]
- Barraud, P.; Tisné, C. To be or not to be modified: Miscellaneous aspects influencing nucleotide modifications in tRNAs. IUBMB Life 2019, 71, 1126–1140. [Google Scholar] [CrossRef]
- Hopper, A.K. Transfer RNA post-transcriptional processing, turnover, and subcellular dynamics in the yeast Saccharomyces cerevisiae. Genetics 2013, 194, 43–67. [Google Scholar] [CrossRef]
- Lin, B.Y.; Chan, P.P.; Lowe, T.M. tRNAviz: Explore and visualize tRNA sequence features. Nucleic Acids Res. 2019, 47, W542–W547. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. GtRNAdb 2.0: An expanded database of transfer RNA genes identified in complete and draft genomes. Nucleic Acids Res. 2016, 44, D184–D189. [Google Scholar] [CrossRef] [PubMed]
- Robertus, J.D.; Ladner, J.E.; Finch, J.T.; Rhodes, D.; Brown, R.S.; Clark, B.F.; Klug, A. Structure of yeast phenylalanine tRNA at 3 A resolution. Nature 1974, 250, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Suddath, F.L.; Quigley, G.J.; McPherson, A.; Sussman, J.L.; Wang, A.H.; Seeman, N.C.; Rich, A. Three-dimensional tertiary structure of yeast phenylalanine transfer RNA. Science 1974, 185, 435–440. [Google Scholar] [CrossRef]
- Giegé, R.; Jühling, F.; Pütz, J.; Stadler, P.; Sauter, C.; Florentz, C. Structure of transfer RNAs: Similarity and variability. Wiley Interdiscip. Rev. RNA 2012, 3, 37–61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ferré-D’Amaré, A.R. The tRNA Elbow in Structure, Recognition and Evolution. Life 2016, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Biela, A.; Hammermeister, A.; Kaczmarczyk, I.; Walczak, M.; Koziej, L.; Lin, T.Y.; Glatt, S. The diverse structural modes of tRNA binding and recognition. J. Biol. Chem. 2023, 299, 104966. [Google Scholar] [CrossRef] [PubMed]
- Jackman, J.E.; Alfonzo, J.D. Transfer RNA modifications: Nature’s combinatorial chemistry playground. Wiley Interdiscip. Rev. RNA 2013, 4, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Ontiveros, R.J.; Stoute, J.; Liu, K.F. The chemical diversity of RNA modifications. Biochem. J. 2019, 476, 1227–1245. [Google Scholar] [CrossRef] [PubMed]
- Cappannini, A.; Ray, A.; Purta, E.; Mukherjee, S.; Boccaletto, P.; Moafinejad, S.N.; Lechner, A.; Barchet, C.; Klaholz, B.P.; Stefaniak, F.; et al. MODOMICS: A database of RNA modifications and related information. 2023 update. Nucleic Acids Res. 2024, 52, D239–D244. [Google Scholar] [CrossRef] [PubMed]
- Machnicka, M.A.; Olchowik, A.; Grosjean, H.; Bujnicki, J.M. Distribution and frequencies of post-transcriptional modifications in tRNAs. RNA Biol. 2014, 11, 1619–1629. [Google Scholar] [CrossRef] [PubMed]
- Motorin, Y.; Helm, M. tRNA stabilization by modified nucleotides. Biochemistry 2010, 49, 4934–4944. [Google Scholar] [CrossRef]
- Lorenz, C.; Lünse, C.E.; Mörl, M. tRNA Modifications: Impact on Structure and Thermal Adaptation. Biomolecules 2017, 7, 35. [Google Scholar] [CrossRef]
- Alexandrov, A.; Chernyakov, I.; Gu, W.; Hiley, S.L.; Hughes, T.R.; Grayhack, E.J.; Phizicky, E.M. Rapid tRNA decay can result from lack of nonessential modifications. Mol. Cell 2006, 21, 87–96. [Google Scholar] [CrossRef]
- Kadaba, S.; Krueger, A.; Trice, T.; Krecic, A.M.; Hinnebusch, A.G.; Anderson, J. Nuclear surveillance and degradation of hypomodified initiator tRNAMet in S. cerevisiae. Genes Dev. 2004, 18, 1227–1240. [Google Scholar] [CrossRef]
- Kimura, S.; Waldor, M.K. The RNA degradosome promotes tRNA quality control through clearance of hypomodified tRNA. Proc. Natl. Acad. Sci. USA 2019, 116, 1394–1403. [Google Scholar] [CrossRef]
- Wichtowska, D.; Turowski, T.W.; Boguta, M. An interplay between transcription, processing, and degradation determines tRNA levels in yeast. Wiley Interdiscip. Rev. RNA 2013, 4, 709–722. [Google Scholar] [CrossRef]
- Sprinzl, M.; Horn, C.; Brown, M.; Ioudovitch, A.; Steinberg, S. Compilation of tRNA sequences and sequences of tRNA genes. Nucleic Acids Res. 1998, 26, 148–153. [Google Scholar] [CrossRef]
- Björk, G.R.; Hagervall, T.G. Transfer RNA Modification: Presence, Synthesis, and Function. EcoSal Plus 2014, 6, 10-1128. [Google Scholar] [CrossRef]
- de Crécy-Lagard, V.; Jaroch, M. Functions of Bacterial tRNA Modifications: From Ubiquity to Diversity. Trends Microbiol. 2021, 29, 41–53. [Google Scholar] [CrossRef]
- Smith, T.J.; Giles, R.N.; Koutmou, K.S. Anticodon stem-loop tRNA modifications influence codon decoding and frame maintenance during translation. Semin. Cell Dev. Biol. 2024, 154, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. The expanding world of tRNA modifications and their disease relevance. Nat. Rev. Mol. Cell Biol. 2021, 22, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Agris, P.F.; Narendran, A.; Sarachan, K.; Väre, V.Y.P.; Eruysal, E. The Importance of Being Modified: The Role of RNA Modifications in Translational Fidelity. Enzymes 2017, 41, 1–50. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G.; Batlle, E.; Ribas de Pouplana, L. Role of tRNA modifications in human diseases. Trends Mol. Med. 2014, 20, 306–314. [Google Scholar] [CrossRef] [PubMed]
- El Yacoubi, B.; Bailly, M.; de Crécy-Lagard, V. Biosynthesis and function of posttranscriptional modifications of transfer RNAs. Annu. Rev. Genet. 2012, 46, 69–95. [Google Scholar] [CrossRef] [PubMed]
- Nedialkova, D.D.; Leidel, S.A. Optimization of Codon Translation Rates via tRNA Modifications Maintains Proteome Integrity. Cell 2015, 161, 1606–1618. [Google Scholar] [CrossRef]
- Urbonavicius, J.; Qian, Q.; Durand, J.M.; Hagervall, T.G.; Björk, G.R. Improvement of reading frame maintenance is a common function for several tRNA modifications. EMBO J. 2001, 20, 4863–4873. [Google Scholar] [CrossRef] [PubMed]
- Urbonavicius, J.; Stahl, G.; Durand, J.M.B.; Ben Salem, S.N.; Qian, Q.; Farabaugh, P.J.; Björk, G.R. Transfer RNA modifications that alter +1 frameshifting in general fail to affect -1 frameshifting. RNA 2003, 9, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, N.; Rodnina, M.V. Thio-Modification of tRNA at the Wobble Position as Regulator of the Kinetics of Decoding and Translocation on the Ribosome. J. Am. Chem. Soc. 2017, 139, 5857–5864. [Google Scholar] [CrossRef] [PubMed]
- Tükenmez, H.; Xu, H.; Esberg, A.; Byström, A.S. The role of wobble uridine modifications in +1 translational frameshifting in eukaryotes. Nucleic Acids Res. 2015, 43, 9489–9499. [Google Scholar] [CrossRef]
- Klassen, R.; Bruch, A.; Schaffrath, R. Independent suppression of ribosomal +1 frameshifts by different tRNA anticodon loop modifications. RNA Biol. 2017, 14, 1252–1259. [Google Scholar] [CrossRef]
- Manickam, N.; Joshi, K.; Bhatt, M.J.; Farabaugh, P.J. Effects of tRNA modification on translational accuracy depend on intrinsic codon-anticodon strength. Nucleic Acids Res. 2016, 44, 1871–1881. [Google Scholar] [CrossRef]
- Su, C.; Jin, M.; Zhang, W. Conservation and Diversification of tRNA t6A-Modifying Enzymes across the Three Domains of Life. Int. J. Mol. Sci. 2022, 23, 13600. [Google Scholar] [CrossRef]
- Endres, L.; Dedon, P.C.; Begley, T.J. Codon-biased translation can be regulated by wobble-base tRNA modification systems during cellular stress responses. RNA Biol. 2015, 12, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.; Pham, P.; Dedon, P.C.; Begley, T.J. Lifestyle modifications: Coordinating the tRNA epitranscriptome with codon bias to adapt translation during stress responses. Genome Biol. 2018, 19, 228. [Google Scholar] [CrossRef]
- Pollo-Oliveira, L.; de Crécy-Lagard, V. Can Protein Expression Be Regulated by Modulation of tRNA Modification Profiles? Biochemistry 2019, 58, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Mitchener, M.M.; Begley, T.J.; Dedon, P.C. Molecular Coping Mechanisms: Reprogramming tRNAs To Regulate Codon-Biased Translation of Stress Response Proteins. Acc. Chem. Res. 2023, 56, 3504–3514. [Google Scholar] [CrossRef]
- Pan, T. Modifications and functional genomics of human transfer RNA. Cell Res. 2018, 28, 395–404. [Google Scholar] [CrossRef]
- Chujo, T.; Tomizawa, K. Human transfer RNA modopathies: Diseases caused by aberrations in transfer RNA modifications. FEBS J. 2021, 288, 7096–7122. [Google Scholar] [CrossRef]
- Zhou, J.B.; Wang, E.D.; Zhou, X.L. Modifications of the human tRNA anticodon loop and their associations with genetic diseases. Cell Mol. Life Sci. 2021, 78, 7087–7105. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.W.; Eargle, J.; Luthey-Schulten, Z. Experimental and computational determination of tRNA dynamics. FEBS Lett. 2010, 584, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Väre, V.Y.P.; Eruysal, E.R.; Narendran, A.; Sarachan, K.L.; Agris, P.F. Chemical and Conformational Diversity of Modified Nucleosides Affects tRNA Structure and Function. Biomolecules 2017, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Roovers, M.; Droogmans, L.; Grosjean, H. Post-Transcriptional Modifications of Conserved Nucleotides in the T-Loop of tRNA: A Tale of Functional Convergent Evolution. Genes 2021, 12, 140. [Google Scholar] [CrossRef] [PubMed]
- Hori, H. Transfer RNA Modification Enzymes with a Thiouridine Synthetase, Methyltransferase and Pseudouridine Synthase (THUMP) Domain and the Nucleosides They Produce in tRNA. Genes 2023, 14, 382. [Google Scholar] [CrossRef] [PubMed]
- Kawai, G.; Yamamoto, Y.; Kamimura, T.; Masegi, T.; Sekine, M.; Hata, T.; Iimori, T.; Watanabe, T.; Miyazawa, T.; Yokoyama, S. Conformational rigidity of specific pyrimidine residues in tRNA arises from posttranscriptional modifications that enhance steric interaction between the base and the 2’-hydroxyl group. Biochemistry 1992, 31, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Yokoyama, S.; Miyazawa, T.; Watanabe, K.; Higuchi, S. NMR analyses on the molecular mechanism of the conformational rigidity of 2-thioribothymidine, a modified nucleoside in extreme thermophile tRNAs. FEBS Lett. 1983, 157, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.R. Stabilization of RNA stacking by pseudouridine. Nucleic Acids Res. 1995, 23, 5020–5026. [Google Scholar] [CrossRef]
- Zhang, X.; Walker, R.C.; Phizicky, E.M.; Mathews, D.H. Influence of Sequence and Covalent Modifications on Yeast tRNA Dynamics. J. Chem. Theory Comput. 2014, 10, 3473–3483. [Google Scholar] [CrossRef]
- Nomura, Y.; Ohno, S.; Nishikawa, K.; Yokogawa, T. Correlation between the stability of tRNA tertiary structure and the catalytic efficiency of a tRNA-modifying enzyme, archaeal tRNA-guanine transglycosylase. Genes Cells 2016, 21, 41–52. [Google Scholar] [CrossRef]
- Davanloo, P.; Sprinzl, M.; Watanabe, K.; Albani, M.; Kersten, H. Role of ribothymidine in the thermal stability of transfer RNA as monitored by proton magnetic resonance. Nucleic Acids Res. 1979, 6, 1571–1581. [Google Scholar] [CrossRef]
- Horie, N.; Hara-Yokoyama, M.; Yokoyama, S.; Watanabe, K.; Kuchino, Y.; Nishimura, S.; Miyazawa, T. Two tRNAIle1 species from an extreme thermophile, Thermus thermophilus HB8: Effect of 2-thiolation of ribothymidine on the thermostability of tRNA. Biochemistry 1985, 24, 5711–5715. [Google Scholar] [CrossRef]
- Griffey, R.H.; Davis, D.R.; Yamaizumi, Z.; Nishimura, S.; Hawkins, B.L.; Poulter, C.D. 15N-labeled tRNA. Identification of 4-thiouridine in Escherichia coli tRNASer1 and tRNATyr2 by 1H-15N two-dimensional NMR spectroscopy. J. Biol. Chem. 1986, 261, 12074–12078. [Google Scholar] [CrossRef]
- Urbonavicius, J.; Durand, J.M.B.; Björk, G.R. Three modifications in the D and T arms of tRNA influence translation in Escherichia coli and expression of virulence genes in Shigella flexneri. J. Bacteriol. 2002, 184, 5348–5357. [Google Scholar] [CrossRef] [PubMed]
- Arnez, J.G.; Steitz, T.A. Crystal structure of unmodified tRNA(Gln) complexed with glutaminyl-tRNA synthetase and ATP suggests a possible role for pseudo-uridines in stabilization of RNA structure. Biochemistry 1994, 33, 7560–7567. [Google Scholar] [CrossRef] [PubMed]
- Westhof, E. Pseudouridines or how to draw on weak energy differences. Biochem. Biophys. Res. Commun. 2019, 520, 702–704. [Google Scholar] [CrossRef] [PubMed]
- Nobles, K.N.; Yarian, C.S.; Liu, G.; Guenther, R.H.; Agris, P.F. Highly conserved modified nucleosides influence Mg2+-dependent tRNA folding. Nucleic Acids Res. 2002, 30, 4751–4760. [Google Scholar] [CrossRef] [PubMed]
- Sowers, L.C.; Shaw, B.R.; Sedwick, W.D. Base stacking and molecular polarizability: Effect of a methyl group in the 5-position of pyrimidines. Biochem. Biophys. Res. Commun. 1987, 148, 790–794. [Google Scholar] [CrossRef]
- Kintanar, A.; Yue, D.; Horowitz, J. Effect of nucleoside modifications on the structure and thermal stability of Escherichia coli valine tRNA. Biochimie 1994, 76, 1192–1204. [Google Scholar] [CrossRef] [PubMed]
- Agris, P.F.; Sierzputowska-Gracz, H.; Smith, C. Transfer RNA contains sites of localized positive charge: Carbon NMR studies of [13C]methyl-enriched Escherichia coli and yeast tRNAPhe. Biochemistry 1986, 25, 5126–5131. [Google Scholar] [CrossRef]
- Steinberg, S.; Cedergren, R. A correlation between N2-dimethylguanosine presence and alternate tRNA conformers. RNA 1995, 1, 886–891. [Google Scholar]
- Helm, M.; Brulé, H.; Degoul, F.; Cepanec, C.; Leroux, J.P.; Giegé, R.; Florentz, C. The presence of modified nucleotides is required for cloverleaf folding of a human mitochondrial tRNA. Nucleic Acids Res. 1998, 26, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Helm, M.; Giegé, R.; Florentz, C. A Watson-Crick base-pair-disrupting methyl group (m1A9) is sufficient for cloverleaf folding of human mitochondrial tRNALys. Biochemistry 1999, 38, 13338–13346. [Google Scholar] [CrossRef]
- Finet, O.; Yague-Sanz, C.; Marchand, F.; Hermand, D. The Dihydrouridine landscape from tRNA to mRNA: A perspective on synthesis, structural impact and function. RNA Biol. 2022, 19, 735–750. [Google Scholar] [CrossRef]
- Catala, M.; Gato, A.; Tisné, C.; Barraud, P. 1H, 15N chemical shift assignments of the imino groups of yeast tRNAPhe: Influence of the post-transcriptional modifications. Biomol. NMR Assign. 2020, 14, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Barraud, P.; Gato, A.; Heiss, M.; Catala, M.; Kellner, S.; Tisné, C. Time-resolved NMR monitoring of tRNA maturation. Nat. Commun. 2019, 10, 3373. [Google Scholar] [CrossRef]
- Yared, M.J.; Yoluç, Y.; Catala, M.; Tisné, C.; Kaiser, S.; Barraud, P. Different modification pathways for m1A58 incorporation in yeast elongator and initiator tRNAs. Nucleic Acids Res. 2023, 51, 10653–10667. [Google Scholar] [CrossRef]
- Basavappa, R.; Sigler, P.B. The 3 A crystal structure of yeast initiator tRNA: Functional implications in initiator/elongator discrimination. EMBO J. 1991, 10, 3105–3111. [Google Scholar] [CrossRef] [PubMed]
- Phizicky, E.M.; Alfonzo, J.D. Do all modifications benefit all tRNAs? FEBS Lett. 2010, 584, 265–271. [Google Scholar] [CrossRef]
- Dalluge, J.J.; Hashizume, T.; Sopchik, A.E.; McCloskey, J.A.; Davis, D.R. Conformational flexibility in RNA: The role of dihydrouridine. Nucleic Acids Res. 1996, 24, 1073–1079. [Google Scholar] [CrossRef]
- Dyubankova, N.; Sochacka, E.; Kraszewska, K.; Nawrot, B.; Herdewijn, P.; Lescrinier, E. Contribution of dihydrouridine in folding of the D-arm in tRNA. Org. Biomol. Chem. 2015, 13, 4960–4966. [Google Scholar] [CrossRef] [PubMed]
- Biedenbänder, T.; de Jesus, V.; Schmidt-Dengler, M.; Helm, M.; Corzilius, B.; Fürtig, B. RNA modifications stabilize the tertiary structure of tRNAfMet by locally increasing conformational dynamics. Nucleic Acids Res. 2022, 50, 2334–2349. [Google Scholar] [CrossRef] [PubMed]
- Takakura, M.; Ishiguro, K.; Akichika, S.; Miyauchi, K.; Suzuki, T. Biogenesis and functions of aminocarboxypropyluridine in tRNA. Nat. Commun. 2019, 10, 5542. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, A.; McCallum, S.A.; Pardi, A. Comparison of the global structure and dynamics of native and unmodified tRNAval. Biochemistry 2005, 44, 6024–6033. [Google Scholar] [CrossRef]
- Godwin, R.C.; Macnamara, L.M.; Alexander, R.W.; Salsbury, F.R., Jr. Structure and Dynamics of tRNAMet Containing Core Substitutions. ACS Omega 2018, 3, 10668–10678. [Google Scholar] [CrossRef]
- Bhaskaran, H.; Taniguchi, T.; Suzuki, T.; Suzuki, T.; Perona, J.J. Structural dynamics of a mitochondrial tRNA possessing weak thermodynamic stability. Biochemistry 2014, 53, 1456–1465. [Google Scholar] [CrossRef]
- Porat, J.; Kothe, U.; Bayfield, M.A. Revisiting tRNA chaperones: New players in an ancient game. RNA 2021, 27, 543–559. [Google Scholar] [CrossRef]
- Keffer-Wilkes, L.C.; Veerareddygari, G.R.; Kothe, U. RNA modification enzyme TruB is a tRNA chaperone. Proc. Natl. Acad. Sci. USA 2016, 113, 14306–14311. [Google Scholar] [CrossRef] [PubMed]
- Keffer-Wilkes, L.C.; Soon, E.F.; Kothe, U. The methyltransferase TrmA facilitates tRNA folding through interaction with its RNA-binding domain. Nucleic Acids Res. 2020, 48, 7981–7990. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, R.; Nureki, O.; Nameki, N.; Okada, N.; Nishimura, S.; Yokoyama, S. Alternative tertiary structure of tRNA for recognition by a posttranscriptional modification enzyme. Cell 2003, 113, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Finer-Moore, J.; Czudnochowski, N.; O’Connell, J.D., 3rd; Wang, A.L.; Stroud, R.M. Crystal Structure of the Human tRNA m(1)A58 Methyltransferase-tRNA(3)(Lys) Complex: Refolding of Substrate tRNA Allows Access to the Methylation Target. J. Mol. Biol. 2015, 427, 3862–3876. [Google Scholar] [CrossRef] [PubMed]
- Porat, J.; Vakiloroayaei, A.; Remnant, B.M.; Talebi, M.; Cargill, T.; Bayfield, M.A. Crosstalk between the tRNA methyltransferase Trm1 and RNA chaperone La influences eukaryotic tRNA maturation. J. Biol. Chem. 2023, 299, 105326. [Google Scholar] [CrossRef]
- Rajkowitsch, L.; Schroeder, R. Dissecting RNA chaperone activity. RNA 2007, 13, 2053–2060. [Google Scholar] [CrossRef]
- Chakshusmathi, G.; Kim, S.D.; Rubinson, D.A.; Wolin, S.L. A La protein requirement for efficient pre-tRNA folding. EMBO J. 2003, 22, 6562–6572. [Google Scholar] [CrossRef]
- Copela, L.A.; Chakshusmathi, G.; Sherrer, R.L.; Wolin, S.L. The La protein functions redundantly with tRNA modification enzymes to ensure tRNA structural stability. RNA 2006, 12, 644–654. [Google Scholar] [CrossRef]
- Vakiloroayaei, A.; Shah, N.S.; Oeffinger, M.; Bayfield, M.A. The RNA chaperone La promotes pre-tRNA maturation via indiscriminate binding of both native and misfolded targets. Nucleic Acids Res. 2017, 45, 11341–11355. [Google Scholar] [CrossRef]
- Kadaba, S.; Wang, X.; Anderson, J.T. Nuclear RNA surveillance in Saccharomyces cerevisiae: Trf4p-dependent polyadenylation of nascent hypomethylated tRNA and an aberrant form of 5S rRNA. RNA 2006, 12, 508–521. [Google Scholar] [CrossRef]
- Tasak, M.; Phizicky, E.M. Initiator tRNA lacking 1-methyladenosine is targeted by the rapid tRNA decay pathway in evolutionarily distant yeast species. PLoS Genet 2022, 18, e1010215. [Google Scholar] [CrossRef]
- Whipple, J.M.; Lane, E.A.; Chernyakov, I.; D’Silva, S.; Phizicky, E.M. The yeast rapid tRNA decay pathway primarily monitors the structural integrity of the acceptor and T-stems of mature tRNA. Genes Dev. 2011, 25, 1173–1184. [Google Scholar] [CrossRef]
- Bowles, I.E.; Jackman, J.E. A tRNA-specific function for tRNA methyltransferase Trm10 is associated with a new tRNA quality control mechanism in Saccharomyces cerevisiae. RNA 2024, 30, 171–187. [Google Scholar] [CrossRef]
- Hori, H. Regulatory Factors for tRNA Modifications in Extreme-Thermophilic Bacterium Thermus thermophilus. Front. Genet. 2019, 10, 204. [Google Scholar] [CrossRef]
- Tomikawa, C.; Yokogawa, T.; Kanai, T.; Hori, H. N7-Methylguanine at position 46 (m7G46) in tRNA from Thermus thermophilus is required for cell viability at high temperatures through a tRNA modification network. Nucleic Acids Res. 2010, 38, 942–957. [Google Scholar] [CrossRef] [PubMed]
- Chernyakov, I.; Whipple, J.M.; Kotelawala, L.; Grayhack, E.J.; Phizicky, E.M. Degradation of several hypomodified mature tRNA species in Saccharomyces cerevisiae is mediated by Met22 and the 5’-3’exonucleases Rat1 and Xrn1. Genes Dev. 2008, 22, 1369–1380. [Google Scholar] [CrossRef]
- Kotelawala, L.; Grayhack, E.J.; Phizicky, E.M. Identification of yeast tRNA Um(44) 2’-O-methyltransferase (Trm44) and demonstration of a Trm44 role in sustaining levels of specific tRNA(Ser) species. RNA 2008, 14, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Dewe, J.M.; Whipple, J.M.; Chernyakov, I.; Jaramillo, L.N.; Phizicky, E.M. The yeast rapid tRNA decay pathway competes with elongation factor 1A for substrate tRNAs and acts on tRNAs lacking one or more of several modifications. RNA 2012, 18, 1886–1896. [Google Scholar] [CrossRef]
- De Zoysa, T.; Phizicky, E.M. Hypomodified tRNA in evolutionarily distant yeasts can trigger rapid tRNA decay to activate the general amino acid control response, but with different consequences. PLoS Genet. 2020, 16, e1008893. [Google Scholar] [CrossRef]
- Han, L.; Phizicky, E.M. A rationale for tRNA modification circuits in the anticodon loop. RNA 2018, 24, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Sokołowski, M.; Klassen, R.; Bruch, A.; Schaffrath, R.; Glatt, S. Cooperativity between different tRNA modifications and their modification pathways. Biochim. Biophys. Acta Gene Regul. Mech. 2018, 1861, 409–418. [Google Scholar] [CrossRef]
- Li, J.; Zhu, W.Y.; Yang, W.Q.; Li, C.T.; Liu, R.J. The occurrence order and cross-talk of different tRNA modifications. Sci. China Life Sci. 2021, 64, 1423–1436. [Google Scholar] [CrossRef] [PubMed]
- Tomikawa, C. 7-Methylguanosine Modifications in Transfer RNA (tRNA). Int. J. Mol. Sci. 2018, 19, 4080. [Google Scholar] [CrossRef] [PubMed]
- Kowalak, J.A.; Dalluge, J.J.; McCloskey, J.A.; Stetter, K.O. The role of posttranscriptional modification in stabilization of transfer RNA from hyperthermophiles. Biochemistry 1994, 33, 7869–7876. [Google Scholar] [CrossRef] [PubMed]
- Droogmans, L.; Roovers, M.; Bujnicki, J.M.; Tricot, C.; Hartsch, T.; Stalon, V.; Grosjean, H. Cloning and characterization of tRNA (m1A58) methyltransferase (TrmI) from Thermus thermophilus HB27, a protein required for cell growth at extreme temperatures. Nucleic Acids Res. 2003, 31, 2148–2156. [Google Scholar] [CrossRef] [PubMed]
- Ohira, T.; Suzuki, T. Transfer RNA modifications and cellular thermotolerance. Mol. Cell 2024, 84, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Kunibayashi, T.; Tomikawa, C.; Ochi, A.; Kanai, T.; Hirata, A.; Iwashita, C.; Hori, H. Pseudouridine at position 55 in tRNA controls the contents of other modified nucleotides for low-temperature adaptation in the extreme-thermophilic eubacterium Thermus thermophilus. Nucleic Acids Res. 2011, 39, 2304–2318. [Google Scholar] [CrossRef]
- Yamagami, R.; Tomikawa, C.; Shigi, N.; Kazayama, A.; Asai, S.I.; Takuma, H.; Hirata, A.; Fourmy, D.; Asahara, H.; Watanabe, K.; et al. Folate-/FAD-dependent tRNA methyltransferase from Thermus thermophilus regulates other modifications in tRNA at low temperatures. Genes Cells 2016, 21, 740–754. [Google Scholar] [CrossRef]
- Meyer, B.; Immer, C.; Kaiser, S.; Sharma, S.; Yang, J.; Watzinger, P.; Weiß, L.; Kotter, A.; Helm, M.; Seitz, H.M.; et al. Identification of the 3-amino-3-carboxypropyl (acp) transferase enzyme responsible for acp3U formation at position 47 in Escherichia coli tRNAs. Nucleic Acids Res. 2020, 48, 1435–1450. [Google Scholar] [CrossRef]
- Jones, J.D.; Franco, M.K.; Tardu, M.; Smith, T.J.; Snyder, L.R.; Eyler, D.E.; Polikanov, Y.; Kennedy, R.T.; Niederer, R.O.; Koutmou, K.S. Conserved 5-methyluridine tRNA modification modulates ribosome translocation. bioRxiv 2023. [Google Scholar] [CrossRef]
- Schultz, S.K.; Katanski, C.D.; Halucha, M.; Pena, N.; Fahlman, R.P.; Pan, T.; Kothe, U. Modifications in the T arm of tRNA globally determine tRNA maturation, function and cellular fitness. bioRxiv 2023. [Google Scholar] [CrossRef]
- Lucas, M.C.; Pryszcz, L.P.; Medina, R.; Milenkovic, I.; Camacho, N.; Marchand, V.; Motorin, Y.; Ribas de Pouplana, L.; Novoa, E.M. Quantitative analysis of tRNA abundance and modifications by nanopore RNA sequencing. Nat. Biotechnol. 2024, 42, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Smoczynski, J.; Yared, M.J.; Meynier, V.; Barraud, P.; Tisné, C. Advances in the Structural and Functional Understanding of m1A RNA Modification. Acc. Chem. Res. 2024, 57, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Schultz, S.K.L.; Kothe, U. tRNA elbow modifications affect the tRNA pseudouridine synthase TruB and the methyltransferase TrmA. RNA 2020, 26, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Thomas, N.K.; Poodari, V.C.; Jain, M.; Olsen, H.E.; Akeson, M.; Abu-Shumays, R.L. Direct Nanopore Sequencing of Individual Full Length tRNA Strands. ACS Nano 2021, 15, 16642–16653. [Google Scholar] [CrossRef] [PubMed]
- Emilsson, V.; Näslund, A.K.; Kurland, C.G. Thiolation of transfer RNA in Escherichia coli varies with growth rate. Nucleic Acids Res. 1992, 20, 4499–4505. [Google Scholar] [CrossRef] [PubMed]
- Emilsson, V.; Näslund, A.K.; Kurland, C.G. Growth-rate-dependent accumulation of twelve tRNA species in Escherichia coli. J. Mol. Biol. 1993, 230, 483–491. [Google Scholar] [CrossRef]
- Dong, H.; Nilsson, L.; Kurland, C.G. Co-variation of tRNA abundance and codon usage in Escherichia coli at different growth rates. J. Mol. Biol. 1996, 260, 649–663. [Google Scholar] [CrossRef]
- Torrent, M.; Chalancon, G.; de Groot, N.S.; Wuster, A.; Madan Babu, M. Cells alter their tRNA abundance to selectively regulate protein synthesis during stress conditions. Sci. Signal. 2018, 11. [Google Scholar] [CrossRef]
- Singhal, R.P.; Vold, B. Changes in transfer ribonucleic acids of Bacillus subtilis during different growth phases. Nucleic Acids Res. 1976, 3, 1249–1262. [Google Scholar] [CrossRef][Green Version]
- Chan, C.T.Y.; Dyavaiah, M.; DeMott, M.S.; Taghizadeh, K.; Dedon, P.C.; Begley, T.J. A quantitative systems approach reveals dynamic control of tRNA modifications during cellular stress. PLoS Genet. 2010, 6, e1001247. [Google Scholar] [CrossRef]
- Pang, Y.L.J.; Abo, R.; Levine, S.S.; Dedon, P.C. Diverse cell stresses induce unique patterns of tRNA up- and down-regulation: TRNA-seq for quantifying changes in tRNA copy number. Nucleic Acids Res. 2014, 42, e170. [Google Scholar] [CrossRef]
- Galvanin, A.; Vogt, L.M.; Grober, A.; Freund, I.; Ayadi, L.; Bourguignon-Igel, V.; Bessler, L.; Jacob, D.; Eigenbrod, T.; Marchand, V.; et al. Bacterial tRNA 2’-O-methylation is dynamically regulated under stress conditions and modulates innate immune response. Nucleic Acids Res. 2020, 48, 12833–12844. [Google Scholar] [CrossRef] [PubMed]
- Preston, M.A.; D’Silva, S.; Kon, Y.; Phizicky, E.M. tRNAHis 5-methylcytidine levels increase in response to several growth arrest conditions in Saccharomyces cerevisiae. RNA 2013, 19, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, R.; Sieg, J.P.; Assmann, S.M.; Bevilacqua, P.C. Genome-wide analysis of the in vivo tRNA structurome reveals RNA structural and modification dynamics under heat stress. Proc. Natl. Acad. Sci. USA 2022, 119, e2201237119. [Google Scholar] [CrossRef] [PubMed]
- Pape, T.; Wintermeyer, W.; Rodnina, M.V. Complete kinetic mechanism of elongation factor Tu-dependent binding of aminoacyl-tRNA to the A site of the E. coli ribosome. EMBO J. 1998, 17, 7490–7497. [Google Scholar] [CrossRef] [PubMed]
- Whitford, P.C.; Geggier, P.; Altman, R.B.; Blanchard, S.C.; Onuchic, J.N.; Sanbonmatsu, K.Y. Accommodation of aminoacyl-tRNA into the ribosome involves reversible excursions along multiple pathways. RNA 2010, 16, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lancaster, L.; Donohue, J.P.; Noller, H.F. How the ribosome hands the A-site tRNA to the P site during EF-G-catalyzed translocation. Science 2014, 345, 1188–1191. [Google Scholar] [CrossRef] [PubMed]
- Rundlet, E.J.; Holm, M.; Schacherl, M.; Natchiar, S.K.; Altman, R.B.; Spahn, C.M.T.; Myasnikov, A.G.; Blanchard, S.C. Structural basis of early translocation events on the ribosome. Nature 2021, 595, 741–745. [Google Scholar] [CrossRef]
- Milicevic, N.; Jenner, L.; Myasnikov, A.; Yusupov, M.; Yusupova, G. mRNA reading frame maintenance during eukaryotic ribosome translocation. Nature 2024, 625, 393–400. [Google Scholar] [CrossRef]
- Ling, C.; Ermolenko, D.N. Structural insights into ribosome translocation. Wiley Interdiscip. Rev. RNA 2016, 7, 620–636. [Google Scholar] [CrossRef]
- Saleh, S.; Farabaugh, P.J. Posttranscriptional modification to the core of tRNAs modulates translational misreading errors. RNA 2023, 30, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Kinghorn, S.M.; O’Byrne, C.P.; Booth, I.R.; Stansfield, I. Physiological analysis of the role of truB in Escherichia coli: A role for tRNA modification in extreme temperature resistance. Microbiology 2002, 148, 3511–3520. [Google Scholar] [CrossRef] [PubMed]
- Björk, G.R.; Neidhardt, F.C. Physiological and biochemical studies on the function of 5-methyluridine in the transfer ribonucleic acid of Escherichia coli. J. Bacteriol. 1975, 124, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Gutgsell, N.; Englund, N.; Niu, L.; Kaya, Y.; Lane, B.G.; Ofengand, J. Deletion of the Escherichia coli pseudouridine synthase gene truB blocks formation of pseudouridine 55 in tRNA in vivo, does not affect exponential growth, but confers a strong selective disadvantage in competition with wild-type cells. RNA 2000, 6, 1870–1881. [Google Scholar] [CrossRef] [PubMed]
- Kersten, H.; Albani, M.; Männlein, E.; Praisler, R.; Wurmbach, P.; Nierhaus, K.H. On the role of ribosylthymine in prokaryotic tRNA function. Eur. J. Biochem. 1981, 114, 451–456. [Google Scholar] [CrossRef]
- Chou, H.J.; Donnard, E.; Gustafsson, H.T.; Garber, M.; Rando, O.J. Transcriptome-wide Analysis of Roles for tRNA Modifications in Translational Regulation. Mol. Cell 2017, 68, 978–992.e4. [Google Scholar] [CrossRef]
- Witzenberger, M.; Burczyk, S.; Settele, D.; Mayer, W.; Welp, L.M.; Heiss, M.; Wagner, M.; Monecke, T.; Janowski, R.; Carell, T.; et al. Human TRMT2A methylates tRNA and contributes to translation fidelity. Nucleic Acids Res. 2023, 51, 8691–8710. [Google Scholar] [CrossRef]
- Girodat, D.; Wieden, H.J.; Blanchard, S.C.; Sanbonmatsu, K.Y. Geometric alignment of aminoacyl-tRNA relative to catalytic centers of the ribosome underpins accurate mRNA decoding. Nat. Commun. 2023, 14, 5582. [Google Scholar] [CrossRef] [PubMed]
- Uhlenbeck, O.C.; Schrader, J.M. Evolutionary tuning impacts the design of bacterial tRNAs for the incorporation of unnatural amino acids by ribosomes. Curr. Opin. Chem. Biol. 2018, 46, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Favre, A.; Hajnsdorf, E.; Thiam, K.; Caldeira de Araujo, A. Mutagenesis and growth delay induced in Escherichia coli by near-ultraviolet radiations. Biochimie 1985, 67, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Favre, A.; Michelson, A.M.; Yaniv, M. Photochemistry of 4-thiouridine in Escherichia coli transfer RNA1Val. J. Mol. Biol. 1971, 58, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Carré, D.S.; Thomas, G.; Favre, A. Conformation and functioning of tRNAs: Cross-linked tRNAs as substrate for tRNA nucleotidyl-transferase and aminoacyl synthetases. Biochimie 1974, 56, 1089–1101. [Google Scholar] [CrossRef]
- Thomas, G.; Thiam, K.; Favre, A. tRNA thiolated pyrimidines as targets for near-ultraviolet-induced synthesis of guanosine tetraphosphate in Escherichia coli. Eur. J. Biochem. 1981, 119, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.; Favre, A. 4-Thiouridine as the target for near-ultraviolet light induced growth delay in Escherichia coli. Biochem. Biophys. Res. Commun. 1975, 66, 1454–1461. [Google Scholar] [CrossRef]
- Thiam, K.; Favre, A. Role of the stringent response in the expression and mechanism of near-ultraviolet induced growth delay. Eur. J. Biochem. 1984, 145, 137–142. [Google Scholar] [CrossRef]
- Kramer, G.F.; Baker, J.C.; Ames, B.N. Near-UV stress in Salmonella typhimurium: 4-thiouridine in tRNA, ppGpp, and ApppGpp as components of an adaptive response. J. Bacteriol. 1988, 170, 2344–2351. [Google Scholar] [CrossRef]
- Ramabhadran, T.V.; Jagger, J. Mechanism of growth delay induced in Escherichia coli by near ultraviolet radiation. Proc. Natl. Acad. Sci. USA 1976, 73, 59–63. [Google Scholar] [CrossRef]
- Ramabhadran, T.V.; Fossum, T.; Jagger, J. Escherichia coli mutant lacking 4-thiouridine in its transfer ribonucleic acid. J. Bacteriol. 1976, 128, 671–672. [Google Scholar] [CrossRef]
- Probst-Rüd, S.; McNeill, K.; Ackermann, M. Thiouridine residues in tRNAs are responsible for a synergistic effect of UVA and UVB light in photoinactivation of Escherichia coli. Environ. Microbiol. 2017, 19, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Gehrig, S.; Eberle, M.E.; Botschen, F.; Rimbach, K.; Eberle, F.; Eigenbrod, T.; Kaiser, S.; Holmes, W.M.; Erdmann, V.A.; Sprinzl, M.; et al. Identification of modifications in microbial, native tRNA that suppress immunostimulatory activity. J. Exp. Med. 2012, 209, 225–233. [Google Scholar] [CrossRef]
- Jöckel, S.; Nees, G.; Sommer, R.; Zhao, Y.; Cherkasov, D.; Hori, H.; Ehm, G.; Schnare, M.; Nain, M.; Kaufmann, A.; et al. The 2’-O-methylation status of a single guanosine controls transfer RNA-mediated Toll-like receptor 7 activation or inhibition. J. Exp. Med. 2012, 209, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, S.; Rimbach, K.; Eigenbrod, T.; Dalpke, A.H.; Helm, M. A modified dinucleotide motif specifies tRNA recognition by TLR7. RNA 2014, 20, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Rimbach, K.; Kaiser, S.; Helm, M.; Dalpke, A.H.; Eigenbrod, T. 2’-O-Methylation within Bacterial RNA Acts as Suppressor of TLR7/TLR8 Activation in Human Innate Immune Cells. J. Innate Immun. 2015, 7, 482–493. [Google Scholar] [CrossRef]
- Jain, V.; Kumar, M.; Chatterji, D. ppGpp: Stringent response and survival. J. Microbiol. 2006, 44, 1–10. [Google Scholar] [PubMed]
- De Bie, L.G.S.; Roovers, M.; Oudjama, Y.; Wattiez, R.; Tricot, C.; Stalon, V.; Droogmans, L.; Bujnicki, J.M. The yggH gene of Escherichia coli encodes a tRNA (m7G46) methyltransferase. J. Bacteriol. 2003, 185, 3238–3243. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, A.; Martzen, M.R.; Phizicky, E.M. Two proteins that form a complex are required for 7-methylguanosine modification of yeast tRNA. RNA 2002, 8, 1253–1266. [Google Scholar] [CrossRef] [PubMed]
- Thongdee, N.; Jaroensuk, J.; Atichartpongkul, S.; Chittrakanwong, J.; Chooyoung, K.; Srimahaeak, T.; Chaiyen, P.; Vattanaviboon, P.; Mongkolsuk, S.; Fuangthong, M. TrmB, a tRNA m7G46 methyltransferase, plays a role in hydrogen peroxide resistance and positively modulates the translation of katA and katB mRNAs in Pseudomonas aeruginosa. Nucleic Acids Res. 2019, 47, 9271–9281. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Liu, Q.; Lelyveld, V.S.; Choe, J.; Szostak, J.W.; Gregory, R.I. Mettl1/Wdr4-Mediated m7G tRNA Methylome Is Required for Normal mRNA Translation and Embryonic Stem Cell Self-Renewal and Differentiation. Mol. Cell 2018, 71, 244–255.e5. [Google Scholar] [CrossRef]
- Schultz, S.K.; Meadows, K.; Kothe, U. Molecular mechanism of tRNA binding by the Escherichia coli N7 guanosine methyltransferase TrmB. J. Biol. Chem. 2023, 299, 104612. [Google Scholar] [CrossRef]
- McGuffey, J.C.; Jackson-Litteken, C.D.; Di Venanzio, G.; Zimmer, A.A.; Lewis, J.M.; Distel, J.S.; Kim, K.Q.; Zaher, H.S.; Alfonzo, J.; Scott, N.E.; et al. The tRNA methyltransferase TrmB is critical for Acinetobacter baumannii stress responses and pulmonary infection. mBio 2023, 14, e0141623. [Google Scholar] [CrossRef]
- Takano, Y.; Takayanagi, N.; Hori, H.; Ikeuchi, Y.; Suzuki, T.; Kimura, A.; Okuno, T. A gene involved in modifying transfer RNA is required for fungal pathogenicity and stress tolerance of Colletotrichum lagenarium. Mol. Microbiol. 2006, 60, 81–92. [Google Scholar] [CrossRef]
- Kumagai, I.; Watanabe, K.; Oshima, T. Thermally induced biosynthesis of 2’-O-methylguanosine in tRNA from an extreme thermophile, Thermus thermophilus HB27. Proc. Natl. Acad. Sci. USA 1980, 77, 1922–1926. [Google Scholar] [CrossRef]
- Babosan, A.; Fruchard, L.; Krin, E.; Carvalho, A.; Mazel, D.; Baharoglu, Z. Nonessential tRNA and rRNA modifications impact the bacterial response to sub-MIC antibiotic stress. Microlife 2022, 3, uqac019. [Google Scholar] [CrossRef]
- Yoluç, Y.; van de Logt, E.; Kellner-Kaiser, S. The Stress-Dependent Dynamics of Saccharomyces cerevisiae tRNA and rRNA Modification Profiles. Genes 2021, 12, 1344. [Google Scholar] [CrossRef]
- Shigi, N.; Suzuki, T.; Terada, T.; Shirouzu, M.; Yokoyama, S.; Watanabe, K. Temperature-dependent biosynthesis of 2-thioribothymidine of Thermus thermophilus tRNA. J. Biol. Chem. 2006, 281, 2104–2113. [Google Scholar] [CrossRef]
- Hou, Y.M.; Masuda, I.; Foster, L.J. tRNA methylation: An unexpected link to bacterial resistance and persistence to antibiotics and beyond. Wiley Interdiscip. Rev. RNA 2020, 11, e1609. [Google Scholar] [CrossRef]
- Burby, P.E.; Nye, T.M.; Schroeder, J.W.; Simmons, L.A. Implementation and Data Analysis of Tn-seq, Whole-Genome Resequencing, and Single-Molecule Real-Time Sequencing for Bacterial Genetics. J. Bacteriol. 2017, 199, 10-1128. [Google Scholar] [CrossRef]
- Fruchard, L.; Babosan, A.; Carvalho, A.; Lang, M.; Li, B.; Duchateau, M.; Giai-Gianetto, Q.; Matondo, M.; Bonhomme, F.; Hatin, I.; et al. Aminoglycoside tolerance in Vibrio cholerae engages translational reprogramming associated to queuosine tRNA modification. bioRxiv 2024. [Google Scholar] [CrossRef]
- Irving, S.E.; Choudhury, N.R.; Corrigan, R.M. The stringent response and physiological roles of (pp)pGpp in bacteria. Nat. Rev. Microbiol. 2021, 19, 256–271. [Google Scholar] [CrossRef]
- Huang, H.Y.; Hopper, A.K. Multiple Layers of Stress-Induced Regulation in tRNA Biology. Life 2016, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.E.; Alexander, R.W.; Pan, T. Misacylation of specific nonmethionyl tRNAs by a bacterial methionyl-tRNA synthetase. Proc. Natl. Acad. Sci. USA 2011, 108, 6933–6938. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kim, D.G.; Kim, B.G.; Yang, W.S.; Hong, J.; Kang, T.; Oh, Y.S.; Kim, K.R.; Han, B.W.; Hwang, B.J.; et al. Promiscuous methionyl-tRNA synthetase mediates adaptive mistranslation to protect cells against oxidative stress. J. Cell Sci. 2014, 127, 4234–4245. [Google Scholar] [CrossRef] [PubMed]
- Lyons, S.M.; Fay, M.M.; Ivanov, P. The role of RNA modifications in the regulation of tRNA cleavage. FEBS Lett. 2018, 592, 2828–2844. [Google Scholar] [CrossRef]
- Shetty, S.; Varshney, U. Regulation of translation by one-carbon metabolism in bacteria and eukaryotic organelles. J. Biol. Chem. 2021, 296, 100088. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yared, M.-J.; Marcelot, A.; Barraud, P. Beyond the Anticodon: tRNA Core Modifications and Their Impact on Structure, Translation and Stress Adaptation. Genes 2024, 15, 374. https://doi.org/10.3390/genes15030374
Yared M-J, Marcelot A, Barraud P. Beyond the Anticodon: tRNA Core Modifications and Their Impact on Structure, Translation and Stress Adaptation. Genes. 2024; 15(3):374. https://doi.org/10.3390/genes15030374
Chicago/Turabian StyleYared, Marcel-Joseph, Agathe Marcelot, and Pierre Barraud. 2024. "Beyond the Anticodon: tRNA Core Modifications and Their Impact on Structure, Translation and Stress Adaptation" Genes 15, no. 3: 374. https://doi.org/10.3390/genes15030374
APA StyleYared, M.-J., Marcelot, A., & Barraud, P. (2024). Beyond the Anticodon: tRNA Core Modifications and Their Impact on Structure, Translation and Stress Adaptation. Genes, 15(3), 374. https://doi.org/10.3390/genes15030374