
Sequence Analysis of Six Candidate Genes in Miniature Schnauzers with Primary Hypertriglyceridemia

 , , ,
, , ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Miniature Schnauzers WGS Cohort
2.2. DNA Isolation and WGS
2.3. Candidate Gene Analysis
2.4. Variant Follow-Up Genotyping
3. Results
3.1. Miniature Schnauzer WGS Cohort
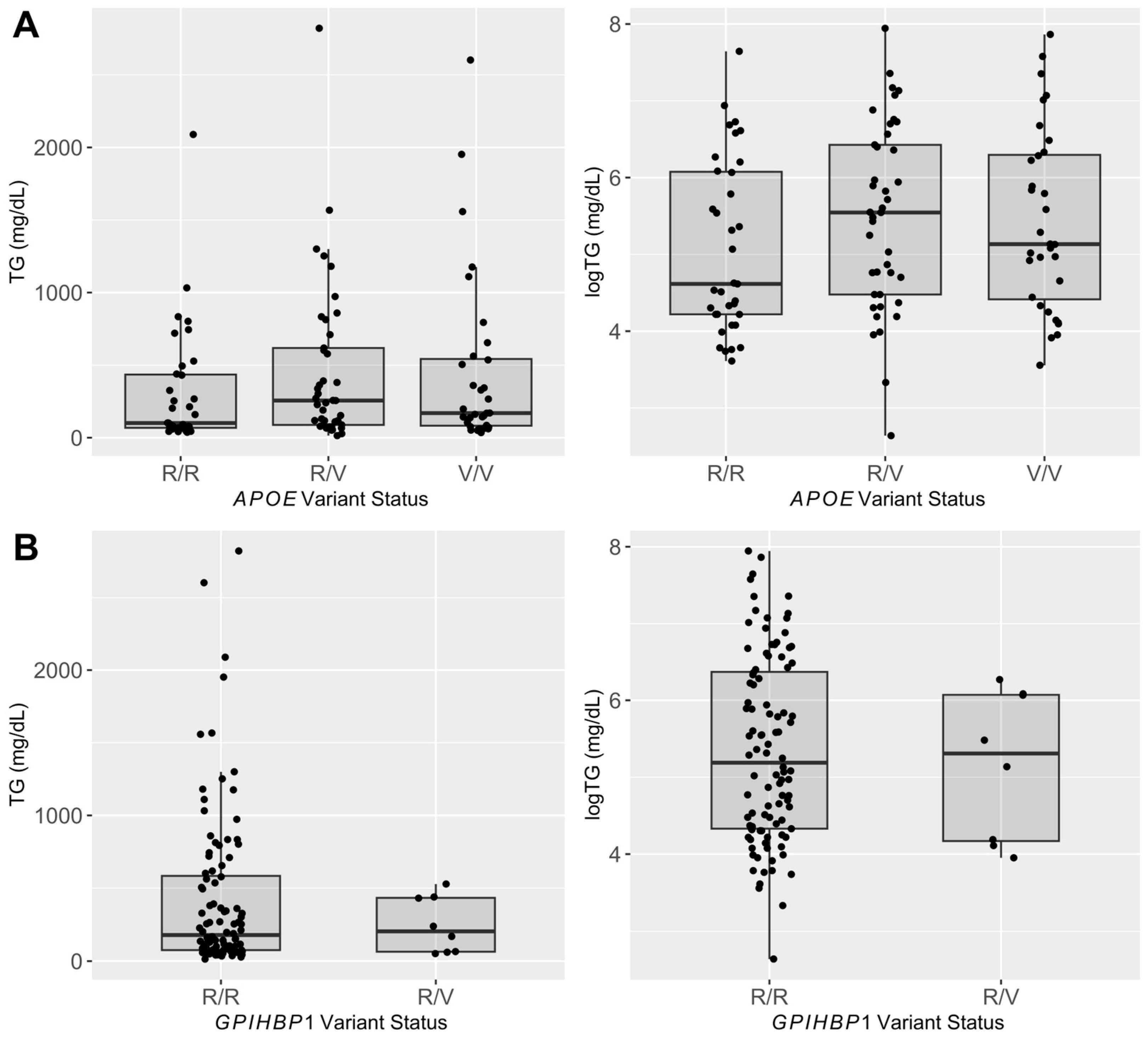
3.2. Candidate Gene Analysis
3.3. Variant Follow-Up Genotyping
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xenoulis, P.G.; Suchodolski, J.S.; Levinski, M.D.; Steiner, J.M. Investigation of hypertriglyceridemia in healthy Miniature Schnauzers. J. Vet. Intern. Med. 2007, 21, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Xenoulis, P.G.; Levinski, M.D.; Suchodolski, J.S.; Steiner, J.M. Serum triglyceride concentrations in Miniature Schnauzers with and without a history of probable pancreatitis. J. Vet. Intern. Med. 2011, 25, 20–25. [Google Scholar] [CrossRef]
- Kutsunai, M.; Kanemoto, H.; Fukushima, K.; Fujino, Y.; Ohno, K.; Tsujimoto, H. The association between gall bladder mucoceles and hyperlipidaemia in dogs: A retrospective case control study. Vet. J. 2014, 199, 76–79. [Google Scholar] [CrossRef]
- Furrow, E.; Jaeger, J.Q.; Parker, V.J.; Hinchcliff, K.W.; Johnson, S.E.; Murdoch, S.J.; de Boer, I.H.; Sherding, R.G.; Brunzell, J.D. Proteinuria and lipoprotein lipase activity in Miniature Schnauzer dogs with and without hypertriglyceridemia. Vet. J. 2016, 212, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Surendran, R.P.; Visser, M.E.; Heemelaar, S.; Wang, J.; Peter, J.; Defesche, J.C.; Kuivenhoven, J.A.; Hosseini, M.; Peterfy, M.; Kastelein, J.J.P.; et al. Mutations in LPL, APOC2, APOA5, GPIHBP1, and LMF1 in patients with severe hypertriglyceridemia. J. Intern. Med. 2012, 272, 185–196. [Google Scholar] [CrossRef]
- Schickel, R. Identification of the Nucleotide Sequence of the Lipoprotein Lipase Gene as well as Its Role in the Development of Hyperlipidemia and Pancreatitis in the Miniature Schnauzer. Ph.D. Thesis, Ludwig-Maximilians University, München, Germany, 2005. [Google Scholar]
- Xenoulis, P.G.; Tate, N.M.; Bishop, M.A.; Steiner, J.M.; Suchodolski, J.; Furrow, E. Sequence analysis of the coding regions of the apolipoprotein C2 (APOC2) gene in Miniature Schnauzers with idiopathic hypertriglyceridemia. Vet. J. 2020, 265, 105559. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Lupski, J.R. Non-coding genetic variants in human disease. Hum. Mol. Genet. 2015, 24, R102–R110. [Google Scholar] [CrossRef]
- Furrow, E.; Lees, G.E.; Brown, C.A.; Cianciolo, R.E. Glomerular lesions in proteinuric Miniature Schnauzer dogs. Vet. Pathol. 2017, 54, 484–489. [Google Scholar] [CrossRef]
- Khalil, Y.A.; Rabès, J.P.; Boileau, C.; Varret, M. APOE gene variants in primary dyslipidemia. Atherosclerosis 2021, 328, 11–22. [Google Scholar] [CrossRef]
- Turro, E.; Astle, W.J.; Megy, K.; Gräf, S.; Greene, D.; Shamardina, O.; Allen, H.L.; Sanchis-Juan, A.; Frontini, M.; Thys, C.; et al. Whole-genome sequencing of patients with rare diseases in a national health system. Nature 2020, 583, 96–102. [Google Scholar] [CrossRef]
- van Steenbeek, F.G.; Hytönen, M.K.; Leegwater, P.A.J.; Lohi, H. The canine era: The rise of a biomedical model. Anim Genet. 2016, 47, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Cullen, J.N.; Friedenberg, S.G. WAGS: User-friendly, rapid, containerized pipelines for processing, variant discovery, and annotation of short read whole genome sequencing data. G3 2023, 27, jkad117. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wallerman, O.; Arendt, M.-L.; Sundström, E.; Karlsson, A.; Nordin, J.; Mäkeläinen, S.; Pielberg, G.R.; Hanson, J.; Ohlsson, Å.; et al. A novel canine reference genome resolves genomic architecture and uncovers transcript complexity. Commun. Biol. 2021, 4, 185. [Google Scholar] [CrossRef] [PubMed]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl variant effect predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed]
- Haberle, V.; Stark, A. Eukaryotic core promoters and the functional basis of transcription initiation. Nat. Rev. Mol. Cell Biol. 2018, 19, 621–637. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, M.B.; Senapathy, P. RNA splice junctions of different classes of eukaryotes: Sequence statistics and functional implications in gene expression. Nucleic Acids Res. 1987, 15, 7155–7174. [Google Scholar] [CrossRef]
- Usui, S.; Mizoguchi, Y.; Yasuda, H.; Arai, N.; Koketsu, Y. Dog age and breeds associated with high plasma cholesterol and triglyceride concentrations. J. Vet. Med. Sci. 2014, 76, 269–272. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wu, S.A.; Kersten, S.; Qi, L. Lipoprotein Lipase and Its Regulators: An Unfolding Story. Trends Endocrinol. Metab. 2021, 32, 48–61. [Google Scholar] [CrossRef]
- LaRosa, J.C.; Levy, R.I.; Herbert, P.N.; Lux, S.E.; Fredrickson, D.S. A specific apoprotein activator for lipoprotein lipase. Biochem. Biophys. Res. Commun. 1970, 41, 57–62. [Google Scholar] [CrossRef]
- Lookene, A.; Beckstead, J.A.; Nilsson, S.; Olivecrona, G.; Ryan, R.O. Apolipoprotein A-V-heparin interactions: Implications for plasma lipoprotein metabolism. J. Biol. Chem. 2005, 280, 25383–25387. [Google Scholar] [CrossRef]
- Peterfy, M.; Ben-Zeev, O.; Mao, H.Z.; Weissglas-Volkov, D.; Aouizerat, B.E.; Pullinger, C.R.; Frost, P.H.; Kane, J.P.; Malloy, M.J.; Reue, K.; et al. Mutations in LMF1 cause combined lipase deficiency and severe hypertriglyceridemia. Nature 2007, 39, 1483–1487. [Google Scholar] [CrossRef]
- Young, S.G.; Fong, L.G.; Beigneux, A.P.; Allan, C.M.; He, C.; Jiang, H.; Nakajima, K.; Meiyappan, M.; Birrane, G.; Ploug, M. GPIHBP1 and lipoprotein lipase, partners in plasma triglyceride metabolism. Cell Metab. 2019, 30, 51–65. [Google Scholar] [CrossRef]
- Pollard, K.S.; Hubisz, M.J.; Rosenbloom, K.R.; Siepel, A. Detection of nonneutral substitution rates on mammalian phylogenies. Genome Res. 2010, 20, 110–121. [Google Scholar] [CrossRef]
- Blanchette, M.; Kent, W.J.; Reimer, C.; Elnitski, L.; Smit, A.F.; Roskin, K.M.; Baertsch, R.; Rosenbloom, K.; Clawson, H.; Green, E.D.; et al. Aligning multiple genomic sequences with the threaded blockset aligner. Genome Res. 2004, 14, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, R.; Capriotti, E.; Fariselli, P.; Martelli, P.L.; Casadio, R. Functional annotations improve the predictive score of human disease-related mutations in proteins. Hum. Mutat. 2009, 30, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan5:genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef]
- Furrow, E.; Patterson, E.E.; Armstrong, P.J.; Osborne, C.A.; Lulich, J.P. Fasting urinary calcium-to-creatinine and oxalate-to-creatinine ratios in dogs with calcium oxalate urolithiasis and breed-matched controls. J. Vet. Intern. Med. 2015, 29, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.E.; Granick, J.L.; Stauthammer, C.D.; Polzin, D.J.; Heinrich, D.A.; Furrow, E. Clinical consequences of hypertriglyceridemia associated proteinuria in Miniature Schnauzers. J. Vet. Intern. Med. 2017, 31, 1740–1748. [Google Scholar] [CrossRef] [PubMed]
- Vincze, T.; Posfai, J.; Roberts, R.J. NEBcutter: A program to cleave DNA with restriction enzymes. Nucleic Acids Res. 2003, 31, 3688–3691. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org (accessed on 18 November 2022).
- Horuchi, K.; Tajima, S.; Menju, M.; Yamamoto, A. Structure and expression of mouse apolipoprotein E gene. J. Biochem. 1989, 106, 98–103. [Google Scholar] [CrossRef]
- Maloney, B.; Ge, Y.W.; Alley, G.M.; Lahiri, D.K. Important differences between human and mouse APOE gene promoters: Limitation of mouse APOE model in studying Alzheimer’s disease. J. Neurochem. 2007, 103, 1237–1257. [Google Scholar] [CrossRef]
- Bae, S.H.; Han, H.W.; Moon, J. Functional analysis of the molecular interactions of TATA box-containing genes and essential genes. PLoS ONE 2015, 10, e0120848. [Google Scholar] [CrossRef] [PubMed]
- Zukunft, J.; Lang, T.; Richter, T.; Hirsch-Ernst, K.I.; Nussler, A.K.; Klein, K.; Schwab, M.; Eichelbaum, M.; Zanger, U.M. A natural CYP2B6 TATA box polymorphism (82T-> C) leading to enhanced transcription and relocation of the transcriptional start site. Mol. Pharmacol. 2005, 67, 1772–1782. [Google Scholar] [CrossRef]
- Matsunaga, A.; Sasaki, J.; Han, H.; Huang, W.; Kugi, M.; Koga, T.; Ichiki, S.; Shinkawa, T.; Arakawa, K. Compound heterozygosity for an apolipoprotein A1 gene promoter mutation and a structural nonsense mutation with apolipoprotein A1 deficiency. Arterioscler Thromb. Vasc. Biol. 1999, 19, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xuemei Zhang Fan, P.; Liu, R.; Huang, Y.; Liang, S.; Liu, Y.; Wu, Y.; Bai, H. Distribution and effect of apo E genotype on plasma lipid and apolipoprotein profiles in overweight/obese and nonobese Chinese subjects. J. Clin. Lab. Anal. 2012, 26, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Kofler, B.M.; Miles, E.A.; Curtis, P.; Armah, C.K.; Tricon, S.; Grew, J.; Napper, F.L.; Farrell, L.; Lietz, G.; Packard, C.J.; et al. Apolipoprotein E genotype and the cardiovascular disease risk phenotype: Impact of sex and adiposity (the FINGEN study). Atherosclerosis 2012, 221, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.D.; Tsongalis, G.J.; Seip, R.L.; Bilbie, C.; Miles, M.; Zoeller, R.; Visich, P.; Gordon, P.; Angelopoulos, T.J.; Pescatello, L.; et al. Apolipoprotein E genotype and changes in serum lipids and maximal oxygen uptake with exercise training. Metabolism 2004, 53, 193–202. [Google Scholar] [CrossRef]
- Tucker, A.J.; Mackay, K.A.; Robinson, L.E.; Graham, T.E.; Bakovic, M.; Duncan, A.M. The effect of whole grain wheat sourdough bread consumption on serum lipids in healthy normoglycemic/normoinsulinemic and hyperglycemic/hyperinsulinemic adults depends on presence of the APOE E3/E3 genotype: A randomized controlled trial. Nutr. Metab. 2010, 7, 37. [Google Scholar] [CrossRef]
- Zeng, Y.; Wen, S.; Huan, L.; Xiong, L.; Zhong, B.; Wang, P. Association of ApoE gene polymorphisms with serum lipid levels and the risk of type 2 diabetes mellitus in the Chinese Han population of central China. PeerJ 2023, 11, e15226. [Google Scholar] [CrossRef]
- Vaz-Drago, R.; Custódio, N.; Carmo-Fonseca, M. Deep intronic mutations and human disease. Hum. Genet. 2017, 136, 1093–1111. [Google Scholar] [CrossRef]
- Dron, J.S.; Wang, J.; McIntyre, A.D.; Cao, H.; Robinson, J.F.; Duell, P.B.; Manjoo, P.; Feng, J.; Movsesyan, I.; Malloy, M.J.; et al. Partial LPL deletions: Rare copy-number variants contributing towards severe hypertriglyceridemia. J. Lipid Res. 2019, 60, 1593–1958. [Google Scholar] [CrossRef] [PubMed]
- Rios, J.J.; Shastry, S.; Jasso, J.; Hauser, N.; Garg, A.; Bensadoun, A.; Cohen, J.C.; Hobbs, H.H. Deletion of GPIHBP1 causing severe hyperchylomicronemia. J. Inherit Metab. Dis. 2012, 35, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Patni, N.; Brothers, J.; Xing, C.; Garg, A. Type 1 hyperlipoproteinemia in a child with large homozygous deletion encompassing GPIHBP1. J. Clin. Lipidol. 2016, 10, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cao, H.; Ban, M.R.; Kennedy, B.A.; Zhu, S.; Anand, S.; Yusuf, S.; Pollex, R.L.; Hegele, R.A. Resequencing genomic DNA of patients with severe hypertriglyceridemia (MIM 144650). Arterioscler Thromb. Vasc. Biol. 2007, 27, 2450–2455. [Google Scholar] [CrossRef]
- Lewis, G.F.; Xiao, C.; Hegele, R.A. Hypertriglyceridemia in the genomic era: A new paradigm. Endocr. Rev. 2015, 36, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, M.C.; van Greevenbroek, M.M.; Stehouwer, C.D.; de Graaf, J.; Stalenhoef, A.F. The genetics of familial combined hyperlipidaemia. Nat. Rev. Endocrinol. 2012, 8, 352–362. [Google Scholar] [CrossRef]
- Kersten, S. Physiological regulation of lipoprotein lipase. Biochim. Biophys. Acta 2014, 1841, 919–933. [Google Scholar] [CrossRef]
- Dron, J.S.; Wang, J.; Cao, H.; McIntyre, A.D.; Iacocca, M.A.; Menard, J.R.; Movsesyan, I.; Malloy, M.J.; Pullinger, C.R.; Kane, J.P.; et al. Severe hypertriglyceridemia is primarily polygenic. J. Clin. Lipidol. 2019, 13, 80–88. [Google Scholar] [CrossRef]
- Dron, J.S.; Wang, J.; McIntyre, A.D.; Cao, H.; Hegele, R.A. The polygenic nature of mild-to-moderate hypertriglyceridemia. J. Clin. Lipidol. 2020, 14, 28–34.e2. [Google Scholar] [CrossRef] [PubMed]
- Xenoulis, P.G.; Cammarata, P.J.; Walzem, R.L.; Suchodolski, J.S.; Steiner, J.M. Effect of a low-fat diet on serum triglyceride and cholesterol concentrations and lipoprotein profiles in Miniature Schnauzers with hypertriglyceridemia. J. Vet. Intern. Med. 2020, 34, 2605–2616. [Google Scholar] [CrossRef] [PubMed]
- de Albuquerque, P.; De Marco, V.; Vendramini, T.H.A.; Amaral, A.R.; Catanozi, S.; Santana, K.G.; Nunes, V.S.; Nakandakare, E.R.; Brunetto, M.A. Supplementation of omega-3 and dietary factors can influence the cholesterolemia and triglyceridemia in hyperlipidemic Schnauzer dogs: A preliminary report. PLoS ONE 2021, 16, e0258058. [Google Scholar] [CrossRef] [PubMed]
- Kawasumi, K.; Kashiwado, N.; Okada, Y.; Sawamura, M.; Sasaki, Y.; Iwazaki, E.; Mori, N.; Yamamoto, I.; Arai, T. Age effects on plasma cholesterol and triglyceride profiles and metabolite concentrations in dogs. BMC Vet. Res. 2014, 10, 57. [Google Scholar] [CrossRef]


{kind=link}
| Gene Symbol | Gene Name | Function |
|---|---|---|
| LPL | Lipoprotein lipase | Hydrolyzes circulating triglycerides [19] |
| APOC2 | Apolipoprotein C-II | Essential cofactor for LPL activity [19,20] |
| APOA5 | Apolipoprotein A-V | Stimulates LPL-mediated triglyceride hydrolysis [19,21] |
| LMF1 | Lipase maturation factor 1 | Essential for proper LPL function [19,22] |
| GPIHBP1 | Glycosylphosphatidylinositol-anchored HDL binding protein 1 | Facilitates LPL toward cell surface and promotes processing of triglyceride-rich lipoproteins [19,23] |
| APOE | Apolipoprotein E | Regulates clearance of chylomicron remnants and VLDL [10] |
| Gene | Description | Variant Type | PhyloP Score | MS HTG | Other MS * | Non-MS Breeds † | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R/R | R/V | V/V | AF | R/R | R/V | V/V | AF | R/R | R/V | V/V | AF | ||||
| APOE | g.111237170_ 111237172del | 5′ UTR | 1.95, 0.83, 0.43 | 6 | 2 | 0 | 0.13 | 14 | 14 | 2 | 0.30 | 613 | 0 | 0 | 0 |
| LMF1 | g.4020228C>A | intronic | NA †† | 1 | 4 | 3 | 0.63 | 8 | 11 | 11 | 0.55 | 610 | 3 | 0 | 0.002 |
| GPIHBP1 | g.37746233C>T | missense | −3.66 | 6 | 2 | 0 | 0.13 | 30 | 0 | 0 | 0 | 599 | 1 | 0 | 0 |
| Variable | Estimate of the Coefficient | Std. Error | p Value |
|---|---|---|---|
| APOE Genotype | 0.18 | 0.14 | 0.20 |
| GPIHBP1 Genotype | −0.26 | 0.42 | 0.54 |
| Age (years) | 0.15 | 0.05 | 0.006 |
| Sex (male) | 0.01 | 0.23 | 0.97 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tate, N.M.; Underwood, M.; Thomas-Hollands, A.; Minor, K.M.; Cullen, J.N.; Friedenberg, S.G.; Mickelson, J.R.; Xenoulis, P.G.; Steiner, J.M.; Furrow, E. Sequence Analysis of Six Candidate Genes in Miniature Schnauzers with Primary Hypertriglyceridemia. Genes 2024, 15, 193. https://doi.org/10.3390/genes15020193
Tate NM, Underwood M, Thomas-Hollands A, Minor KM, Cullen JN, Friedenberg SG, Mickelson JR, Xenoulis PG, Steiner JM, Furrow E. Sequence Analysis of Six Candidate Genes in Miniature Schnauzers with Primary Hypertriglyceridemia. Genes. 2024; 15(2):193. https://doi.org/10.3390/genes15020193
Chicago/Turabian StyleTate, Nicole M., Michaela Underwood, Alison Thomas-Hollands, Katie M. Minor, Jonah N. Cullen, Steven G. Friedenberg, James R. Mickelson, Panagiotis G. Xenoulis, Joerg M. Steiner, and Eva Furrow. 2024. "Sequence Analysis of Six Candidate Genes in Miniature Schnauzers with Primary Hypertriglyceridemia" Genes 15, no. 2: 193. https://doi.org/10.3390/genes15020193
APA StyleTate, N. M., Underwood, M., Thomas-Hollands, A., Minor, K. M., Cullen, J. N., Friedenberg, S. G., Mickelson, J. R., Xenoulis, P. G., Steiner, J. M., & Furrow, E. (2024). Sequence Analysis of Six Candidate Genes in Miniature Schnauzers with Primary Hypertriglyceridemia. Genes, 15(2), 193. https://doi.org/10.3390/genes15020193