
Gene Selection of Methionine-Dependent Melanoma and Independent Melanoma by Variable Selection Using Tensor Decomposition



Highlights
- We introduced tensor decomposition-based feature extraction for gene selection from the gene expression profiles determined RNA sequencing.
- An enrichment analysis of the selected gene set revealed findings consistent with prior studies on methionine dependency in melanoma.
- This research offers new insights into the molecular mechanisms of melanoma, which could lead to improved diagnostic and therapeutic strategies.
- Our method has the potential to reveal novel insights based on transcriptomic and other large-scale molecular datasets.
Abstract
1. Introduction
2. Materials and Methods
2.1. Tensor Decomposition
2.2. Gene Expression Profile
2.3. TDbasedUFE
2.4. Enrichment Analysis
3. Results
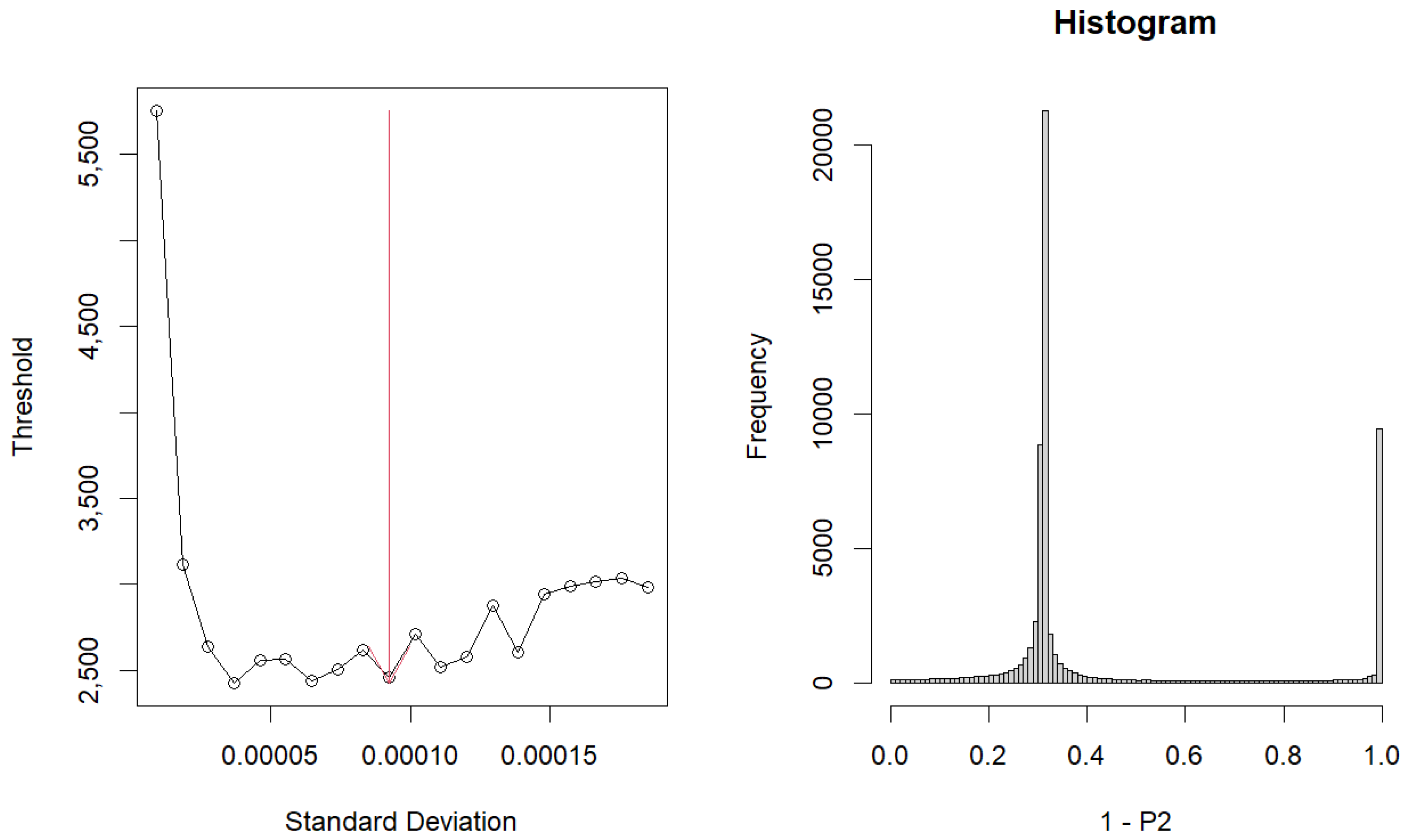
3.1. Gene Selection
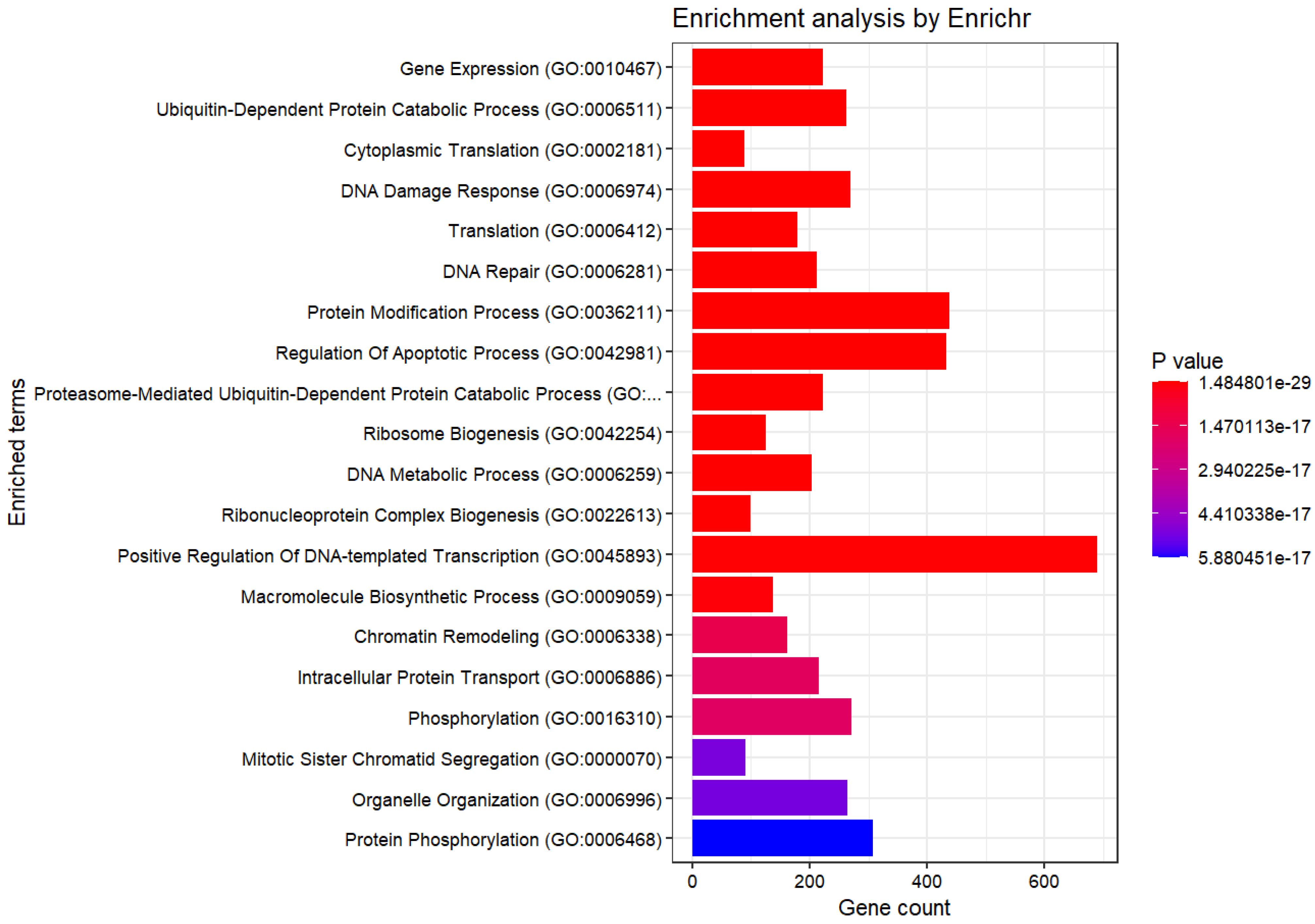
3.2. Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perrone, C.E.; Malloy, V.L.; Orentreich, D.S.; Orentreich, N. Metabolic adaptations to methionine restriction that benefit health and lifespan in rodents. Exp. Gerontol. 2013, 48, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Kreis, W.; Hession, C. Biological effects of enzymatic deprivation of L-methionine in cell culture and an experimental tumor. Cancer Res. 1973, 33, 1866–1869. [Google Scholar] [PubMed]
- Halpern, B.C.; Clark, B.R.; Hardy, D.N.; Halpern, R.M.; Smith, R.A. The effect of replacement of methionine by homocystine on survival of malignant and normal adult mammalian cells in culture. Proc. Natl. Acad. Sci. USA 1974, 71, 1133–1136. [Google Scholar] [CrossRef]
- Hall, C.A.; Begley, J.A.; Chu, R.C. Methionine dependency of cultured human lymphocytes. Proc. Soc. Exp. Biol. Med. 1986, 182, 215–220. [Google Scholar] [CrossRef]
- Taguchi, Y.; Turki, T. Application note: TDbasedUFE and TDbasedUFEadv: Bioconductor packages to perform tensor decomposition based unsupervised feature extraction. Front. Artif. Intell. 2023, 6, 1237542. [Google Scholar] [CrossRef]
- Taguchi, Y.; Turki, T. Tensor decomposition-based unsupervised feature extraction applied to single-cell gene expression analysis. Front. Genet. 2019, 10, 864. [Google Scholar] [CrossRef]
- Taguchi, Y. Unsupervised Feature Extraction Applied to Bioinformatics: A PCA Based and TD Based Approach; Springer International Publishing AG: Cham, Switzerland, 2019. [Google Scholar]
- Garg, S.; Morehead, L.C.; Bird, J.T.; Graw, S.; Gies, A.; Storey, A.J.; Tackett, A.J.; Edmondson, R.D.; Mackintosh, S.G.; Byrum, S.D.; et al. Characterization of methionine dependence in melanoma cells. bioRxiv 2023. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene set knowledge discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Martínez, Y.; Li, X.; Liu, G.; Bin, P.; Yan, W.; Más, D.; Valdivié, M.; Hu, C.A.A.; Ren, W.; Yin, Y. The role of methionine on metabolism, oxidative stress, and diseases. Amino Acids 2017, 49, 2091–2098. [Google Scholar] [CrossRef]
- Kokkinakis, D.M.; Brickner, A.G.; Kirkwood, J.M.; Liu, X.; Goldwasser, J.E.; Kastrama, A.; Sander, C.; Bocangel, D.; Chada, S. Mitotic arrest, apoptosis, and sensitization to chemotherapy of melanomas by methionine deprivation stress. Mol. Cancer Res. 2006, 4, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, P. Methionine dependence of cancer. Biomolecules 2020, 10, 568. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, S.M.; Gao, X.; Dai, Z.; Locasale, J.W. Methionine metabolism in health and cancer: A nexus of diet and precision medicine. Nat. Rev. Cancer 2019, 19, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Xuan, M.; Gu, X.; Li, J.; Huang, D.; Xue, C.; He, Y. Polyamines: Their significance for maintaining health and contributing to diseases. Cell Commun. Signal. 2023, 21, 348. [Google Scholar] [CrossRef]
- Strekalova, E.; Malin, D.; Weisenhorn, E.M.; Russell, J.D.; Hoelper, D.; Jain, A.; Coon, J.J.; Lewis, P.W.; Cryns, V.L. S-adenosylmethionine biosynthesis is a targetable metabolic vulnerability of cancer stem cells. Breast Cancer Res. Treat. 2019, 175, 39–50. [Google Scholar] [CrossRef]
- Strekalova, E.; Malin, D.; Good, D.M.; Cryns, V.L. Methionine deprivation induces a targetable vulnerability in triple-negative breast cancer cells by enhancing TRAIL receptor-2 expression. Clin. Cancer Res. 2015, 21, 2780–2791. [Google Scholar] [CrossRef]
- Beetstra, S.; Suthers, G.; Dhillon, V.; Salisbury, C.; Turner, J.; Altree, M.; McKinnon, R.; Fenech, M. Methionine-dependence phenotype in the de novo pathway in BRCA1 and BRCA2 mutation carriers with and without breast cancer. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 2565–2571. [Google Scholar] [CrossRef]
- Jeon, H.; Kim, J.H.; Lee, E.; Jang, Y.J.; Son, J.E.; Kwon, J.Y.; Lim, T.g.; Kim, S.; Park, J.H.Y.; Kim, J.E.; et al. Methionine deprivation suppresses triple-negative breast cancer metastasis in vitro and in vivo. Oncotarget 2016, 7, 67223. [Google Scholar] [CrossRef] [PubMed]
- Villa, E.; Sahu, U.; O’Hara, B.P.; Ali, E.S.; Helmin, K.A.; Asara, J.M.; Gao, P.; Singer, B.D.; Ben Sahra, I. mTORC1 stimulates cell growth through SAM synthesis and m6A mRNA-dependent control of protein synthesis. Mol. Cell 2021, 81, 2076–2093. [Google Scholar] [CrossRef]
- Hassan, Y.A.; Helmy, M.W.; Ghoneim, A.I. Combinatorial antitumor effects of amino acids and epigenetic modulations in hepatocellular carcinoma cell lines. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2021, 394, 2245–2257. [Google Scholar] [CrossRef]
- Walvekar, A.S.; Laxman, S. Methionine at the heart of anabolism and signaling: Perspectives from budding yeast. Front. Microbiol. 2019, 10, 2624. [Google Scholar] [CrossRef]
- Wu, X.; Tu, B.P. Selective regulation of autophagy by the Iml1-Npr2-Npr3 complex in the absence of nitrogen starvation. Mol. Biol. Cell 2011, 22, 4124–4133. [Google Scholar] [CrossRef]
- Sutter, B.M.; Wu, X.; Laxman, S.; Tu, B.P. Methionine inhibits autophagy and promotes growth by inducing the SAM-responsive methylation of PP2A. Cell 2013, 154, 403–415. [Google Scholar] [CrossRef]
- Brown, T. Genomes, 4th ed.; Garland Science: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Shiraki, N.; Shiraki, Y.; Tsuyama, T.; Obata, F.; Miura, M.; Nagae, G.; Aburatani, H.; Kume, K.; Endo, F.; Kume, S. Methionine metabolism regulates maintenance and differentiation of human pluripotent stem cells. Cell Metab. 2014, 19, 780–794. [Google Scholar] [CrossRef]
- Borini Etichetti, C.M.; Arel Zalazar, E.; Cocordano, N.; Girardini, J. Beyond the Mevalonate Pathway: Control of Post-Prenylation Processing by Mutant p53. Front. Oncol. 2020, 10, 595034. [Google Scholar] [CrossRef]
- Benavides, M.A.; Oelschlager, D.K.; Zhang, H.G.; Stockard, C.R.; Vital Reyes, V.S.; Katkoori, V.R.; Manne, U.; Wang, W.; Bland, K.I.; Grizzle, W.E. Methionine inhibits cellular growth dependent on the p53 status of cells. Am. J. Surg. 2007, 193, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | Overlap | Adjusted p-Value |
|---|---|---|
| Gene Expression (GO:0010467) | 223/296 | |
| Ubiquitin-Dependent Protein Catabolic Process (GO:0006511) | 262/367 | |
| Cytoplasmic Translation (GO:0002181) | 89/93 | |
| DNA Damage Response (GO:0006974) | 270/384 | |
| Translation (GO:0006412) | 179/234 | |
| DNA Repair (GO:0006281) | 212/292 | |
| Protein Modification Process (GO:0036211) | 437/711 | |
| Regulation Of Apoptotic Process (GO:0042981) | 432/705 | |
| Proteasome-Mediated Ubiquitin-Dependent Protein Catabolic Process (GO:0043161) | 222/319 | |
| Ribosome Biogenesis (GO:0042254) | 125/155 | |
| DNA Metabolic Process (GO:0006259) | 203/288 | |
| Ribonucleoprotein Complex Biogenesis (GO:0022613) | 100/118 | |
| Positive Regulation Of DNA-templated Transcription (GO:0045893) | 690/1243 | |
| Macromolecule Biosynthetic Process (GO:0009059) | 138/183 | |
| Chromatin Remodeling (GO:0006338) | 162/228 | |
| Intracellular Protein Transport (GO:0006886) | 216/325 | |
| Phosphorylation (GO:0016310) | 272/429 | |
| Mitotic Sister Chromatid Segregation (GO:0000070) | 91/111 | |
| Organelle Organization (GO:0006996) | 265/418 | |
| Protein Phosphorylation (GO:0006468) | 308/500 |
| Term | Overlap | Adjusted p-Value |
|---|---|---|
| Cell Cycle Overview | 91/107 | |
| Brest Cancer | 91/108 | |
| Pancreatic Ductal Carcinoma | 94/1117 | |
| Melanoma | 110/145 | |
| Proteins with Altered Expression in Cancer Metabolic Reprogramming | 72/85 | |
| Metabolic Effects of Oncogenes and Tumor Suppressor in Cancer Cells | 60/68 | |
| Hepatocellular Carcinoma | 88/112 | |
| Chronic Myeloid Leukemia | 61/70 | |
| Endometrioid Endometrial Cancer | 75/92 | |
| Protein Involved in Melanoma | 160/238 |
| Term | Overlap | Adjusted p-Value |
|---|---|---|
| HIV-1 Nef: negative effector of Fas and TNF Homo sapiens h HivnefPathway | 43/51 | |
| Control of Gene Expression by Vitamin D Receptor Homo sapiens h vdrPathway | 25/27 | |
| Influence of Ras and Rho proteins on G1 to S Transition Homo sapiens h RacCycDPathway | 25/28 | |
| Mechanism of Gene Regulation by Peroxisome Proliferators via PPARa Homo sapiens h pparaPathway | 40/52 | |
| Ceramide Signaling Pathway Homo sapiens h ceramide Pathway | 28/33 | |
| Integrin Signaling Pathway Homo sapiens h integrin Pathway | 28/33 | |
| Cell Cycle: G1/S Check Point Homo sapiens h g1 Pathway | 23/26 | |
| Inhibition of Cellular Proliferation by Gleevec Homo sapiens h Gleevec pathway | 20/22 | |
| Skeletal muscle hypertrophy is regulated via AKT/mTOR pathway Homo sapiens h igf1mtor pathway | 22/25 |
| Term | Overlap | Adjusted p-Value |
|---|---|---|
| E2F Targets | 180/200 | |
| G2-M Checkpoint | 176/200 | |
| Myc Targets V1 | 176/200 | |
| Mitotic Spindle | 172/199 | |
| mTORC1 Signaling | 166/200 | |
| Oxidative Phosphorylation | 163/200 | |
| Unfolded Protein Response | 98/113 | |
| UV Response Dn | 116/144 | |
| Adipogenesis | 148/200 |
| Term | Overlap | Adjusted p-Value |
|---|---|---|
| Direct p53 effectors Homo sapiens 67c3b75d-6191-11e5-8ac5-06603eb7f303 | 103/136 | |
| ErbB1 downstream signaling Homo sapiens 30d60550-6192-11e5-8ac5-06603eb7f303 | 83/105 | |
| PDGFR-beta signaling pathway Homo sapiens c901a3e4-6194-11e5-8ac5-06603eb7f303 | 95/128 | |
| Signaling events mediated by Hepatocyte Growth Factor Receptor (c-Met) Homo sapiens ac39d2b9-6195-11e5-8ac5-06603eb7f303 | 63/77 | |
| Validated targets of C-MYC transcriptional activation Homo sapiens 61d3b115-6196-11e5-8ac5-06603eb7f303 | 64/79 | |
| ATR signaling pathway Homo sapiens 8991cbac-618b-11e5-8ac5-06603eb7f303 | 37/39 | |
| Signaling events mediated by focal adhesion kinase Homo sapiens 8fb80085-6195-11e5-8ac5-06603eb7f303 | 50/58 | |
| p53 pathway Homo sapiens a0de862d-6194-11e5-8ac5-06603eb7f303 | 49/57 | |
| Aurora A signaling Homo sapiens f131cf8e-618b-11e5-8ac5-06603eb7f303 | 30/31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, K.; Taguchi, Y.-h. Gene Selection of Methionine-Dependent Melanoma and Independent Melanoma by Variable Selection Using Tensor Decomposition. Genes 2024, 15, 1543. https://doi.org/10.3390/genes15121543
Kobayashi K, Taguchi Y-h. Gene Selection of Methionine-Dependent Melanoma and Independent Melanoma by Variable Selection Using Tensor Decomposition. Genes. 2024; 15(12):1543. https://doi.org/10.3390/genes15121543
Chicago/Turabian StyleKobayashi, Kenta, and Y-h. Taguchi. 2024. "Gene Selection of Methionine-Dependent Melanoma and Independent Melanoma by Variable Selection Using Tensor Decomposition" Genes 15, no. 12: 1543. https://doi.org/10.3390/genes15121543
APA StyleKobayashi, K., & Taguchi, Y.-h. (2024). Gene Selection of Methionine-Dependent Melanoma and Independent Melanoma by Variable Selection Using Tensor Decomposition. Genes, 15(12), 1543. https://doi.org/10.3390/genes15121543