
Development of a Sensitive, Easy and High-Throughput Compliant Protocol for Maize and Soybean DNA Extraction and Quantitation Using a Plant-Specific Universal Taqman Minor Groove Binder Probe


Abstract
:1. Introduction
2. Materials and Methods
2.1. Maize and Soybean Samples
2.2. Reagents
- EBPT2 extraction buffer: 1% SDS, 1% PVP, 5% Tween 20, 0.5% Triton X-100, 50 mM EDTA, 100 mM NaCl, 100 mM Tris-HCl pH 8.
- RNase stock: Ribonuclease A at 20 mg/mL (Sigma Aldrich, St. Louis, MO, USA).
- Proteinase K stock: 20 mg/mL (Takara Bio Inc., Otsu, Japan).
- A total of 5 M potassium acetate stock [20].
- Binding Buffer BBU stock solution: 2.8 M guanidine-HCl, 65% ethanol.
- Wash Buffer CZ: 0.5 M guanidine-HCl, 40% ethanol.
- TE buffer [20].
- Genomic DNA Purification Trays (P/N 4318641, Invitrogen™, Carlsbad, CA, USA).
2.3. Instrumentations
- ABI PRISM® 6100 Nucleic Acid PrepStation (Thermo Fisher Scientific, Waltham, MA, USA).
- Refrigerated microcentrifuge.
- Thermomixer.
2.4. GUST2 DNA Extraction Protocol
- In a 1.5 mL microcentrifuge tube, mix 100 mg of flour with 660 µL of EBPT2 extraction buffer.
- Immediately add 10 µL of Rnase A and 10 µL of proteinase K and incubate at 65 °C with shaking (900 rpm) for 60 min (for example, in an Eppendorf Thermomixer Comfort).
- Add 260 µL of ice-cold 5 M potassium acetate, vortex for 30 s, and incubate in ice for 10 min.
- Centrifuge at 8000× g at 4 °C for 10 min.
- Transfer 260 µL of the supernatant into fresh 1.5 mL tubes and add 390 µL (1.5 volumes) of Binding Buffer BBU. Mix for at least 1 min.
- In the meanwhile, assemble the Genomic DNA Purification Tray onto the PrepStation and pre-wet the wells with 40 µL of Wash Buffer.
- Load the entire resulting volume from step 5 into the well and apply 20% vacuum for 90 s.
- First wash: buffer CZ, 600 µL, 20% vacuum for 90 s.
- Second wash: 60% ethanol, 600 µL, 20% vacuum for 90 s.
- Drying: 90% vacuum for 30 s (this removes residual ethanol).
- Add 100 µL of TE buffer.
- Incubate with 0% vacuum for 120 s.
- Elute using a 20% vacuum for 120 s.
2.5. TransPrep Extraction Protocol
2.6. NK603 CTAB Extraction Protocol
2.7. Primers, Probes, and Real-Time PCR
2.8. Construction of the 18S rRNA Standard Reference Plasmid (p18S)
3. Results
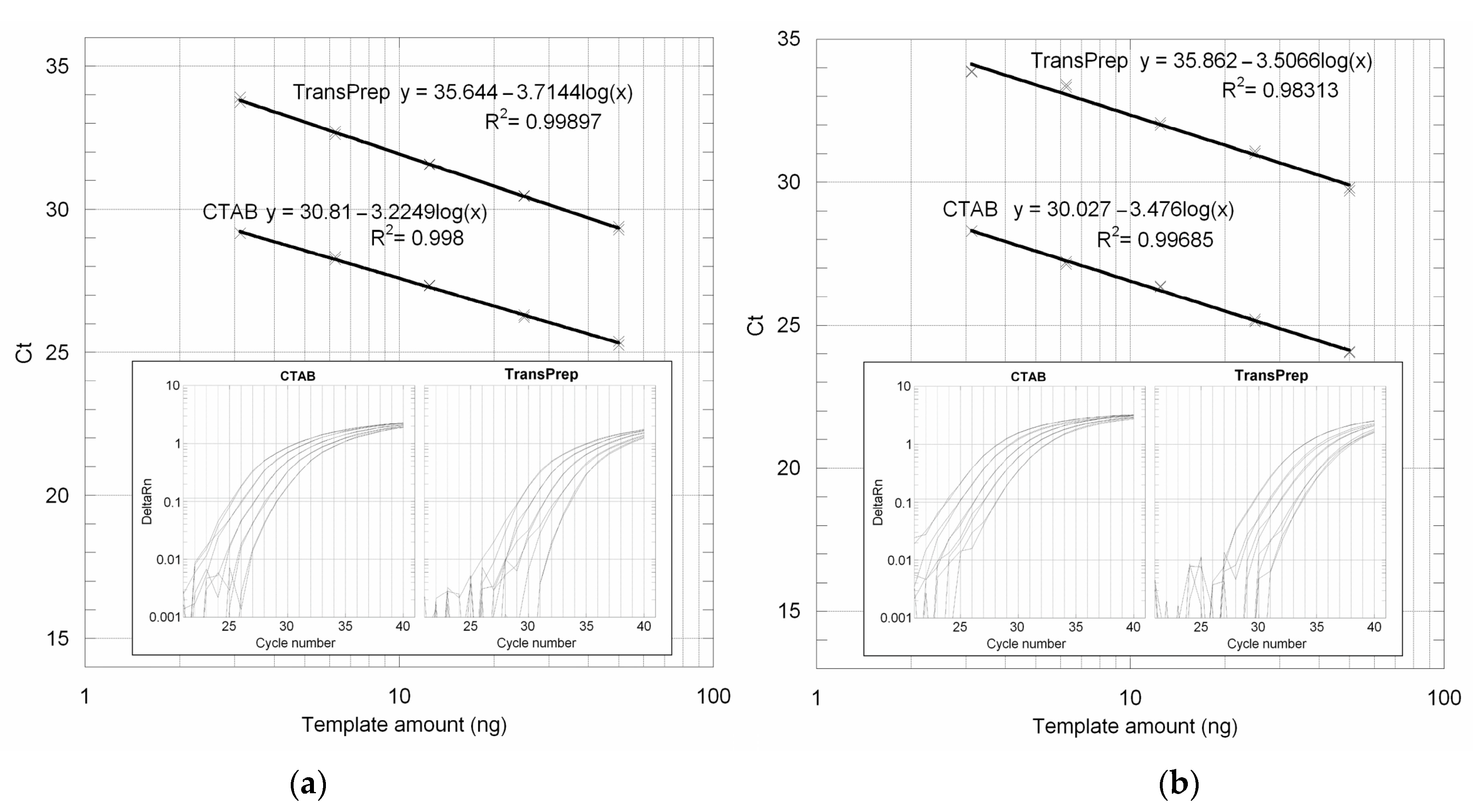
3.1. TransPrep DNA Extraction Protocol
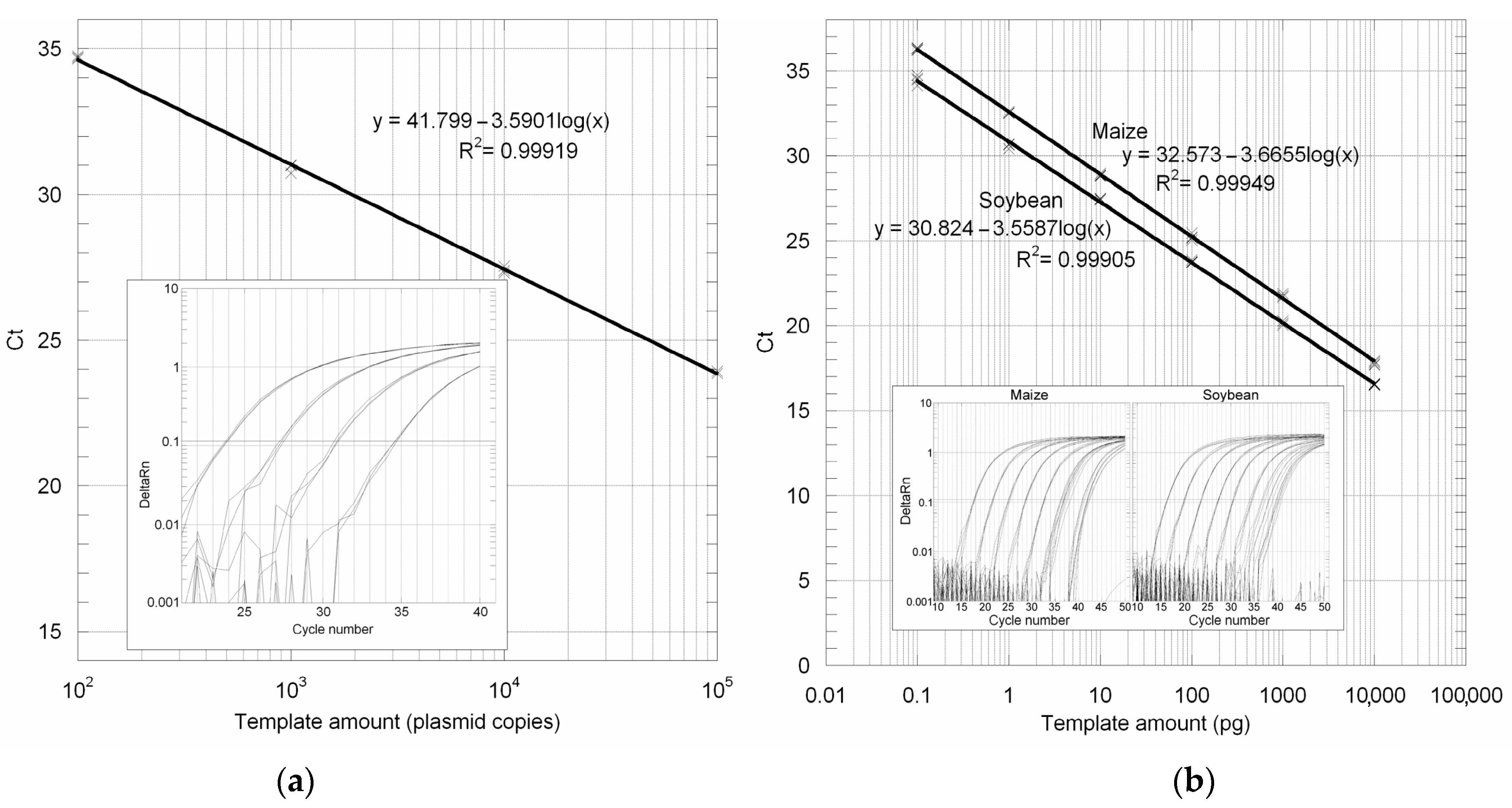
3.2. GUST2 DNA Extraction Protocol Evaluation Using the 18S rRNA Gene PCR Assay
3.3. Absolute LOD and LOQ Determination
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Al-Rabadi, G.J.; Tranchant, C.C.; Almajwal, A.; Kubow, S.; Alli, I. Occurrence, Types, Properties and Interactions of Phenolic Compounds with Other Food Constituents in Oil-Bearing Plants. Crit. Rev. Food Sci. Nutr. 2017, 58, 3209–3218. [Google Scholar] [CrossRef] [PubMed]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A Plant DNA Minipreparation: Version II. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR Inhibitors—Occurrence, Properties and Removal. J. Appl. Microbiol. 2012, 113, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.G.; Thompson, W.F. Rapid Isolation of High Molecular Weight Plant DNA. Nucleic Acids Res. 1980, 8, 4321. [Google Scholar] [CrossRef]
- Lipp, M.; Bluth, A.; Eyquem, F.; Kruse, L.; Schimmel, H.; Van Den Eede, G.; Anklam, E. Validation of a Method Based on Polymerase Chain Reaction for the Detection of Genetically Modified Organisms in Various Processed Foodstuffs. Eur. Food Res. Technol. 2001, 212, 497–504. [Google Scholar] [CrossRef]
- Sajali, N.; Wong, S.C.; Hanapi, U.K.; Jamaluddin, S.A.B.; Tasrip, N.A.; Mohd Desa, M.N. The Challenges of DNA Extraction in Different Assorted Food Matrices: A Review. J. Food. Sci. 2018, 83, 2409–2414. [Google Scholar] [CrossRef]
- Gryson, N. Effect of Food Processing on Plant DNA Degradation and PCR-Based GMO Analysis: A Review. Anal. Bioanal Chem. 2010, 396, 2003–2022. [Google Scholar] [CrossRef]
- Schenk, J.J.; Becklund, L.E.; Carey, S.J.; Fabre, P.P. What Is the “Modified” CTAB Protocol? Characterizing Modifications to the CTAB DNA Extraction Protocol. Appl. Plant Sci. 2023, 11, e11517. [Google Scholar] [CrossRef]
- Vogelstein, B.; Gillespie, D. Preparative and Analytical Purification of DNA from Agarose. Proc. Natl. Acad. Sci. USA 1979, 76, 615–619. [Google Scholar] [CrossRef]
- Porteous, L.A.; Armstrong, J.L. A Simple Mini-Method to Extract DNA Directly from Soil for Use with Polymerase Chain Reaction Amplification. Curr. Microbiol. 1993, 27, 115–118. [Google Scholar] [CrossRef]
- Alaeddini, R. Forensic Implications of PCR Inhibition—A Review. Forensic. Sci. Int. Genet. 2012, 6, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Elphinstone, M.S.; Hinten, G.N.; Anderson, M.J.; Nock, C.J. An Inexpensive and High-Throughput Procedure to Extract and Purify Total Genomic DNA for Population Studies. Mol. Ecol. Notes 2003, 3, 317–320. [Google Scholar] [CrossRef]
- Kikuchi, T.; Karim, N.; Masuya, H.; Ota, Y.; Kubono, T. An Inexpensive High-Throughput Method to Extract High Yields of Good Quality DNA from Fungi. Mol. Ecol. Resour. 2009, 9, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, N.V.; Dewaard, J.R.; Hebert, P.D.N. An Inexpensive, Automation-Friendly Protocol for Recovering High-Quality DNA. Mol. Ecol. Notes 2006, 6, 998–1002. [Google Scholar] [CrossRef]
- Schiebelhut, L.M.; Abboud, S.S.; Gómez Daglio, L.E.; Swift, H.F.; Dawson, M.N. A Comparison of DNA Extraction Methods for High-Throughput DNA Analyses. Mol. Ecol. Resour. 2017, 17, 721–729. [Google Scholar] [CrossRef]
- Vilanova, S.; Alonso, D.; Gramazio, P.; Plazas, M.; García-Fortea, E.; Ferrante, P.; Schmidt, M.; Díez, M.J.; Usadel, B.; Giuliano, G.; et al. SILEX: A Fast and Inexpensive High-Quality DNA Extraction Method Suitable for Multiple Sequencing Platforms and Recalcitrant Plant Species. Plant Methods 2020, 16, 110. [Google Scholar] [CrossRef]
- Applied Biosystems Application Note. Isolation, Detection, and Quantitation of Genetically Modified DNA in Food Materials. Available online: https://assets.thermofisher.com/TFS-Assets/LSG/Application-Notes/cms_040125.pdf (accessed on 25 July 2023).
- Paoletti, C.; Mazzara, M.; Puumalaainen, J.; Rasulo, D.; Van Den Eede, G. Event-Specific Method for the Quantitation of Maize Line GA21 Using Real-Time PCR: Validation Report and Protocol. Available online: http://gmo-crl.jrc.ec.europa.eu/gmomethods/docs/QT-EVE-ZM-007.pdf (accessed on 25 July 2023).
- Joint Research Centre—European Commission Biotechnology & GMOs Unit. Event-Specific Method for the Quantification of Maize Line NK603 Using Real-Time PCR. Protocol. Available online: https://gmo-crl.jrc.ec.europa.eu/summaries/NK603-WEB-ProtocolValidation.pdf (accessed on 2 September 2023).
- Sambrook, J.; Fritsch, E.R.; Maniatis, T. Molecular Cloning, a Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Kuribara, H.; Shindo, Y.; Matsuoka, T.; Takubo, K.; Futo, S.; Aoki, N.; Hirao, T.; Akiyama, H.; Goda, Y.; Toyoda, M.; et al. Novel Reference Molecules for Quantitation of Genetically Modified Maize and Soybean. J. AOAC Int. 2002, 85, 1077–1089. [Google Scholar] [CrossRef]
- Clustal, W. Multiple Sequence Alignment—Kyoto University Bioinformatics Center. Available online: https://www.genome.jp/tools-bin/clustalw (accessed on 26 July 2023).
- Leitch, I.J.; Johnston, E.; Pellicer, J.; Hidalgo, O.; Bennett, M.D. Plant DNA C-Values Database (Release 7.1, April 2019). Available online: https://cvalues.science.kew.org/ (accessed on 26 July 2023).
- Pikkart, M.J.; Villeponteau, B. Suppression of PCR Amplification by High Levels of RNA. Biotechniques 1993, 14, 24–25. [Google Scholar]
- Doyle, J.J.; Beachy, R.N. Ribosomal Gene Variation in Soybean (Glycine) and Its Relatives. Theor. Appl. Genet. 1985, 70, 369–376. [Google Scholar] [CrossRef]
- Varsanyi-Breiner, A.; Gusella, J.F.; Keys, C.; Housman, D.E.; Sullivan, D.; Brisson, N.; Verma, D.P.S. The Organization of a Nuclear DNA Sequence from a Higher Plant: Molecular Cloning and Characterization of Soybean Ribosomal DNA. Gene 1979, 7, 317–334. [Google Scholar] [CrossRef]
- Friedrich, H.; Hemleben, V.; Meagher, R.B.; Key, J.L. Purification and Restriction Endonuclease Mapping of Soybean 18 S and 25 S Ribosomal RNA Genes. Planta 1979, 146, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Buescher, P.J.; Phillips, R.L.; Brambl, R. Ribosomal RNA Contents of Maize Genotypes with Different Ribosomal RNA Gene Numbers. Biochem. Genet. 1984, 22, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, S.A.; Sinclair, J.H. Intraspecific Variation of Ribosomal Gene Redundancy in ZEA MAYS. Genetics 1975, 80, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Ingle, J.; Sinclair, J. Ribosomal RNA Genes and Plant Development. Nature 1972, 235, 30–32. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.O.; Bendich, A.J. Ribosomal RNA Genes in Plants: Variability in Copy Number and in the Intergenic Spacer. Plant Mol. Biol. 1987, 9, 509–520. [Google Scholar] [CrossRef]
- Hübner, P.; Waiblinger, H.; Pietsch, K.; Brodmann, P. Validation of PCR Methods for Quantitation of Genetically Modified Plants in Food. J. AOAC Int. 2001, 84, 1855–1864. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Target (Ref) | Name | Orientation | Sequence (5′-3′) |
|---|---|---|---|
| zSSIIb [21] | SSIIb 1-5′ SSIIb 1-3′ SSIIb-Taq | forward primer reverse primer TaqMan probe | CTC CCA ATC CTT TGA CAT CTG C TCG ATT TCT CTC TTG GTG ACA GG AGC AAA GTC AGA GCG CTG CAA TGC A |
| Le1 [21] | Le1n02-5′ Le1n02-3′ Le1-Taq | forward primer reverse primer TaqMan probe | GCC CTC TAC TCC ACC CCC A GCC CAT CTG CAA GCC TTT TT AGC TTC GCC GCT TCC TTC AAC TTC AC |
| 18S rDNA (this study) | uniPLA FW uniPLA RV uniPLA pro | forward primer reverse primer TaqMan MGB probe | CGG GGG CATT CGT ATT TCA TA GAC GGT ATC TGA TCG TCT TCG AG AGG ATG TTT TCA TTA ATC AAG AAC GA |
| Measured Values | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Calculated Values | 18S rRNA Gene Copies | |||||||||
| DNA | Genome Copies | Mean CT and SD | Total | per Haploid Genome | ||||||
| (pg) | Maize | Soybean | Maize | Soybean | Maize | Soybean | Maize | Soybean | ||
| 10,000 | 1835 | 4444 | 17.79 | 0.11 | 16.54 | 0.03 | 4,875,865 | 10,853,777 | 1329 | 1221 |
| 1000 | 183 | 444 | 21.80 | 0.10 | 20.15 | 0.10 | 372,780 | 1,073,526 | 1016 | 1208 |
| 100 | 18.3 | 44.4 | 25.20 | 0.17 | 23.76 | 0.07 | 41,897 | 106,086 | 1142 | 1193 |
| 10 | 1.83 | 4.44 | 28.86 | 0.08 | 27.45 | 0.04 | 4021 | 9899 | 1096 | 1114 |
| 1 | 0.18 | 0.44 | 32.52 | 0.05 | 30.54 | 0.36 | 384 | 1370 | 1048 | 1542 |
| 0.1 | 0.018 | 0.044 | 36.28 | 0.06 | 34.48 | 0.26 | 34 | 109 | 937 | 1231 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ambra, R.; Marcelli, M.; D’Orso, F. Development of a Sensitive, Easy and High-Throughput Compliant Protocol for Maize and Soybean DNA Extraction and Quantitation Using a Plant-Specific Universal Taqman Minor Groove Binder Probe. Genes 2023, 14, 1797. https://doi.org/10.3390/genes14091797
Ambra R, Marcelli M, D’Orso F. Development of a Sensitive, Easy and High-Throughput Compliant Protocol for Maize and Soybean DNA Extraction and Quantitation Using a Plant-Specific Universal Taqman Minor Groove Binder Probe. Genes. 2023; 14(9):1797. https://doi.org/10.3390/genes14091797
Chicago/Turabian StyleAmbra, Roberto, Marco Marcelli, and Fabio D’Orso. 2023. "Development of a Sensitive, Easy and High-Throughput Compliant Protocol for Maize and Soybean DNA Extraction and Quantitation Using a Plant-Specific Universal Taqman Minor Groove Binder Probe" Genes 14, no. 9: 1797. https://doi.org/10.3390/genes14091797
APA StyleAmbra, R., Marcelli, M., & D’Orso, F. (2023). Development of a Sensitive, Easy and High-Throughput Compliant Protocol for Maize and Soybean DNA Extraction and Quantitation Using a Plant-Specific Universal Taqman Minor Groove Binder Probe. Genes, 14(9), 1797. https://doi.org/10.3390/genes14091797