
Cloning and Functional Verification of Endogenous U6 Promoters for the Establishment of Efficient CRISPR/Cas9-Based Genome Editing in Castor (Ricinus communis)

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Plasmid Construction
2.3. Transient Expression with the Particle Delivery System
2.4. Confocal Microscopy
2.5. Nucleotide Acession Numbers
3. Results and Discussion
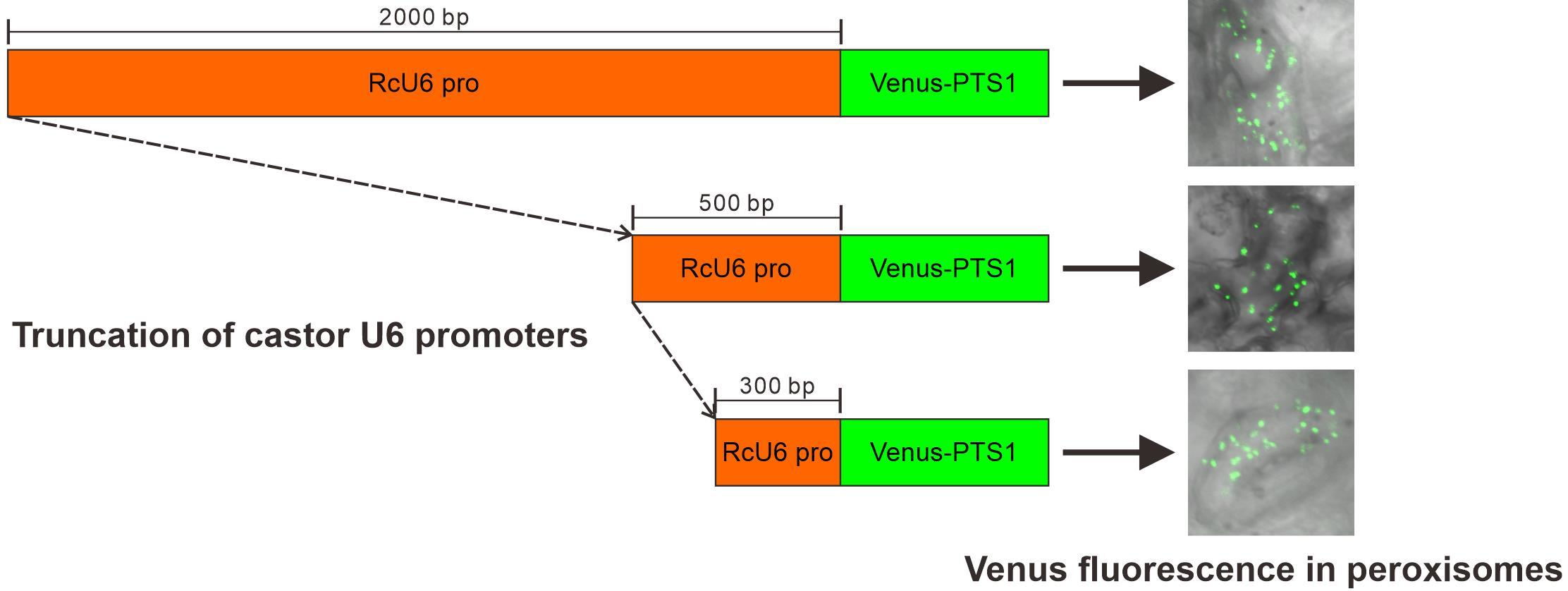
3.1. Castor U6 snRNA Gene Identification and Promoter Cloning
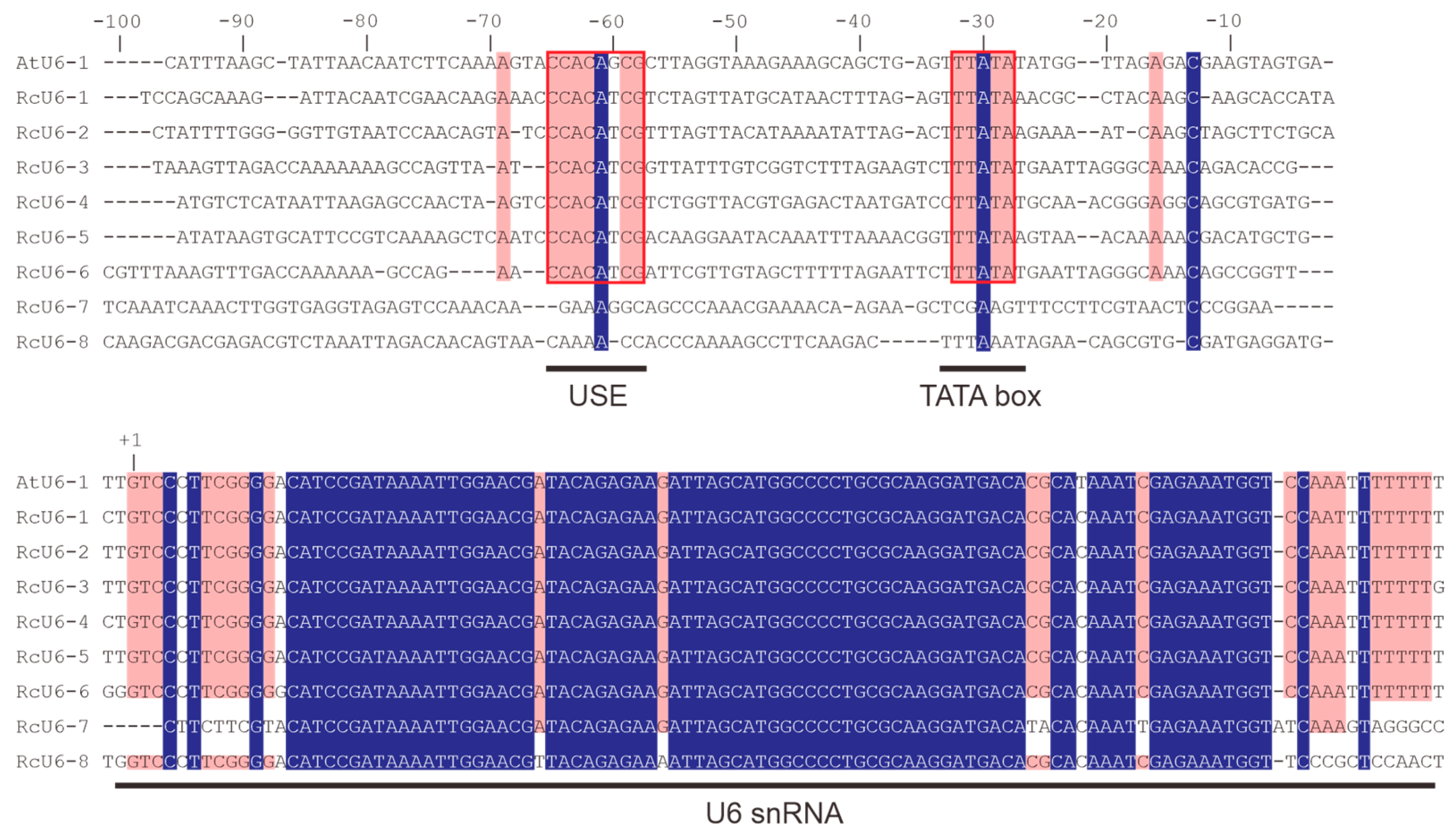
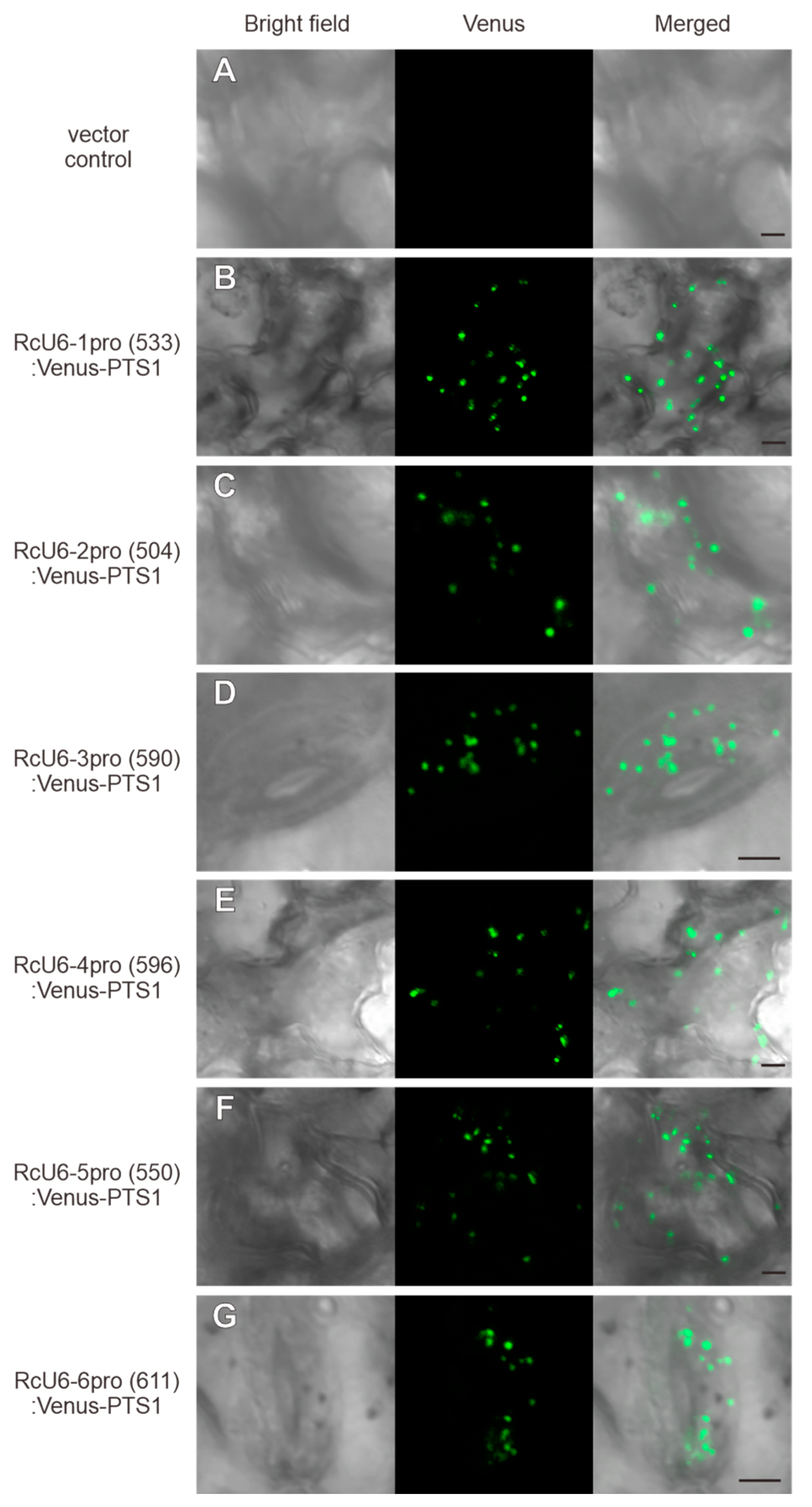
3.2. Functional Analysis of the Rc U6 Gene Promoters
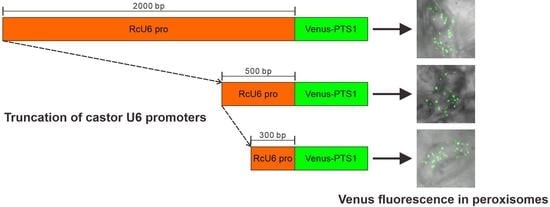
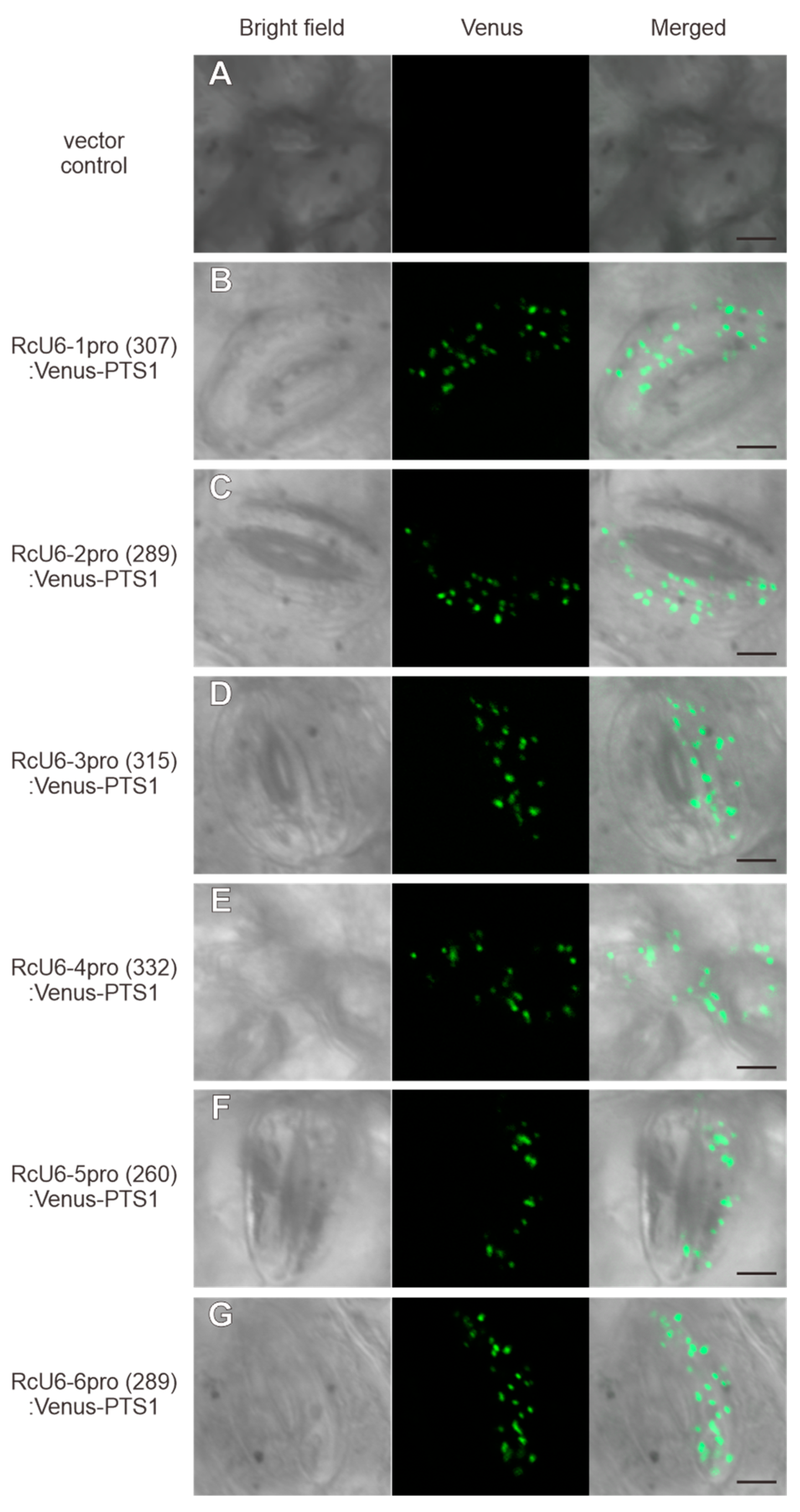
3.3. Truncation of Rc U6 Promoters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patel, V.R.; Dumancas, G.G.; Kasi Viswanath, L.C.; Maples, R.; Subong, B.J.J. Castor Oil: Properties, Uses, and Optimization of Processing Parameters in Commercial Production. Lipid Insights 2016, 9, 1–12. [Google Scholar] [CrossRef]
- Nitbani, F.O.; Tjitda, P.J.P.; Wogo, H.E.; Detha, A.I.R. Preparation of Ricinoleic Acid from Castor Oil: A Review. J. Oleo Sci. 2022, 71, 781–793. [Google Scholar] [CrossRef]
- Kunduru, K.R.; Basu, A.; Haim Zada, M.; Domb, A.J. Castor Oil-Based Biodegradable Polyesters. Biomacromolecules 2015, 16, 2572–2587. [Google Scholar] [CrossRef]
- Chakraborty, I.; Chatterjee, K. Polymers and Composites Derived from Castor Oil as Sustainable Materials and Degradable Biomaterials: Current Status and Emerging Trends. Biomacromolecules 2020, 21, 4639–4662. [Google Scholar] [CrossRef]
- Timko, J.A.; Amsalu, A.; Acheampong, E.; Teferi, M.K. Local Perceptions about the Effects of Jatropha (Jatropha curcas) and Castor (Ricinus communis) Plantations on Households in Ghana and Ethiopia. Sustain. Sci. Pract. Policy 2014, 6, 7224–7241. [Google Scholar] [CrossRef]
- Xu, W.; Chen, Z.; Ahmed, N.; Han, B.; Cui, Q.; Liu, A. Genome-Wide Identification, Evolutionary Analysis, and Stress Responses of the GRAS Gene Family in Castor Beans. Int. J. Mol. Sci. 2016, 17, 1004. [Google Scholar] [CrossRef]
- Han, B.; Fu, L.; Zhang, D.; He, X.; Chen, Q.; Peng, M.; Zhang, J. Interspecies and Intraspecies Analysis of Trehalose Contents and the Biosynthesis Pathway Gene Family Reveals Crucial Roles of Trehalose in Osmotic-Stress Tolerance in Cassava. Int. J. Mol. Sci. 2016, 17, 1077. [Google Scholar] [CrossRef]
- Polito, L.; Bortolotti, M.; Battelli, M.G.; Calafato, G.; Bolognesi, A. Ricin: An Ancient Story for a Timeless Plant Toxin. Toxins 2019, 11, 324. [Google Scholar] [CrossRef]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu. Rev. Plant Biol. 2019, 70, 667–697. [Google Scholar] [CrossRef]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G.; et al. Draft Genome Sequence of the Oilseed Species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef]
- Lu, J.; Pan, C.; Fan, W.; Liu, W.; Zhao, H.; Li, D.; Wang, S.; Hu, L.; He, B.; Qian, K.; et al. A Chromosome-Level Genome Assembly of Wild Castor Provides New Insights into Its Adaptive Evolution in Tropical Desert. Genom. Proteom. Bioinform. 2022, 20, 42–59. [Google Scholar] [CrossRef]
- Alexandrov, O.S.; Petrov, N.R.; Varlamova, N.V.; Khaliluev, M.R. An Optimized Protocol for In Vitro Indirect Shoot Organogenesis of Impala Bronzovaya and Zanzibar Green Ricinus communis L. Varieties. Horticulturae 2021, 7, 105. [Google Scholar] [CrossRef]
- Cable, J.; Ronald, P.C.; Voytas, D.; Zhang, F.; Levy, A.A.; Takatsuka, A.; Arimura, S.-I.; Jacobsen, S.E.; Toki, S.; Toda, E.; et al. Plant Genome Engineering from Lab to Field-a Keystone Symposia Report. Ann. N. Y. Acad. Sci. 2021, 1506, 35–54. [Google Scholar] [CrossRef] [PubMed]
- Kor, S.D.; Chowdhury, N.; Keot, A.K.; Yogendra, K.; Chikkaputtaiah, C.; Sudhakar Reddy, P. RNA Pol III Promoters-Key Players in Precisely Targeted Plant Genome Editing. Front. Genet. 2022, 13, 989199. [Google Scholar] [CrossRef]
- Long, L.; Guo, D.-D.; Gao, W.; Yang, W.-W.; Hou, L.-P.; Ma, X.-N.; Miao, Y.-C.; Botella, J.R.; Song, C.-P. Optimization of CRISPR/Cas9 Genome Editing in Cotton by Improved SgRNA Expression. Plant Methods 2018, 14, 85. [Google Scholar] [CrossRef] [PubMed]
- Massel, K.; Lam, Y.; Hintzsche, J.; Lester, N.; Botella, J.R.; Godwin, I.D. Endogenous U6 Promoters Improve CRISPR/Cas9 Editing Efficiencies in Sorghum Bicolor and Show Potential for Applications in Other Cereals. Plant Cell Rep. 2022, 41, 489–492. [Google Scholar] [CrossRef]
- Johansen, I.E.; Liu, Y.; Jørgensen, B.; Bennett, E.P.; Andreasson, E.; Nielsen, K.L.; Blennow, A.; Petersen, B.L. High Efficacy Full Allelic CRISPR/Cas9 Gene Editing in Tetraploid Potato. Sci. Rep. 2019, 9, 17715. [Google Scholar] [CrossRef]
- Riu, Y.-S.; Kim, G.H.; Chung, K.W.; Kong, S.-G. Enhancement of the CRISPR/Cas9-Based Genome Editing System in Lettuce (Lactuca sativa L.) Using the Endogenous U6 Promoter. Plants 2023, 12, 878. [Google Scholar] [CrossRef]
- Nakagawa, T.; Nakamura, S.; Tanaka, K.; Kawamukai, M.; Suzuki, T.; Nakamura, K.; Kimura, T.; Ishiguro, S. Development of R4 Gateway Binary Vectors (R4pGWB) Enabling High-Throughput Promoter Swapping for Plant Research. Biosci. Biotechnol. Biochem. 2008, 72, 624–629. [Google Scholar] [CrossRef]
- Kanai, M.; Hayashi, M.; Kondo, M.; Nishimura, M. The Plastidic DEAD-Box RNA Helicase 22, HS3, Is Essential for Plastid Functions Both in Seed Development and in Seedling Growth. Plant Cell Physiol. 2013, 54, 1431–1440. [Google Scholar] [CrossRef]
- Didychuk, A.L.; Butcher, S.E.; Brow, D.A. The Life of U6 Small Nuclear RNA, from Cradle to Grave. RNA 2018, 24, 437–460. [Google Scholar] [CrossRef] [PubMed]
- Giles, K.E.; Caputi, M.; Beemon, K.L. Packaging and Reverse Transcription of SnRNAs by Retroviruses May Generate Pseudogenes. RNA 2004, 10, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Guerineau, F.; Waugh, R. The U6 Small Nuclear RNA Gene Family of Potato. Plant Mol. Biol. 1993, 22, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Wilson, F.M.; Harrison, K.; Armitage, A.D.; Simkin, A.J.; Harrison, R.J. CRISPR/Cas9-Mediated Mutagenesis of Phytoene Desaturase in Diploid and Octoploid Strawberry. Plant Methods 2019, 15, 45. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, G.; Liu, Z. Efficient Genome Editing of Wild Strawberry Genes, Vector Development and Validation. Plant Biotechnol. J. 2018, 16, 1868–1877. [Google Scholar] [CrossRef]
- Lorence, A.; Verpoorte, R. Gene Transfer and Expression in Plants. Methods Mol. Biol. 2004, 267, 329–350. [Google Scholar]
- Taylor, N.J.; Fauquet, C.M. Microparticle Bombardment as a Tool in Plant Science and Agricultural Biotechnology. DNA Cell Biol. 2002, 21, 963–977. [Google Scholar] [CrossRef]
- Gunnery, S.; Mathews, M.B. Functional mRNA Can Be Generated by RNA Polymerase III. Mol. Cell. Biol. 1995, 15, 3597–3607. [Google Scholar] [CrossRef]
- Hayashi, M.; Aoki, M.; Kato, A.; Kondo, M.; Nishimura, M. Transport of Chimeric Proteins That Contain a Carboxy-Terminal Targeting Signal into Plant Microbodies. Plant J. 1996, 10, 225–234. [Google Scholar] [CrossRef]
- Nagai, T.; Ibata, K.; Park, E.S.; Kubota, M.; Mikoshiba, K.; Miyawaki, A. A Variant of Yellow Fluorescent Protein with Fast and Efficient Maturation for Cell-Biological Applications. Nat. Biotechnol. 2002, 20, 87–90. [Google Scholar] [CrossRef]
- Nakamura, S.; Mano, S.; Tanaka, Y.; Ohnishi, M.; Nakamori, C.; Araki, M.; Niwa, T.; Nishimura, M.; Kaminaka, H.; Nakagawa, T.; et al. Gateway Binary Vectors with the Bialaphos Resistance Gene, bar, as a Selection Marker for Plant Transformation. Biosci. Biotechnol. Biochem. 2010, 74, 1315–1319. [Google Scholar] [CrossRef] [PubMed]
- Manghwar, H.; Lindsey, K.; Zhang, X.; Jin, S. CRISPR/Cas System: Recent Advances and Future Prospects for Genome Editing. Trends Plant Sci. 2019, 24, 1102–1125. [Google Scholar] [CrossRef] [PubMed]
- Bernard, G.; Gagneul, D.; Alves Dos Santos, H.; Etienne, A.; Hilbert, J.-L.; Rambaud, C. Efficient Genome Editing Using CRISPR/Cas9 Technology in Chicory. Int. J. Mol. Sci. 2019, 20, 1155. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Chromosome No. | Region (bp) | Sequence Identity |
|---|---|---|---|
| RcU6-1 | 8 | 15,427,322–15,427,419 | 96/98 (98%) |
| RcU6-2 | 8 | 16,221,161–16,221,064 | 97/98 (99%) |
| RcU6-3 | 6 | 19,921,764–19,921,668 | 96/97 (99%) |
| RcU6-4 | 6 | 21,889,600–21,889,503 | 97/98 (99%) |
| RcU6-5 | 1 | 35,147,644–35,147,547 | 97/98 (99%) |
| RcU6-6 | 10 | 15,041,917–15,041,820 | 96/98 (98%) |
| RcU6-7 | 1 | 32,438,768–32,438,688 | 77/81 (95%) |
| RcU6-8 | 7 | 28,298,413–28,298,329 | 82/85 (95%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanai, M.; Hikino, K.; Mano, S. Cloning and Functional Verification of Endogenous U6 Promoters for the Establishment of Efficient CRISPR/Cas9-Based Genome Editing in Castor (Ricinus communis). Genes 2023, 14, 1327. https://doi.org/10.3390/genes14071327
Kanai M, Hikino K, Mano S. Cloning and Functional Verification of Endogenous U6 Promoters for the Establishment of Efficient CRISPR/Cas9-Based Genome Editing in Castor (Ricinus communis). Genes. 2023; 14(7):1327. https://doi.org/10.3390/genes14071327
Chicago/Turabian StyleKanai, Masatake, Kazumi Hikino, and Shoji Mano. 2023. "Cloning and Functional Verification of Endogenous U6 Promoters for the Establishment of Efficient CRISPR/Cas9-Based Genome Editing in Castor (Ricinus communis)" Genes 14, no. 7: 1327. https://doi.org/10.3390/genes14071327
APA StyleKanai, M., Hikino, K., & Mano, S. (2023). Cloning and Functional Verification of Endogenous U6 Promoters for the Establishment of Efficient CRISPR/Cas9-Based Genome Editing in Castor (Ricinus communis). Genes, 14(7), 1327. https://doi.org/10.3390/genes14071327