

Occurrence of Sex Chromosomes in Fish of the Genus Ancistrus with a New Description of Multiple Sex Chromosomes in the Ecuadorian Endemic Ancistrus clementinae (Loricariidae)
 , , ,
, , ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Morphological Identification
2.2. Molecular Identification of Samples and Phylogenetic Reconstruction
2.3. Cytogenetic Procedures
2.4. Images Capture and Processing
3. Results
3.1. Molecular Identification of Samples and Phylogenetic Reconstruction
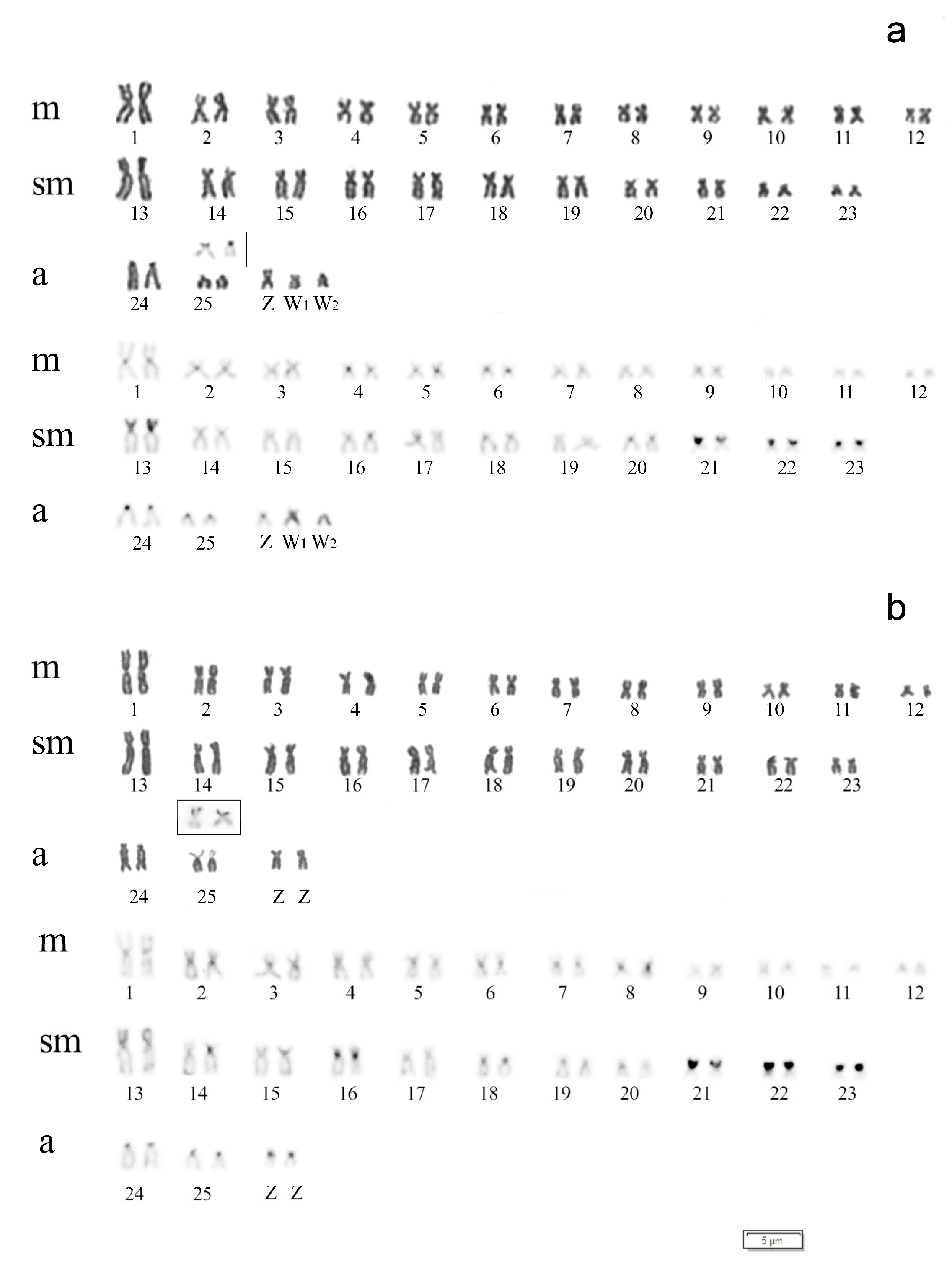
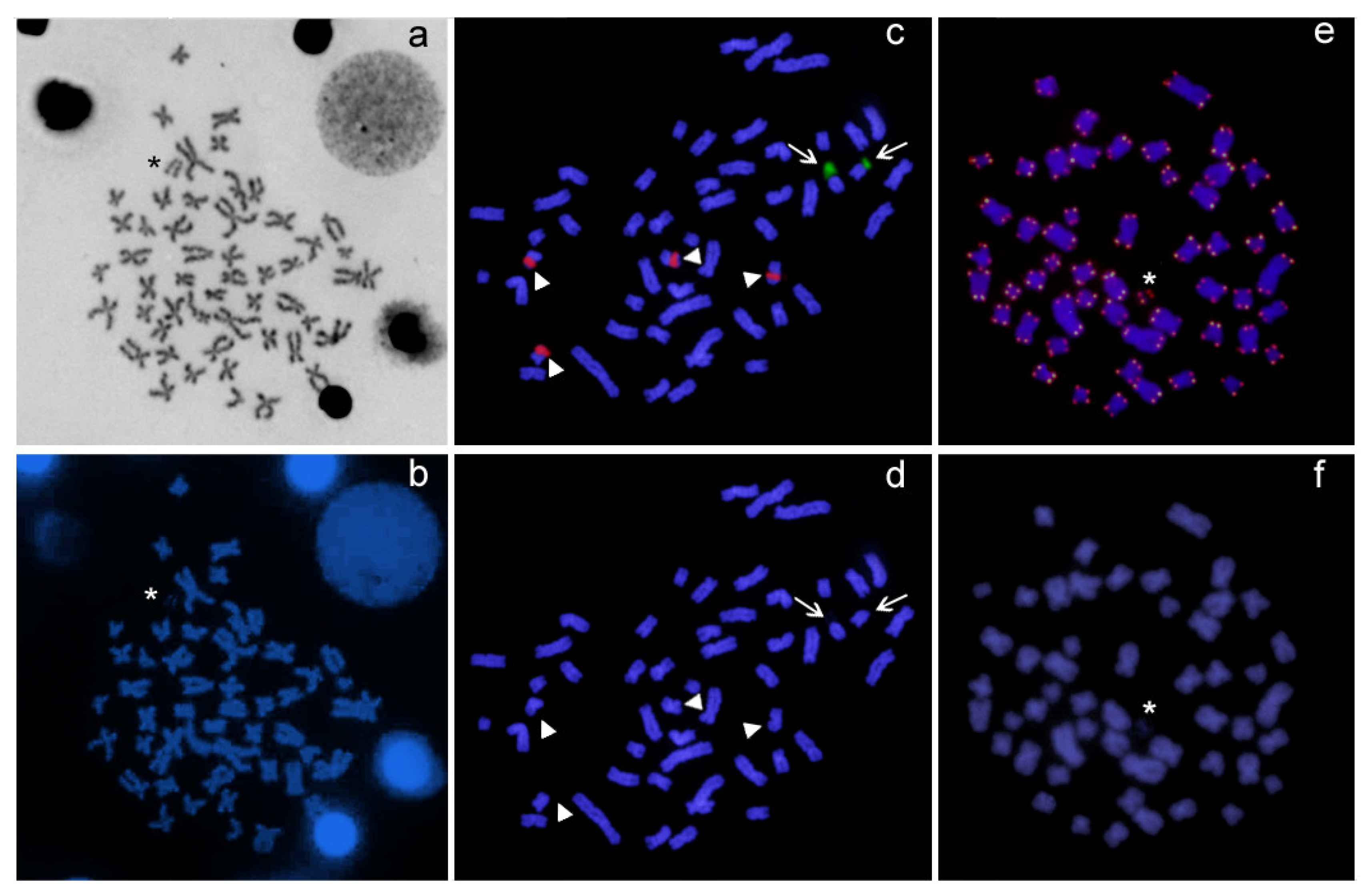
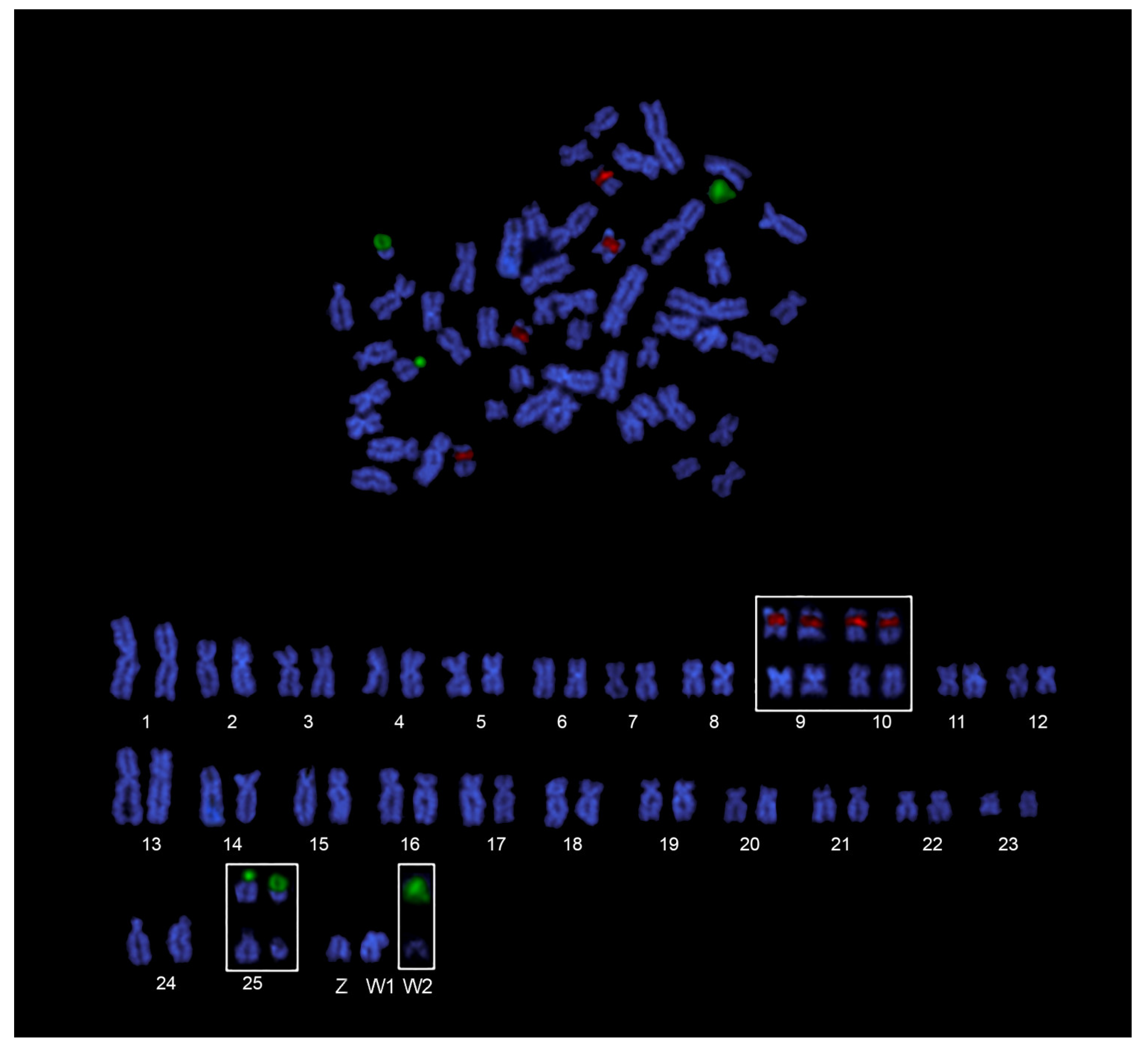
3.2. Cytogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Eschmeyer Catalog of Fishes: Genera/Species by Family/Subfamily. Available online: https://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 30 November 2022).
- Artoni, R.F.; Bertollo, L.A. Trends in the Karyotype Evolution of Loricariidae Fish (Siluriformes). Hereditas 2001, 134, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Ferraris, C.J. Checklist of Catfishes, Recent and Fossil (Osteichthyes: Siluriformes), and Catalogue of Siluriform Primary Types. Zootaxa 2007, 1418, 1–628. [Google Scholar] [CrossRef]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016; 752p. [Google Scholar]
- Roxo, F.F.; Ochoa, L.E.; Sabaj, M.H.; Lujan, N.K.; Covain, R.; Silva, G.S.C.; Melo, B.F.; Albert, J.S.; Chang, J.; Foresti, F.; et al. Phylogenomic Reappraisal of the Neotropical Catfish Family Loricariidae (Teleostei: Siluriformes) Using Ultraconserved Elements. Mol. Phylogenet. Evol. 2019, 135, 148–165. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, J.W. Phylogenetic Relationships of the Suckermouth Armoured Catfishes (Loricariidae) with Emphasis on the Hypostominae and the Ancistrinae. Zool. J. Linn. Soc. 2004, 141, 1–80. [Google Scholar] [CrossRef]
- De Oliveira, R.R.; Feldberg, E.; Dos Anjos, M.B.; Zuanon, J. Mechanisms of Chromosomal Evolution and Its Possible Relation to Natural History Characteristics in Ancistrus Catfishes (Siluriformes: Loricariidae). J. Fish Biol. 2009, 75, 2209–2225. [Google Scholar] [CrossRef]
- Lujan, N.K.; Armbruster, J.W.; Lovejoy, N.R.; López-Fernández, H. Multilocus Molecular Phylogeny of the Suckermouth Armored Catfishes (Siluriformes: Loricariidae) with a Focus on Subfamily Hypostominae. Mol. Phylogenet. Evol. 2015, 82 Pt A, 269–288. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. Family Loricariidae—Armored Catfishes. Available online: https://www.fishbase.se/Nomenclature/ValidNameList.php?syng=Ancistrus&syns=&vtitle=Scientific+Names+where+Genus+Equals+%3Ci%3EAncistrus%3C%2Fi%3E&crit2=CONTAINS&crit1=EQUAL (accessed on 23 December 2022).
- Taphorn, D.C.; Armbruster, J.W.; Rodríguez-Olarte, D. Ancistrus falconensis N. Sp. and A. gymnorhynchus Kner (Siluriformes: Loricariidae) from Central Venezuelan Caribbean Coastal Streams. Zootaxa 2010, 2345, 19–32. [Google Scholar] [CrossRef]
- Taphorn, D.C.; Armbruster, J.W.; Villa-Navarro, F.; Ray, C.K. Trans-Andean Ancistrus (Siluriformes: Loricariidae). Zootaxa 2013, 3641, 343–370. [Google Scholar] [CrossRef]
- Provenzano, R.F.; Barriga-Salazar, R. Species of Ancistrus (Siluriformes, Loricariidae) from Ecuador, with the Description of a New Species from the Amazon River Basin. Zootaxa 2018, 4527, 211–238. [Google Scholar] [CrossRef]
- Jiménez Prado, P. Guía de Peces Para Aguas Continentales En La Vertiente Occidental Del Ecuador; Pontificia Universidad Católica del Ecuador Sede Esmeraldas (PUCESE), Universidad del Azuay (UDA) and Museo Ecuatoriano de Ciencias Naturales (MECN) del Instituto Nacional de Biodiversidad: Esmeraldas, Ecuador, 2015; 420p. [Google Scholar]
- Dignall, J.G. Ancistrus clementinae Loricariidae Cat-eLog. Available online: https://www.planetcatfish.com/common/species.php?species_id=1941 (accessed on 4 November 2022).
- Rossi, A.R. Fish Cytogenetics: Present and Future. Genes 2021, 12, 983. [Google Scholar] [CrossRef]
- Arai, R. Fish Karyotypes: A Check List; Springer: Tokyo, Japan, 2011; 348p. [Google Scholar]
- Glugoski, L.; Deon, G.; Schott, S.; Vicari, M.R.; Nogaroto, V.; Moreira-Filho, O. Comparative Cytogenetic Analyses in Ancistrus Species (Siluriformes: Loricariidae). Neotrop. Ichthyol. 2020, 18, e200013. [Google Scholar] [CrossRef]
- Mariotto, S.; Miyazawa, C.S. Ancistrus cf. dubius (Siluriformes, Ancistrinae), a Complex of Species. 1. Chromosomic Characterization of Four Populations and Occurrence of Sexual Chromosomes of Type XX/XY, in the Pantanal Basin of Mato Grosso, Brazil. Caryologia 2006, 59, 299–304. [Google Scholar] [CrossRef]
- Mariotto, S.; Artoni, F.R.; Miyazawa, C.S. Occurrence of Sexual Chromosome, of the Type ZZ/ZW, in Ancistrus cf. dubius (Loricariidae, Ancistrinae) of the Paraguay River Basin, Mato Grosso, Brazil. Caryologia 2004, 57, 327–331. [Google Scholar] [CrossRef]
- Mariotto, S.; Centofante, L.; Miyazawa, C.S.; Bertollo, L.A.C.; Moreira Filho, O. Chromosome Polymorphism in Ancistrus cuiabae Knaack, 1999 (Siluriformes: Loricariidae: Ancistrini). Neotrop. Ichthyol. 2009, 7, 595–600. [Google Scholar] [CrossRef]
- de Oliveira, R.R.; Feldberg, E.; dos Anjos, M.B.; Zuanon, J. Occurrence of Multiple Sexual Chromosomes (XX/XY1Y2 and Z1Z1Z2Z2/Z1Z2W1W2) in Catfishes of the Genus Ancistrus (Siluriformes: Loricariidae) from the Amazon Basin. Genetica 2008, 134, 243–249. [Google Scholar] [CrossRef]
- Santos da Silva, K.; Glugoski, L.; Vicari, M.R.; de Souza, A.C.P.; Noronha, R.C.R.; Pieczarka, J.C.; Nagamachi, C.Y. Chromosomal Diversification in Ancistrus Species (Siluriformes: Loricariidae) Inferred from Repetitive Sequence Analysis. Front. Genet. 2022, 13, 838462. [Google Scholar] [CrossRef]
- Mariotto, S.; Centofante, L.; Vicari, M.R.; Artoni, R.F.; Moreira-Filho, O. Chromosomal Diversification in Ribosomal DNA Sites in Ancistrus Kner, 1854 (Loricariidae, Ancistrini) from Three Hydrographic Basins of Mato Grosso, Brazil. Comp. Cytogenet. 2011, 5, 289–300. [Google Scholar] [CrossRef]
- Mariotto, S.; Centofante, L.; Moreira-Filho, O. Diversity and Chromosomal Evolution in the genus Ancistrus Kner, 1854 (Loricariidae: Ancistrini) from Three Hydrographic Basins of Mato Grosso State, Brazil. Neotrop. Ichthyol. 2013, 11, 125–131. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA Barcoding Australia’s Fish Species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Milana, V.; Fusari, A.; Rossi, A.; Sola, L. Molecular and Morphological Identification of an Uncommon Centrolophid Fish. Cent. Eur. J. Biol. 2011, 6, 360. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple Sequence Alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 2, Unit 2.3. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Lozano, R.; Ruiz Rejon, C.; Ruiz Rejon, M. A Method for Increasing the Number of Mitoses Available for Cytogenetic Analysis in Rainbow Trout. Stain Technol. 1988, 63, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Leary, S.L.; Underwood, W.; Anthony, R.; Cartner, S.; Grandin, T.; Greenacre, C.; Gwaltney-Brant, S.; McCracking, M.A.; Meyer, R.; Miller, D.; et al. AVMA Guidelines for the Euthanasia of Animals, 2020th ed.; American Veterinary Medical Association: Schaumburg, IL, USA, 2020. [Google Scholar]
- Nirchio, M.; Oliveira, C. Citogenética de Peces; Universidad de Oriente: Cumaná, Venezuela, 2006; 212p. [Google Scholar]
- Sumner, A.T. A Simple Technique for Demonstrating Centromeric Heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef] [PubMed]
- Howell, W.M.; Black, D.A. Controlled Silver-Staining of Nucleolus Organizer Regions with a Protective Colloidal Developer: A 1-Step Method. Experientia 1980, 36, 1014–1015. [Google Scholar] [CrossRef]
- Pinkel, D.; Straume, T.; Gray, J.W. Cytogenetic analysis using quantitative, high-sensitivity, fluorescence hybridization. Proc. Natl. Acad. Sci. USA 1986, 83, 2934–2938. [Google Scholar] [CrossRef]
- Soares, L.B.; Paim, F.G.; Ramos, L.P.; Foresti, F.; Oliveira, C. Molecular Cytogenetic Analysis and the Establishment of a Cell Culture in the Fish Species Hollandichthys multifasciatus (Eigenmann & Norris, 1900)(Characiformes, Characidae). Genet. Mol. Biol. 2021, 44, e20200260. [Google Scholar] [CrossRef]
- Sassi, F.M.C.; Toma, G.A.; Cioffi, M.B. FISH-in Fish Chromosomes. In Cytogenetics and Molecular Cytogenetics; Liehr, T., Ed.; CRC Press: Boca Raton, FL, USA, 2023; pp. 281–296. [Google Scholar] [CrossRef]
- Utsunomia, R.; de Silva, D.M.Z.A.; Ruiz-Ruano, F.J.; Araya-Jaime, C.; Pansonato-Alves, J.C.; Scacchetti, P.C.; Hashimoto, D.T.; Oliveira, C.; Trifonov, V.A.; Porto-Foresti, F.; et al. Uncovering the Ancestry of B Chromosomes in Moenkhausia sanctaefilomenae (Teleostei, Characidae). PLoS ONE 2016, 11, e0150573. [Google Scholar] [CrossRef]
- Pendas, A.M.; Moran, P.; Martinez, J.L.; Garcia-Vazquez, E. Applications of 5S rDNA in Atlantic Salmon, Brown Trout, and in Atlantic Salmon Brown Trout Hybrid Identification. Mol. Ecol. 1995, 4, 275–276. [Google Scholar] [CrossRef]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved Telomere Detection Using a Telomere Repeat Probe (TTAGGG)n Generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for Centromeric Position on Chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Favarato, R.M.; da Silva, M.; de Oliveira, R.R.; Artoni, R.F.; Feldberg, E.; Matoso, D.A. Cytogenetic Diversity and the Evolutionary Dynamics of rDNA Genes and Telomeric Sequences in the Ancistrus Genus (Loricariidae: Ancistrini). Zebrafish 2016, 13, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Bueno, V.; Konerat, J.T.; Zawadzki, C.H.; Venere, P.C.; Blanco, D.R.; Margarido, V.P. Divergent Chromosome Evolution in Hypostominae Tribes (Siluriformes: Loricariidae): Correlation of Chromosomal Data with Morphological and Molecular Phylogenies. Zebrafish 2018, 15, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Sember, A.; Nguyen, P.; Perez, M.F.; Altmanová, M.; Ráb, P.; de Bello Cioffi, M. Multiple Sex Chromosomes in Teleost Fishes from a Cytogenetic Perspective: State of the Art and Future Challenges. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20200098. [Google Scholar] [CrossRef]
- de Cioffi, M.B.; Yano, C.F.; Sember, A.; Bertollo, L.A.C. Chromosomal Evolution in Lower Vertebrates: Sex Chromosomes in Neotropical Fishes. Genes 2017, 8, 258. [Google Scholar] [CrossRef]
- Sassi, F.M.C.; Deon, G.A.; Moreira-filho, O.; Vicari, M.R.; Bertollo, L.A.C.; Liehr, T.; de Oliveira, E.A.; Cioffi, M.B. Multiple Sex Chromosomes and Evolutionary Relationships in Amazonian Catfishes: The Outstanding Model of the Genus Harttia (Siluriformes: Loricariidae). Genes 2020, 11, 1179. [Google Scholar] [CrossRef]
- Chalopin, D.; Volff, J.-N.; Galiana, D.; Anderson, J.L.; Schartl, M. Transposable Elements and Early Evolution of Sex Chromosomes in Fish. Chromosome Res. 2015, 23, 545–560. [Google Scholar] [CrossRef]
- de Cioffi, M.B.; Kejnovský, E.; Marquioni, V.; Poltronieri, J.; Molina, W.F.; Diniz, D.; Bertollo, L.A.C. The Key Role of Repeated DNAs in Sex Chromosome Evolution in Two Fish Species with ZW Sex Chromosome System. Mol. Cytogenet. 2012, 5, 28. [Google Scholar] [CrossRef]
- Marajó, L.; Viana, P.F.; Ferreira, A.M.V.; Py-Daniel, L.H.R.; de Cioffi, M.B.; Sember, A.; Feldberg, E. Chromosomal Rearrangements and the First Indication of an ♀X1X1X2X2 /♂X1X2Y Sex Chromosome System in Rineloricaria Fishes (Teleostei: Siluriformes). J. Fish Biol. 2022, 1–12. [Google Scholar] [CrossRef]
- Blanco, D.R.; Vicari, M.R.; Lui, R.L.; Bertollo, L.A.C.; Traldi, J.B.; Moreira-Filho, O. The Role of the Robertsonian Rearrangements in the Origin of the XX/XY1Y2 Sex Chromosome System and in the Chromosomal Differentiation in Harttia Species (Siluriformes, Loricariidae). Rev. Fish Biol. Fish. 2013, 23, 127–134. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, J.; Zhang, J.; Wang, Z.; Wang, Y.; Cai, M. Cytogenetic Characterization and Description of an X1X1X2X2/X1X2Y Sex Chromosome System in Collichthys lucidus (Richardson, 1844). Acta Oceanol. Sin. 2018, 37, 34–39. [Google Scholar] [CrossRef]
- Prizon, A.C.; Bruschi, D.P.; Borin-Carvalho, L.A.; Cius, A.; Barbosa, L.M.; Ruiz, H.B.; Zawadzki, C.H.; Fenocchio, A.S.; Portela-Castro, A.L. Hidden Diversity in the Populations of the Armored Catfish Ancistrus Kner, 1854 (Loricariidae, Hypostominae) from the Paraná River Basin Revealed by Molecular and Cytogenetic Data. Front. Genet. 2017, 8, 185. [Google Scholar] [CrossRef]
- De Oliveira, R.R.; Feldberg, E.; dos Anjos, M.B.; Zuanon, J. Karyotype Characterization and ZZ/ZW Sex Chromosome Heteromorphism in Two Species of the Catfish Genus Ancistrus Kner, 1854 (Siluriformes: Loricariidae) from the Amazon Basin. Neotrop. Ichthyol. 2007, 5, 301–306. [Google Scholar] [CrossRef]
- Konerat, J.T.; Bueno, V.; Margarido, V.P.; Portela-Castro, A.L.B.; Martins-Santos, I.C. Diversity of Sex Chromosome Systems in Ancistrini (Loricariidae, Hypostominae): ZZ/ZW in Ancistrus taunayi Miranda Ribeiro, 1918. Cytogenet. Genome Res. 2015, 146, 306–310. [Google Scholar] [CrossRef]
- Kapuscinski, J. DAPI: A DNA-Specific Fluorescent Probe. Biotech. Histochem. 1995, 70, 220–233. [Google Scholar] [CrossRef]
- Gornung, E. Twenty Years of Physical Mapping of Major Ribosomal RNA Genes across the Teleosts: A Review of Research. Cytogenet. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary Trends in Animal Ribosomal DNA Loci: Introduction to a New Online Database. Chromosoma 2018, 127, 141–150. [Google Scholar] [CrossRef]
- Blanco, D.R.; Vicari, M.R.; Lui, R.L.; Traldi, J.B.; Bueno, V.; Martinez, J.d.F.; Brandão, H.; Oyakawa, O.T.; Moreira Filho, O. Karyotype Diversity and Evolutionary Trends in Armored Catfish Species of the Genus Harttia (Siluriformes: Loricariidae). Zebrafish 2017, 14, 169–176. [Google Scholar] [CrossRef]
- Paim, F.G.; Nirchio, M.; Oliveira, C.; Rossi, A.R. Sex Chromosomes and Internal Telomeric Sequences in Dormitator latifrons (Richardson 1844) (Eleotridae: Eleotrinae): An Insight into Their Origin in the Genus. Genes 2020, 11, 659. [Google Scholar] [CrossRef]
- Diniz, D.; Laudicina, A.; Bertollo, L.A.C. Chromosomal Location of 18S and 5S rDNA Sites in Triportheus Fish Species (Characiformes, Characidae). Genet. Mol. Biol. 2009, 32, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Diniz, D.; Moreira-Filho, O.; Bertollo, L.A.C. Molecular Cytogenetics and Characterization of a ZZ/ZW Sex Chromosome System in Triportheus nematurus (Characiformes, Characidae). Genetica 2008, 133, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Yano, C.F.; Poltronieri, J.; Bertollo, L.A.C.; Artoni, R.F.; Liehr, T.; de Cioffi, M.B. Chromosomal Mapping of Repetitive DNAs in Triportheus trifurcatus (Characidae, Characiformes): Insights into the Differentiation of the Z and W Chromosomes. PLoS ONE 2014, 9, e90946. [Google Scholar] [CrossRef]
- Artoni, R.F.; Bertollo, L.A.C. Evolutionary Aspects of the ZZ/ZW Sex Chromosome System in the Characidae Fish, Genus Triportheus. A Monophyletic State and NOR Location on the W Chromosome. Heredity 2002, 89, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Gandhi, D.; Kumon, T.; Yamashita, Y.M. Ribosomal DNA Instability as a Potential Cause of Karyotype Evolution. Mol. Biol. Evol. 2022, 39, msac221. [Google Scholar] [CrossRef] [PubMed]
- Montiel, E.E.; Badenhorst, D.; Lee, L.; Valenzuela, N. Evolution and dosage compensation of nucleolar organizing regions (NORs) mediated by mobile elements in turtles with female (ZZ/ZW) but not with male (XX/XY) heterogamety. J. Evol. Biol. 2022, 35, 1709–1720. [Google Scholar] [CrossRef]
- Valente, G.T.; Mazzuchelli, J.; Ferreira, I.A.; Poletto, A.B.; Fantinatti, B.E.A.; Martins, C. Cytogenetic Mapping of the Retroelements Rex1, Rex3 and Rex6 among Cichlid Fish: New Insights on the Chromosomal Distribution of Transposable Elements. Cytogenet. Genome Res. 2011, 133, 34–42. [Google Scholar] [CrossRef] [PubMed]
- García, E.; Cross, I.; Portela-Bens, S.; Rodríguez, M.E.; García-Angulo, A.; Molina, B.; Cuadrado, A.; Liehr, T.; Rebordinos, L. Integrative Genetic Map of Repetitive DNA in the Sole Solea senegalensis Genome Shows a Rex Transposon Located in a Proto-Sex Chromosome. Sci. Rep. 2019, 9, 17146. [Google Scholar] [CrossRef]
- Ayres-Alves, T.; Cardoso, A.L.; Nagamachi, C.Y.; de Sousa, L.M.; Pieczarka, J.C.; Noronha, R.C.R. Karyotypic Evolution and Chromosomal Organization of Repetitive DNA Sequences in Species of Panaque, Panaqolus, and Scobinancistrus (Siluriformes and Loricariidae) from the Amazon Basin. Zebrafish 2017, 14, 251–260. [Google Scholar] [CrossRef]
- Pety, A.M.; Cardoso, A.L.; Nagamachi, C.Y.; Pieczarka, J.C.; de Sousa, L.M.; Noronha, R.C.R. In Situ Localization of Ribosomal Sites in Peckoltia and Ancistomus (Loricariidae: Hypostominae) from the Amazon Basin. Zebrafish 2018, 15, 263–269. [Google Scholar] [CrossRef]
- Rosa, K.O.; Ziemniczak, K.; de Barros, A.V.; Nogaroto, V.; Almeida, M.C.; Cestari, M.M.; Artoni, R.F.; Vicari, M.R. Numeric and Structural Chromosome Polymorphism in Rineloricarialima (Siluriformes: Loricariidae): Fusion Points Carrying 5S rDNA or Telomere Sequence Vestiges. Rev. Fish Biol. Fish. 2012, 22, 739–749. [Google Scholar] [CrossRef]
- Barros, A.V.; Wolski, M.A.V.; Nogaroto, V.; Almeida, M.C.; Moreira-Filho, O.; Vicari, M.R. Fragile Sites, Dysfunctional Telomere and Chromosome Fusions: What Is 5S rDNA Role? Gene 2017, 608, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Deon, G.A.; Glugoski, L.; Hatanaka, T.; de Sassi, F.M.C.; Nogaroto, V.; Bertollo, L.A.C.; Liehr, T.; Al-Rikabi, A.; Moreira Filho, O.; de Cioffi, M.B.; et al. Evolutionary Breakpoint Regions and Chromosomal Remodeling in Harttia (Siluriformes: Loricariidae) Species Diversification. Genet. Mol. Biol. 2022, 45, e20210170. [Google Scholar] [CrossRef]
- Deon, G.A.; Glugoski, L.; Vicari, M.R.; Nogaroto, V.; de Sassi, F.M.C.; de Cioffi, M.B.; Liehr, T.; Bertollo, L.A.C.; Moreira-Filho, O. Highly Rearranged Karyotypes and Multiple Sex Chromosome Systems in Armored Catfishes from the Genus Harttia (Teleostei, Siluriformes). Genes 2020, 11, 1366. [Google Scholar] [CrossRef]
- de Sassi, F.M.C.; Moreira-Filho, O.; Deon, G.A.; Sember, A.; Bertollo, L.A.C.; Liehr, T.; Oliveira, V.C.S.; Viana, P.F.; Feldberg, E.; Vicari, M.R.; et al. Adding New Pieces to the Puzzle of Karyotype Evolution in Harttia (Siluriformes, Loricariidae): Investigation of Amazonian Species. Biology 2021, 10, 922. [Google Scholar] [CrossRef]
- Ditcharoen, S.; Antonio Carlos Bertollo, L.; Ráb, P.; Hnátková, E.; Franco Molina, W.; Liehr, T.; Tanomtong, A.; Triantaphyllidis, C.; Ozouf-Costaz, C.; Tongnunui, S.; et al. Genomic Organization of Repetitive DNA Elements and Extensive Karyotype Diversity of Silurid Catfishes (Teleostei: Siluriformes): A Comparative Cytogenetic Approach. Int. J. Mol. Sci. 2019, 20, 3545. [Google Scholar] [CrossRef]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity 2005, 95, 118–128. [Google Scholar] [CrossRef]
- Zimmer, E.A.; Martins, S.L.; Beverly, S.M.; Kan, Y.W.; Wilson, A.C. Rapid duplication and loss of genes coding for the alpha chains of hemoglobin. Proc. Natl. Acad. Sci. USA 1980, 77, 2158–2162. [Google Scholar] [CrossRef]
- Dover, G.A. Molecular drive: A cohesive model of species evolution. Nature 1982, 199, 111–117. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nirchio, M.; Oliveira, C.; de Bello Cioffi, M.; de Menezes Cavalcante Sassi, F.; Valdiviezo, J.; Paim, F.G.; Soares, L.B.; Rossi, A.R. Occurrence of Sex Chromosomes in Fish of the Genus Ancistrus with a New Description of Multiple Sex Chromosomes in the Ecuadorian Endemic Ancistrus clementinae (Loricariidae). Genes 2023, 14, 306. https://doi.org/10.3390/genes14020306
Nirchio M, Oliveira C, de Bello Cioffi M, de Menezes Cavalcante Sassi F, Valdiviezo J, Paim FG, Soares LB, Rossi AR. Occurrence of Sex Chromosomes in Fish of the Genus Ancistrus with a New Description of Multiple Sex Chromosomes in the Ecuadorian Endemic Ancistrus clementinae (Loricariidae). Genes. 2023; 14(2):306. https://doi.org/10.3390/genes14020306
Chicago/Turabian StyleNirchio, Mauro, Claudio Oliveira, Marcelo de Bello Cioffi, Francisco de Menezes Cavalcante Sassi, Jonathan Valdiviezo, Fabilene Gomes Paim, Leticia Batista Soares, and Anna Rita Rossi. 2023. "Occurrence of Sex Chromosomes in Fish of the Genus Ancistrus with a New Description of Multiple Sex Chromosomes in the Ecuadorian Endemic Ancistrus clementinae (Loricariidae)" Genes 14, no. 2: 306. https://doi.org/10.3390/genes14020306
APA StyleNirchio, M., Oliveira, C., de Bello Cioffi, M., de Menezes Cavalcante Sassi, F., Valdiviezo, J., Paim, F. G., Soares, L. B., & Rossi, A. R. (2023). Occurrence of Sex Chromosomes in Fish of the Genus Ancistrus with a New Description of Multiple Sex Chromosomes in the Ecuadorian Endemic Ancistrus clementinae (Loricariidae). Genes, 14(2), 306. https://doi.org/10.3390/genes14020306