
Mutations Linked to Insecticide Resistance Not Detected in the Ace-1 or VGSC Genes in Nyssorhynchus darlingi from Multiple Localities in Amazonian Brazil and Peru


 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
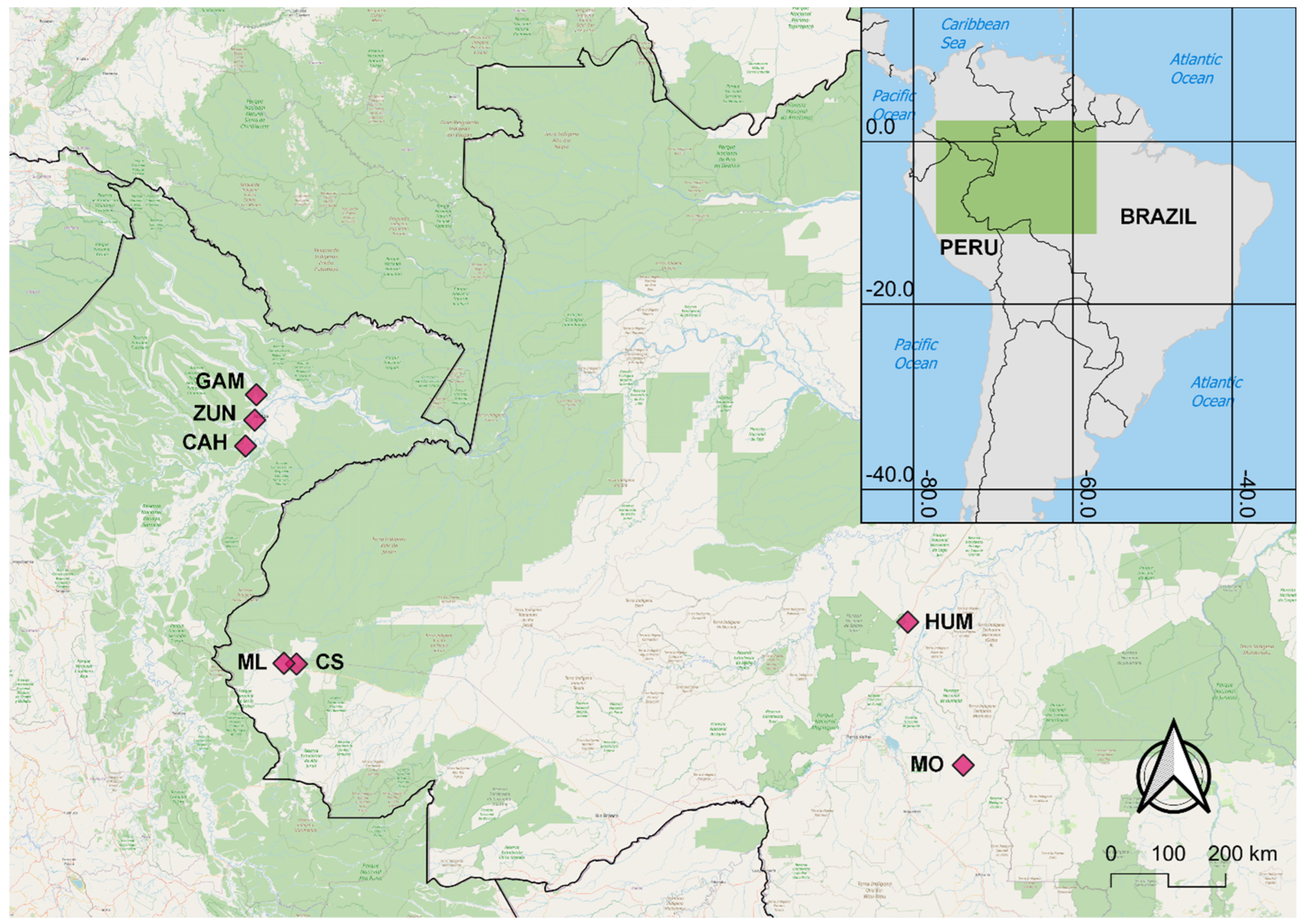
2.1. Sample Collection
2.2. Molecular Analysis
3. Results
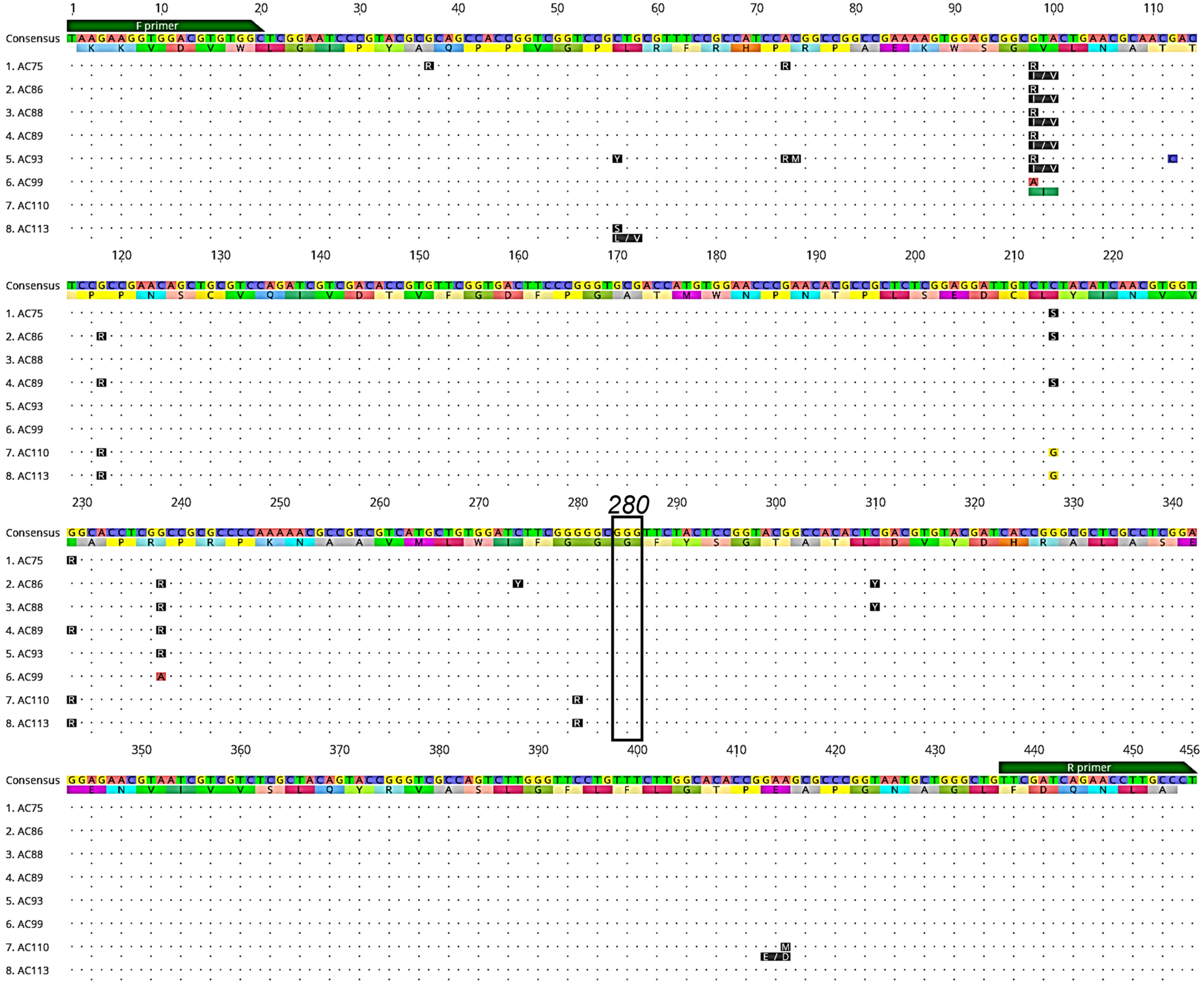
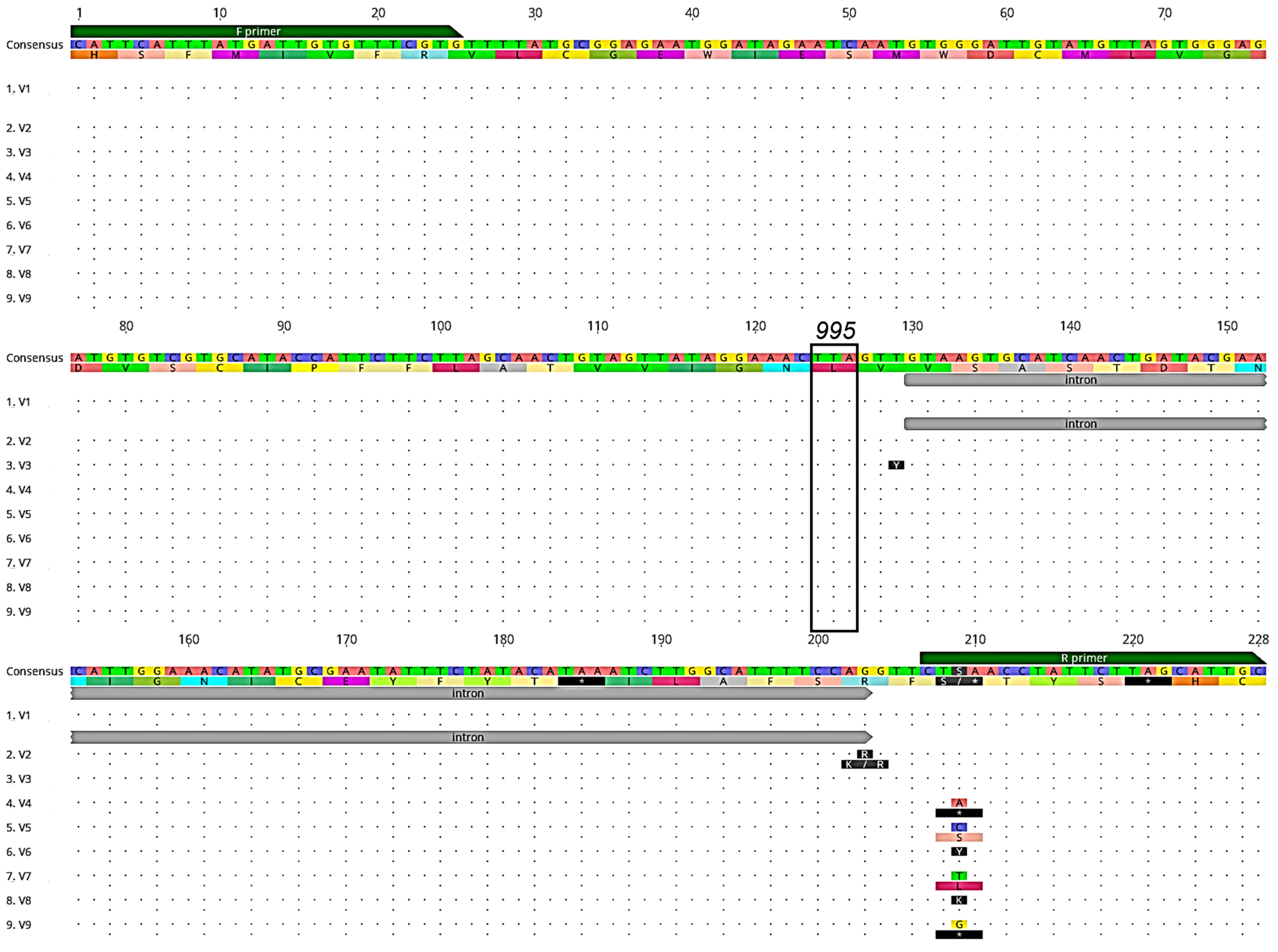
3.1. Detection of Voltage-Gated Sodium Channel (VGSC) Mutations
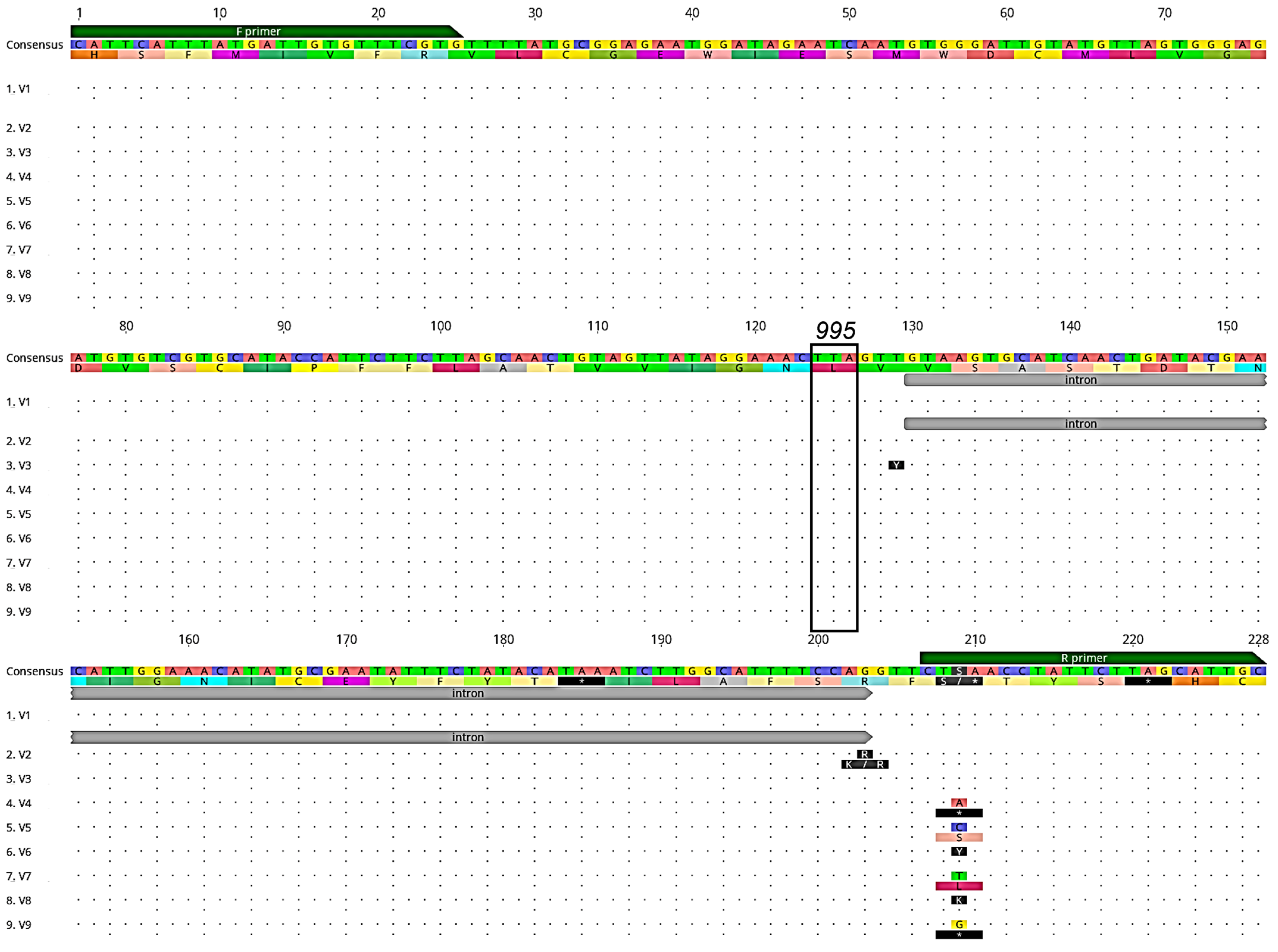
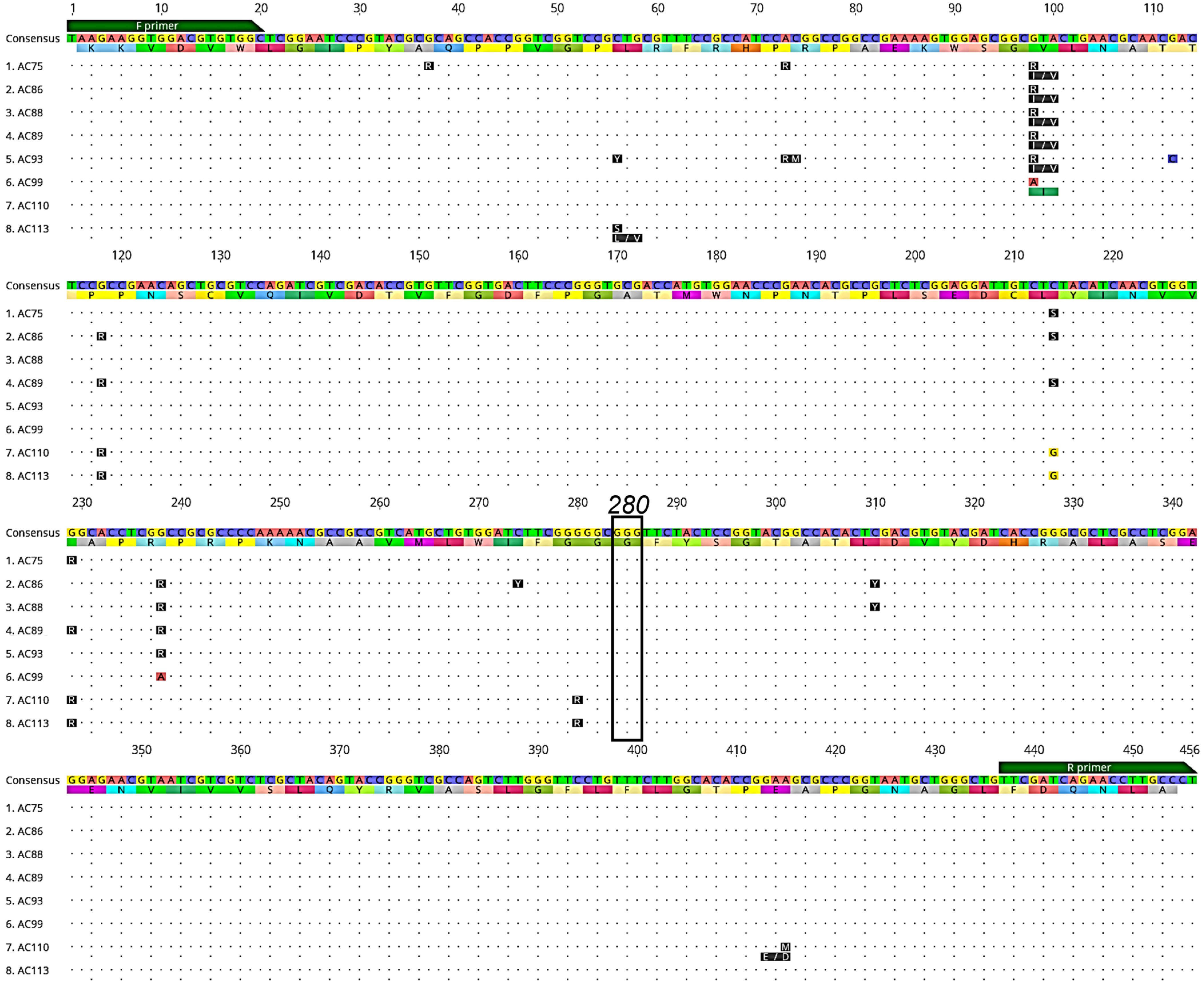
3.2. Detection of Acetylcholinesterase-1 (Ace-1) Mutations
3.3. Data Management
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alexander, N.; DE LA Hoz, F.; Cruz, J.; Hall, A.J.; Prieto, G.; Arroyo, J.A.; Rodríguez, M.; Suárez, M.; Pérez, L.; Cotacio, M.C.; et al. Case-control study of mosquito nets against malaria in the Amazon region of Colombia. Am. J. Trop. Med. Hyg. 2005, 73, 140–148. [Google Scholar] [CrossRef]
- Sherrard-Smith, E.; Skarp, J.E.; Beale, A.D.; Fornadel, C.; Norris, L.C.; Moore, S.J.; Mihreteab, S.; Charlwood, J.D.; Bhatt, S.; Winskill, P.; et al. Mosquito feeding behavior and how it influences residual malaria transmission across Africa. Proc. Natl. Acad. Sci. USA 2019, 116, 15086–15095. [Google Scholar] [CrossRef]
- Parker, B.S.; Paredes Olortegui, M.; Penataro Yori, P.; Escobedo, K.; Florin, D.; Rengifo Pinedo, S.; Cardenas Greffa, R.; Capcha Vega, L.; Rodriguez Ferrucci, H.; Pan, W.K.; et al. Hyperendemic malaria transmission in areas of occupation-related travel in the Peruvian Amazon. Malar. J. 2013, 12, 178. [Google Scholar] [CrossRef]
- Carrasco-Escobar, G.; Gamboa, D.; Castro, M.C.; Bangdiwala, S.I.; Rodriguez, H.; Contreras-Mancilla, J.; Alava, F.; Speybroeck, N.; Lescano, A.G.; Vinetz, J.M.; et al. Micro-epidemiology and spatial heterogeneity of P. vivax parasitaemia in riverine communities of the Peruvian Amazon: A multilevel analysis. Sci. Rep. 2017, 7, 8082. [Google Scholar] [CrossRef]
- Moreno, J.E.; Rubio-Palis, Y.; Paez, E.; Perez, E.; Sanchez, V. Abundance, biting behaviour and parous rate of anopheline mosquito species in relation to malaria incidence in gold-mining areas of southern Venezuela. Med. Vet. Entomol. 2007, 21, 339–349. [Google Scholar] [CrossRef]
- Moreno, M.; Saavedra, M.P.; Bickersmith, S.A.; Lainhart, W.; Tong, C.; Alava, F.; Vinetz, J.M.; Conn, J.E. Implications for changes in Anopheles darlingi biting behaviour in three communities in the peri-Iquitos region of Amazonian Peru. Malar. J. 2015, 14, 290. [Google Scholar] [CrossRef]
- Durnez, L.; Coosemans, M. Residual transmission of malaria: An old issue for new approaches. In Anopheles Mosquitoes—New Insights into Malaria Vectors; Manguin, S., Ed.; Intech Open: Rijeka, Croatia, 2013; pp. 671–704. [Google Scholar]
- Sousa, J.O.; de Albuquerque, B.C.; Coura, J.R.; Suárez-Mutis, M.C. Use and retention of long-lasting insecticidal nets (LLINs) in a malaria risk area in the Brazilian Amazon: A 5-year follow-up intervention. Malar. J. 2019, 18, 100. [Google Scholar] [CrossRef]
- World Health Organization. World Malaria Report 2015; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Soto-Calle, V.; Rosas-Aguirre, A.; Llanos-Cuentas, A.; Abatih, E.; DeDeken, R.; Rodriguez, H.; Rosanas-Urgell, A.; Gamboa, D.; Alessandro, U.D.; Erhart, A.; et al. Spatio-temporal analysis of malaria incidence in the Peruvian Amazon Region between 2002 and 2013. Sci. Rep. 2017, 7, 40350. [Google Scholar] [CrossRef]
- World Health Organization. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021; p. 322. [Google Scholar]
- Gabaldón-Figueira, J.C.; Villegas, L.; Grillet, M.E.; Lezaun, J.; Pocaterra, L.; Bevilacqua, M.; Paniz-Mondolfi, A.; González, O.N.; Chaccour, C. Malaria in Venezuela: Gabaldón’s legacy scattered to the winds. Lancet Glob. Health 2021, 9, e584–e585. [Google Scholar] [CrossRef]
- Ministerio de Salud. Documento Technico: Plan Hacia la Malaria en el Peru 2022–2030; Ministerio de Salud: Lima, Pero, 2022; p. 60.
- World Health Organization. World Malaria Report 2020: 20 Years of Global Progress and Challenges; World Health Organization: Geneva, Switzerland, 2020; p. 247. [Google Scholar]
- Şengül Demirak, M.; Canpolat, E. Plant-based bioinsecticides for mosquito control: Impact on insecticide resistance and disease transmission. Insects 2022, 13, 162. [Google Scholar] [CrossRef]
- Orjuela, L.I.; Álvarez-Diaz, D.A.; Morales, J.A.; Grisales, N.; Ahumada, M.L.; Venegas, H.J.; Quiñones, M.L.; Yasnot, M.F. Absence of knockdown mutations in pyrethroid and DDT resistant populations of the main malaria vectors in Colombia. Malar. J. 2019, 18, 384. [Google Scholar] [CrossRef] [PubMed]
- Quinones, M.L.; Norris, D.E.; Conn, J.E.; Moreno, M.; Burkot, T.R.; Bugoro, H.; Keven, J.B.; Cooper, R.; Yan, G.; Rosas, A.; et al. Insecticide resistance in areas under investigation by the International Centers of Excellence for Malaria Research: A challenge for malaria control and elimination. Am. J. Trop. Med. Hyg. 2015, 93, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Lol, J.C.; Castañeda, D.; Mackenzie-Impoinvil, L.; Romero, C.G.; Lenhart, A.; Padilla, N.R. Development of molecular assays to detect target-site mechanisms associated with insecticide resistance in malaria vectors from Latin America. Malar. J. 2019, 18, 202. [Google Scholar] [CrossRef]
- Baia-da-Silva, D.C.; Brito-Sousa, J.D.; Rodovalho, S.R.; Peterka, C.; Moresco, G.; Lapouble, O.M.M.; Melo, G.C.; Sampaio, V.S.; Alecrim, M.; Pimenta, P.; et al. Current vector control challenges in the fight against malaria in Brazil. Rev. Soc. Bras. Med. Trop. 2019, 52, e20180542. [Google Scholar] [CrossRef]
- Hiwat, H.; Bretas, G. Ecology of Anopheles darlingi Root with respect to vector importance: A review. Parasit. Vectors 2011, 4, 177. [Google Scholar] [CrossRef]
- World Health Organization. Test Procedures for Insecticide Resistance Monitoring in Malaria Vectors, Bio-Efficacy and Persistence of Insecticides on Treated Surfaces: Report of the WHO Informal Consultation; World Health Organization: Geneva, Switzerland, 1998; p. 43. [Google Scholar]
- Lenhart, A.; Chan, A.; Vizcaino, L.; Brogdon, W. Manual for Evaluating Insecticide Resistance Using the CDC Bottle Bioassay; CDC: Atlanta, GA, USA, 2023; p. 31. [Google Scholar]
- Brogdon, W.; Chan, A. Guideline for Evaluating Insecticide Resistance in Vectors Using the CDC Bottle Bioassa; CDC: Atlanta, GA, USA, 2011; p. 28. [Google Scholar]
- World Health Organization. Global Report on Insecticide Resistance in Malaria Vectors: 2010–2016; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Weill, M.; Lutfalla, G.; Mogensen, K.; Chandre, F.; Berthomieu, A.; Berticat, C.; Pasteur, N.; Philips, A.; Fort, P.; Raymond, M. Insecticide resistance in mosquito vectors. Nature 2003, 423, 136–137. [Google Scholar] [CrossRef]
- Davies, T.G.; Field, L.M.; Usherwood, P.N.; Williamson, M.S. DDT, pyrethrins, pyrethroids and insect sodium channels. IUBMB Life 2007, 59, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Luen, S.C.; Shalaby, A.M. Preliminary note on the development of DDT-resistance in Anopheles culicifacies Giles in Panchmahals District, Gujerat State, India. Bull. World Health Organ. 1962, 26, 128–134. [Google Scholar]
- World Health Organization. Division of Vector, B.; Control. Instructions for Determining the Susceptibility or Resistance of Mosquito Larvae to Insecticides; World Health Organization: Geneva, Switzerland, 1981. [Google Scholar]
- Suarez, M.F.; Quiñones, M.L.; Palacios, J.D.; Carrillo, A. First record of DDT resistance in Anopheles darlingi. J. Am. Mosq. Control Assoc. 1990, 6, 72–74. [Google Scholar]
- Carrasco, D.; Lefèvre, T.; Moiroux, N.; Pennetier, C.; Chandre, F.; Cohuet, A. Behavioural adaptations of mosquito vectors to insecticide control. Curr. Opin. Insect Sci. 2019, 34, 48–54. [Google Scholar] [CrossRef]
- Roberts, D.R.; Alecrim, W.D. Behavioral response of Anopheles darlingi to DDT-sprayed house walls in Amazonia. Bull. Pan Am. Health Organ. 1991, 25, 210–217. [Google Scholar] [PubMed]
- Panini, M.; Manicardi, G.C.; Moores, G.D.; Mazzoni, E. An overview of the main pathways of metabolic resistance in insects. Invertebrate Surviv. J. 2016, 13, 326–335. [Google Scholar]
- Vontas, J.; Katsavou, E.; Mavridis, K. Cytochrome P450-based metabolic insecticide resistance in Anopheles and Aedes mosquito vectors: Muddying the waters. Pestic. Biochem. Physiol. 2020, 170, 104666. [Google Scholar] [CrossRef] [PubMed]
- Lucas, E.R.; Rockett, K.A.; Lynd, A.; Essandoh, J.; Grisales, N.; Kemei, B.; Njoroge, H.; Hubbart, C.; Rippon, E.J.; Morgan, J.; et al. A high throughput multi-locus insecticide resistance marker panel for tracking resistance emergence and spread in Anopheles gambiae. Sci. Rep. 2019, 9, 13335. [Google Scholar] [CrossRef]
- Riveron, J.M.; Tchouakui, M.; Mugenzi, L.; Menze, B.D.; Chiang, M.-C.; Wondji, C.S. Insecticide resistance in malaria vectors: An update at a global scale. In Towards Malaria Elimination; Sylvie, M., Vas, D., Eds.; IntechOpen: Rijeka, Croatia, 2018. [Google Scholar]
- Silva, A.P.; Santos, J.M.; Martins, A.J. Mutations in the voltage-gated sodium channel gene of anophelines and their association with resistance to pyrethroids—A review. Parasit. Vectors 2014, 7, 450. [Google Scholar] [CrossRef] [PubMed]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Mavridis, K.; Wipf, N.; Müller, P.; Traoré, M.M.; Muller, G.; Vontas, J. Detection and monitoring of insecticide resistance mutations in Anopheles gambiae: Individual vs. pooled specimens. Genes 2018, 9, 479. [Google Scholar] [CrossRef]
- Kisinza, W.; Kabula, B.; Tungu, P.; Sindato, C.; Mweya, C.; Massue, D.; Emidi, B.; Kitau, J.; Chacha, M.; Batengana, B.; et al. Detection and Monitoring of Insecticide Resistance in Malaria Vectors in Tanzania Mainland; Technical Report of the National Institute for Medical Research, Tanzania: Muheza, Tanzania, 2011. [Google Scholar]
- World Health Organization. Malaria Threats Map. Available online: http://www.who.int/teams/global-malaria-programme/surveillance/malaria-threats-map (accessed on 13 June 2023).
- Laporta, G.Z.; Ilacqua, R.C.; Bergo, E.S.; Chaves, L.S.M.; Rodovalho, S.R.; Moresco, G.G.; Figueira, E.A.G.; Massad, E.; de Oliveira, T.M.P.; Bickersmith, S.A.; et al. Malaria transmission in landscapes with varying deforestation levels and timelines in the Amazon: A longitudinal spatiotemporal study. Sci. Rep. 2021, 11, 6477. [Google Scholar] [CrossRef]
- Sallum, M.A.M.; Conn, J.E.; Bergo, E.S.; Laporta, G.Z.; Chaves, L.S.M.; Bickersmith, S.A.; de Oliveira, T.M.P.; Figueira, E.A.G.; Moresco, G.; Olívêr, L.; et al. Vector competence, vectorial capacity of Nyssorhynchus darlingi and the basic reproduction number of Plasmodium vivax in agricultural settlements in the Amazonian Region of Brazil. Malar. J. 2019, 18, 117. [Google Scholar] [CrossRef]
- Prussing, C.; Moreno, M.; Saavedra, M.P.; Bickersmith, S.A.; Gamboa, D.; Alava, F.; Schlichting, C.D.; Emerson, K.J.; Vinetz, J.M.; Conn, J.E. Decreasing proportion of Anopheles darlingi biting outdoors between long-lasting insecticidal net distributions in peri-Iquitos, Amazonian Peru. Malar. J. 2018, 17, 86. [Google Scholar] [CrossRef]
- Consoli, R.A.; Lourenco-de-Oliveira, R. Principais Mosquitos de Importância Sanitária no Brasil; Editora Fiocruz: Fundação Oswaldo Cruz, Brazil, 1994; p. 228. [Google Scholar]
- Faran, M.E.; Linthicum, K.J. A handbook of the Amazonian species of Anopheles (Nyssorhynchus) (Diptera: Culicidae). Mosq. Syst. 1981, 13, 1–81. [Google Scholar]
- Forattini, O.P. Entomologia Medica; Faculdade de Higiene e Sáude Publica: São Paulo, Brazil, 1962; Volume 1, p. 622. [Google Scholar]
- Berman, H.B. Stat Trek: Random Number Generator. Available online: http://stattrek.com/statistics/random-number-generator.aspx (accessed on 21 January 2021).
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Loureiro, A.C.; Araki, A.S.; Bruno, R.V.; Lima, J.B.P.; Ladeia-Andrade, S.; Santacoloma, L.; Martins, A.J. Molecular diversity of genes related to biological rhythms (period and timeless) and insecticide resistance (Na V and ace-1) in Anopheles darlingi. Mem. Inst. Oswaldo Cruz 2023, 118, e220159. [Google Scholar] [CrossRef]
- Lol, J.C.; Castellanos, M.E.; Liebman, K.A.; Lenhart, A.; Pennington, P.M.; Padilla, N.R. Molecular evidence for historical presence of knock-down resistance in Anopheles albimanus, a key malaria vector in Latin America. Parasit. Vectors 2013, 6, 268. [Google Scholar] [CrossRef]
- Mackenzie-Impoinvil, L.; Weedall, G.D.; Lol, J.C.; Pinto, J.; Vizcaino, L.; Dzuris, N.; Riveron, J.; Padilla, N.; Wondji, C.; Lenhart, A. Contrasting patterns of gene expression indicate differing pyrethroid resistance mechanisms across the range of the New World malaria vector Anopheles albimanus. PLoS ONE 2019, 14, e0210586. [Google Scholar] [CrossRef] [PubMed]
- Braga, T.A.; Loureiro, A.C.; Lima, J.B.P.; Martins, A.J. Insecticide resistance in Anopheles albitarsis s.s. from a rice production field, with the first record of Kdr mutation. Res. Sq. 2021; preprint. [Google Scholar] [CrossRef]
- Alimi, T.O.; Fuller, D.O.; Quinones, M.L.; Xue, R.D.; Herrera, S.V.; Arevalo-Herrera, M.; Ulrich, J.N.; Qualls, W.A.; Beier, J.C. Prospects and recommendations for risk mapping to improve strategies for effective malaria vector control interventions in Latin America. Malar. J. 2015, 14, 519. [Google Scholar] [CrossRef] [PubMed]
- Floch, H. [Antimalarial campaign in French Guiana. III. DDT campaigns and their results]. Riv. Malariol. 1955, 34, 77–92. [Google Scholar] [PubMed]
- Angella, A.F.; Gil, L.H.; Silva, L.H.; Ribolla, P.E. Population structure of the malaria vector Anopheles darlingi in Rondonia, Brazilian Amazon, based on mitochondrial DNA. Mem. Inst. Oswaldo Cruz 2007, 102, 953–958. [Google Scholar] [CrossRef]
- Ranson, H. Current and future prospects for preventing malaria transmission via the use of insecticides. Cold Spring Harb. Perspect. Med. 2017, 7, a026823. [Google Scholar] [CrossRef]
- Souris, M.; Marcombe, S.; Laforet, J.; Brey, P.T.; Corbel, V.; Overgaard, H.J. Modeling spatial variation in risk of presence and insecticide resistance for malaria vectors in Laos. PLoS ONE 2017, 12, e0177274. [Google Scholar] [CrossRef]
- Metelo-Matubi, E.; Zanga, J.; Binene, G.; Mvuama, N.; Ngamukie, S.; Nkey, J.; Schopp, P.; Bamba, M.; Irish, S.; Nguya-Kalemba-Maniania, J.; et al. The effect of a mass distribution of insecticide-treated nets on insecticide resistance and entomological inoculation rates of Anopheles gambiae s.l. in Bandundu City, Democratic Republic of Congo. Pan Afr. Med. J. 2021, 40, 118. [Google Scholar] [CrossRef] [PubMed]
- Sanou, A.; Nelli, L.; Guelbéogo, W.M.; Cissé, F.; Tapsoba, M.; Ouédraogo, P.; Sagnon, N.; Ranson, H.; Matthiopoulos, J.; Ferguson, H.M. Insecticide resistance and behavioural adaptation as a response to long-lasting insecticidal net deployment in malaria vectors in the Cascades region of Burkina Faso. Sci. Rep. 2021, 11, 17569. [Google Scholar] [CrossRef]
- Sangbakembi-Ngounou, C.; Costantini, C.; Longo-Pendy, N.M.; Ngoagouni, C.; Akone-Ella, O.; Rahola, N.; Cornelie, S.; Kengne, P.; Nakouné, E.R.; Komas, N.P.; et al. Diurnal biting of malaria mosquitoes in the Central African Republic indicates residual transmission may be “out of control”. Proc. Natl. Acad. Sci. USA 2022, 119, e2104282119. [Google Scholar] [CrossRef]
- Vezenegho, S.B.; Adde, A.; Pommier de Santi, V.; Issaly, J.; Carinci, R.; Gaborit, P.; Dusfour, I.; Girod, R.; Briolant, S. High malaria transmission in a forested malaria focus in French Guiana: How can exophagic Anopheles darlingi thwart vector control and prevention measures? Mem. Inst. Oswaldo Cruz 2016, 111, 561–569. [Google Scholar] [CrossRef]
- Iyer, M.; Skelton, J.; de Wildt, G.; Meza, G. A qualitative study on the use of long-lasting insecticidal nets (LLINs) for the prevention of malaria in the Peruvian Amazon. Malar. J. 2019, 18, 301. [Google Scholar] [CrossRef]
- Chaves, L.S.M.; Bergo, E.S.; Conn, J.E.; Laporta, G.Z.; Prist, P.R.; Sallum, M.A.M. Anthropogenic landscape decreases mosquito biodiversity and drives malaria vector proliferation in the Amazon rainforest. PLoS ONE 2021, 16, e0245087. [Google Scholar] [CrossRef]
- Escobar, D.; Ascencio, K.; Ortiz, A.; Palma, A.; Fontecha, G. Distribution and phylogenetic diversity of Anopheles species in malaria endemic areas of Honduras in an elimination setting. Parasit. Vectors 2020, 13, 333. [Google Scholar] [CrossRef] [PubMed]
- Mosnier, E.; Dusfour, I.; Lacour, G.; Saldanha, R.; Guidez, A.; Gomes, M.S.; Sanna, A.; Epelboin, Y.; Restrepo, J.; Davy, D.; et al. Resurgence risk for malaria, and the characterization of a recent outbreak in an Amazonian border area between French Guiana and Brazil. BMC Infect. Dis. 2020, 20, 373. [Google Scholar] [CrossRef]
- Dusfour, I.; Carinci, R.; Issaly, J.; Gaborit, P.; Girod, R. A survey of adult anophelines in French Guiana: Enhanced descriptions of species distribution and biting responses. J. Vector Ecol. 2013, 38, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Pommier de Santi, V.; Dusfour, I.; de Parseval, E.; Lespinet, B.; Nguyen, C.; Gaborit, P.; Carinci, R.; Hyvert, G.; Girod, R.; Briolant, S. Risk of daytime transmission of malaria in the French Guiana rain forest. Med. Sante Trop. 2017, 27, 111–112. [Google Scholar] [CrossRef] [PubMed]
- Hiwat, H.; Issaly, J.; Gaborit, P.; Somai, A.; Samjhawan, A.; Sardjoe, P.; Soekhoe, T.; Girod, R. Behavioral heterogeneity of Anopheles darlingi (Diptera: Culicidae) and malaria transmission dynamics along the Maroni River, Suriname, French Guiana. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Liebman, K.A.; Pinto, J.; Valle, J.; Palomino, M.; Vizcaino, L.; Brogdon, W.; Lenhart, A. Novel mutations on the ace-1 gene of the malaria vector Anopheles albimanus provide evidence for balancing selection in an area of high insecticide resistance in Peru. Malar. J. 2015, 14, 74. [Google Scholar] [CrossRef]
- Reid, M.C.; McKenzie, F.E. The contribution of agricultural insecticide use to increasing insecticide resistance in African malaria vectors. Malar. J. 2016, 15, 107. [Google Scholar] [CrossRef]
- Chouaïbou, M.S.; Fodjo, B.K.; Fokou, G.; Allassane, O.F.; Koudou, B.G.; David, J.P.; Antonio-Nkondjio, C.; Ranson, H.; Bonfoh, B. Influence of the agrochemicals used for rice and vegetable cultivation on insecticide resistance in malaria vectors in southern Côte d’Ivoire. Malar. J. 2016, 15, 426. [Google Scholar] [CrossRef]
- Miller, M.W.; Tren, R. Implications of public-health insecticide resistance and replacement costs for malaria control: Challenges and policy options for endemic countries and donors. Res. Rep. Trop. Med. 2012, 3, 1–19. [Google Scholar] [CrossRef]
- Campos, K.B.; Martins, A.J.; Rodovalho, C.M.; Bellinato, D.F.; Dias, L.D.S.; Macoris, M.; Andrighetti, M.T.M.; Lima, J.B.P.; Obara, M.T. Assessment of the susceptibility status of Aedes aegypti (Diptera: Culicidae) populations to pyriproxyfen and malathion in a nation-wide monitoring of insecticide resistance performed in Brazil from 2017 to 2018. Parasit. Vectors 2020, 13, 531. [Google Scholar] [CrossRef]
- Lopes, R.P.; Lima, J.B.P.; Martins, A.J. Insecticide resistance in Culex quinquefasciatus Say, 1823 in Brazil: A review. Parasit. Vectors 2019, 12, 591. [Google Scholar] [CrossRef]
- Santos, R.L.; Fayal Ada, S.; Aguiar, A.E.; Vieira, D.B.; Póvoa, M.M. [Evaluation of the residual effect of pyrethroids on Anopheles in the Brazilian Amazon]. Rev. Saude Publica 2007, 41, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, A.; Galardo, A.K.R.; Lima, L.A.; Câmara, D.C.P.; Müller, J.N.; Barroso, J.F.S.; Lapouble, O.M.M.; Rodovalho, C.M.; Ribeiro, K.A.N.; Lima, J.B.P. Efficacy of insecticides used in indoor residual spraying for malaria control: An experimental trial on various surfaces in a “test house”. Malar. J. 2019, 18, 345. [Google Scholar] [CrossRef]
- Kamareddine, L. The biological control of the malaria vector. Toxins 2012, 4, 748–767. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Locality | State/Dept | Country | Latitude | Longitude | Collection Year | Ace-1 N | VGSC N |
|---|---|---|---|---|---|---|---|
| Cruzeiro do Sul (CS) | Acre | Brazil | −7.631889 | −72.688722 | 2015 | 49 | 50 |
| Humaitá (HUM) | Amazonas | Brazil | −6.980687 | −63.100031 | 2016 | 49 | 49 |
| Machadinho d’Oeste (MO) | Rondônia | Brazil | −9.193528 | −62.230107 | 2015 | 50 | 50 |
| Mâncio Lima (ML) | Acre | Brazil | −7.620124 | −72.885559 | 2015 | 50 | 50 |
| Gamitanacocha (GAM) | Loreto | Peru | −3.426000 | −73.318000 | 2018 | 50 | 50 |
| Zungarococha (ZUN) | Loreto | Peru | −3.824560 | −73.343880 | 2019; 2021 | 106 | 107 |
| Cahuide (CAH) | Loreto | Peru | −4.230350 | −73.487833 | 2014; 2019 | 139 | 139 |
| TOTAL | 493 | 495 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bickersmith, S.A.; Jurczynski, J.D.; Sallum, M.A.M.; Chaves, L.S.M.; Bergo, E.S.; Rodriguez, G.A.D.; Morante, C.A.; Rios, C.T.; Saavedra, M.P.; Alava, F.; et al. Mutations Linked to Insecticide Resistance Not Detected in the Ace-1 or VGSC Genes in Nyssorhynchus darlingi from Multiple Localities in Amazonian Brazil and Peru. Genes 2023, 14, 1892. https://doi.org/10.3390/genes14101892
Bickersmith SA, Jurczynski JD, Sallum MAM, Chaves LSM, Bergo ES, Rodriguez GAD, Morante CA, Rios CT, Saavedra MP, Alava F, et al. Mutations Linked to Insecticide Resistance Not Detected in the Ace-1 or VGSC Genes in Nyssorhynchus darlingi from Multiple Localities in Amazonian Brazil and Peru. Genes. 2023; 14(10):1892. https://doi.org/10.3390/genes14101892
Chicago/Turabian StyleBickersmith, Sara A., John D. Jurczynski, Maria Anice Mureb Sallum, Leonardo S. M. Chaves, Eduardo S. Bergo, Gloria A. D. Rodriguez, Clara A. Morante, Carlos T. Rios, Marlon P. Saavedra, Freddy Alava, and et al. 2023. "Mutations Linked to Insecticide Resistance Not Detected in the Ace-1 or VGSC Genes in Nyssorhynchus darlingi from Multiple Localities in Amazonian Brazil and Peru" Genes 14, no. 10: 1892. https://doi.org/10.3390/genes14101892
APA StyleBickersmith, S. A., Jurczynski, J. D., Sallum, M. A. M., Chaves, L. S. M., Bergo, E. S., Rodriguez, G. A. D., Morante, C. A., Rios, C. T., Saavedra, M. P., Alava, F., Gamboa, D., Vinetz, J. M., & Conn, J. E. (2023). Mutations Linked to Insecticide Resistance Not Detected in the Ace-1 or VGSC Genes in Nyssorhynchus darlingi from Multiple Localities in Amazonian Brazil and Peru. Genes, 14(10), 1892. https://doi.org/10.3390/genes14101892