
Selection and Validation of Reference Genes for qRT-PCR Gene Expression Analysis in Kengyilia melanthera

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. RNA Extraction and cDNA Synthesis
2.3. Primer Design
2.4. Quantitative RT-PCR Amplification
2.5. Analysis of Reference Gene Candidates’ Expression
2.6. Validation of Reference Genes
3. Results
3.1. Primer Specificity and Amplification Efficiency
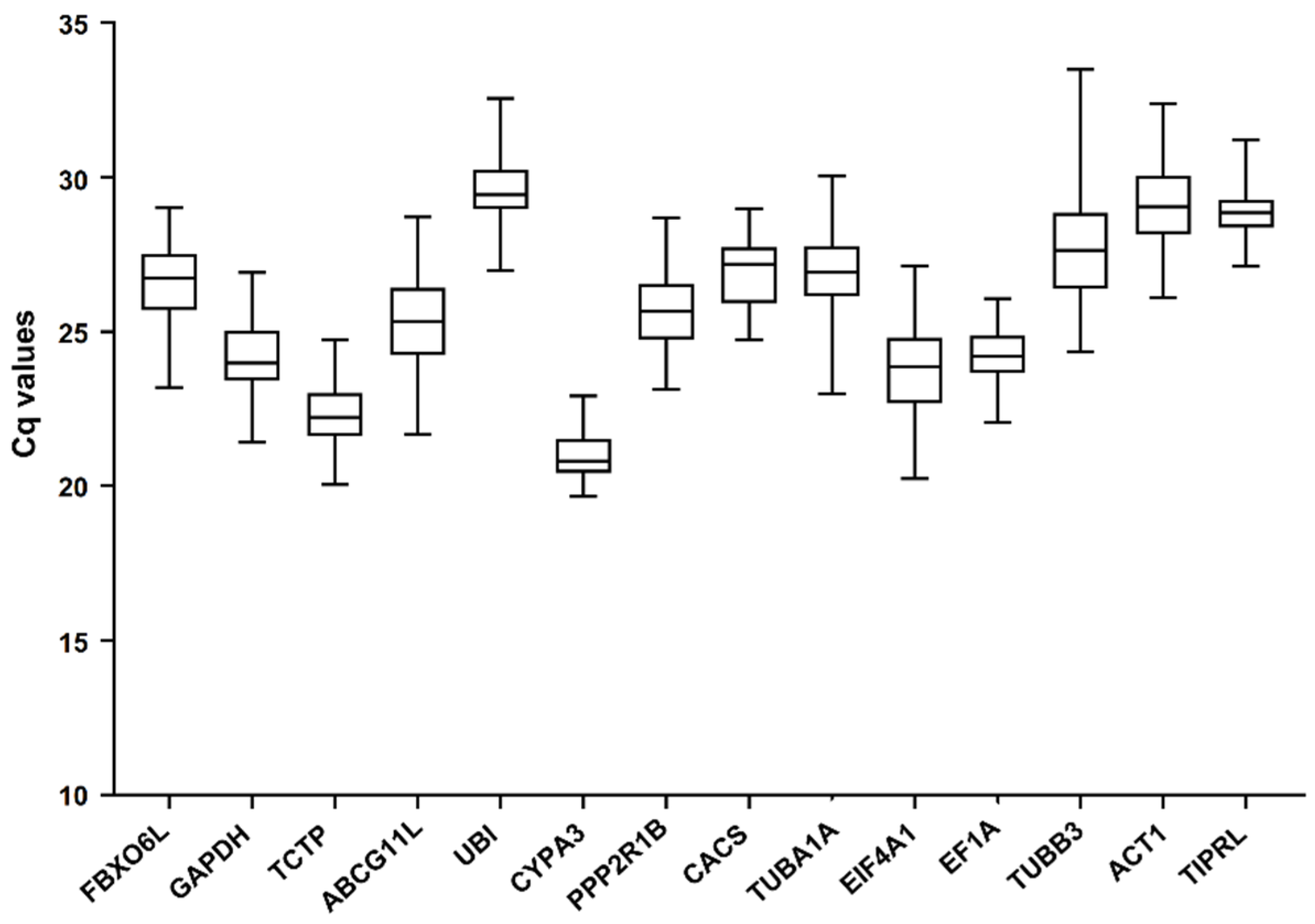
3.2. Expression Profile of the 14 Reference Gene Candidates in Response to Different Treatments
3.3. Analysis of Reference Genes Stability
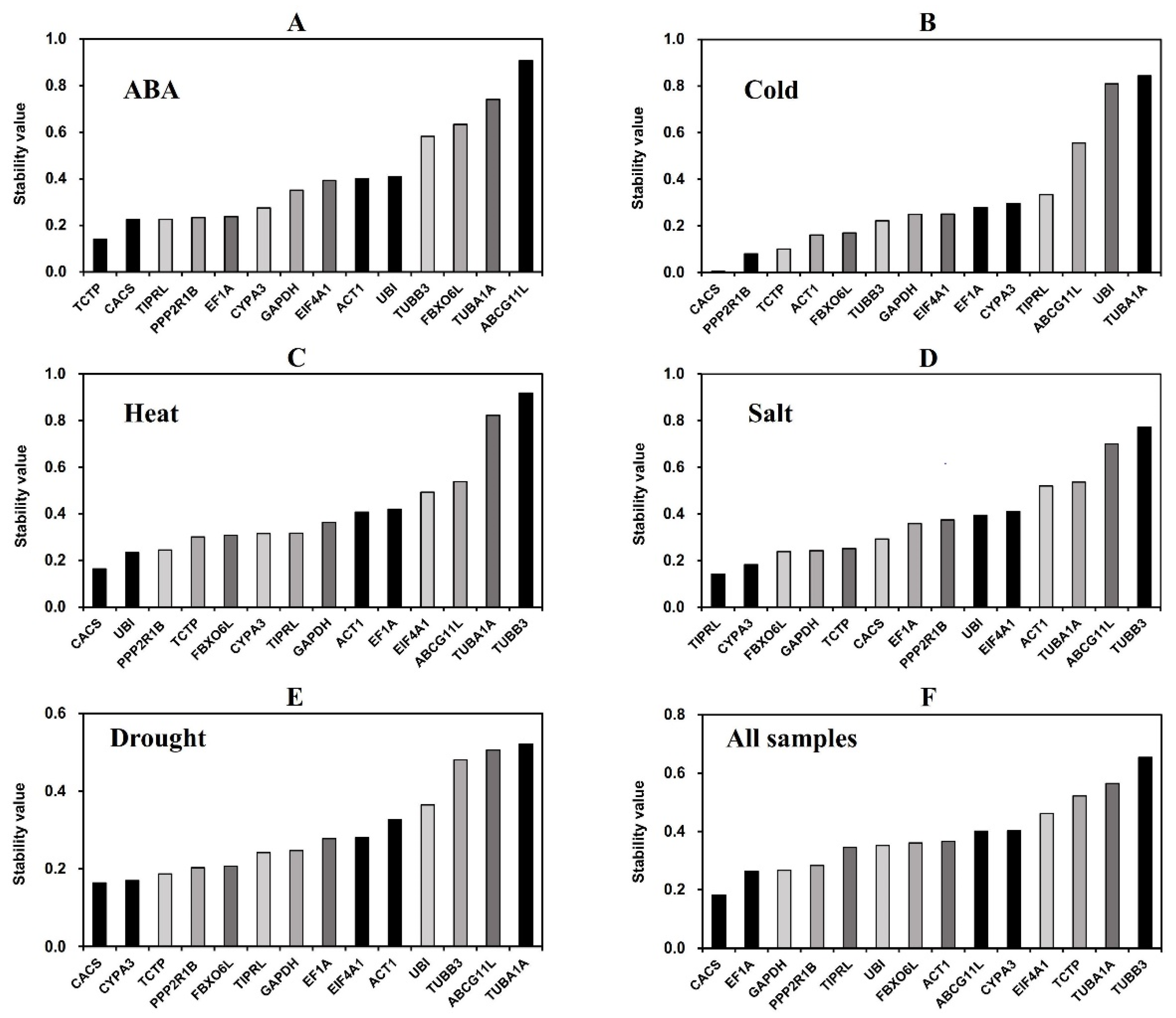
3.3.1. GeNorm Analysis
3.3.2. NormFinder Analysis
3.3.3. BestKeeper Analysis
3.3.4. RefFinder Analysis
3.4. Validation of The Reference Genes Identified from This Study
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yen, C.; Yang, J.L. Kengyilia gobicola, a new taxon from West China. Can. J. Bot. 1990, 68, 1894–1897. [Google Scholar]
- Yen, C.; Yang, J.; Bernard, R.B. Biosystematics of Triticeae: Volume III. Kengyilia Douglasdeweya Agropyron Australopyrum Anthosachne; China Agriculture Press: Beijing, China, 2013. [Google Scholar]
- Wan, Y.F.; Yen, C.; Yang, J.L. The diversity of head-scab resistance in Triticeae and their relation to ecological conditions. Euphytica 1997, 97, 277–281. [Google Scholar]
- Dewey, D.R. The Genomic System of Classification as a Guide to Intergeneric Hybridization with the Perennial Triticeae. In Gene Manipulation in Plant Improvement; Springer: Boston, MA, USA, 1984; pp. 209–279. [Google Scholar]
- Crespo-Herrera, L.A.; Garkava-Gustavsson, L.; Åhman, I. A systematic review of rye (Secale cereale L.) as a source of resistance to pathogens and pests in wheat (Triticum aestivum L.). Hereditas 2017, 154, 14. [Google Scholar]
- Fatih, A.M.B. Analysis of the breeding potential of wheat-Agropyron and wheat-Elymus derivatives: I. Agronomic and quality characteristics. Hereditas 1983, 98, 287–295. [Google Scholar]
- Subbarao, G.V.; Kishii, M.; Bozal-Leorri, A.; Ortiz-Monasterio, I.; Gao, X.; Ibba, M.I.; Karwat, H.; Gonzalez-Moro, M.; Gonzalez-Murua, C.; Tadashi, Y.; et al. Enlisting wild grass genes to combat nitrification in wheat farming: A nature-based solution. Proc. Natl. Acad. Sci. USA 2021, 118, e2106595118. [Google Scholar]
- Graybosch, R.; Bai, G.; Amand, P.S.; Sarath, G.; Evolution, C. Persistence of rye (Secale cereale L.) chromosome arm 1RS in wheat (Triticum aestivum L.) breeding programs of the Great Plains of North America. Genet. Resour. Crop Evol. 2019, 66, 941–950. [Google Scholar]
- Jin, Y.H.; Liu, F.; Huang, W.; Sun, Q.; Huang, X.Z. Identification of reliable reference genes for qRT-PCR in the ephemeral plant Arabidopsis pumila based on full-length transcriptome data. Sci. Rep. 2019, 9, 8408. [Google Scholar]
- Song, H.Y.; Mao, W.M.; Duan, Z.H.; Que, Q.M.; Zhou, W.; Chen, X.Y.; Li, P. Selection and validation of reference genes for measuring gene expression in Toona ciliata under different experimental conditions by quantitative real-time PCR analysis. BMC Plant Biol. 2020, 20, 450. [Google Scholar]
- Zhang, J.C.; Xie, W.G.; Yu, X.X.; Zhang, Z.Y.; Zhao, Y.Q.; Wang, N.; Wang, Y.R. Selection of suitable reference genes for rt-qpcr gene expression analysis in siberian wild rye (Elymus sibiricus) under different experimental conditions. Genes 2019, 10, 451. [Google Scholar]
- Zhang, Y.X.; Han, X.J.; Chen, S.S.; Zheng, L.; He, X.L.; Liu, M.Y.; Qiao, G.R.; Wang, Y.; Zhuo, R.Y. Selection of suitable reference genes for quantitative real-time PCR gene expression analysis in Salix matsudana under different abiotic stresses. Sci. Rep. 2017, 7, 40290. [Google Scholar]
- Ye, J.; Jin, C.-F.; Li, N.; Liu, M.-H.; Fei, Z.-X.; Dong, L.-Z.; Li, L.; Li, Z.-Q. Selection of suitable reference genes for qRT-PCR normalisation under different experimental conditions in Eucommia ulmoides Oliv. Sci. Rep. 2018, 8, 15043. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar]
- Zhong, M.Y.; Yang, X.Y.; Hu, Y.Y.; Huang, L.K.; Peng, Y.; Li, Z.; Liu, Q.X.; Wang, X.; Zhang, X.Q.; Nie, G. Identification of candidate reference genes for quantitative RT-PCR in Miscanthus sinensis subjected to various abiotic stresses. Mol. Biol. Rep. 2020, 47, 2913–2927. [Google Scholar]
- Kozera, B.; Rapacz, M. Reference genes in real-time PCR. J. Appl. Genet. 2013, 54, 391–406. [Google Scholar]
- Hruz, T.; Wyss, M.; Docquier, M.; Pfaffl, M.W.; Masanetz, S.; Borghi, L.; Verbrugghe, P.; Kalaydjieva, L.; Bleuler, S.; Laule, O. RefGenes: Identification of reliable and condition specific reference genes for RT-qPCR data normalization. BMC Genom. 2011, 12, 156. [Google Scholar]
- Joseph, J.T.; Poolakkalody, N.J.; Shah, J.M. Plant reference genes for development and stress response studies. J. Biosci. 2018, 43, 173–187. [Google Scholar]
- Xu, W.; Dong, Y.N.; Yu, Y.C.; Xing, Y.X.; Li, X.W.; Zhang, X.; Hou, X.J.; Sun, X.L. Identification and evaluation of reliable reference genes for quantitative real-time PCR analysis in tea plants under differential biotic stresses. Sci. Rep. 2020, 10, 2429. [Google Scholar]
- Zeng, A.; Xu, Y.; Song, L.; Li, J.; Yan, J. Validation of suitable reference genes for qRT-PCR in cabbage (Brassica oleracea L.) under different abiotic stress experimental conditions. J. Plant Biochem. Biotechnol. 2021, 30, 184–195. [Google Scholar]
- Wan, Q.; Chen, S.; Shan, Z.; Yang, Z.; Chen, L.; Zhang, C.; Yuan, S.; Hao, Q.; Zhang, X.; Qiu, D. Stability evaluation of reference genes for gene expression analysis by RT-qPCR in soybean under different conditions. PLoS ONE 2017, 12, e0189405. [Google Scholar]
- Wang, Z.L.; Xu, J.Y.; Liu, Y.H.; Chen, J.Y.; Lin, H.F.; Huang, Y.L.; Bian, X.H.; Zhao, Y.C. Selection and validation of appropriate reference genes for real-time quantitative PCR analysis in Momordica charantia. Phytochemistry 2019, 164, 1–11. [Google Scholar]
- Dheda, K.; Huggett, J.; Chang, J.; Kim, L.; Bustin, S.; Johnson, M.; Rook, G.; Zumla, A. The implications of using an inappropriate reference gene for real-time reverse transcription PCR data normalization. Anal. Biochem. 2005, 344, 141–143. [Google Scholar]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar]
- Xie, F.L.; Xiao, P.; Chen, D.L.; Xu, L.; Zhang, B.H. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar]
- Chen, Y.; Hu, B.; Tan, Z.; Liu, J.; Yang, Z.; Li, Z.; Huang, B. Selection of reference genes for quantitative real-time PCR normalization in creeping bentgrass involved in four abiotic stresses. Plant Cell Rep. 2015, 34, 1825–1834. [Google Scholar]
- Wong, M.L.; Medrano, J.F. Real-time PCR for mRNA quantitation. Biotechniques 2005, 39, 75–85. [Google Scholar]
- Purev, M.; Kim, Y.-J.; Kim, M.K.; Pulla, R.K.; Yang, D.-C. Isolation of a novel catalase (Cat1) gene from Panax ginseng and analysis of the response of this gene to various stresses. Plant Physiol. Biochem. 2010, 48, 451–460. [Google Scholar]
- Guénin, S.; Mauriat, M.; Pelloux, J.; Van Wuytswinkel, O.; Bellini, C.; Gutierrez, L. Normalization of qRT-PCR data: The necessity of adopting a systematic, experimental conditions-specific, validation of references. J. Exp. Bot. 2009, 60, 487–493. [Google Scholar]
- Gao, G.; Tang, Z.L.; Deng, J.B.; Gou, X.m.; Wang, Q.; Zhang, Y.; Ding, C.B.; Zhang, L.; Zhou, Y.H.; Yang, R.W. Phylogenetic relationships and Y genome origin in Kengyilia (Triticeae: Poaceae) based on single copy gene DMC1. Biologia 2016, 71, 298–304. [Google Scholar]
- Zeng, J.; Cao, G.; Liu, J.; Zhang, H.-Q.; Zhou, Y.-H. C-banding analysis of eight species of Kengyilia (Poaceae: Triticeae). J. Appl. Genet. 2008, 49, 11–21. [Google Scholar]
- Zhang, L.; Zheng, Y.L.; Wei, Y.M.; Liu, S.G.; Zhou, Y.H. The genetic diversity and similarities among Kengyilia species based on random amplified microsatellite polymorphism (RAMP). Genet. Resour. Crop Evol. 2005, 52, 1011–1017. [Google Scholar]
- Gordon, S.P.; Tseng, E.; Salamov, A.; Zhang, J.; Meng, X.; Zhao, Z.; Kang, D.; Underwood, J.; Grigoriev, I.V.; Figueroa, M. Widespread Polycistronic Transcripts in Fungi Revealed by Single-Molecule mRNA Sequencing. PLoS ONE 2015, 10, e0132628. [Google Scholar]
- Thomas, S.; Underwood, J.G.; Tseng, E.; Holloway, A.K.; Bench to Basinet CvDC Informatics Subcommittee. Long-Read Sequencing of Chicken Transcripts and Identification of New Transcript Isoforms. PLoS ONE 2014, 9, e94650. [Google Scholar]
- Ma, R.; Xu, S.; Zhao, Y.; Xia, B.; Wang, R. Selection and validation of appropriate reference genes for quantitative real-time PCR analysis of gene expression in Lycoris aurea. Front. Plant Sci. 2016, 7, 536. [Google Scholar]
- Pu, Q.; Li, Z.; Nie, G.; Zhou, J.; Liu, L.; Peng, Y. Selection and validation of reference genes for quantitative real-time PCR in white clover (Trifolium repens L.) involved in five abiotic stresses. Plants 2020, 9, 996. [Google Scholar]
- Zhu, P.H.; Ma, Y.Y.; Zhu, L.Z.; Chen, Y.; Li, R.; Ji, K.S. Selection of Suitable Reference Genes in Pinus massoniana Lamb. Under Different Abiotic Stresses for qPCR Normalization. Forests 2019, 10, 632. [Google Scholar]
- Zhou, H.J.; Costaguta, G.; Payne, G.S. Clathrin Adaptor Complex-interacting Protein Irc6 Functions through the Conserved C-Terminal Domain. Sci. Rep. 2019, 9, 4436. [Google Scholar]
- Kong, Q.; Yuan, J.; Gao, L.; Zhao, S.; Jiang, W.; Huang, Y.; Bie, Z. Identification of suitable reference genes for gene expression normalization in qRT-PCR analysis in watermelon. PLoS ONE 2014, 9, e90612. [Google Scholar]
- Migocka, M.; Papierniak, A. Identification of suitable reference genes for studying gene expression in cucumber plants subjected to abiotic stress and growth regulators. Mol. Breed. 2011, 28, 343–357. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Primer Sequence F/R (5′–3′) | Amplicon Length (bp) | Tm (°C) | Efficiency (%) | R2 |
|---|---|---|---|---|---|
| EF1A | F: TGATATGCGCCCTGTTGATGT | 128 | 59.2 | 93.9 | 0.995 |
| R: GCAGCCTACAGATAACATTCCA | |||||
| GAPDH | F: CTGTTCTCAAACCCCTCCGT | 85 | 60.2 | 93.1 | 0.985 |
| R: GATCCGGCCGAAACCATTGA | |||||
| ACT1 | F: CCCAAGGCCAATCGTGAGAA | 97 | 60.3 | 106.5 | 0.999 |
| R: CATACAGCGAGAGGACAGCC | |||||
| UBI | F: AACTTCAAAGGCGCAGATTCG | 165 | 59.5 | 93.9 | 0.994 |
| R: TGATAGTCTTGCCTGTGAGGG | |||||
| TUBB3 | F: GGGCATGGATGAGATGGAGTT | 143 | 61.3 | 104.1 | 0.995 |
| R: GTGGCTTATGCAGCACCTCCT | |||||
| TIPRL | F: TGAACGAAGACACCATGCAAAC | 81 | 59.8 | 99.5 | 0.996 |
| R: CAAGGTCGATCCGGTCATCA | |||||
| CACS | F: AAATGGCGTGGGCTCCTTATT | 125 | 60.2 | 100.3 | 0.998 |
| R: TCTGATCTGCCCTCTGCTAGT | |||||
| PPP2R1B | F: GCTCTGATCCCGTCAGTTGT | 131 | 59.9 | 99.5 | 0.998 |
| R: TGATGGAGTTCAGGCGCAAT | |||||
| TUBA1A | F: TCCTTGTGCCGCCTATCTTG | 89 | 59.9 | 99.7 | 0.998 |
| R: AACCCAACACCCAGACACAA | |||||
| EIF4A1 | F: GTGACCCGTGAAGATGAGAGG | 189 | 59.7 | 99.7 | 0.992 |
| R: CCCTCCCCACAGACAAGAAA | |||||
| CYPA3 | F: AAGTTGGCGTGAGTCGTGTT | 91 | 60.2 | 99.1 | 0.999 |
| R: CAGTCCACCTGAAACCCTCC | |||||
| TCTP | F: TGCTCTGCTCTATGGTGTTCA | 152 | 59.4 | 101.1 | 0.991 |
| R: CGAGGCACTGACCAAAACAC | |||||
| ABCG11L | F: CTACCGCCTGCTGTTCTTCA | 197 | 60.2 | 92.4 | 0.999 |
| R: GCTACCCAGCAACCCAGTTTA | |||||
| FBXO6L | F: ACGCAGAGACAGAAACCGAG | 151 | 60.1 | 91.1 | 0.995 |
| R: GCAAACAGTGCGGAAACGAA | |||||
| CAT1 | F: GATGAGTCCTCGATGGCGTG | 84 | 60.2 | 99.2 | 0.999 |
| R: CTTTGCCGATAAGAGGGGAGAA |
| Rank | ABA | Cold | Heat | Salt | Drought | All Samples |
|---|---|---|---|---|---|---|
| 1 | EF1A (1.22 ± 0.31) | TIPRL (0.80 ± 0.23) | FBXO6L (0.95 ± 0.25) | FBXO6L (1.34 ± 0.36) | PPP2R1B (1.29 ± 0.34) | TIPRL (2.12 ± 0.61) |
| 2 | TIPRL (1.51 ± 0.44) | CACS (1.07 ± 0.27) | TIPRL (1.45 ± 0.42) | GAPDH (1.68 ± 0.40) | FBXO6L (1.42 ± 0.40) | EF1A (2.81 ± 0.68) |
| 3 | CACS (1.76 ± 0.49) | CYPA3 (1.17 ± 0.24) | GAPDH (2.01 ± 0.48) | TIPRL (1.69 ± 0.47) | TUBA1A (1.56 ± 0.43) | UBI (2.89 ± 0.86) |
| 4 | TCTP (1.94 ± 0.43) | TCTP (1.22 ± 0.27) | TCTP (2.07 ± 0.44) | UBI (1.89 ± 0.56) | CACS (1.63 ± 0.45) | CYPA3 (2.9 ± 0.61) |
| 5 | CYPA3 (2.01 ± 0.44) | ACT1 (1.30 ± 0.36) | EF1A (2.22 ± 0.53) | PPP2R1B (1.90 ± 0.49) | GAPDH (1.65 ± 0.42) | CACS (3.32 ± 0.9) |
| 6 | FBXO6L (2.03 ± 0.55) | TUBB3 (1.38 ± 0.35) | CACS (2.29 ± 0.61) | CYPA3 (2.07 ± 0.43) | ACT1 (1.78 ± 0.54) | TCTP (3.41 ± 0.76) |
| 7 | ACT1 (2.10 ± 0.64) | FBXO6L (1.50 ± 0.38) | UBI (2.32 ± 0.67) | ACT1 (2.09 ± 0.59) | EF1A (1.82 ± 0.44) | FBXO6L (3.56 ± 0.95) |
| 8 | UBI (2.32 ± 0.69) | PPP2R1B (1.67 ± 0.40) | CYPA3 (2.43 ± 0.51) | TCTP (2.13 ± 0.48) | TIPRL (1.82 ± 0.54) | ACT1 (3.61 ± 1.05) |
| 9 | PPP2R1B (2.40 ± 0.65) | EF1A (2.04 ± 0.48) | ACT1 (2.65 ± 0.77) | CACS (2.15 ± 0.59) | TCTP (2.00 ± 0.47) | GAPDH (3.63 ± 0.87) |
| 10 | TUBB3 (2.55 ± 0.70) | GAPDH (2.09 ± 0.48) | PPP2R1B (2.78 ± 0.70) | TUBA1A (2.35 ± 0.65) | TUBB3 (2.05 ± 0.58) | PPP2R1B (4.06 ± 1.04) |
| 11 | TUBA1A (2.65 ± 0.71) | EIF4A1 (2.20 ± 0.48) | EIF4A1 (3.11 ± 0.75) | EF1A (2.66 ± 0.64) | CYPA3 (2.12 ± 0.44) | TUBA1A (4.23 ± 1.13) |
| 12 | EIF4A1 (2.72 ± 0.69) | UBI (2.89 ± 0.83) | TUBA1A (3.27 ± 0.89) | EIF4A1 (3.18 ± 0.75) | UBI (2.38 ± 0.73) | TUBB3 (4.84 ± 1.34) |
| 13 | GAPDH (3.03 ± 0.74) | ABCG11L (3.24 ± 0.75) | ABCG11L (4.18 ± 1.03) | ABCG11L (3.47 ± 0.89) | EIF4A1 (2.77 ± 0.67) | EIF4A1 (5.02 ± 1.2) |
| 14 | ABCG11L (4.35 ± 1.15) | TUBA1A (4.89 ± 1.20) | TUBB3 (4.77 ± 1.37) | TUBB3 (4.20 ± 1.19) | ABCG11L (3.23 ± 0.85) | ABCG11L (5.35 ± 1.35) |
| Rank | ABA | Cold | Heat | Salt | Drought | All Samples |
|---|---|---|---|---|---|---|
| 1 | TCTP | CACS | CACS | TIPRL | CACS | CACS |
| 2 | TIPRL | TCTP | FBXO6L | CYPA3 | FBXO6L | PPP2R1B |
| 3 | CACS | PPP2R1B | TIPRL | FBXO6L | PPP2R1B | CYPA3 |
| 4 | EF1A | FBXO6L | TCTP | GAPDH | TCTP | EF1A |
| 5 | CYPA3 | CYPA3 | CYPA3 | TCTP | CYPA3 | GAPDH |
| 6 | PPP2R1B | ACT1 | PPP2R1B | CACS | EF1A | TIPRL |
| 7 | GAPDH | GAPDH | UBI | PPP2R1B | TIPRL | FBXO6L |
| 8 | EIF4A1 | TUBB3 | GAPDH | EF1A | GAPDH | TCTP |
| 9 | ACT1 | EIF4A1 | EF1A | UBI | EIF4A1 | UBI |
| 10 | UBI | TIPRL | ACT1 | ACT1 | ACT1 | ACT1 |
| 11 | FBXO6L | EF1A | EIF4A1 | EIF4A1 | TUBA1A | EIF4A1 |
| 12 | TUBB3 | ABCG11L | ABCG11L | TUBA1A | UBI | TUBA1A |
| 13 | TUBA1A | UBI | TUBA1A | ABCG11L | TUBB3 | ABCG11L |
| 14 | ABCG11L | TUBA1A | TUBB3 | TUBB3 | ABCG11L | TUBB3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Yang, J.; Wang, X.; Xiong, Y.; Xiong, Y.; Dong, Z.; Lei, X.; Yan, L.; Ma, X. Selection and Validation of Reference Genes for qRT-PCR Gene Expression Analysis in Kengyilia melanthera. Genes 2022, 13, 1445. https://doi.org/10.3390/genes13081445
Zhao J, Yang J, Wang X, Xiong Y, Xiong Y, Dong Z, Lei X, Yan L, Ma X. Selection and Validation of Reference Genes for qRT-PCR Gene Expression Analysis in Kengyilia melanthera. Genes. 2022; 13(8):1445. https://doi.org/10.3390/genes13081445
Chicago/Turabian StyleZhao, Junming, Jian Yang, Xiaoyun Wang, Yanli Xiong, Yi Xiong, Zhixiao Dong, Xiong Lei, Lijun Yan, and Xiao Ma. 2022. "Selection and Validation of Reference Genes for qRT-PCR Gene Expression Analysis in Kengyilia melanthera" Genes 13, no. 8: 1445. https://doi.org/10.3390/genes13081445
APA StyleZhao, J., Yang, J., Wang, X., Xiong, Y., Xiong, Y., Dong, Z., Lei, X., Yan, L., & Ma, X. (2022). Selection and Validation of Reference Genes for qRT-PCR Gene Expression Analysis in Kengyilia melanthera. Genes, 13(8), 1445. https://doi.org/10.3390/genes13081445