
Genome-Wide Analysis of the Protein Phosphatase 2C Genes in Tomato


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of SlPP2C Members
2.2. Phylogenetic Analysis
2.3. Chromosomal Location, Gene Structure, and Sequence Alignment
2.4. Cis-Element Prediction for PP2C Gene Promoter
2.5. Expression Analysis of SlPP2Cs of Tomato Tissue
2.6. Transcriptional Profiling of SlPP2C Genes in Tomato Infected with Ralstonia solanacearum
2.7. Bacterial Strain of Ralstonia solanacearum
2.8. Plant Material and Treatments
2.9. RNA Isolation and Real-Time PCR
3. Results
3.1. Identification of PP2C Genes in Tomato
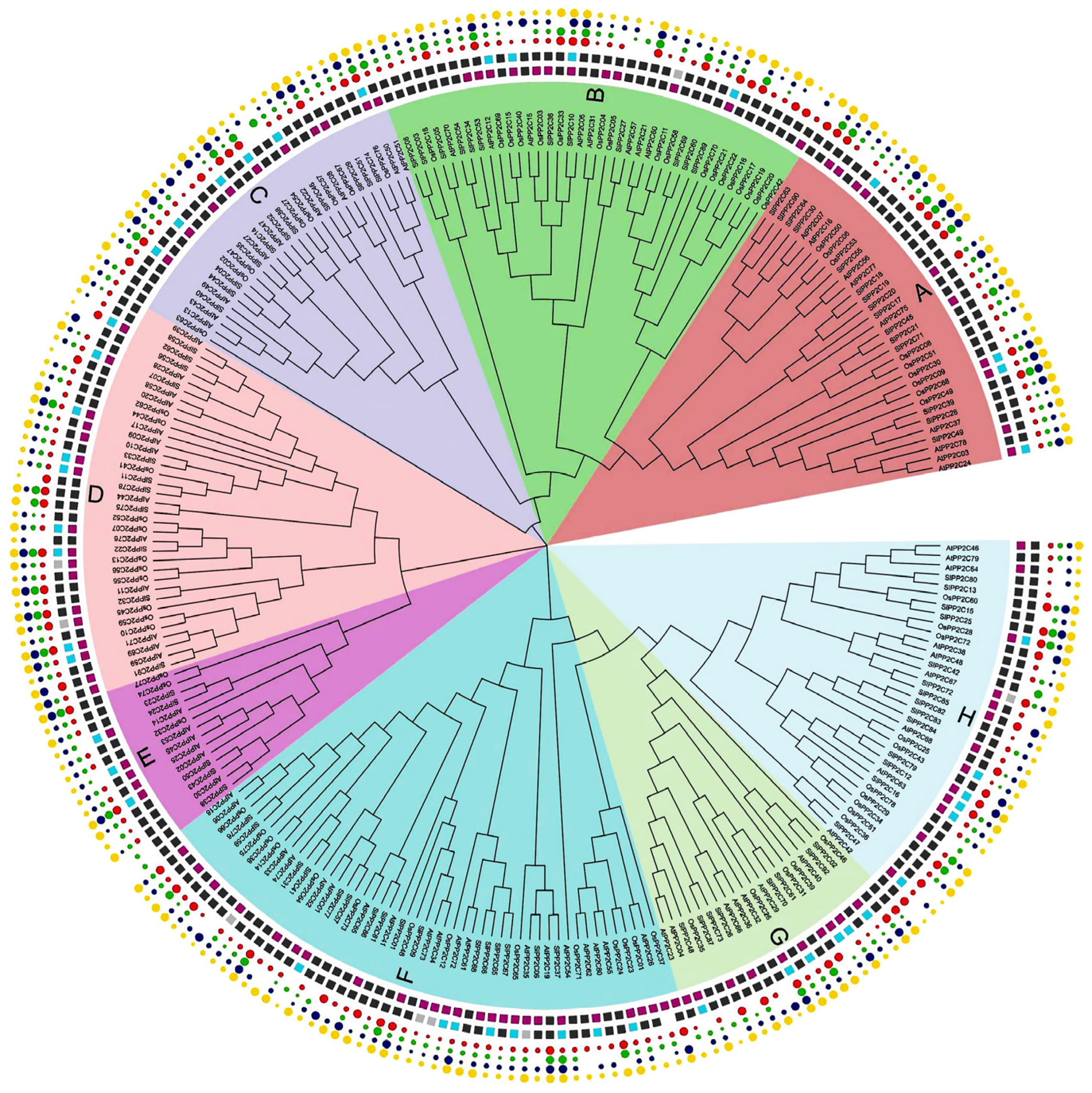
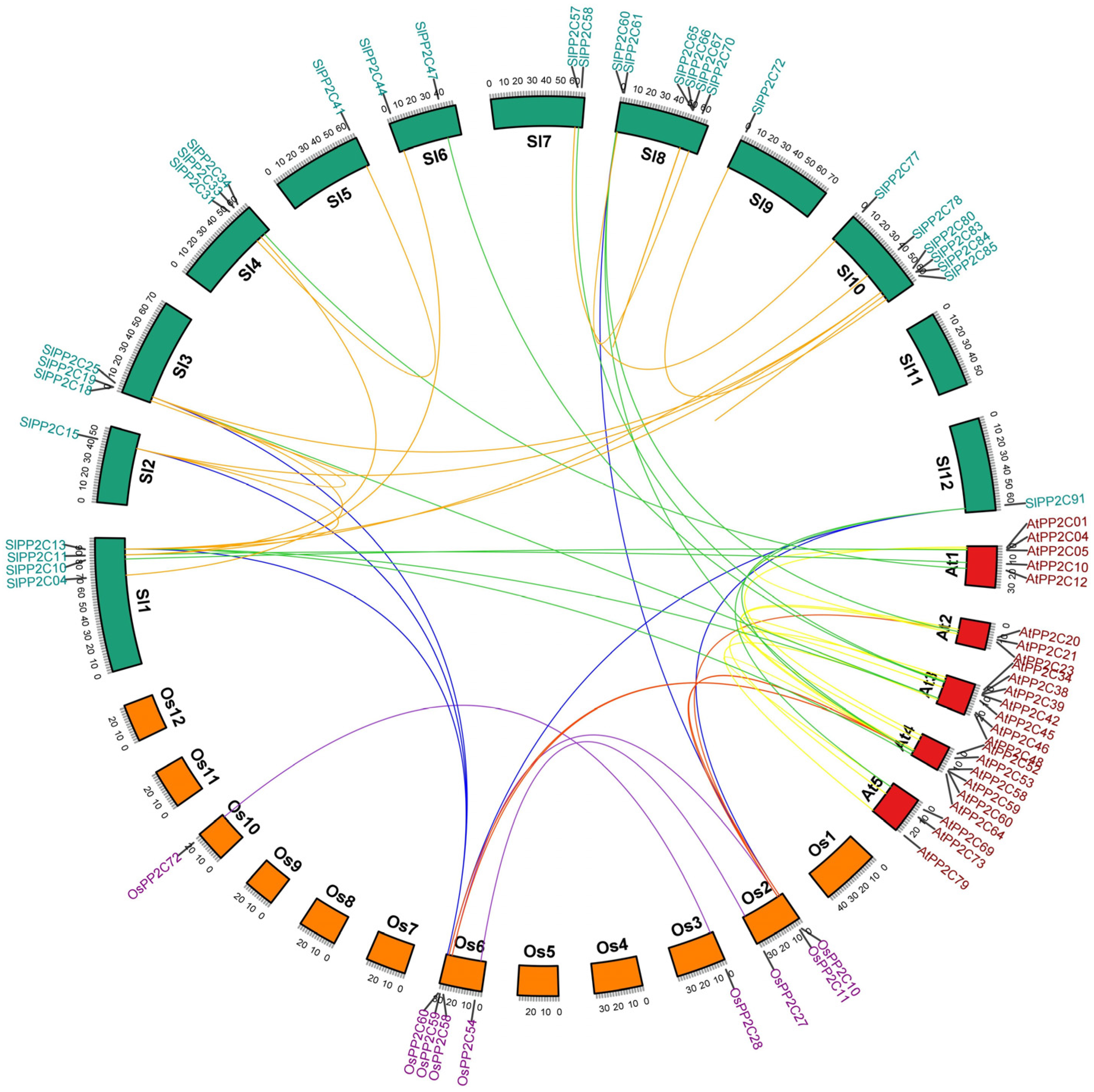
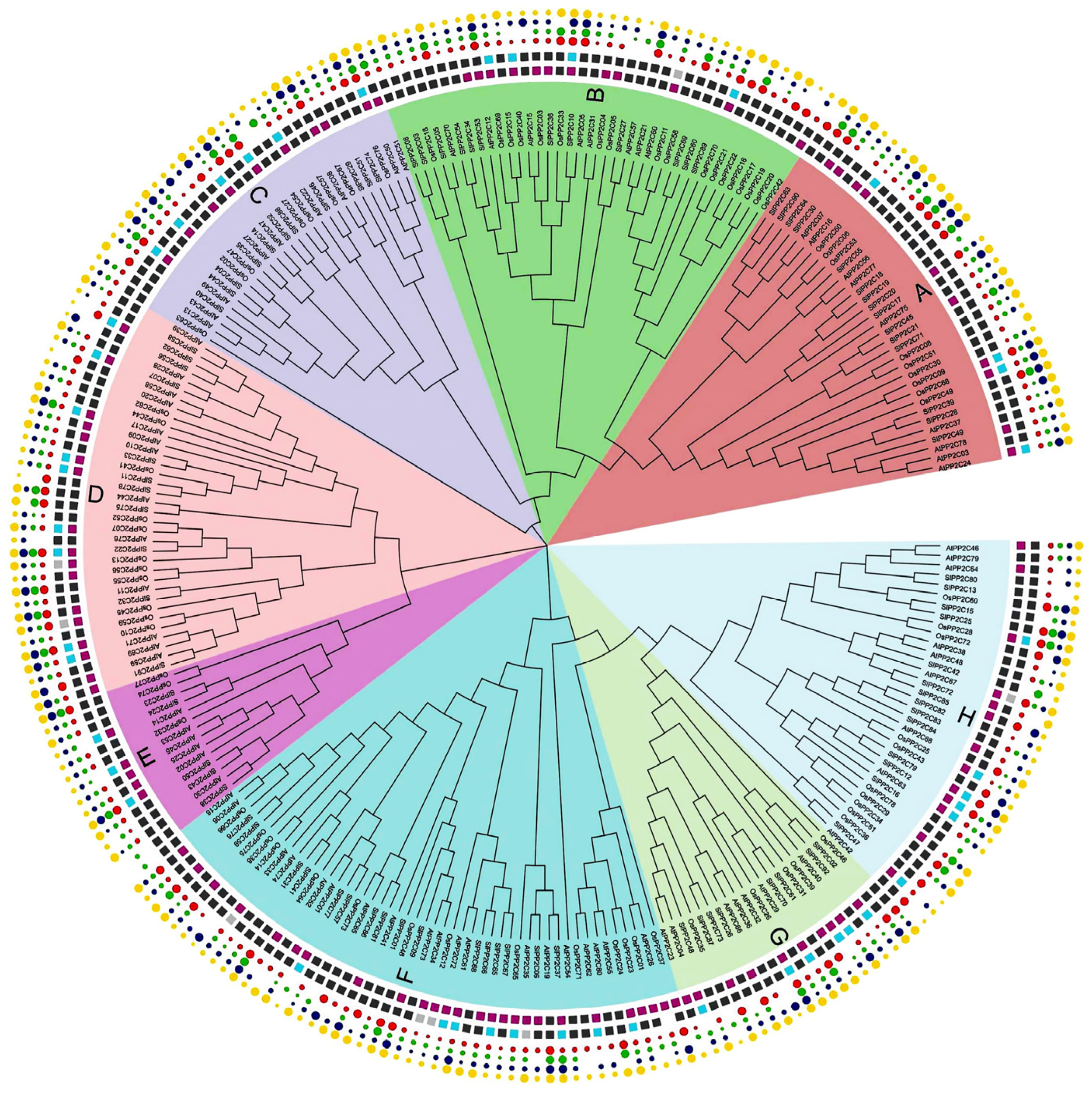
3.2. Phylogenetic and Comparative Synteny Analysis
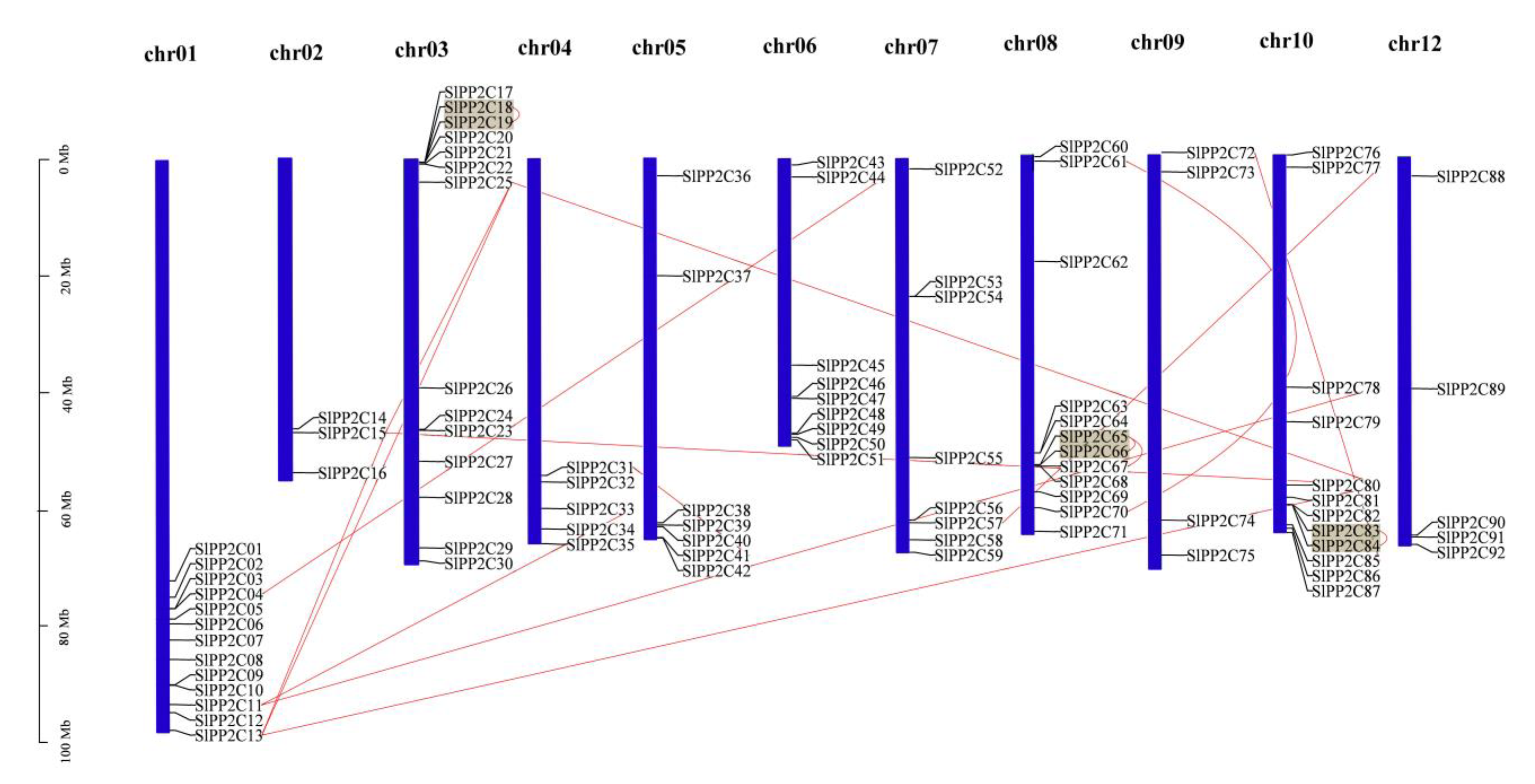
3.3. Chromosomal Localization and Duplication of SlPP2C Genes
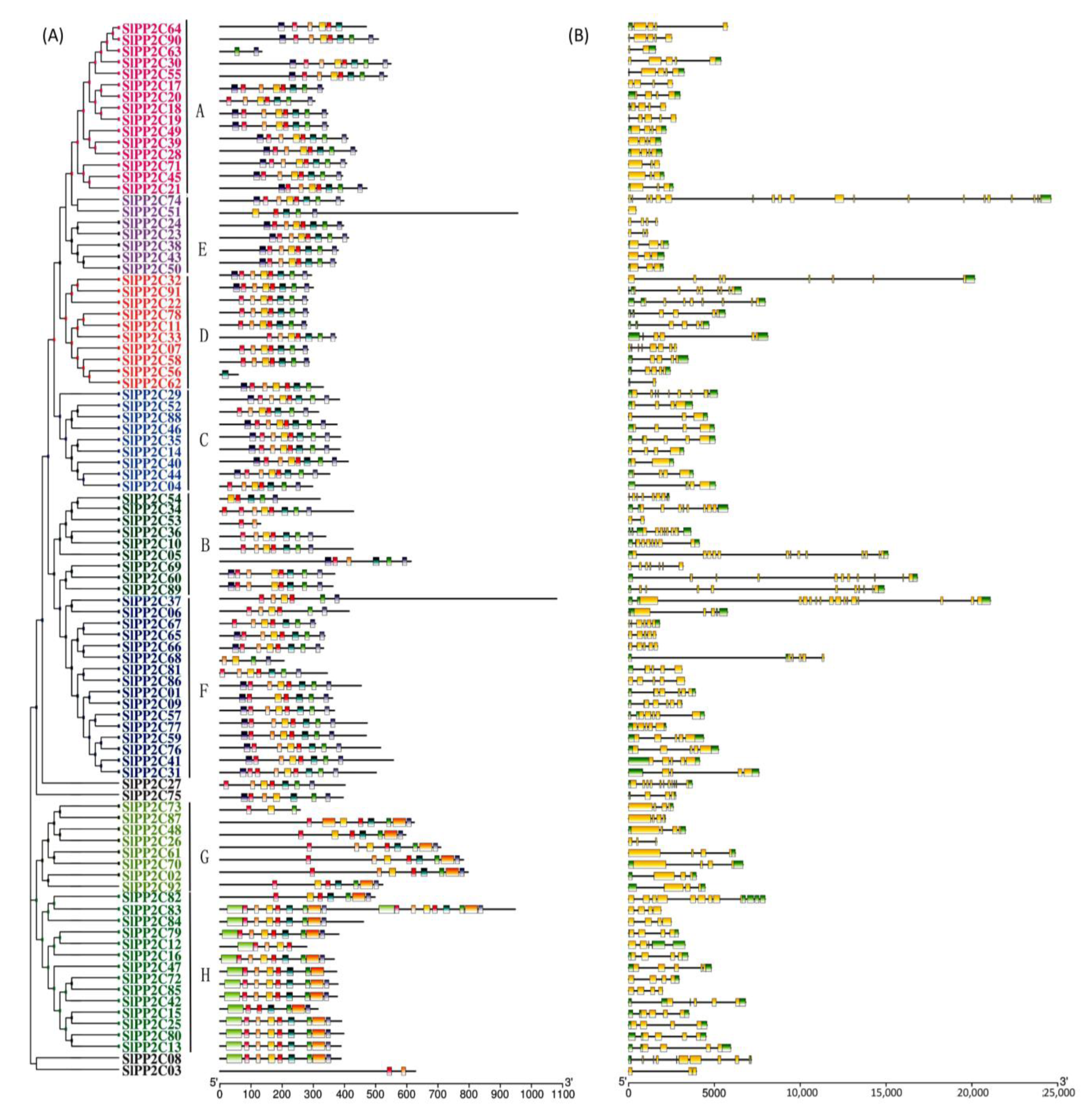
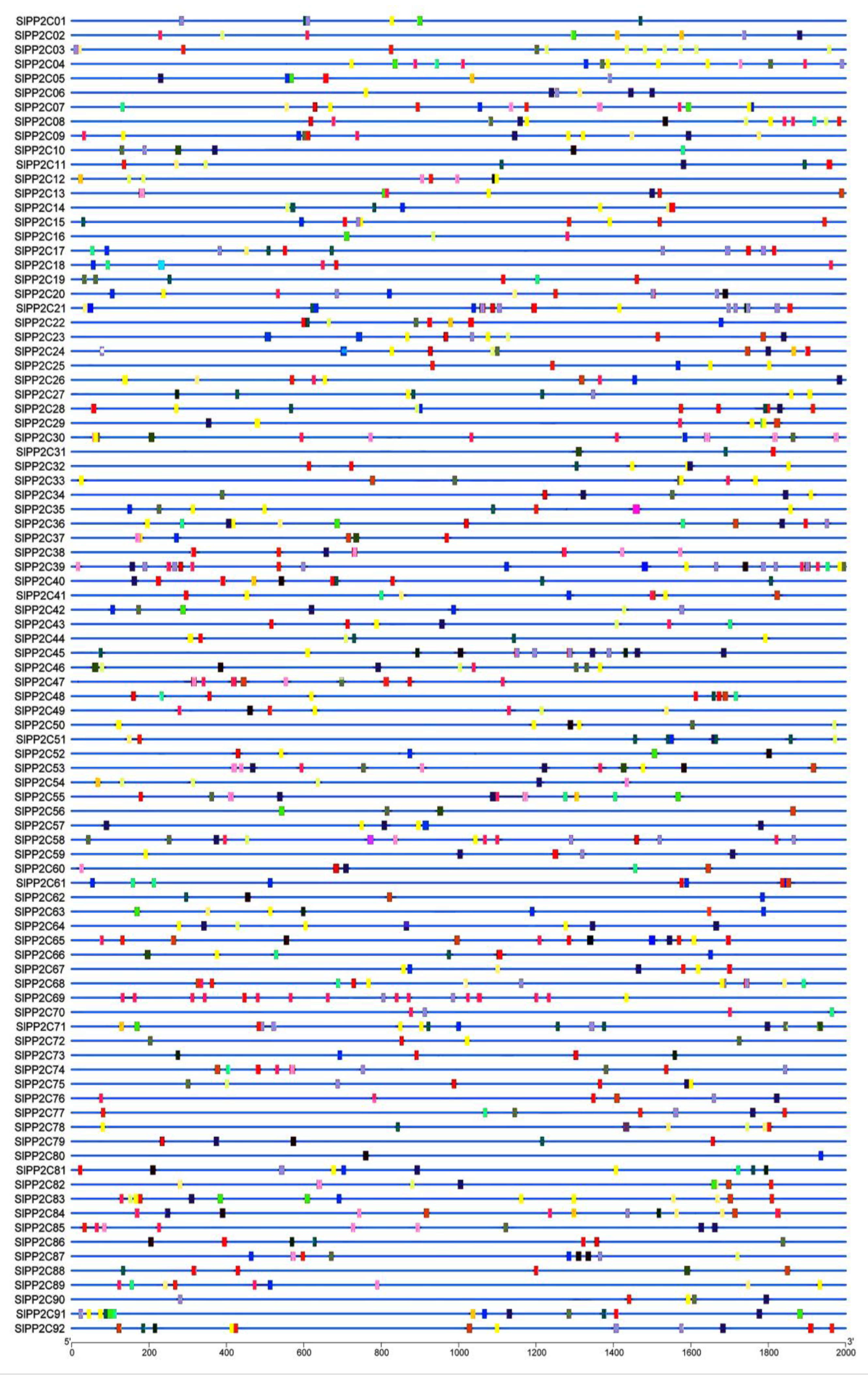
3.4. Conserved Motif and Gene Structure of SlPP2Cs
3.5. Cis-Elements in the Promoters of Tomato PP2C Genes
3.6. Expression of SlPP2C Genes in Different Tomato Tissues
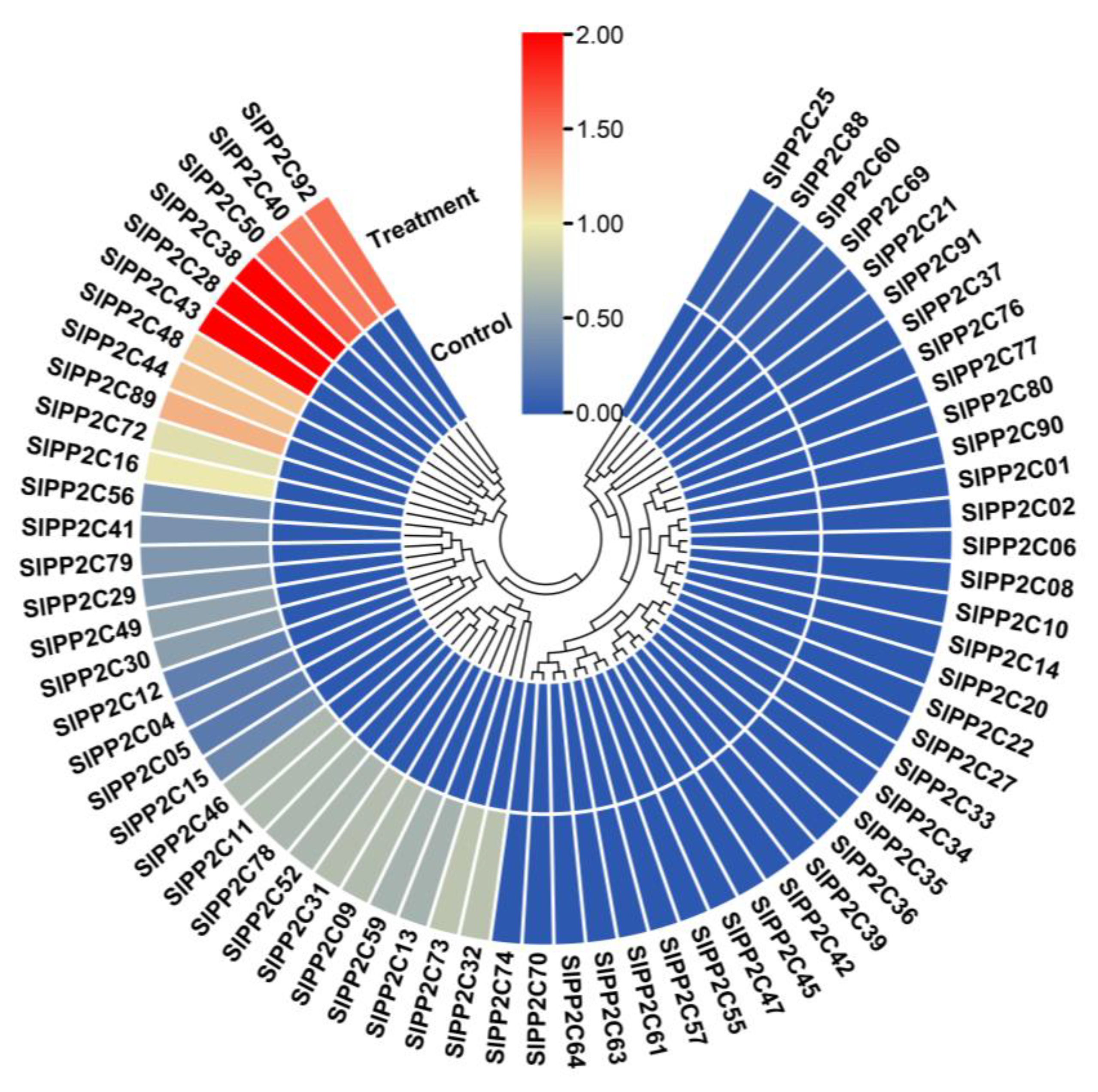
3.7. Transcriptional Profiling of SlPP2C Genes of Tomato Infected with Ralstonia solanacearum
3.8. Analysis of SlPP2C Gene Expression in Tomato under Ralstonia solanacearum Infection
4. Discussion
4.1. Evolution of the SlPP2C Gene Family
4.2. Expression of SlPP2Cs
4.3. Possible Function of SlPP2Cs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Long, L.; Gao, W.; Xu, L. GbMPK3, a mitogen-activated protein kinase from cotton, enhances drought and oxidative stress tolerance in tobacco. Plant Cell Tissue Organ Cult. 2014, 116, 153–162. [Google Scholar] [CrossRef]
- Sheng, L. Protein phosphatases and signaling cascades in higher plants. Trends Plant Sci. 1998, 3, 271–275. [Google Scholar]
- Mizoguchi, T.; Ichimura, K.; Shinozaki, K. Environmental stress response in plants: The role of mitogen-activated protein kinases. Trends Biotechnol. 1997, 15, 15–19. [Google Scholar] [CrossRef]
- Boudsocq, M.; Barbier-Brygoo, H.; Lauriere, C. Identification of nine sucrose nonfermenting 1-related protein kinases 2 activated by hyperosmotic and saline stresses in Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 41758–41766. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.Y.; Wu, W.H. AtCPK23 functions in Arabidopsis responses to drought and salt stresses. Plant Mol. Biol. 2007, 65, 511–518. [Google Scholar] [CrossRef]
- Franz, S.; Ehlert, B.; Liese, A.; Kurth, J.; Cazalé, A.C.; Romeis, T. Calcium-dependent protein kinase CPK21 functions in abiotic stress response in Arabidopsis thaliana. Mol. Plant 2011, 4, 83–96. [Google Scholar] [CrossRef]
- Wei, K.; Pan, S. Maize protein phosphatase gene family: Identification and molecular characterization. BMC Genom. 2014, 15, 773. [Google Scholar] [CrossRef] [Green Version]
- Kerk, D.; Templeton, G.; Moorhead, G.B. Evolutionary radiation pattern of novel protein phosphatases revealed by analysis of protein data from the completely sequenced genomes of humans, green algae, and higher plants. Plant Physiol. 2008, 146, 351–367. [Google Scholar] [CrossRef] [Green Version]
- Cohen, P. The structure and regulation of protein phosphatases. Annu. Rev. Biochem. 1989, 58, 453–508. [Google Scholar] [CrossRef]
- Singh, A.; Pandey, A.; Srivastava, A.K.; Tran, L.S.; Pandey, G.K. Plant protein phosphatases 2C: From genomic diversity to functional multiplicity and importance in stress management. Crit. Rev. Biotechnol. 2016, 36, 1023–1035. [Google Scholar] [CrossRef]
- Chae, L.; Pandey, G.K.; Luan, S.; Cheong, Y.H.; Kim, K. Protein kinases and phosphatases for stress signal transduction in plants. In Abiotic Stress Adaptation in Plants; Springer: Berlin/Heidelberg, Germany, 2010; pp. 123–163. [Google Scholar]
- Mackintosh, C.; Coggins, J.; Cohen, P. Plant protein phosphatases. subcellular distribution, detection of protein phosphatase 2C and identification of protein phosphatase 2A as the major quinate dehydrogenase phosphatase. Biochem. J. 1991, 273, 733–738. [Google Scholar] [CrossRef] [Green Version]
- Schweighofer, A.; Hirt, H.; Meskiene, I. Plant PP2C phosphatases: Emerging functions in stress signaling. Trends Plant Sci. 2004, 9, 236–243. [Google Scholar] [CrossRef]
- Cao, J.; Jiang, M.; Li, P.; Chu, Z. Genome-wide identification and evolutionary analyses of the PP2C gene family with their expression profiling in response to multiple stresses in Brachypodium distachyon. BMC Genom. 2016, 17, 175. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Giri, J.; Kapoor, S.; Tyagi, A.K.; Pandey, G.K. Protein phosphatase complement in rice: Genome-wide identification and transcriptional analysis under abiotic stress conditions and reproductive development. BMC Genom. 2010, 11, 435. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, H.; Kondo, S.; Tanaka, T.; Imamura, C.; Muramoto, N.; Hattori, E.; Ogawa, K.; Mitsukawa, N.; Ohto, C. Overexpression of a novel Arabidopsis PP2C isoform, AtPP2CF1, enhances plant biomass production by increasing inflorescence stem growth. J. Exp. Bot. 2014, 65, 5385–5400. [Google Scholar] [CrossRef] [Green Version]
- Manohar, M.; Wang, D.; Manosalva, P.M.; Choi, H.W.; Kombrink, E.; Klessig, D.F. Members of the abscisic acid co-receptor PP2C protein family mediate salicylic acid –abscisic acid crosstalk. Plant Direct 2017, 1, e00020. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, K.; Suzuki, N.; Kuwamura, M.; Nishikawa, Y.; Nakatani, M.; Ohtawa, H.; Takezawa, D.; Seki, M.; Tanaka, M.; Taji, T.; et al. Group A PP2Cs evolved in land plants as key regulators of intrinsic desiccation tolerance. Nat. Commun. 2013, 4, 2219. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, P.L.; Leube, M.P.; Grill, E. Molecular cloning in Arabidopsis thaliana of a new protein phosphatase 2C (PP2C) with homology to ABI1 and ABI2. Plant Mol. Biol. 1998, 38, 879–883. [Google Scholar] [CrossRef]
- Gosti, F.; Beaudoin, N.; Serizet, C.; Webb, A.A.; Vartanian, N.; Giraudat, J. ABI1 protein phosphatase 2C is a negative regulator of abscisic acid signaling. Plant Cell 1999, 11, 1897–1910. [Google Scholar] [CrossRef] [Green Version]
- Merlot, S.; Gosti, F.; Guerrier, D.; Vavasseur, A.; Giraudat, J. The ABI1 and ABI2 protein phosphatases 2C act in a negative feedback regulatory loop of the abscisic acid signalling pathway. Plant J. 2010, 25, 295–303. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Q.; Jiang, L.; Kai, W.; Liang, B.; Wang, J.; Du, Y.; Zhai, X.; Wang, J.; Zhang, Y.; et al. Suppressing type 2C protein phosphatases alters fruit ripening and the stress response in tomato. Plant Cell Physiol. 2018, 59, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Y.; Sun, X.; Gao, S.; Qin, F.; Dai, M. Deletion of an endoplasmic reticulum stress response element in a ZmPP2C-A gene facilitates drought tolerance of maize seedlings. Mol. Plant 2017, 10, 456–469. [Google Scholar] [CrossRef] [Green Version]
- Arshad, M.; Mattsson, J. A putative poplar PP2C-encoding gene negatively regulates drought and abscisic acid responses in transgenic Arabidopsis thaliana. Trees 2014, 28, 531–543. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, P.; Sun, L.; Li, Q.; Dai, S.; Sun, Y. Transcriptional regulation of PaPYLs, PaPP2Cs and PaSnRK2s during sweet cherry fruit development and in response to abscisic acid and auxin at onset of fruit ripening. Plant Growth Regul. 2015, 75, 455–464. [Google Scholar] [CrossRef]
- Zhang, F.; Wei, Q.; Shi, J.; Jin, X.; He, Y.; Zhang, Y.; Luo, Q.; Wang, Y.; Chang, J.; Yang, G.; et al. Brachypodium distachyon BdPP2CA6 interacts with BdPYLs and BdSnRK2 and positively regulates salt tolerance in transgenic Arabidopsis. Front. Plant Sci. 2017, 8, 264. [Google Scholar] [CrossRef] [Green Version]
- Schweighofer, A.; Kazanaviciute, V.; Scheikl, E.; Teige, M.; Doczi, R.; Hirt, H.; Schwanninger, M.; Kant, M.; Schuurink, R.; Mauch, F.; et al. The PP2C-type phosphatase AP2C1, which negatively regulates MPK4 and MPK6, modulates innate immunity, jasmonic acid, and ethylene levels in Arabidopsis. Plant Cell 2007, 19, 2213–2224. [Google Scholar] [CrossRef] [Green Version]
- Shubchynskyy, V.; Boniecka, J.; Schweighofer, A.; Simulis, J.; Kvederaviciute, K.; Stumpe, M.; Mauch, F.; Balazadeh, S.; Mueller-Roeber, B.; Boutrot, F.; et al. Protein phosphatase AP2C1 negatively regulates basal resistance and defense responses to Pseudomonas syringae. J. Exp. Bot. 2017, 68, 1169–1183. [Google Scholar]
- Sidonskaya, E.; Schweighofer, A.; Shubchynskyy, V.; Kammerhofer, N.; Hofmann, J.; Wieczorek, K.; Meskiene, I. Plant resistance against the parasitic nematode Heterodera schachtii is mediated by MPK3 and MPK6 kinases, which are controlled by the MAPK phosphatase AP2C1 in Arabidopsis. J. Exp. Bot. 2016, 67, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Umbrasaite, J.; Schweighofer, A.; Kazanaviciute, V.; Magyar, Z.; Ayatollahi, Z.; Unterwurzacher, V.; Choopayak, C.; Boniecka, J.; Murray, J.A.; Bogre, L.; et al. MAPK phosphatase AP2C3 induces ectopic proliferation of epidermal cells leading to stomata development in Arabidopsis. PLoS ONE 2010, 5, 15357. [Google Scholar] [CrossRef] [Green Version]
- Tovar-Mendez, A.; Miernyk, J.A.; Hoyos, E.; Randall, D.D. A functional genomic analysis of Arabidopsis thaliana PP2C clade D. Protoplasma 2014, 251, 265–271. [Google Scholar] [CrossRef]
- Min, W.L.; Jelenska, J.; Greenberg, J.T. Arabidopsis proteins important for modulating defense responses to Pseudomonas syringae that secrete HopW1-1. Plant J. 2010, 54, 452–465. [Google Scholar]
- Xue, T.; Wang, D.; Zhang, S. Genome-wide and expression analysis of protein phosphatase 2C in rice and Arabidopsis. BMC Genom. 2008, 9, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Han, J.; Wang, E.; Xiao, J.; Hu, R.; Yang, G.; He, G. Genome-wide identification and homoeologous expression analysis of PP2C genes in wheat (Triticum aestivum L.). Front. Genet. 2019, 10, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, M.S.; Khan, N.; Pervaiz, T.; Zhongjie, L.; Nasim, M.; Jogaiah, S.; Mushtaq, N.; Jiu, S.; Jinggui, F. Genome-wide identification, evolution, and molecular characterization of the PP2C gene family in woodland strawberry. Gene 2019, 702, 27–35. [Google Scholar] [CrossRef]
- Khan, N.; Ke, H.; Hu, C.M.; Naseri, E.; Haider, M.S.; Ayaz, A.; Amjad, K.W.; Wang, J.; Hou, X. Genome-wide identification, evolution, and transcriptional profiling of PP2C gene family in Brassica rapa. BioMed Res. Int. 2019, 2019, 2965035. [Google Scholar] [CrossRef] [Green Version]
- Hosmani, P.S.; Flores-Gonzalez, M.; van de Geest, H.; Maumus, F.; Bakker, L.V.; Schijlen, E.; van Haarst, J.; Cordewener, J.; Sanchez-Perez, G.; Peters, S.; et al. An improved de novo assembly and annotation of the tomato reference genome using single-molecule sequencing, Hi-C proximity ligation and optical maps. bioRxiv 2019, 767764. [Google Scholar] [CrossRef]
- Wang, Y.F.; Liao, Y.Q.; Wang, Y.P. Genome-wide identification and expression analysis of StPP2C gene family in response to multiple stresses in potato (Solanum tuberosum L.). J. Integr. Agric. 2020, 19, 1609–1624. [Google Scholar] [CrossRef]
- Jaina, M.; Sara, C.; Lowri, W. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, 412–419. [Google Scholar]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 1, 200–204. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Pozo, N.; Menda, N.; Edwards, J.D.; Saha, S.; Tecle, I.Y.; Strickler, S.R.; Bombarely, A.; Fisher-York, T.; Pujar, A.; Foerster, H.; et al. The Sol Genomics Network (SGN)—From genotype to phenotype to breeding. Nucleic Acids Res. 2015, 43, 1036–1041. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Borkm, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, 458–460. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2019, 48, 265–268. [Google Scholar] [CrossRef] [Green Version]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the swiss bioinformatics resource portal, as designed by its users. Nucleic Acids Res. 2021, 49, 216–227. [Google Scholar] [CrossRef]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef] [Green Version]
- Hung, J.H.; Weng, Z. Sequence alignment andhomology search with BLAST and ClustalW. Cold Spring Harb. Protoc. 2016, 11, pdb–rot093088. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 6, 1547–1549. [Google Scholar] [CrossRef]
- Gu, Z.; Cavalcanti, A.; Chen, F.C.; Bouman, P.; Li, W.H. Extent of gene duplication in the genomes of drosophila, nematode, and yeast. Mol. Biol. Evol. 2002, 3, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z. KaKs_calculator 3.0: Calculating selective pressure on coding and non-coding sequences. Genom. Proteom. Bioinform. 2022. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.P.; Lin, J.J.; Li, W.H. Positional distribution of transcription factor binding sites in Arabidopsis thaliana. Sci. Rep. 2016, 6, 25164. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Fei, Z.; Je-Gun, J.; Tang, X. Tomato Functional Genomics Database: A comprehensive resource and analysis package for tomato functional genomics. Nucleic Acids Res. 2011, 39, 1156–1163. [Google Scholar] [CrossRef] [Green Version]
- Kelman, A. The relationship of the pathogenicity of Pseudomonas solanacearum to colony appearance on a tetrazolium medium. Phytopathology 1954, 44, 693–695. [Google Scholar]
- Lu, K.; Li, T.; He, J.; Chang, W.; Zhang, R.; Liu, M. qPrimerDB: A thermodynamics-based gene-specific qPCR primer database for 147 organisms. Nucleic Acids Res. 2018, 46, 1229–1236. [Google Scholar] [CrossRef]
- Ding, X.; Li, J.; Pan, Y.; Zhang, Y.; Ni, L.; Wang, Y.; Zhang, X. Genome-wide identification and expression analysis of the UGLcAE gene family in tomato. Int. J. Mol. Sci. 2018, 19, 1583. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Le Berre, M.; Gerlach, J.Q.; Dziembała, I.; Kilcoyne, M. Calculating half maximal inhibitory concentration (IC50) values from glycomics microarray data using GraphPad Prism. Methods Mol. Biol. 2022, 2460, 89–111. [Google Scholar]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Blanc, G.; Wolfe, K.H. Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes. Plant Cell 2004, 16, 1667–1678. [Google Scholar] [CrossRef] [Green Version]
- Cheung, J.; Estivill, X.; Khaja, R.; MacDonald, J.R.; Lau, K.; Tsui, L.C.; Scherer, S.W. Genome-wide detection of segmental duplications and potential assembly errors in the human genome sequence. Genome Biol. 2003, 4, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denny, T.P. Ralstonia solanacearum—A plant pathogen in touch with its host. Trends Microbiol. 2000, 8, 486–489. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [Green Version]
- Haider, M.S.; Kurjogi, M.M.; Khalil-Ur-Rehman, M.; Fiaz, M.; Pervaiz, T.; Jiu, S.; Haifeng, J.; Chen, W.; Fang, J. Grapevine immune signaling network in response to drought stress as revealed by transcriptomic analysis. Plant Physiol. Biochem. 2017, 121, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Kerk, D.; Bulgrien, J.; Smith, D.W.; Barsam, B.; Veretnik, S.; Gribskov, M. The complement of protein phosphatase catalytic subunits encoded in the genome of Arabidopsis. Plant Physiol. 2002, 129, 908–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djami-Tchatchou, A.T.; Maake, M.P.; Piater, L.A.; Dubery, I.A. Isonitrosoacetophenone drives transcriptional reprogramming in Nicotiana tabacum cells in support of innate immunity and defense. PLoS ONE 2015, 10, e0117377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Park, M.; Yeom, S.I. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef]
- Yang, Q.; Liu, K.; Niu, X.; Wang, Q.; Wan, Y.; Yang, F.; Li, G.; Wang, Y.; Wang, R. Genome-wide identification of PP2C genes and their expression profiling in response to drought and cold stresses in Medicago truncatula. Sci. Rep. 2018, 8, 12841. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, S.; Grill, E.; Meskiene, I.; Schweighofer, A. Type 2C protein phosphatases in plants. FEBS J. 2013, 280, 681–693. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [Green Version]
- Juretic, N.; Hoen, D.R.; Huynh, M.L.; Harrison, P.M.; Bureau, T.E. The evolutionary fate of MULE-mediated duplications of host gene fragments in rice. Genome Res. 2005, 15, 1292–1297. [Google Scholar] [CrossRef] [Green Version]
- Pervez, M.A.; Ayub, C.M.; Khan, H.A.; Shahid, M.A.; Ashraf, I. Effect of drought stress on growth, yield and seed quality of tomato (Lycopersicon esculentum L.). Pak. J. Agric. Sci. 2009, 46, 174–178. [Google Scholar]
- Zhang, P.; Senge, M.; Dai, Y. Effects of salinity stress at different growth stages on tomato growth, yield, and water-use efficiency. Commun. Soil Sci. Plant Anal. 2017, 48, 624–634. [Google Scholar] [CrossRef]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef]
- Rubio, S.; Rodrigues, A.; Saez, A.; Dizon, M.B.; Galle, A.; Kim, T.; Santiago, J.; Flexas, J.; Schroeder, J.I.; Rodriguez, P.L. Triple loss of function of protein phosphatases type 2C leads to partial constitutive response to endogenous abscisic acid. Plant Physiol. 2009, 150, 1345–1355. [Google Scholar] [CrossRef] [Green Version]
- Saez, A.; Apostolova, N.; Gonzalez-Guzman, M.; Gonzalez-Garcia, M.P.; Nicolas, C.; Lorenzo, O.; Rodriguez, P.L. Gain-of-function and loss-of-function phenotypes of the protein phosphatase 2C HAB1 reveal its role as a negative regulator of abscisic acid signalling. Plant J. 2004, 37, 354–369. [Google Scholar] [CrossRef]
- Sun, H.L.; Wang, X.J.; Ding, W.H.; Zhu, S.Y.; Zhao, R.; Zhang, Y.X.; Xin, Q.; Wang, X.F.; Zhang, D.P. Identification of an important site for function of the type2C protein phosphatase ABI2 in abscisic acid signalling in Arabidopsis. J. Exp. Bot. 2011, 62, 5713–5725. [Google Scholar] [CrossRef] [Green Version]
- Allen, G.J.; Kuchitsu, K.; Chu, S.P.; Murata, Y.; Schroeder, J.I. Arabidopsis abi1-1 and abi2-1 phosphatase mutations reduce abscisic acid-induced cytoplasmic calcium rises in guard cells. Plant Cell 1999, 11, 1785–1798. [Google Scholar]
- Kuhn, J.M.; Boisson-Dernier, A.; Dizon, M.B.; Maktabi, M.H.; Schroeder, J.I. The protein phosphatase AtPP2CA negatively regulates abscisic acid signal transduction in Arabidopsis, and effects of abh1 on AtPP2CA mRNA. Plant Physiol. 2006, 140, 127–139. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhu, Y.; Zhai, H.; Cai, H.; Ji, W.; Luo, X.; Li, J.; Bai, X. AtPP2CG1, a protein phosphatase 2C, positively regulates salt tolerance of Arabidopsis in abscisic acid-dependent manner. Biochem. Biophys. Res. Commun. 2012, 422, 710–715. [Google Scholar] [CrossRef]
- Haider, M.S.; Kurjogi, M.M.; Khalil-Ur-Rehman, M. Drought stress revealed physiological, biochemical and gene-expressional variations in ‘Yoshihime’ peach (Prunus persica L.) cultivar. J. Plant Interact. 2018, 13, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Choopayak, C. Characterization and Functional Analysis of a Novel PP2C Phosphatase AP2C2 from Arabidopsis; University of Vienna: Vienna, Austria, 2008. [Google Scholar]
- Chen, C.; Yu, Y.; Ding, X.; Liu, B.; Duanmu, H.; Zhu, D.; Sun, X.; Cao, L. Genome-wide analysis and expression profiling of PP2C clade D under saline and alkali stresses in wild soybean and Arabidopsis. Protoplasma 2017, 255, 643–654. [Google Scholar] [CrossRef]
- Ali, M.; Javaid, A.; Naqvi, S.H.; Batcho, A.; Kayani, W.K.; Lal, A.; Sajid, I.A.; Nwogwugwu, J.O. Biotic stress triggered small RNA and RNAi defense response in plants. Mol. Biol. Rep. 2020, 47, 5511–5522. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Xiao, D.; Cui, Y.; Xu, F.; Xu, X.; Gao, G.; Wang, Y.; Guo, Z.; Wang, D.; Wang, N.N. Senescence-suppressed protein phosphatase directly interacts with the cytoplasmic domain of senescence-associated receptor-like kinase and negatively regulates leaf senescence in Arabidopsis. Plant Physiol. 2015, 169, 1275–1291. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.S.; Sun, H.; Wang, Z.F.; Duan, M.; Huang, S.D.; Yang, J.; Huang, J.; Zhang, H.S. A novel nuclear protein phosphatase 2C negatively regulated by ABL1 is involved in abiotic stress and panicle development in rice. Mol. Biotechnol. 2013, 54, 703–710. [Google Scholar] [CrossRef]
- Li, W.; Cui, X.; Meng, Z.; Huang, X.; Xie, Q.; Wu, H.; Jin, H.; Zhang, D.; Liang, W. Transcriptional regulation of Arabidopsis MIR168a and ARGONAUTE1 homeostasis in ABA and abiotic stress responses. Plant Physiol. 2012, 158, 1279–1292. [Google Scholar] [CrossRef] [Green Version]
- Maestrini, P.; Cavallini, A.; Rizzo, M.; Giordani, T.; Bernardi, R.; Durante, M.; Natali, L. Isolation and expression analysis of low temperature induced genes in white poplar (Populus alba). Plant Physiol. 2009, 166, 1544–1556. [Google Scholar] [CrossRef]
- Wen, F.; Zhu, H.; Li, P.; Jiang, M.; Mao, W.; Ong, C.; Chu, Z. Genome-wide evolutionary characterization and expression analyses of WRKY family genes in Brachypodium distachyon. DNA Res. 2014, 21, 327–339. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Identifier | Chromosome | AA | PIs/MW | GRAVY | Instability | Subcellular |
|---|---|---|---|---|---|---|---|
| Location | Index | Localization | |||||
| SlPP2C01 | Solyc01g065700.3.1 | chr01: 72,289,658–72,295,088 | 361 | 6.47/39,992.45 | −0.266 | 35.78 | Cyto |
| SlPP2C02 | Solyc01g066870.3.1 | chr01: 750,92,877–75,095,866 | 522 | 5.09/57,125.35 | −0.202 | 42.69 | Cyto |
| SlPP2C03 | Solyc01g067980.1.1 | chr01: 77,027,259–77,032,781 | 202 | 6.19/20,650.27 | −0.001 | 31.43 | Extr. Nucl |
| SlPP2C04 | Solyc01g068000.4.1 | chr01: 77,050,710–77,057,722 | 297 | 4.89/32,913.15 | −0.202 | 54.88 | Nucl |
| SlPP2C05 | Solyc01g079720.3.1 | chr01: 78,822,308–78,843,121 | 613 | 6/68,185.28 | −0.282 | 45.77 | Cyto |
| SlPP2C06 | Solyc01g080400.3.1 | chr01: 79,645,915–79,653,551 | 414 | 4.81/45,150.79 | −0.406 | 46.18 | Nucl |
| SlPP2C07 | Solyc01g087460.3.1 | chr01: 82,424,646–82,428,543 | 372 | 6.17/41,583.98 | −0.655 | 38.39 | Nuc |
| SlPP2C08 | Solyc01g094230.4.1 | chr01: 85,781,740–85,791,508 | 627 | 11.39/64,450.04 | −0.476 | 35.66 | Nucl |
| SlPP2C09 | Solyc01g100040.3.1 | chr01: 90,110,898–90,115,136 | 367 | 6.22/40,919.44 | −0.258 | 38.73 | Nucl |
| SlPP2C10 | Solyc01g100110.4.1 | chr01: 90,196,815–90,202,470 | 427 | 6.91/46,135.74 | −0.146 | 43.86 | Cyto. Nucl |
| SlPP2C11 | Solyc01g105280.3.1 | chr01: 93,506,692–93,513,168 | 283 | 6.8/30,954.82 | −0.426 | 36.53 | Cyto. Nuc |
| SlPP2C12 | Solyc01g107300.4.1 | chr01: 94,888,760–94,892,503 | 278 | 7.79/31,335.87 | −0.128 | 46.13 | Cyto |
| SlPP2C13 | Solyc01g111730.3.1 | chr01: 97,915,696–97,923,710 | 388 | 8.23/42,809.56 | −0.263 | 44.55 | Chlo. Nucl |
| SlPP2C14 | Solyc02g082490.4.1 | chr02: 46,189,450–46193937 | 384 | 5.67/41,815.49 | −0.313 | 50.46 | Nucl |
| SlPP2C15 | Solyc02g083420.3.1 | chr02: 46,831,821–46,836,739 | 390 | 7.74/43,034.89 | −0.226 | 48.53 | Nucl |
| SlPP2C16 | Solyc02g092750.3.1 | chr02: 53,716,175–53,721,010 | 366 | 5.45/40,529.01 | −0.229 | 39.32 | Nucl |
| SlPP2C17 | Solyc03g006930.3.1 | chr03: 1,502,552–1,506,122 | 331 | 6.09/36,843.85 | −0.25 | 59.95 | Cyto |
| SlPP2C18 | Solyc03g006940.3.1 | chr03: 1,509,616–1,512,540 | 345 | 4.95/37,872.93 | −0.185 | 57 | Nucl |
| SlPP2C19 | Solyc03g006950.3.1 | chr03: 1,514,383–1,518,178 | 347 | 6.02/38,049.53 | −0.147 | 53.69 | Nucl |
| SlPP2C20 | Solyc03g006960.4.1 | chr03: 1,519,421–1,523,583 | 304 | 5.7/33,397.94 | −0.255 | 56.99 | Cyto |
| SlPP2C21 | Solyc03g007230.4.1 | chr03: 1,791,778–17,95,346 | 397 | 5.72/43,979.53 | −0.424 | 50.8 | Nucl |
| SlPP2C22 | Solyc03g007270.3.1 | chr03: 1,828,441–1,838,542 | 299 | 4.96/32,489.18 | −0.432 | 43.54 | Nucl |
| SlPP2C23 | Solyc03g013100.1.1 | chr03: 47,516,649–47,518,201 | 114 | 6.51/12,939.8 | −0.337 | 35.03 | Mito |
| SlPP2C24 | Solyc03g013140.1.1 | chr03: 47,332,347–47,334,696 | 163 | 5.81/18,565.16 | −0.428 | 20.18 | Cyto |
| SlPP2C25 | Solyc03g033340.3.1 | chr03: 4,912,004–4,918,389 | 397 | 7.65/43,970.18 | −0.217 | 47.54 | Chlo. Nucl |
| SlPP2C26 | Solyc03g065190.1.1 | chr03: 40,216,058–40,218,346 | 123 | 9.55/14,453 | −0.293 | 32.52 | Cyto. Extr. Nucl |
| SlPP2C27 | Solyc03g082960.2.1 | chr03: 52,814,842–52,820,023 | 395 | 6.29/43,427.5 | −0.203 | 44.71 | Nucl |
| SlPP2C28 | Solyc03g096670.3.1 | chr03: 58,989,562–58,992,297 | 406 | 6/44,683.73 | −0.397 | 52.59 | Cyto. Nucl |
| SlPP2C29 | Solyc03g118890.4.1 | chr03: 67,672,668–67,679,525 | 329 | 6.22/36,093.73 | −0.248 | 42.26 | Cyto |
| SlPP2C30 | Solyc03g121880.4.1 | chr03: 69,911,090–69,918,573 | 548 | 5.86/59,716.58 | −0.095 | 38.86 | Cyto |
| SlPP2C31 | Solyc04g056560.4.1 | chr04: 54,528,741–54,538,177 | 401 | 4.87/43,785.08 | −0.266 | 32.37 | Nucl |
| SlPP2C32 | Solyc04g064500.4.1 | chr04: 55,645,130–55,673,295 | 372 | 4.95/40,544.4 | −0.481 | 45.68 | Nucl |
| SlPP2C33 | Solyc04g074190.3.1 | chr04: 60,189,483–60,198,295 | 278 | 6.32/30,483.44 | −0.345 | 49.68 | Cyto. Nucl |
| SlPP2C34 | Solyc04g079120.3.1 | chr04: 63,704,094–63,711,682 | 428 | 5.28/46,166.21 | −0.296 | 47.94 | Nucl |
| SlPP2C35 | Solyc04g082600.3.1 | chr04: 66,227,666–66,234,622 | 387 | 4.85/42,607.25 | −0.265 | 47.37 | Chlo. Nucl |
| SlPP2C36 | Solyc05g009070.4.1 | chr05: 3,216,982–3,221,083 | 339 | 8.01/37,083.37 | −0.231 | 35.43 | Nucl |
| SlPP2C37 | Solyc05g018300.3.1 | chr05: 20,358,675–20,387,085 | 1080 | 4.81/119,917.55 | −0.203 | 42.53 | Nucl |
| SlPP2C38 | Solyc05g052520.3.1 | chr05: 62,745,936–62,749,176 | 396 | 5.56/42,845.56 | −0.233 | 53.55 | Nucl |
| SlPP2C39 | Solyc05g052980.4.1 | chr05: 63,149,350–63,151,994 | 437 | 5.69/48,147.65 | −0.281 | 52.66 | Nucl |
| SlPP2C40 | Solyc05g053290.3.1 | chr05: 63,421,852–63,425,476 | 411 | 4.73/44,751.23 | −0.109 | 43.03 | Nucl |
| SlPP2C41 | Solyc05g055790.4.1 | chr05: 65,291,032–65,295,556 | 499 | 5.23/55,284.13 | −0.375 | 52.06 | Nucl |
| SlPP2C42 | Solyc05g055980.4.1 | chr05: 65,383,352–65,389,926 | 314 | 6.91/35,158.87 | −0.366 | 43.45 | Chlo. Nucl |
| SlPP2C43 | Solyc06g007190.4.1 | chr06: 1,265,243–1,268,111 | 412 | 7.97/4,4889.01 | −0.182 | 44.84 | Nucl |
| SlPP2C44 | Solyc06g009390.3.1 | chr06: 3,311,448–3,316,710 | 352 | 4.94/38,881.06 | −0.11 | 56.6 | Nucl |
| SlPP2C45 | Solyc06g051940.4.1 | chr06: 35,603,081–35,605,882 | 471 | 5.8/51,885.49 | −0.36 | 43.54 | Nucl |
| SlPP2C46 | Solyc06g065610.3.1 | chr06: 40,985,604–40,992,533 | 375 | 5.59/40,805.91 | −0.342 | 45.11 | Nucl |
| SlPP2C47 | Solyc06g065920.4.1 | chr06: 41,312,022–41,318,633 | 374 | 8.51/41,712.42 | −0.298 | 46.39 | Nucl |
| SlPP2C48 | Solyc06g076100.3.1 | chr06: 47,243,874–47,248,381 | 708 | 5.41/79,199.49 | −0.537 | 38.2 | Nucl |
| SlPP2C49 | Solyc06g076400.3.1 | chr06: 47,471,880–47,474,601 | 410 | 5.12/44,859.67 | −0.346 | 46.42 | Nucl |
| SlPP2C50 | Solyc06g082080.3.1 | chr06: 48,014,366–48,017,179 | 379 | 8.18/40,564.02 | −0.241 | 52.55 | Nucl |
| SlPP2C51 | Solyc06g082700.1.1 | chr06: 48,429,677–48,430,277 | 142 | 6.09/15,813.08 | −0.125 | 26.13 | Cell membrane. Chlo. Cyto |
| SlPP2C52 | Solyc07g007220.3.1 | chr07: 1,956,469–1,961,556 | 383 | 5.07/42,272.7 | −0.264 | 47.32 | Nucl |
| SlPP2C53 | Solyc07g024010.2.1 | chr07: 23,836,770–23,838,030 | 136 | 6.07/15,409.59 | −0.258 | 24.63 | Chlo |
| SlPP2C54 | Solyc07g024020.2.1 | chr07: 23,848,754–23,851,910 | 321 | 5.09/34,819.28 | −0.321 | 40.61 | Nucl |
| SlPP2C55 | Solyc07g040990.4.1 | chr07: 51,484,565–51,489,081 | 536 | 5.38/59,196.43 | −0.267 | 47.81 | Nucl |
| SlPP2C56 | Solyc07g053760.4.1 | chr07: 62,192,070–62,195,463 | 286 | 6.67/31,303.63 | −0.331 | 37.81 | Nucl |
| SlPP2C57 | Solyc07g054300.3.1 | chr07: 62,654,289–62,659,599 | 478 | 5.38/53,166.78 | −0.45 | 44.34 | Nucl |
| SlPP2C58 | Solyc07g062970.3.1 | chr07: 65,595,765–65,600,127 | 282 | 5.67/30,943.93 | −0.327 | 28.7 | Nucl |
| SlPP2C59 | Solyc07g066260.3.1 | chr07: 67,742,920–67,748,833 | 515 | 5.75/56,463.59 | −0.371 | 38.51 | Chlo. Nucl |
| SlPP2C60 | Solyc08g006060.3.1 | chr08: 803,615–827,195 | 368 | 4.99/40,229.15 | −0.398 | 38.43 | Nucl |
| SlPP2C61 | Solyc08g007000.3.1 | chr08: 1,574,889–1,583,560 | 781 | 5.23/86,116.34 | −0.491 | 47.03 | Chlo |
| SlPP2C62 | Solyc08g044610.3.1 | chr08: 18,794,957–18,799,764 | 59 | 6.71/6738.81 | 0.012 | 42.27 | Chlo. Nucl. |
| SlPP2C63 | Solyc08g062640.3.1 | chr08: 51,623,577–51,625,677 | 134 | 7.61/15,444.01 | −0.244 | 40.43 | Nucl. |
| SlPP2C64 | Solyc08g062650.2.1 | chr08: 51,624,426–51,632,488 | 469 | 4.89/51,638.63 | −0.057 | 38.97 | Nucl |
| SlPP2C65 | Solyc08g065500.2.1 | chr08: 53,632,222–53,634,431 | 336 | 5.06/36,816.81 | −0.137 | 40.85 | Nucl |
| SlPP2C66 | Solyc08g065540.3.1 | chr08: 53,677,271–53,679,628 | 332 | 5.59/36,782.71 | −0.185 | 39.99 | Nucl |
| SlPP2C67 | Solyc08g065670.4.1 | chr08: 53,898,474–53,901,007 | 306 | 5.22/33,454.16 | −0.075 | 40.12 | Nucl |
| SlPP2C68 | Solyc08g065680.3.1 | chr08: 53,924,816–53,927,725 | 205 | 5.24/22,537.35 | −0.166 | 44.21 | Nucl |
| SlPP2C69 | Solyc08g074230.1.1 | chr08: 58,350,631–58,355,073 | 271 | 4.76/30,183.9 | −0.518 | 37.99 | Nucl |
| SlPP2C70 | Solyc08g077150.3.1 | chr08: 61,046,865–61,055,706 | 796 | 5.19/87,391.85 | −0.456 | 50.57 | Chlo |
| SlPP2C71 | Solyc08g082260.2.1 | chr08: 65,099,205–65,101,710 | 393 | 6.17/44,173.95 | −0.55 | 45.52 | Nucl |
| SlPP2C72 | Solyc09g007080.3.1 | chr09: 727,617–731,731 | 378 | 9.06/41,932.11 | −0.266 | 41.45 | Chlo. Cyto. Mito. Nucl |
| SlPP2C73 | Solyc09g010780.3.1 | chr09: 4,070,320–4,073,958 | 623 | 5.4/69,387.69 | −0.394 | 29.6 | Chlo |
| SlPP2C74 | Solyc09g065650.3.1 | chr09: 63,821,833–63,855,480 | 955 | 5.43/109,110.03 | −0.408 | 38.77 | Nucl |
| SlPP2C75 | Solyc09g090280.3.1 | chr09: 69,809,431–69,813,531 | 257 | 9.78/28,883.02 | −0.807 | 42.35 | Chlo. Nucl |
| SlPP2C76 | Solyc10g005640.4.1 | chr10: 513,071–520,386 | 556 | 5.56/61,356.83 | −0.502 | 41.89 | Nucl |
| SlPP2C77 | Solyc10g008490.3.1 | chr10: 2,617,430–2,620,523 | 469 | 5.02/51,815.01 | −0.455 | 46.09 | Nucl |
| SlPP2C78 | Solyc10g047290.2.1 | chr10: 40,367,471–40,374,422 | 281 | 8.32/30,574.56 | −0.431 | 40.11 | Nucl |
| SlPP2C79 | Solyc10g049630.2.1 | chr10: 46,299,689–46,303,378 | 381 | 9.06/42,379.44 | −0.334 | 48.38 | Nucl |
| SlPP2C80 | Solyc10g055650.2.1 | chr10: 57,152,293–57,156,799 | 388 | 8.67/43,047.9 | −0.271 | 42.35 | Nucl. Pero |
| SlPP2C81 | Solyc10g076320.3.1 | chr10: 59,239,617–59,243,965 | 344 | 6.05/38,842.14 | −0.424 | 40.32 | Chlo. Nucl |
| SlPP2C82 | Solyc10g078800.3.1 | chr10: 60,493,362–60,504,498 | 947 | 5.26/105,611.28 | −0.386 | 45.49 | Nucl |
| SlPP2C83 | Solyc10g078810.1.1 | chr10: 60,504,641–60,507,279 | 438 | 4.93/48,492.24 | −0.355 | 39.39 | Nucl |
| SlPP2C84 | Solyc10g078820.2.1 | chr10: 60,508,870–60,512,342 | 460 | 4.99/51,210.32 | −0.422 | 46.42 | Nucl |
| SlPP2C85 | Solyc10g084410.2.1 | chr10: 63,960,722–63,963,486 | 376 | 9.42/41,906.86 | −0.346 | 47.56 | Nucl |
| SlPP2C86 | Solyc10g085370.3.1 | chr10: 64,570,827–64,575,317 | 453 | 5.86/51,847.37 | −0.654 | 36.8 | Nucl |
| SlPP2C87 | Solyc10g086490.2.1 | chr10: 65,295,550–65,298,502 | 596 | 5.67/66,435.52 | −0.392 | 39 | Cell membrane. Chlo |
| SlPP2C88 | Solyc12g010450.3.1 | chr12: 3,470,481–3,476,558 | 316 | 5.75/34,814.63 | −0.272 | 43.32 | Cyto |
| SlPP2C89 | Solyc12g042570.2.1 | chr12: 39,939,943–39,959,306 | 362 | 4.92/39,315.91 | −0.512 | 39.46 | Nucl |
| SlPP2C90 | Solyc12g096020.3.1 | chr12: 65,098,300–65,101,758 | 508 | 4.62/55,681.53 | −0.235 | 45.3 | Nucl |
| SlPP2C91 | Solyc12g096520.3.1 | chr12: 65,400,452–65,408,533 | 293 | 5.04/31,552.91 | −0.349 | 34.47 | Cyto |
| SlPP2C92 | Solyc12g099600.2.1 | chr12: 66,688,468–66,694,648 | 497 | 5.35/53,856.74 | −0.189 | 47.82 | Nucl |
| Subgroup of PP2C Genes | Numbers of AtPP2Cs | Numbers of OsPP2Cs | Numbers of SlPP2Cs |
|---|---|---|---|
| A | 9 | 9 | 15 |
| B | 8 | 18 | 12 |
| C | 9 | 8 | 11 |
| D | 13 | 11 | 11 |
| E | 6 | 3 | 5 |
| F | 19 | 14 | 16 |
| G | 7 | 5 | 8 |
| H | 9 | 10 | 14 |
| Gene ID | Ka | Ks | Ka/Ks | Divergence Time (Myr) |
|---|---|---|---|---|
| SlPP2C04/44 | 0.09 | 0.69 | 0.12 | 23 |
| SlPP2C11/78 | 0.1 | 0.54 | 0.19 | 18 |
| SlPP2C13/80 | 0.08 | 0.63 | 0.12 | 21 |
| SlPP2C15/25 | 0.07 | 0.6 | 0.11 | 20 |
| SlPP2C15/12 | 0.12 | 2.04 | 0.06 | 68 |
| SlPP2C15/80 | 0.14 | 2.2 | 0.06 | 73.33 |
| SlPP2C18/19 | 0.08 | 0.09 | 0.93 | 3 |
| SlPP2C25/13 | 0.14 | 2.01 | 0.07 | 67 |
| SlPP2C33/11 | 0.19 | 1.68 | 0.11 | 56 |
| SlPP2C41/31 | 0.28 | 0.85 | 0.33 | 28.33 |
| SlPP2C57/57 | 0.09 | 0.75 | 0.12 | 25 |
| SlPP2C61/70 | 0.09 | 0.72 | 0.12 | 24 |
| SlPP2C66/65 | 0.08 | 0.37 | 0.22 | 12.33 |
| SlPP2C67/65 | 0.02 | 0.07 | 0.26 | 2.33 |
| SlPP2C80/25 | 0.15 | 1.93 | 0.08 | 64.33 |
| SlPP2C84/83 | 0.08 | 0.12 | 0.64 | 4 |
| SlPP2C85/72 | 0.12 | 0.82 | 0.15 | 27.33 |
| Motif | Width | Multilevel Consensus Sequence |
|---|---|---|
 1 1 | 15 | FLILASDGLWDVLSN |
 2 2 | 21 | VIQGETLYVANVGDSRAVLCR |
 3 3 | 15 | TFFGVYDGHGGPGAA |
 4 4 | 20 | VWRVKGGLAVSRAIGDKYLK |
 5 5 | 15 | AIQLSVDHKPNREDE |
 6 6 | 15 | RGSHDBISVIVVFLD |
 7 7 | 50 | HEGGDLGGRQDGLLWYKDLGQHANGEFSMAVVQANNLLEDQSQVESGPLS |
 8 8 | 41 | AVDIVHSYPRGGIARRLVKAALQEAAKKREMRYSDLKKIDR |
 9 9 | 20 | SQQGRRGEMEDAHIVWPBFC |
 10 10 | 14 | KKALRKAFLKTDEE |
| Cis-Element | Number of Genes | Functions of Cis-Elements | |
|---|---|---|---|
 | circadian | 19 | circadian control |
 | GT1-motif | 57 | light responsiveness |
 | G-box | 70 | light responsiveness |
 | MRE | 36 | light responsiveness |
 | ACE | 21 | light responsiveness |
 | 3-AF1 | 10 | light responsive |
 | Sp1 | 9 | light responsive |
 | 4cl-CMA2b | 1 | light responsive |
 | AAAC-motif | 3 | light responsive |
 | CGTCA-motif | 60 | MeJA-responsiveness |
 | TGACG-motif | 60 | MeJA-responsiveness |
 | ABRE | 70 | abscisic acid responsiveness |
 | TGA- | 26 | auxin-responsive |
 | AuxRR-core | 11 | auxin responsiveness |
 | TCA- | 47 | salicylic acid responsiveness |
 | SARE | 1 | salicylic acid responsiveness |
 | GARE-motif | 21 | gibberellin-responsive |
 | P-box | 31 | gibberellin-responsive |
 | TATC-box | 16 | gibberellin-responsiveness |
 | LTR | 24 | low-temperature responsiveness |
 | MBS | 38 | drought-inducibility |
 | MBSI | 8 | flavonoid biosynthetic genes regulation |
 | MSA-like | 4 | cell cycle regulation |
 | O2-site | 25 | zein metabolism regulation |
 | Box III | 4 | protein binding site |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, J.; Ni, L.; Xia, X.; Chen, S.; Zhang, Y.; Lang, M.; Li, M.; Liu, B.; Pan, Y.; Li, J.; et al. Genome-Wide Analysis of the Protein Phosphatase 2C Genes in Tomato. Genes 2022, 13, 604. https://doi.org/10.3390/genes13040604
Qiu J, Ni L, Xia X, Chen S, Zhang Y, Lang M, Li M, Liu B, Pan Y, Li J, et al. Genome-Wide Analysis of the Protein Phosphatase 2C Genes in Tomato. Genes. 2022; 13(4):604. https://doi.org/10.3390/genes13040604
Chicago/Turabian StyleQiu, Jianfang, Lei Ni, Xue Xia, Shihao Chen, Yan Zhang, Min Lang, Mengyu Li, Binman Liu, Yu Pan, Jinhua Li, and et al. 2022. "Genome-Wide Analysis of the Protein Phosphatase 2C Genes in Tomato" Genes 13, no. 4: 604. https://doi.org/10.3390/genes13040604
APA StyleQiu, J., Ni, L., Xia, X., Chen, S., Zhang, Y., Lang, M., Li, M., Liu, B., Pan, Y., Li, J., & Zhang, X. (2022). Genome-Wide Analysis of the Protein Phosphatase 2C Genes in Tomato. Genes, 13(4), 604. https://doi.org/10.3390/genes13040604