

Genetic Diversity Analysis and Core Germplasm Collection Construction of Camellia oleifera Based on Fruit Phenotype and SSR Data

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Fruit Phenotype Determination
2.3. DNA Extraction and PCR Analysis
2.4. Construction of Core Collection
2.5. SSR Analysis and Statistical Analysis
3. Results
3.1. Analysis of Fruit Phenotypic Variation
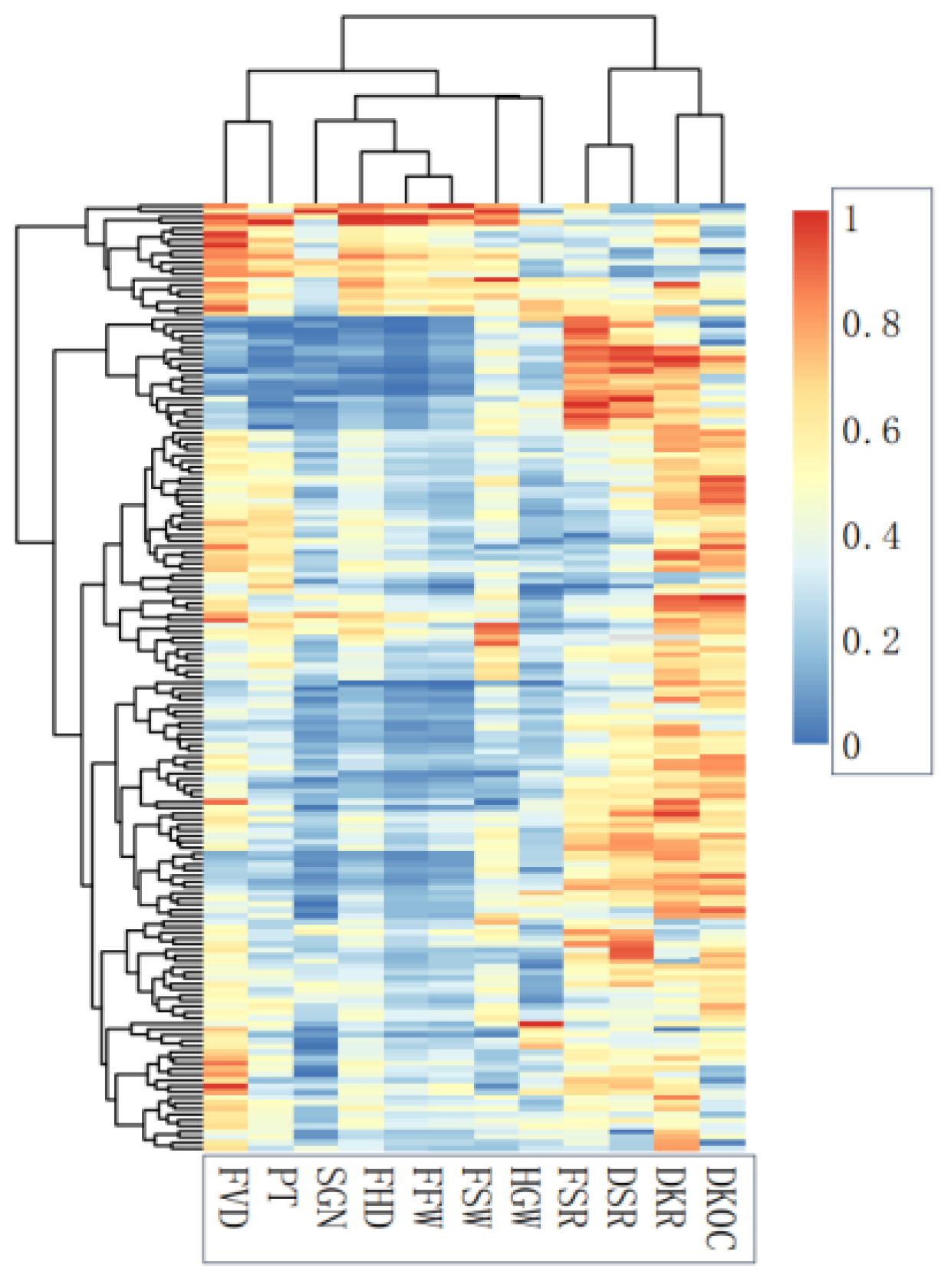
3.2. Correlation and Classification of Fruit Phenotype and Economic Characters
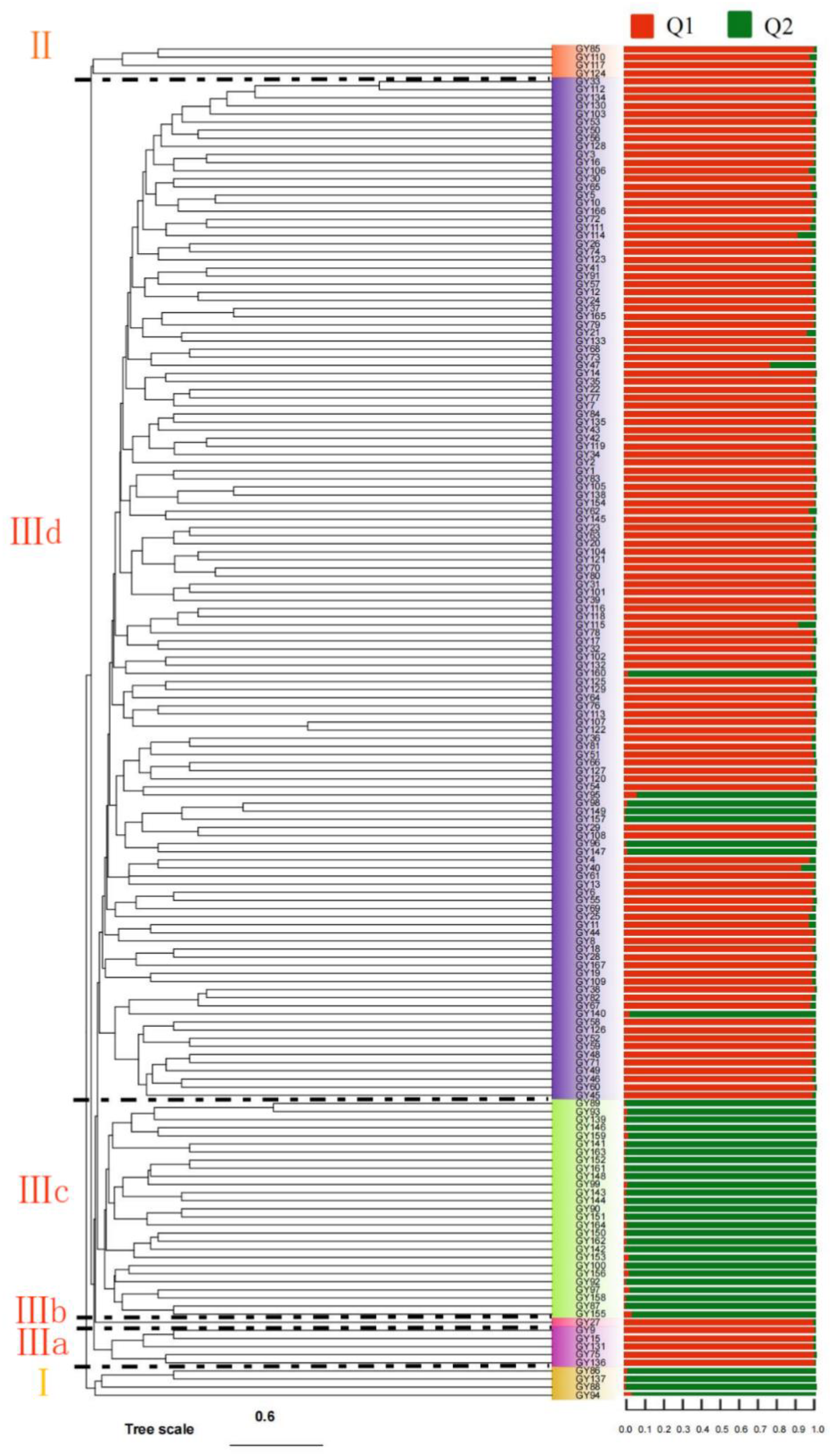
3.3. Genetic Diversity and Cluster Analysis
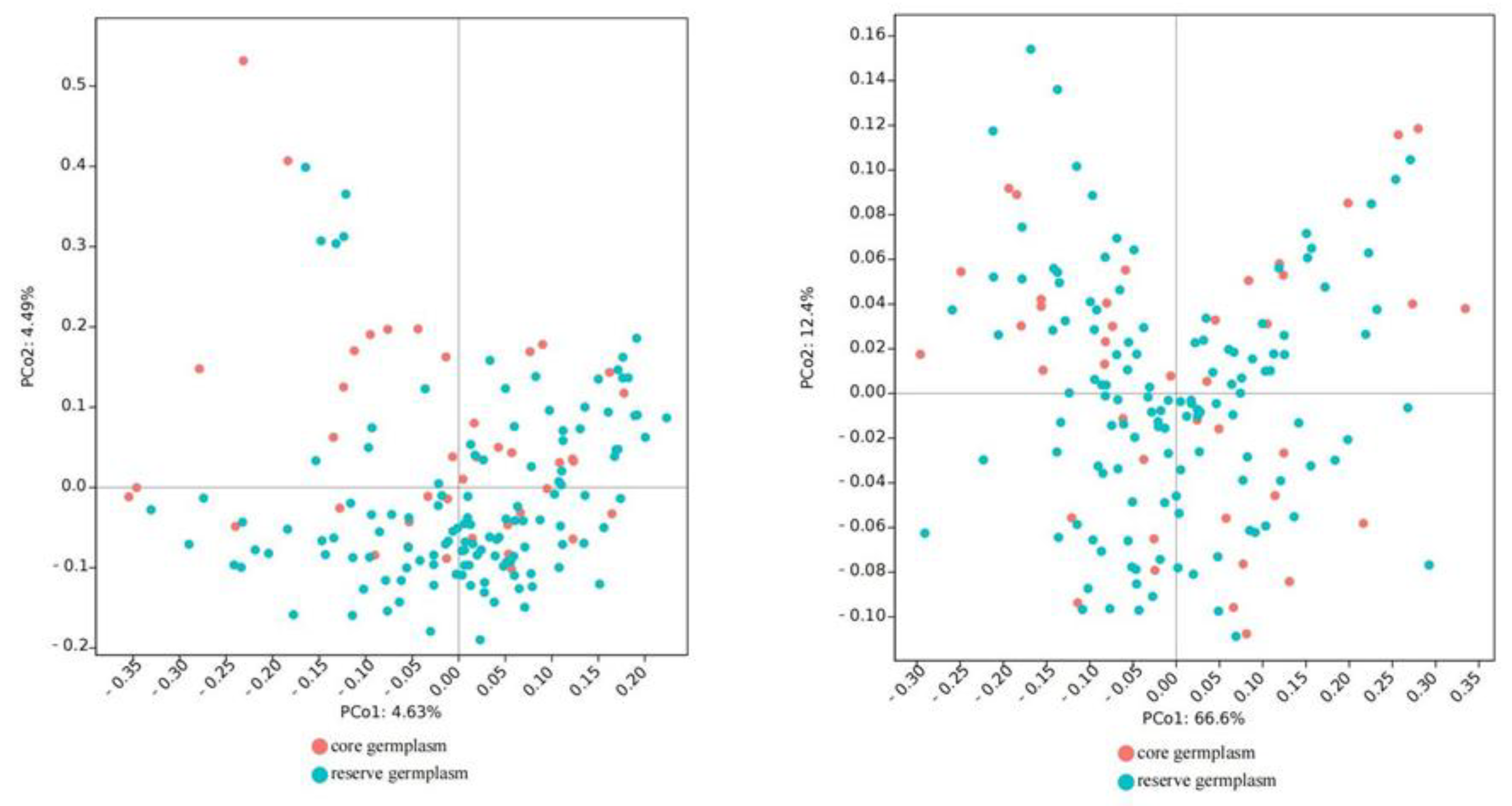
3.4. Construction of Core Collection
4. Discussion
4.1. Fruit Phenotypic Variation
4.2. Correlation Analysis and Cluster Analysis
4.3. Genetic Diversity and Genetic Structure Analysis
4.4. Construction of Core Collections
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, P.; Wang, K.; Wang, Y.; Hu, Z.; Yan, C.; Huang, H.; Ma, X.; Cao, Y.; Long, W.; Liu, W.; et al. The genome of oil-Camellia and population genomics analysis provide insights into seed oil domestication. Genome Biol. 2022, 23, 14. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Tan, X.; Liu, Z.; Lin, Q.; Zhang, L.; Yuan, J.; Zeng, Y.; Wu, L. In Vitro Propagation of Camellia oleifera Abel. Using Hypocotyl, Cotyledonary Node, and Radicle Explants. HortScience 2016, 51, 416–421. [Google Scholar] [CrossRef]
- Li, J.; Gao, G.; Li, B.; Li, B.; Lu, Q. Genetic Analysis of Prunus salicina L. by Random Amplified Polymorphic DNA (RAPD) and Intersimple Sequence Repeat (ISSR). Genet. Res. 2022, 2022, 2409324. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Shi, Y.Z.; Shoda, M.; Kotobuki, K.; Matsuta, N.; Hayashi, T.; Ban, Y.; Yamamoto, T. Identification of Asian Pear Varieties by SSR Analysis. Breed. Sci. 2002, 52, 115–121. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, H.; Tong, B.; Han, B.; Liu, D.; Zhang, P.; Hu, D. EST-SSR marker-based investigation on genetic diversity and genetic structure of Juglans mandshurica Maxim. in Shandong Province of China. Genet. Resour. Crop Evol. 2022, 1–11. [Google Scholar] [CrossRef]
- Chikh-Rouhou, H.; Mezghani, N.; Mnasri, S.; Mezghani, N.; Garcés-Claver, A. Assessing the genetic diversity and population structure of a tunisian melon (Cucumis melo L.) collection using phenotypic traits and SSR molecular markers. Agronomy 2021, 11, 1121. [Google Scholar] [CrossRef]
- Nyabera, L.A.; Nzuki, I.W.; Runo, S.M.; Amwayi, P.W. Assessment of genetic diversity of pumpkins (Cucurbita spp.) from western Kenya using SSR molecular markers. Mol. Biol. Rep. 2021, 48, 2253–2260. [Google Scholar] [CrossRef]
- He, Z.; Liu, C.; Wang, X.; Wang, R.; Chen, Y.; Tian, Y. Assessment of genetic diversity in Camellia oleifera Abel. accessions using morphological traits and simple sequence repeat (SSR) markers. Breed. Sci. 2020, 70, 586–593. [Google Scholar] [CrossRef]
- Razi, S.; Soleimani, A.; Zeinalabedini, M.; Vazifeshenas, M.R.; Martinez-Gomez, P.; Kermani, A.M.; Raiszadeh, A.R.; Tayari, M.; Martinez-Garcia, P.J. Development of a Multipurpose Core Collection of New Promising Iranian Pomegranate (Punica granatum L.) Genotypes Based on Morphological and Pomological Traits. Horticulturae 2021, 7, 350. [Google Scholar] [CrossRef]
- Xue, H.F.; Yu, X.C.; Fu, P.Y.; Liu, B.Y.; Zhang, S.; Li, J.; Zhai, W.J.; Lu, N.; Zhao, X.Y.; Wang, J.H.; et al. Construction of the Core Collection of Catalpa fargesii f. duclouxii (Huangxinzimu) Based on Molecular Markers and Phenotypic Traits. Forests 2021, 12, 1518. [Google Scholar] [CrossRef]
- Sivalingam, P.N.; Singh, D.; Chauhan, S.; Changal, H.K.; Bhan, C.; Mohapatra, T.; More, T.A.; Sharma, S.K. Establishment of the core collection of Ziziphus mauritiana Lam. from India. Plant Genet. Resour.-Charact. Util. 2014, 12, 140–142. [Google Scholar] [CrossRef]
- Gomes, R.L.F.; Costa, M.F.; Alves-Pereira, A.; Bajay, M.M.; Viana, J.P.G.; Valente, S.E.d.S.; Lopes, Â.C.d.; Zucchi, M.I.; Pinheiro, J.B. A lima bean core collection based on molecular markers. Sci. Agric. 2019, 77. [Google Scholar] [CrossRef]
- Lv, J.; Li, C.; Zhou, C.; Chen, J.; Li, F.; Weng, Q.; Li, M.; Wang, Y.; Chen, S.; Chen, J.; et al. Genetic diversity analysis of a breeding population of Eucalyptus cloeziana F. Muell. (Myrtaceae) and extraction of a core germplasm collection using microsatellite markers. Ind. Crops Prod. 2020, 145, 112157. [Google Scholar] [CrossRef]
- Boccacci, P.; Aramini, M.; Ordidge, M.; van Hintum, T.J.L.; Marinoni, D.T.; Valentini, N.; Sarraquigne, J.-P.; Solar, A.; Rovira, M.; Bacchetta, L.; et al. Comparison of selection methods for the establishment of a core collection using SSR markers for hazelnut (Corylus avellana L.) accessions from European germplasm repositories. Tree Genet. Genomes 2021, 17, 48. [Google Scholar] [CrossRef]
- Zhong, Y.; Wang, Y.; Sun, Z.; Niu, J.; Shi, Y.; Huang, K.; Chen, J.; Chen, J.; Luan, M. Genetic Diversity of a Natural Population of Akebia trifoliata (Thunb.) Koidz and Extraction of a Core Collection Using Simple Sequence Repeat Markers. Front. Genet. 2021, 12, 716498. [Google Scholar] [CrossRef]
- Sun, Y.; Dong, S.; Liu, Q.; Chen, J.; Pan, J.; Zhang, J. Selection of a core collection of Prunus sibirica L. germplasm by a stepwise clustering method using simple sequence repeat markers. PLoS ONE 2021, 16, e0260097. [Google Scholar] [CrossRef]
- Wang, X.; Cao, Z.; Gao, C.; Li, K. Strategy for the construction of a core collection for Pinus yunnanensis Franch. to optimize timber based on combined phenotype and molecular marker data. Genet. Resour. Crop Evol. 2021, 68, 3219–3240. [Google Scholar] [CrossRef]
- Sa, K.J.; Kim, D.M.; Oh, J.S.; Park, H.; Hyun, D.Y.; Lee, S.; Rhee, J.H.; Lee, J.K. Construction of a core collection of native Perilla germplasm collected from South Korea based on SSR markers and morphological characteristics. Sci. Rep. 2021, 11, 23891. [Google Scholar] [CrossRef]
- Munda, S.; Saikia, R.J.; Begum, T.; Bhandari, S.; Gogoi, A.; Sarma, N.; Tamang, R.; Lal, M. Evaluation of Genetic Diversity Based on Microsatellites and Phytochemical Markers of Core Collection of Cymbopogon winterianus Jowitt Germplasm. Plants 2022, 11, 528. [Google Scholar] [CrossRef]
- Yang, L.; Gao, C.; Xie, J.J.; Qiu, J.; Deng, Q.E.; Zhou, Y.C.; Liao, D.S.; Deng, C.Y. Fruit economic characteristics and yields of 40 superior Camellia oleifera Abel plants in the low-hot valley area of Guizhou Province, China. Sci. Rep. 2022, 12, 7068. [Google Scholar] [CrossRef]
- Zhang, Z.J. Study on Germplasm Resources and Elite Germplasm Selection of Camellia oleifera in Sanjiang Area. Master’s Thesis, Beijing Forestry University, Beijing, China, 2018. [Google Scholar]
- Ren, W. Genetic diversity and cross compatibility of elite genotypes in Camellia oleifera. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2018. [Google Scholar]
- Chen, Y.N.; Dai, X.G.; Hou, J.; Guan, H.W.; Wang, Y.X.; Li, Y.; Yin, T.M. DNA fingerprinting of oil camellia cultivars with SSR markers. Tree Genet. Genomes 2016, 12, 7. [Google Scholar] [CrossRef]
- Jia, B.G.; Lin, Q.; Zhang, L.; Tan, X.F.; Lei, X.L.; Hu, X.Y.; Shao, F.G. Development of 15 genic-SSR markers in oil-tea tree (Camellia oleifera) based on transcriptome sequencing. Genetika 2014, 46, 789–797. [Google Scholar] [CrossRef]
- Jia, B.G.; Lin, Q.; Feng, Y.Z.; Hu, X.Y.; Tan, X.F.; Shao, F.G.; Zhang, L. Development and cross-species transferability of unigene-derived microsatellite markers in an edible oil woody plant, Camellia oleifera (Theaceae). Genet. Mol. Res. 2015, 14, 6906–6916. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-W.; Chung, H.-K.; Cho, G.-T.; Ma, K.-H.; Chandrabalan, D.; Gwag, J.-G.; Kim, T.-S.; Cho, E.-G.; Park, Y.-J. PowerCore: A program applying the advanced M strategy with a heuristic search for establishing core sets. Bioinformatics 2007, 23, 2155–2162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.M.; Xi, J.W.; Hong, J.Y.; Xia, G.H. Phenotypic diversity of fruit and leaf traits of Carya cathayensis in Dabie Mountains. For. Res. 2020, 33, 10. [Google Scholar]
- Li, B.; Gu, W.-F. Review on genetic diversity in Pinus. Yi Chuan = Hered. 2003, 25, 740–748. [Google Scholar]
- Aspinwall, M.J.; Loik, M.E.; Resco de Dios, V.; Tjoelker, M.G.; Payton, P.R.; Tissue, D.T. Utilizing intraspecific variation in phenotypic plasticity to bolster agricultural and forest productivity under climate change. Plant Cell Environ. 2015, 38, 1752–1764. [Google Scholar] [CrossRef]
- Zhou, R.; Wu, Z.; Cao, X.; Jiang, F.L. Genetic diversity of cultivated and wild tomatoes revealed by morphological traits and SSR markers. Genet. Mol. Res. 2015, 14, 13868–13879. [Google Scholar] [CrossRef]
- Li, M.; Zhao, Z.; Miao, X.J. Genetic variability of wild apricot (Prunus armeniaca L.) populations in the Ili Valley as revealed by ISSR markers. Genet. Resour. Crop Evol. 2013, 60, 2293–2302. [Google Scholar] [CrossRef]
- Guliyev, N.; Sharifova, S.; Ojaghi, J.; Abbasov, M.; Akparov, Z. Genetic diversity among melon (Cucumis melo L.) accessions revealed by morphological traits and ISSR markers. Turk. J. Agric. For. 2018, 42, 393–401. [Google Scholar] [CrossRef]
- Gao, S.; Wang, B.; Liu, F.; Zhao, J.; Yuan, J.; Xiao, S.; Masabni, J.; Zou, F.; Yuan, D. Variation in Fruit Morphology and Seed Oil Fatty Acid Composition of Camellia oleifera Collected from Diverse Regions in Southern China. Horticulturae 2022, 8, 818. [Google Scholar] [CrossRef]
- Xie, Y.; Yao, X.; Li, Z.; Huang, Y. Analysis of Genetic Difference and Relationship of Camellia meiocarpa Native Varieties by Morphology and AFLP Markers. For. Res. 2014, 27, 201–207. [Google Scholar]
- Chen, K. Camellia meiocarpa and Camellia oleifera Phylogenetic Relationship and Fruit Size Differentiation Transcriptome Analysis. Masters’s Thesis, Nanchang University, Nanchang, China, 2020. [Google Scholar]
- Liang, D.Y.; Zhang, X.X.; Wang, C.; Wang, X.W.; Li, K.L.; Liu, G.F.; Zhao, X.Y.; Qu, G.Z. Evaluation of Betula platyphylla Families Based on Growth and Wood Property Traits. For. Sci. 2018, 64, 663–670. [Google Scholar] [CrossRef]
- Yang, L.; Gao, C.; Wei, H.L.; Long, L.; Qiu, J. Evaluation of the economic characteristics of the fruit of 45 superior Camellia weiningensis YK Li. trees. PLoS ONE 2022, 17, e0268802. [Google Scholar]
- Gong, W.; Xiao, S.; Wang, L.; Liao, Z.; Chang, Y.; Mo, W.; Hu, G.; Li, W.; Zhao, G.; Zhu, H. Chromosome-level genome of Camellia lanceoleosa provides a valuable resource for understanding genome evolution and self-incompatibility. Plant J. 2022, 110, 881–898. [Google Scholar] [CrossRef]
- Zhao, Y.; Ruan, C.J.; Ding, G.J.; Mopper, S. Genetic relationships in a germplasm collection of Camellia japonica and Camellia oleifera using SSR analysis. Genet. Mol. Res. GMR 2017, 16, 16019526. [Google Scholar] [CrossRef]
- Li, Q.; Su, X.; Ma, H.; Du, K.; Yang, M.; Chen, B.; Fu, S.; Fu, T.; Xiang, C.; Zhao, Q. Development of genic SSR marker resources from RNA-seq data in Camellia japonica and their application in the genus Camellia. Sci. Rep. 2021, 11, 9919. [Google Scholar] [CrossRef]
- Jan, S.; Khan, M.N.; Jan, S.; Zaffar, A.; Rashid, R.; Khan, M.A.; Sheikh, F.A.; Bhat, M.A.; Mir, R.R. Trait phenotyping and molecular marker characterization of barley (Hordeum vulgare L.) germplasm from Western Himalayas. Genet. Resour. Crop Evol. 2022, 69, 661–676. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.-T.; Wei, J.-T.; Li, Y.; Tigabu, M.; Zhao, X.-Y. Genetic Improvement of Pinus koraiensis in China: Current Situation and Future Prospects. Forests 2020, 11, 148. [Google Scholar] [CrossRef]
- Velázquez-Barrera, M.E.; Ramos-Cabrer, A.M.; Pereira-Lorenzo, S.; Ríos-Mesa, D.J. Genetic pool of the cultivated pear tree (Pyrus spp.) in the Canary Islands (Spain), studied using SSR molecular markers. Agronomy 2022, 12, 1711. [Google Scholar] [CrossRef]
- Han, P.; Tian, X.M.; Wang, Y.; Huang, C.; Ma, Y.Z.; Zhou, X.F.; Yu, Y.; Zhang, D.W.; Xu, H.J.; Cao, Y.; et al. Construction of a core germplasm bank of upland cotton (Gossypium hirsutum L.) based on phenotype, genotype and favorable alleles. Genet. Resour. Crop Evol. 2022, 69, 2399–2411. [Google Scholar] [CrossRef]
- Mahmoodi, R.; Dadpour, M.R.; Hassani, D.; Zeinalabedini, M.; Vendramin, E.; Leslie, C.A. Composite core set construction and diversity analysis of Iranian walnut germplasm using molecular markers and phenotypic traits. PLoS ONE 2021, 16, e0248623. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Oh, Y.; Lee, G.-A.; Kwon, Y.S.; Kim, S.A.; Kwon, S.-I.; Doi, Y.-S.; Choi, C. Genetic Diversity, Structure, and Core Collection of Korean Apple Germplasm Using Simple Sequence Repeat Markers. Hortic. J. 2019, 88, 329–337. [Google Scholar] [CrossRef]
- Belaj, A.; Dominguez-Garcia, M.D.; Atienza, S.G.; Urdiroz, N.M.; De la Rosa, R.; Satovic, Z.; Martin, A.; Kilian, A.; Trujillo, I.; Valpuesta, V.; et al. Developing a core collection of olive (Olea europaea L.) based on molecular markers (DArTs, SSRs, SNPs) and agronomic traits. Tree Genet. Genomes 2012, 8, 365–378. [Google Scholar] [CrossRef]
- Mahmoodi, R.; Dadpour, M.R.; Hassani, D.; Zeinalabedini, M.; Vendramin, E.; Micali, S.; Nahandi, F.Z. Development of a core collection in Iranian walnut (Juglans regia L.) germplasm using the phenotypic diversity. Sci. Hortic. 2019, 249, 439–448. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collection Place | North Latitude/ (°) | East Longitude / (°) | Altitude/m | Germplasm Code |
|---|---|---|---|---|
| Liping in Guizhou | 26.3355 | 109.1819 | 438 | GY85-164 |
| Jinping in Guizhou | 26.6096 | 109.3642 | 515 | GY1-35 |
| Yuping in Guizhou | 27.3125 | 108.9194 | 536 | GY165-167 |
| Guizhou Academy of Forestry | 26.5241 | 106.7426 | 1221 | GY36-84 |
| Marker Name | Forward (5′-3′) | Reverse (3′-5′) | Tm/ ℃ | References |
|---|---|---|---|---|
| CoUg3417 | CGGGAATCAAAAAGCTAGG | AGTGGGTGGTGAACACCAA | 56 | [24] |
| CoUg4931 | CTTCATCTCCGTTTGCTCT | GACTTTGCCTCCTTTTGTG | 55 | [25] |
| CoUg7330 | ACAACCATCTTCCTCTCCCC | CGTCGTCCCTGTTCACCTCT | 60 | [25] |
| CoUg11592 | TACCGTGCTTGTGTATTCC | GTGTTTTGTGTTGCTGCCT | 55 | [24] |
| CoUg3402 | ACTCTTGTGGGTGAATGTTG | GCTGGTAGGTTGGTTATGTT | 56 | [25] |
| CoUg5179 | AATGGAGAATGAATGGACAG | GCAGAAAGTGATATTGGGTG | 55 | [25] |
| CoUg11169 | GTCTGGTGGCGTTGCTTGCT | GTCTGCTGATCCGATGGCTG | 62 | [24] |
| CoUg17436 | TTGAGGGTGAAGTCGATGA | AAGGAGTTGGTGAGTAGCA | 55 | [24] |
| CoUg8099 | TGGGGATTGCTCAAAAGTGT | AGGGTGGCTGTGCTGGTATT | 58 | [24] |
| CoUg8134 | CCAGAGCCAGGAGGAAGTA | GAGAGAGGGGGTAGAATGA | 58 | [24] |
| CoUg13753 | CACATCATTAGGGTCGTTG | GGTTTTCACTCTTCAGCAG | 55 | [24] |
| CoUg4364 | GTGGTCCTGGAGATCTGTCC | TTTCGCTCTATCCGTTGTTC | 57 | [25] |
| Phenotypic Traits | Mean ± SD | MS | F Value | |||
|---|---|---|---|---|---|---|
| Among Different Varieties | Within the Varieties | Random Errors | Among Different Varieties | Within the Varieties | ||
| FHD/mm | 24.509 ± 4.646 | 442.992 | 26.111 | 6.901 | 64.081 ** | 2.435 ** |
| FVD/mm | 26.734 ± 4.544 | 398.788 | 24.009 | 7.517 | 52.933 ** | 2.357 ** |
| FFW/g | 9.388 ± 5.123 | 518.612 | 33.615 | 8.943 | 57.828 ** | 2.71 ** |
| SGN | 2.795 ± 1.726 | 45.910 | 4.002 | 1.446 | 31.735 ** | 2.184 ** |
| FSW/g | 4.195 ± 2.388 | 93.382 | 9.422 | 2.455 | 37.895 ** | 2.724 ** |
| PT/mm | 2.404 ± 0.873 | 17.038 | 0.483 | 0.226 | 75.322 ** | 2.071 ** |
| FSI | 0.92 ± 0.108 | 0.194 | 0.009 | 0.006 | 33.598 ** | 1.439 ** |
| Phenotypic Traits | Coefficient of Variation/ % | Variance Component | Percentage of Variance Component/ % | Phenotypic Differentiation Coefficient/ % | ||||
|---|---|---|---|---|---|---|---|---|
| among Different Varieties | within the Varieties | among Different Varieties | within the Varieties | Random Errors | among Different Varieties | within the Varieties | ||
| FHD/mm | 18.95 | 11.15 | 12.986 | 1.786 | 6.9013 | 59.92 | 8.24 | 87.91 |
| FVD/mm | 17.00 | 10.55 | 11.675 | 1.534 | 7.5173 | 56.33 | 7.4 | 88.39 |
| FFW/g | 54.56 | 30.14 | 15.108 | 2.294 | 8.9427 | 57.35 | 8.71 | 86.82 |
| SGN | 61.76 | 42.31 | 1.306 | 0.238 | 1.4455 | 43.68 | 7.95 | 84.60 |
| FSW/g | 56.92 | 36.42 | 2.616 | 0.648 | 2.4552 | 45.74 | 11.33 | 80.15 |
| PT/mm | 36.32 | 19.73 | 0.516 | 0.024 | 0.2261 | 67.35 | 3.13 | 95.56 |
| FSI | 11.79 | 8.15 | 0.006 | 0.000 | 0.0057 | 48.75 | 2.59 | 94.96 |
| Marker Name | Na | Ne | I | Ho | He | PIC | MAF |
|---|---|---|---|---|---|---|---|
| CoUg11592 | 8 | 2.830 | 1.285 | 0.952 | 0.647 | 0.592 | 0.500 |
| CoUg3402 | 6 | 2.689 | 1.257 | 0.867 | 0.628 | 0.590 | 0.548 |
| CoUg3417 | 27 | 14.228 | 2.864 | 0.982 | 0.930 | 0.927 | 0.123 |
| CoUg8134 | 19 | 10.662 | 2.573 | 0.994 | 0.906 | 0.903 | 0.186 |
| CoUg5179 | 20 | 9.627 | 2.481 | 0.897 | 0.896 | 0.890 | 0.156 |
| CoUg8099 | 14 | 4.085 | 1.751 | 0.915 | 0.755 | 0.736 | 0.398 |
| CoUg17436 | 23 | 11.578 | 2.716 | 0.976 | 0.914 | 0.909 | 0.180 |
| CoUg4364 | 19 | 10.267 | 2.541 | 1.000 | 0.903 | 0.895 | 0.153 |
| CoUg4931 | 16 | 10.236 | 2.467 | 1.000 | 0.902 | 0.895 | 0.138 |
| CoUg11169 | 18 | 10.043 | 2.508 | 1.000 | 0.900 | 0.892 | 0.174 |
| CoUg13753 | 18 | 9.735 | 2.401 | 0.994 | 0.897 | 0.889 | 0.135 |
| CoUg7330 | 26 | 12.008 | 2.767 | 1.000 | 0.917 | 0.911 | 0.171 |
| Mean | 17.833 | 8.999 | 2.301 | 0.965 | 0.850 | 0.836 | 0.238 |
| Method | Sampling Rate (%) | Evaluation Parameters | ||||
|---|---|---|---|---|---|---|
| Trait Retention Ratio (%) | Mean Difference (%) | Variance Difference (%) | Coincidence Rate of Range (%) | Rate of Variable Coefficient Change (%) | ||
| QGA | 10 | 82.38 | 8.33 | 25 | 83.64 | 121.58 |
| 15 | 93.77 | 0 | 41.67 | 87.03 | 123.96 | |
| 20 | 93.77 | 0 | 41.67 | 87.83 | 118.55 | |
| 25 | 99.16 | 8.33 | 75 | 100 | 128.66 | |
| 30 | 97.50 | 0 | 8.33 | 94.9 | 116.87 | |
| PowerCore | 10 | 87.27 | 5.7 | 31.77 | 84.61 | 116.03 |
| 15 | 94.55 | 3.11 | 30.21 | 88.92 | 115.99 | |
| 20 | 96.36 | 4.28 | 26.63 | 91.08 | 111.26 | |
| 25 | 97.27 | 3.08 | 22.14 | 91.82 | 111.77 | |
| 30 | 99.09 | 1.68 | 18.59 | 92.18 | 109.43 | |
| Method | QGA | PowerCore | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sampling Rate | 100% | 10% | 15% | 20% | 25% | 30% | 10% | 15% | 20% | 25% | 30% |
| Na | 17.833 | 11.333 | 12.250 | 13.750 | 13.833 | 14.500 | 12.667 | 14.417 | 15.250 | 15.833 | 16.750 |
| Ne | 8.999 | 7.811 | 7.945 | 8.442 | 8.364 | 8.451 | 9.206 | 9.686 | 9.825 | 9.857 | 9.592 |
| I | 2.301 | 2.122 | 2.161 | 2.209 | 2.215 | 2.228 | 2.277 | 2.326 | 2.336 | 2.350 | 2.352 |
| Ho | 0.965 | 0.979 | 0.979 | 0.962 | 0.973 | 0.962 | 0.978 | 0.973 | 0.974 | 0.973 | 0.969 |
| He | 0.850 | 0.835 | 0.841 | 0.842 | 0.846 | 0.839 | 0.864 | 0.862 | 0.859 | 0.861 | 0.857 |
| uHe | 0.852 | 0.862 | 0.859 | 0.855 | 0.857 | 0.848 | 0.893 | 0.880 | 0.872 | 0.872 | 0.866 |
| Traits | Original Collection | Core Collection | Reserve Collection | t1 | t2 | |||
|---|---|---|---|---|---|---|---|---|
| Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | Range | |||
| FHD | 24.22 ± 3.75 | 17.04–36.41 | 24.61 ± 4.45 | 17.5–36.22 | 24.09 ± 3.51 | 17.04–36.41 | 0.581 | −0.298 |
| FVD | 26.57 ± 3.57 | 18.34–34.75 | 26.64 ± 4.02 | 18.65–34.31 | 26.55 ± 3.43 | 18.34–34.75 | 0.110 | −0.055 |
| FFW | 9.07 ± 4.06 | 3.20–25.64 | 9.69 ± 5.02 | 3.36–24.89 | 8.87 ± 3.69 | 3.20–25.64 | 0.826 | −0.433 |
| SGN | 2.72 ± 1.20 | 1.10–8.33 | 2.8 ± 1.23 | 1.10–6.4 | 2.69 ± 1.19 | 1.10–8.33 | 0.408 | −0.198 |
| FSW | 4.10 ± 1.74 | 1.34–11.62 | 4.39 ± 2.21 | 1.51–11.62 | 4.01 ± 1.55 | 1.34–10.86 | 0.900 | −0.48 |
| PT | 2.35 ± 0.72 | 0.89–4.67 | 2.44 ± 0.84 | 0.93–4.67 | 2.32 ± 0.68 | 0.89–3.99 | 0.730 | −0.372 |
| FSI | 0.92 ± 0.08 | 0.74–1.16 | 0.93 ± 0.08 | 0.76–1.13 | 0.91 ± 0.08 | 0.74–1.16 | 0.868 | −0.426 |
| HGW | 163.28 ± 63.54 | 48.69–444.58 | 171.4 ± 80.61 | 48.69–444.58 | 160.63 ± 57.03 | 59.02–337.88 | 0.693 | −0.368 |
| FSR | 0.47 ± 0.09 | 0.26–0.69 | 0.47 ± 0.10 | 0.26–0.69 | 0.46 ± 0.08 | 0.27–0.67 | 0.188 | −0.097 |
| DSR | 0.25 ± 0.05 | 0.12–0.38 | 0.25 ± 0.05 | 0.14–0.36 | 0.25 ± 0.05 | 0.12–0.38 | −0.494 | 0.240 |
| DKR | 0.64 ± 0.06 | 0.46–0.76 | 0.63 ± 0.06 | 0.51–0.74 | 0.64 ± 0.06 | 0.46–0.76 | −0.901 | 0.431 |
| DKOC | 0.42 ± 0.09 | 0.20–0.60 | 0.40 ± 0.10 | 0.21–0.60 | 0.42 ± 0.08 | 0.20–0.58 | −1.306 | 0.651 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Liang, D.; Song, Z.; Tan, Y.; Guo, X.; Wang, D. Genetic Diversity Analysis and Core Germplasm Collection Construction of Camellia oleifera Based on Fruit Phenotype and SSR Data. Genes 2022, 13, 2351. https://doi.org/10.3390/genes13122351
Zhu Y, Liang D, Song Z, Tan Y, Guo X, Wang D. Genetic Diversity Analysis and Core Germplasm Collection Construction of Camellia oleifera Based on Fruit Phenotype and SSR Data. Genes. 2022; 13(12):2351. https://doi.org/10.3390/genes13122351
Chicago/Turabian StyleZhu, Yunzheng, Deyang Liang, Zejun Song, Yi Tan, Xiaolan Guo, and Delu Wang. 2022. "Genetic Diversity Analysis and Core Germplasm Collection Construction of Camellia oleifera Based on Fruit Phenotype and SSR Data" Genes 13, no. 12: 2351. https://doi.org/10.3390/genes13122351
APA StyleZhu, Y., Liang, D., Song, Z., Tan, Y., Guo, X., & Wang, D. (2022). Genetic Diversity Analysis and Core Germplasm Collection Construction of Camellia oleifera Based on Fruit Phenotype and SSR Data. Genes, 13(12), 2351. https://doi.org/10.3390/genes13122351