

Analysis of Genetic Diversity and Phylogenetic Relationships of Wheat (Triticum aestivum L.) Genotypes Using Phenological, Molecular and DNA Barcoding Markers

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Design: Morphometric and Phenotypic Markers
2.3. DNA-Based Molecular Genetic Diversity Analyses: SCoT and ISSR Markers
2.4. DNA Barcoding Analysis: Plastid rbcL and matK Genes
2.5. Statistical Analysis
3. Results and Discussion
3.1. Phenological Markers
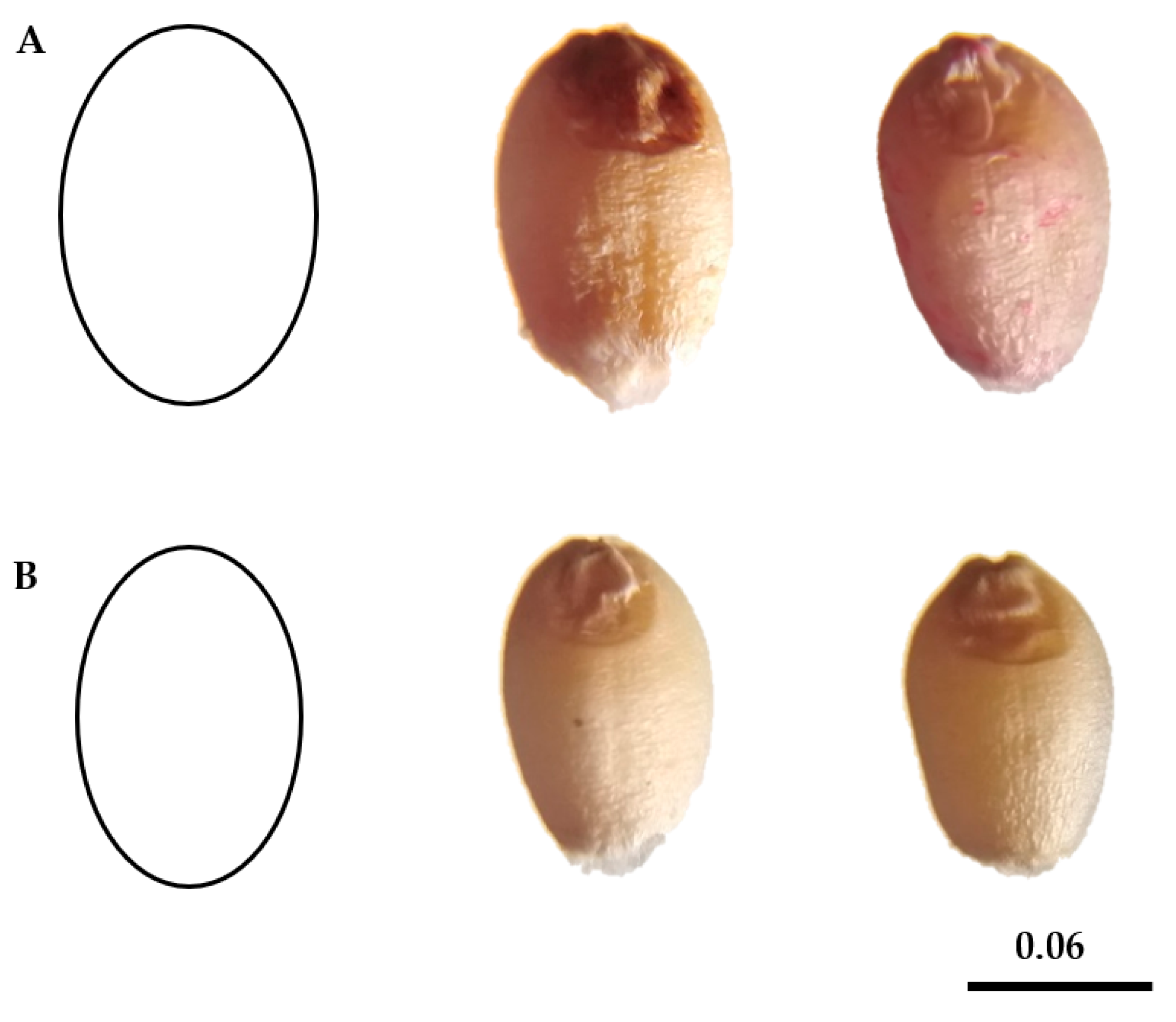
3.1.1. Phenological, Morphological and Geometric Traits Characterization
3.1.2. Availability and Hereditability of the Wheat Genotypes
3.1.3. Phenotypic and Genotypic Correlation Coefficients
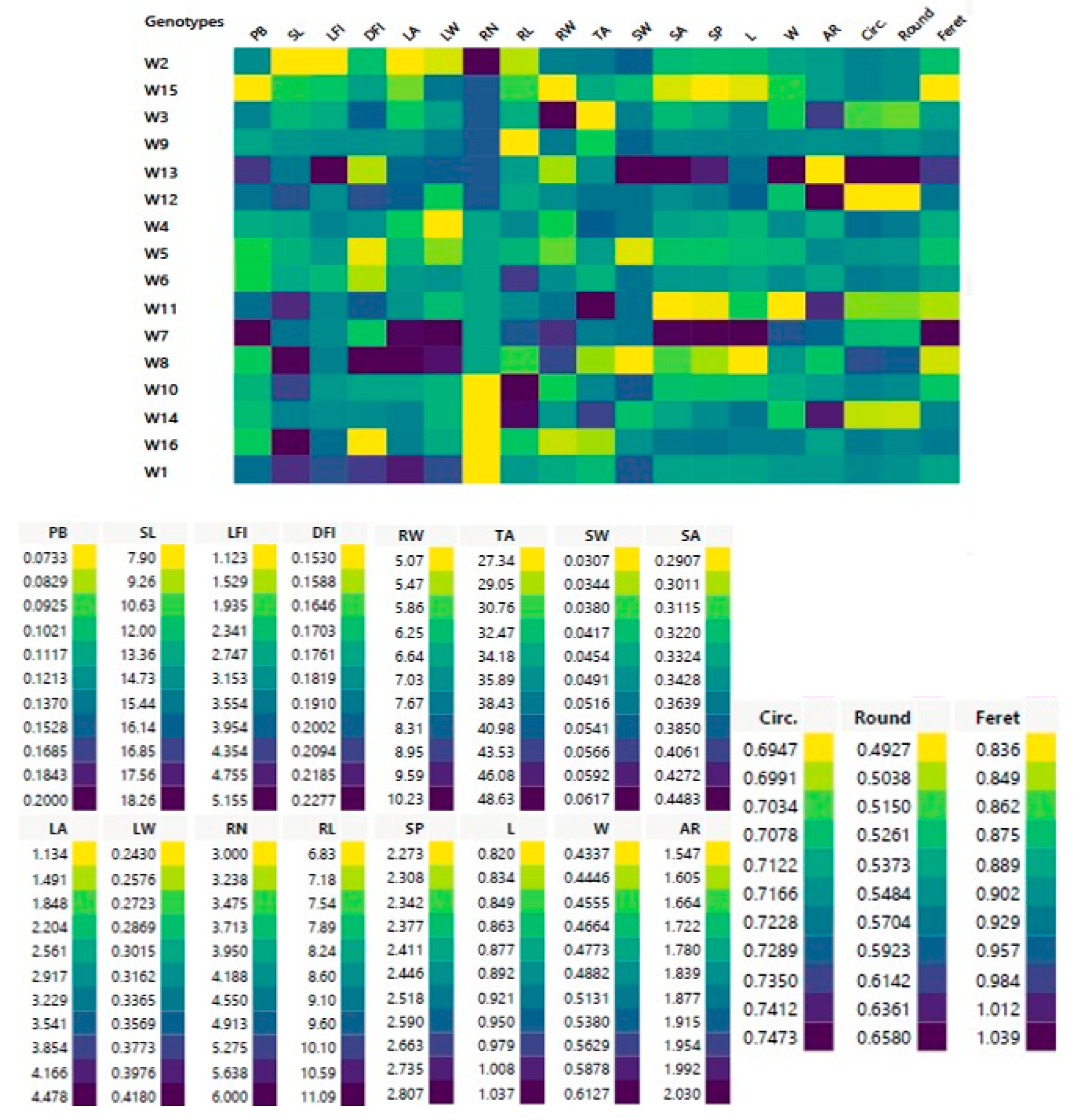
3.1.4. Hierarchical Co-Clustering Analysis Based on Morphological and Phenological Markers
3.2. Molecular Genetic Diversity Analyses: SCoT and ISSR Markers
3.2.1. ISSR Markers
3.2.2. SCoT Markers
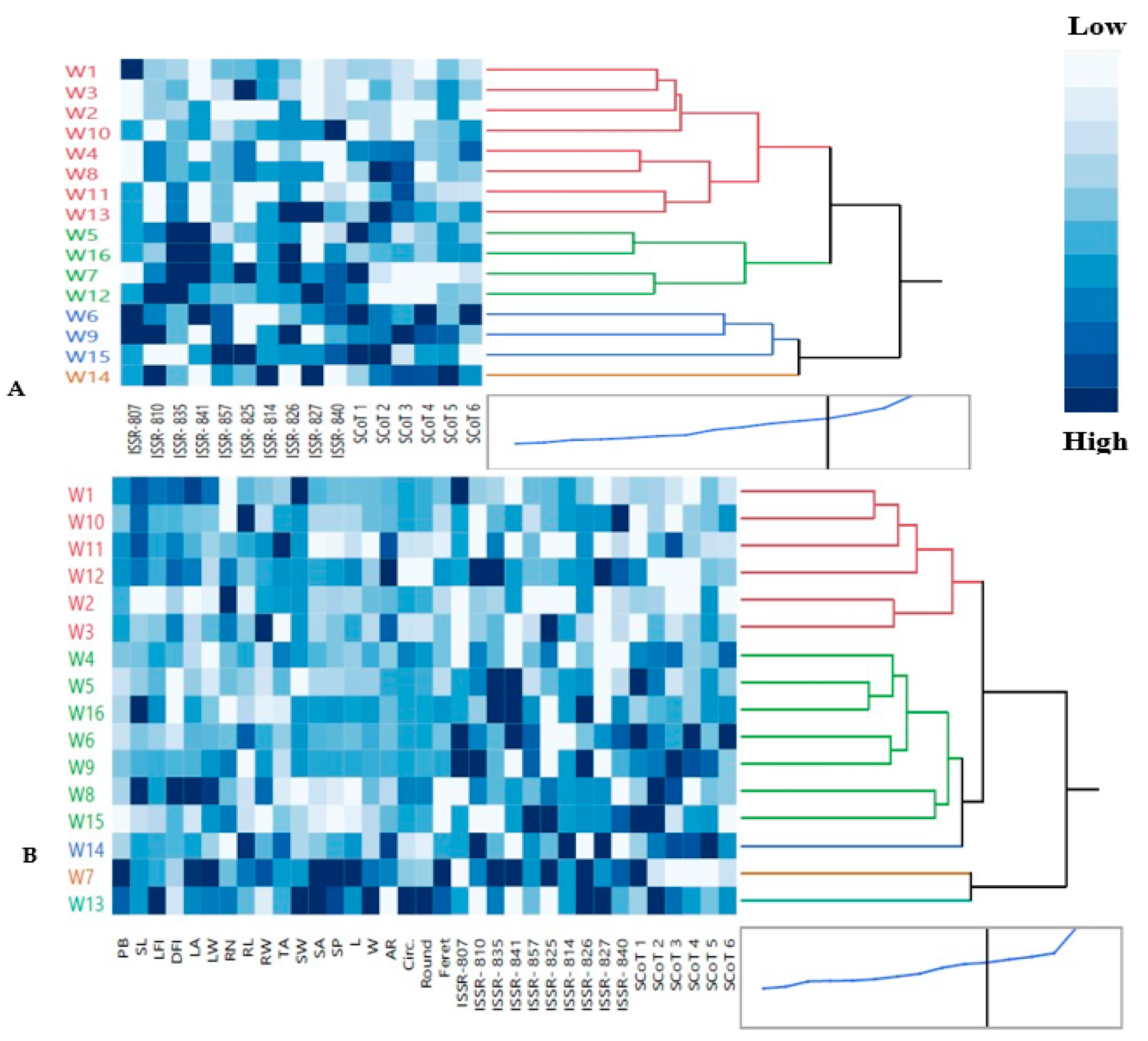
3.2.3. ISSR and SCoT Analysis
3.2.4. Multivariate Analysis Based on ISSR and SCoT Combined Data: Hierarchical Co-Clustering Analysis
3.3. Relationships among Morphological and Genetic Attributes
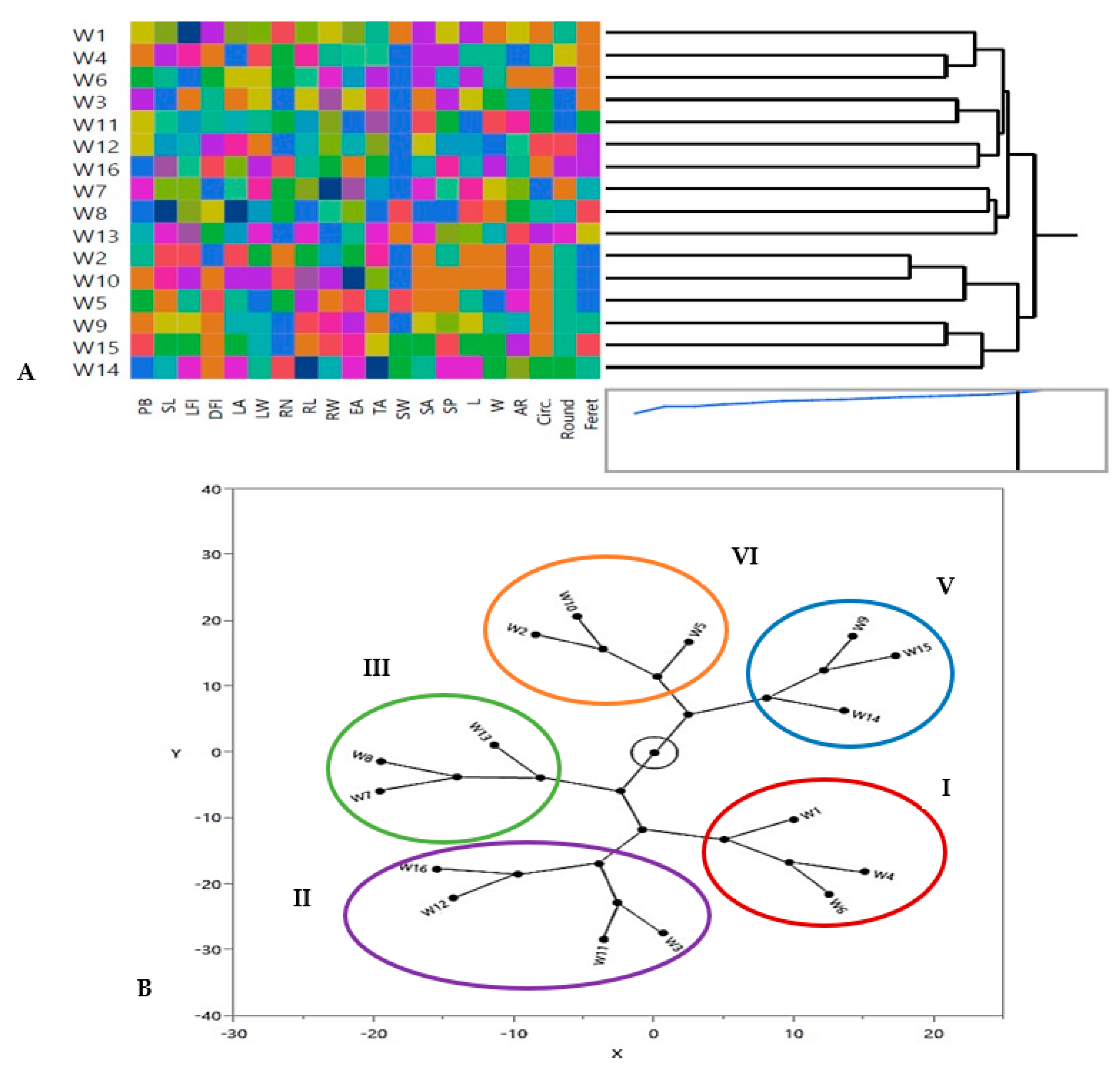
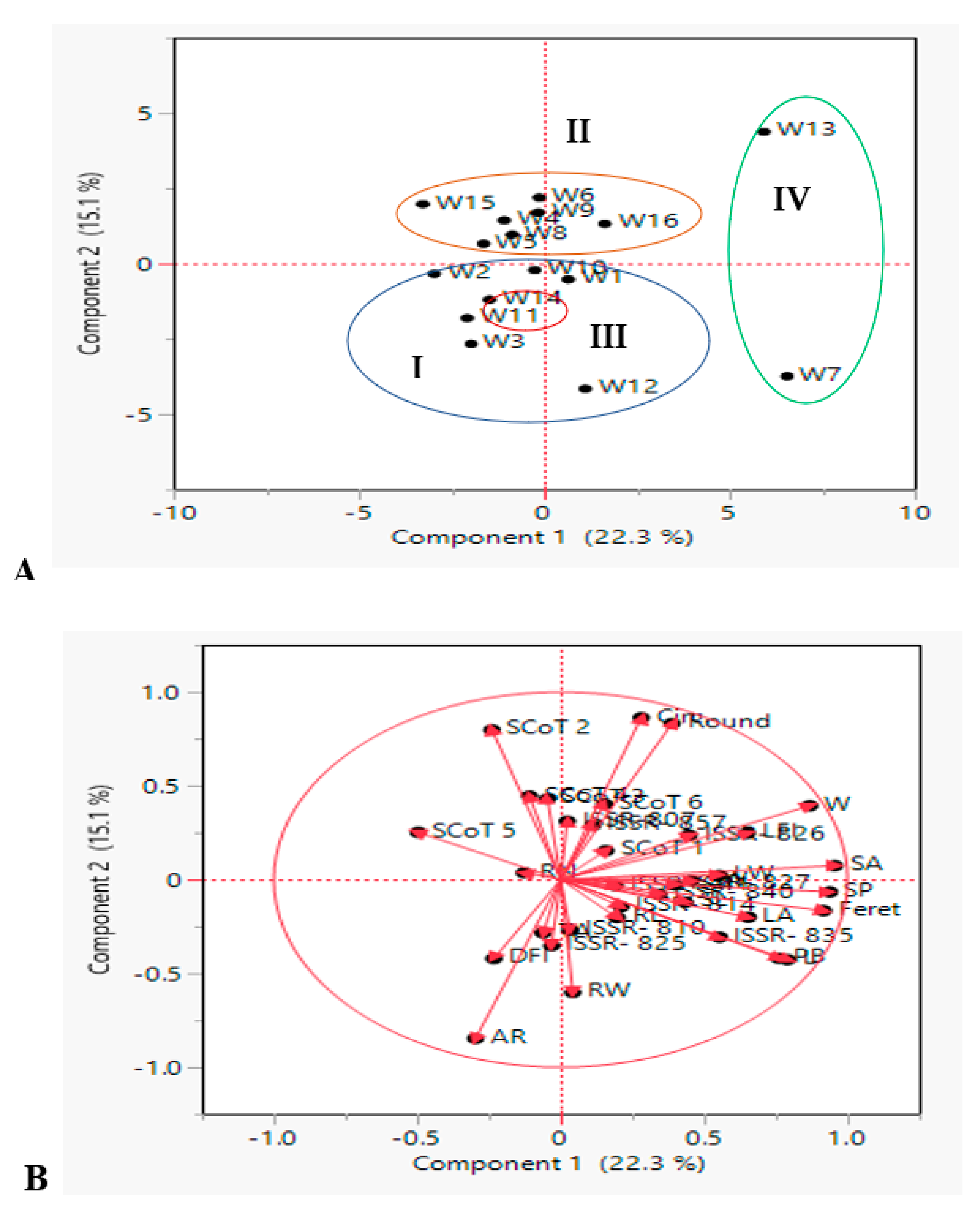
3.3.1. Hierarchical Co-Clustering Analysis
3.3.2. Principal Component Analysis (PCA)
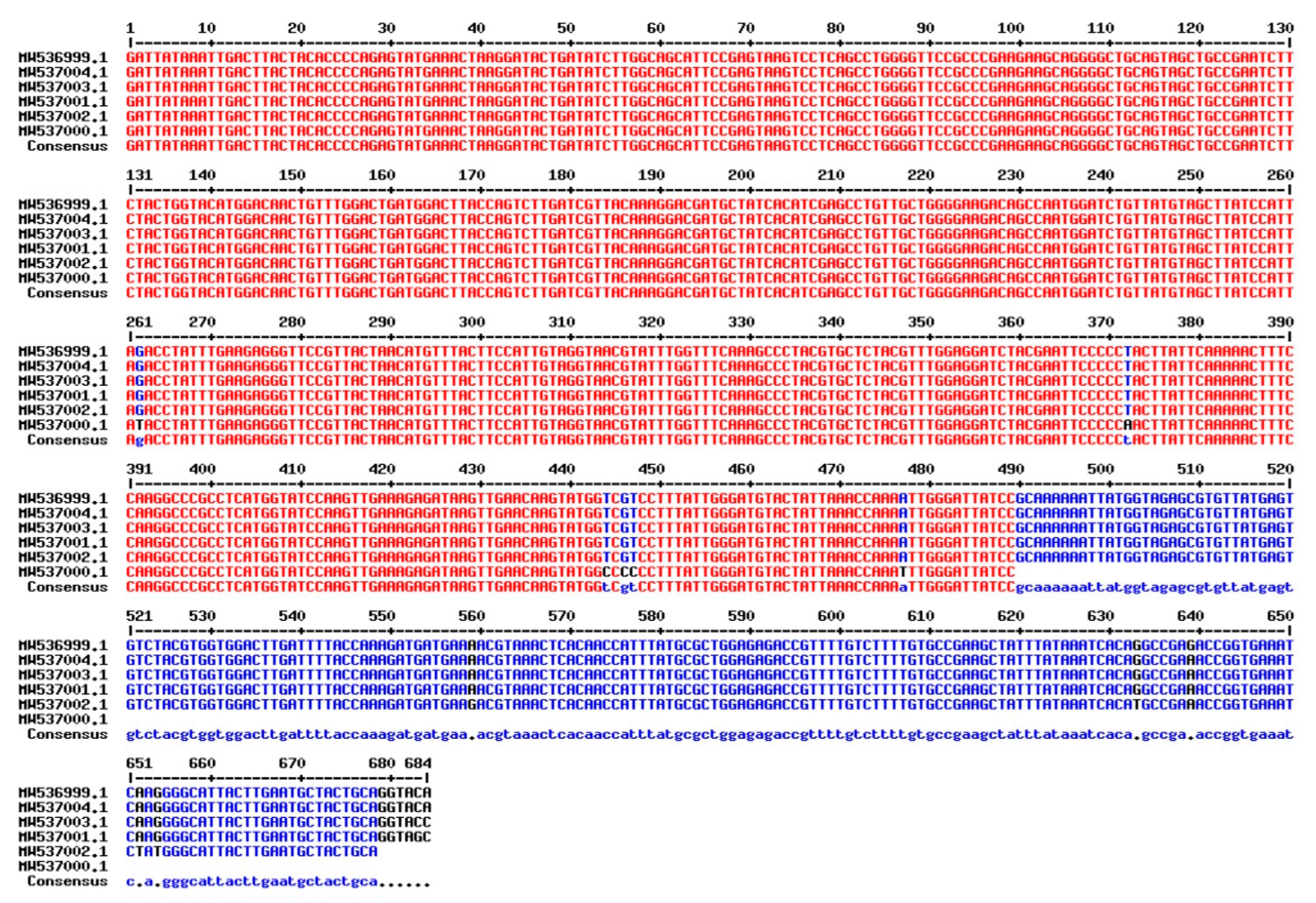
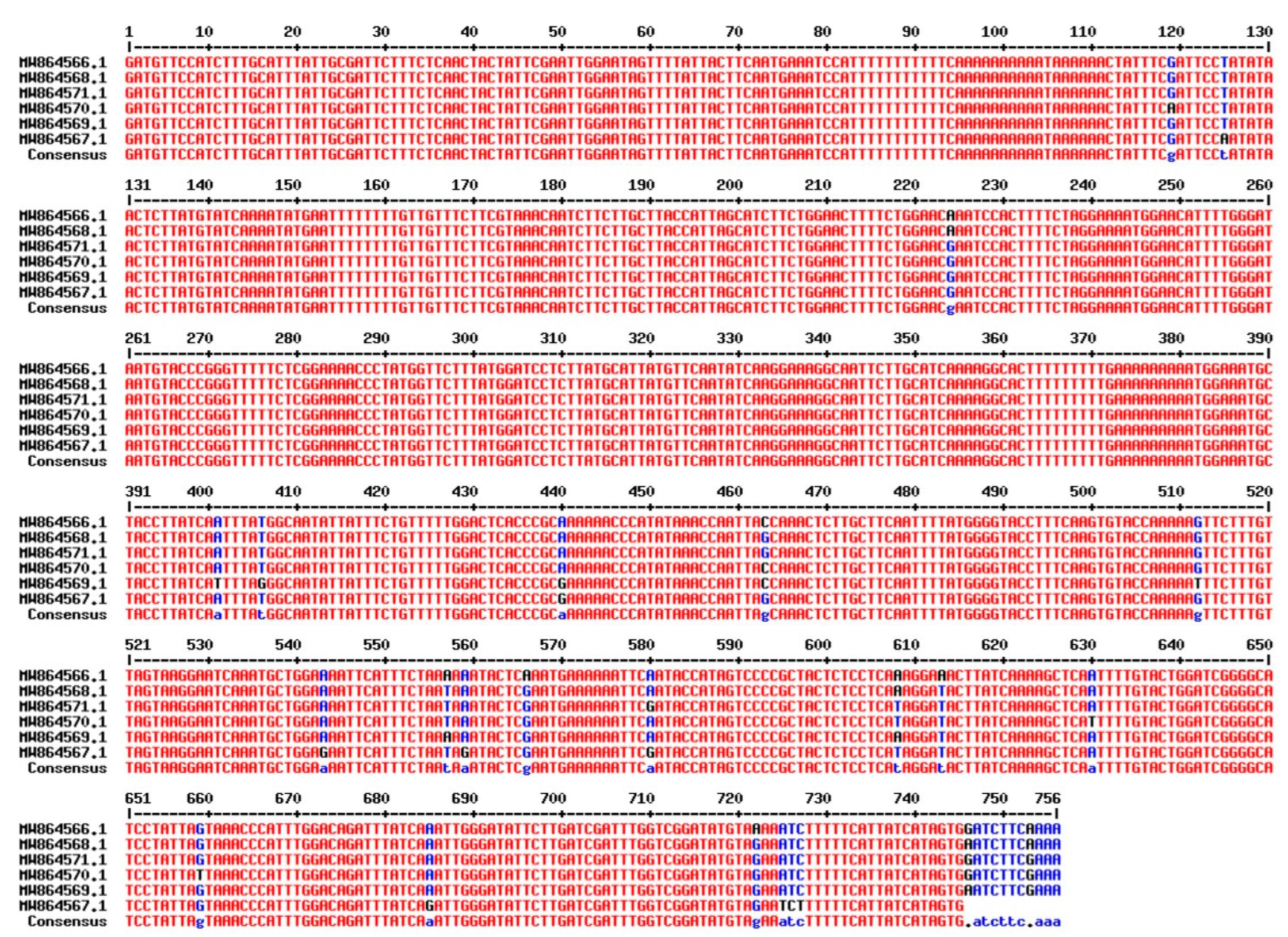
3.4. DNA Barcoding: Plastid rbcL and matK loci Sequencing
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef] [PubMed]
- Venske, E.; dos Santos, R.S.; Busanello, C.; Gustafson, P.; de Oliveira, A.C. Bread wheat: A role model for plant domestication and breeding. Hereditas 2019, 156, 16. [Google Scholar] [CrossRef] [PubMed]
- Martín-Gómez, J.J.; Rewicz, A.; Goriewa-Duba, K.; Wiwart, M.; Tocino, Á.; Cervantes, E. Morphological Description and Classification of Wheat Kernels Based on Geometric Models. Agronomy 2019, 9, 399. [Google Scholar] [CrossRef]
- Rauf, K.; Rahman, R.; Saeed, A.; Ali, M.; Noureen, F.; Amir, R.; Gul, A. Next-Generation Sequencing in Bread Wheat. In Climate Change and Food Security with Emphasis on Wheat; Ozturk, M., Gul, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 311–320. ISBN 978-0-12-819527-7. [Google Scholar] [CrossRef]
- Shitsukawa, N.; Tahira, C.; Kassai, K.; Hirabayashi, C.; Shimizu, T.; Takumi, S.; Mochida, K.; Kawaura, K.; Ogihara, Y.; Murai, K. Genetic and epigenetic alteration among three homoeologous genes of a class E MADS box gene in hexaploid wheat. Plant Cell. 2007, 19, 1723–1737. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Perez, E.; Shaw, P.; Moore, G. The Ph1 locus is needed to ensure specific somatic and meiotic centromere association. Nature 2001, 411, 204–207. [Google Scholar] [CrossRef]
- Charmet, G. Wheat domestication: Lessons for the future. Comptes Rendus Biol. 2011, 334, 212–220. [Google Scholar] [CrossRef]
- FAO: Food and Agriculture Organization of United Nations. 2020. Available online: http://www.fao.org/statistics (accessed on 8 November 2022).
- FAO: Global information and early warning system on food and agriculture (GIEWS) Country Brief Egypt. Available online: https://www.fao.org/giews/countrybrief/country.jsp?code=EGY&lang=ar (accessed on 16 November 2021).
- Fu, Y.B.; Yang, M.H.; Zeng, F.; Biligetu, B. Searching for an accurate marker-based prediction of an individual quantitative trait in molecular plant breeding. Front. Plant Sci. 2017, 8, 1182. [Google Scholar] [CrossRef]
- Sharma, D.; Prajapat, R.K.; Mathur, M.; Upadhyay, T.K.; Maloo, S.R.; Joshi, A.; Surolia, D.K.; Dadarwal, R.S.; Khatik, N. Molecular Marker-Assisted Breeding for Crop Improvement. In Crop Improvement: Biotechnological Advances; Kumar, P., Kumar, T.A., Eds.; Taylor and Francis Group: Boca Raton, FL, USA, 2021; pp. 126–134. [Google Scholar] [CrossRef]
- Jiang, G. Molecular Markers and Marker-Assisted Breeding in Plants. In Plant Breeding from Laboratories to Fields; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef]
- Kadirvel, P.; Senthilvel, S.; Geethanjali, S.; Sujatha, M.; Varaprasad, K.S. Genetic markers, trait mapping and marker-assisted selection in plant breeding. In Plant Biology and Biotechnology; Bahadur, B., Venkat Rajam, M., Sahijram, L., Krishnamurthy, K., Eds.; Springer: New Delhi, India, 2015. [Google Scholar]
- Khayatnezhad, M.; Zaefizadeh, M.; Gholamin, R.; Somarin, S.j.; Mahmoodabad, R.Z. Study of Morphological Traits of Wheat Cultivars Through Factor Analysis. Am-Euras. J. Agric. Environ. Sci. 2010, 9, 460–464. [Google Scholar]
- Ghodke, P.H.; Ramakrishnan, S.; Shirsat, D.V.; Vani, G.K.; Arora, A. Morphological characterization of wheat genotypes for stay green and physiological traits by multivariate analysis under drought stress. Plant Physiol. Rep. 2019, 24, 305–315. [Google Scholar] [CrossRef]
- Varsha, J.; Verma, P.; Saini, P.; Singh, V.; Yashvee, S. Genetic variability of wheat (Triticum aestivum L.) genotypes for agro-morphological traits and their correlation and path analysis. J. Pharmacogn. Phytochem. 2019, 8, 2290–2294. [Google Scholar]
- Luković, K.; Prodanović, S.; Perišić, V.; Milovanović, M.; Perišić, V.; Rajičić, V.; Zečević, V. Multivariate analysis of morphological traits and the most important productive traits of wheat in extreme wet conditions. Appl. Ecol. Environ. Res. 2020, 18, 5857–5871. [Google Scholar] [CrossRef]
- Ouaja, M.; Bahri, B.A.; Aouini, L.; Ferjaoui, S.; Medini, M.; Marcel, T.C.; Hamza, S. Morphological characterization and genetic diversity analysis of Tunisian durum wheat (Triticum turgidum var. durum) accessions. BMC Genom. Data 2021, 22, 3. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Abdullah; Zubaida, B.; Amin, N.; Khan, R.I.; Shafqat, N.; Masood, R.; Waseem, S.; Tahir, J.; Ahmed, I.; et al. Agro-Morphological, Yield, and Genotyping-by-Sequencing Data of Selected Wheat (Triticum aestivum) Germplasm from Pakistan. Front. Genet. 2021, 12, 617–772. [Google Scholar] [CrossRef]
- Siyal, A.L.; Chang, A.G.; Shaikh, N.; Sootaher, J.K.; Jatt, T.; Siyal, F.K.; Chang, M.S. Screening of Wheat Genotypes for Morphological, Physiological and Phenological Traits Under Climatic Condition. Eur. J. Biol. Biotechnol. 2021, 2, 87–91. [Google Scholar] [CrossRef]
- Thompson, D.W. On Growth and Form; Cambridge University Press: London, UK, 1917; p. 1116. [Google Scholar]
- Falconer, D.S. Introduction to Quantitative Genetics, 2nd ed.; Longmans Green: London, UK, 1981; pp. 1–133. [Google Scholar]
- Taneva, K.; Bozhanova, V.; Petrova, I. Variability, heritability and genetic advance of some grain quality traits and grain yield in durum wheat genotypes. Bulg. J. Agric. Sci. 2019, 25, 288–295. [Google Scholar]
- Grover, A.; Sharma, P.C. Development and use of molecular markers: Past and present. Crit. Rev. Biotechnol. 2016, 36, 290–302. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Maackill, D.J. Start Codon Targted (SCoT) polymorphism: A simple novel DNA marker technique for generating gene–targeted markers in plants. Plant Mol. Bio. 2009, 27, 86–93. [Google Scholar] [CrossRef]
- Sofalian, O.; Chaparzadeh, N.; Dolati, M. Genetic diversity in spring wheat landraces from north-west of Iran assessed by ISSR. Bot. Hort. Cluj 2009, 37, 252–256. [Google Scholar]
- Gowayed, S.M.H.; Abd El-Moneim, D. Detection of genetic divergence among some wheat (Triticum aestivum L.) genotypes using molecular and biochemical indicators under salinity stress. PLoS ONE 2021, 16, e0248890. [Google Scholar] [CrossRef]
- Pasqualone, A.; Lotti, C.; Bruno, A.; Vita, P.; Fonzo, N.; Blanco, A. Use of ISSR markers for cultivar identification in durum wheat. Options Mediterraneennes. Ser. A Sem. Medit. 2000, 40, 157–161. [Google Scholar]
- Etminan, A.; Pour-Aboughadareh, A.; Mohammadi, R.; Ahmadi-Rad, A.; Noori, A.; Mahdavian, Z. Applicability of start codon targeted (SCoT) and inter-simple sequence repeat (ISSR) markers for genetic diversity analysis in durum wheat genotypes. Biotechnol. Biotechnol. Equip. 2016, 30, 1075–1081. [Google Scholar] [CrossRef]
- Nosair, H.R. Genetic diversity studies on seven Egyptian wheat (Triticum aestivum L.) cultivars using SCoT and ISSR polymorphism markers. Taeckholmia 2020, 40, 143–151. [Google Scholar] [CrossRef]
- Shaban, A.S.; Arab, S.A.; Basuoni, M.M.; Abozahra, M.S.; Abdelkawy, A.M.; Mohamed, M.M. SCoT, ISSR, and SDS-PAGE Investigation of Genetic Diversity in Several Egyptian Wheat Genotypes under Normal and Drought Conditions. Int. J. Agron. 2022, 2022, 7024028. [Google Scholar] [CrossRef]
- Chen, S.; Yao, H.; Han, J.; Liu, C.; Song, J.; Shi, L.; Zhu, Y.; Ma, X.; Gao, T.; Pang, X. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS ONE 2010, 5, e8613. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Luo, Z.; Cui, S.; Qiao, J.; Zhang, Z.; Zhang, L.; Fu, J.; Ma, X. Molecular Identification and Targeted Quantitative Analysis of Medicinal Materials from Uncaria Species by DNA Barcoding and LC-MS/MS. Molecules 2019, 24, 175. [Google Scholar] [CrossRef]
- Ganopoulos, I.; Kapazoglou, A.; Bosmali, I.; Xanthopoulou, A.; Nianiou-Obeidat, I.; Tsaftaris, A.; Madesis, P. Application of the ITS2 region for barcoding plants of the genus Triticum L. and Aegilops L. Cereal Res. Commun. 2017, 45, 381–389. [Google Scholar] [CrossRef]
- Viglietti, G.; Galla, G.; Porceddu, A.; Barcaccia, G.; Curk, F.; Luro, F.; Scarpa, G.M. Karyological Analysis and DNA Barcoding of Pompia Citron: A First Step toward the Identification of Its Relatives. Plants 2019, 8, 83. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Williams, J.G.K.; Kubelk, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucl. Acid. Res. 1990, 18, 6231–6235. [Google Scholar] [CrossRef]
- Liu, B.H. Statistical Genomics: Linkage, Mapping and QTL Analysis; CRC Press: Boca Raton, FL, USA, 1998. [Google Scholar]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Tessier, C.; David, J.; This, P.; Boursiquot, J.M.; Charrier, A. Optimization of the choice of molecular markers for varietal identification in Vitis vinifera L. Theor. Appl. Genet. 1999, 98, 171–177. [Google Scholar] [CrossRef]
- Prevost, A.; Wilkinson, M.J. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet. 1999, 98, 107–112. [Google Scholar] [CrossRef]
- Utz, F.H. PLABSTAT: A Computer Program for Statistical Analysis of Plant Breeding Experiments; University of Hohenheim: Stuttgart, Germany, 2001. [Google Scholar]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. iMEC: Online marker efficiency calculator. Appl. Plant Sci. 2018, 6, e01159. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]
- Trethowan, R.M. Defining a genetic ideotype for crop improvement. Methods Mol. Biol. 2014, 1145, 1–20. [Google Scholar]
- Hyles, J.; Bloomfield, M.T.; Hunt, J.R.; Trethowan, R.M.; Trevaskis, B. Phenology and related traits for wheat adaptation. Heredity 2020, 125, 417–430. [Google Scholar] [CrossRef]
- Narayanan, S.; Vara, P.P. Characterization of a spring wheat association mapping panel for root traits. Agron. J. 2014, 106, 1593–1604. [Google Scholar] [CrossRef]
- Chen, Y.; Palta, J.; Prasad, P.V.P.; Siddique, K.H.M. Phenotypic variability in bread wheat root systems at the early vegetative stage. BMC Plant Biol. 2020, 20, 185. [Google Scholar] [CrossRef]
- Khodadadi, M.; Fotokian, M.H.; Miransari, M. Genetic diversity of wheat (Triticum aestivum L.) genotypes based on cluster and principal component analyses for breeding strategies. Aust. J. Crop Sci. 2011, 5, 17. [Google Scholar]
- Lynch, J. Root architecture and plant productivity. Plant Physiol. 1995, 109, 7. [Google Scholar] [CrossRef] [PubMed]
- Waines, J.G.; Ehdaie, B. Domestication and crop physiology: Roots of green revolution wheat. Ann Bot. 2007, 100, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Du, Y.; Wang, J.; Wu, A.; Qiao, S.; Xu, B.; Zhang, S.; Siddique, K.H.M.; Chen, Y. Moderate drought stress affected root growth and grain yield in old, modern and newly released cultivars of winter wheat. Front Plant Sci. 2017, 8, 672. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, J.A.; Wingen, L.U.; Griffiths, M.; Pound, M.P.; Gaju, O.; Foulkes, M.J.; Gouis, J.L.; Griffiths, S.; Bennett, M.J.; King, J.; et al. Phenotyping pipeline reveals major seedling root growth QTL in hexaploid wheat. J. Exp. Bot. 2015, 66, 2283–2292. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.S.; Reynolds, M.P. Common genetic basis for canopy temperature depression under heat and drought stress associated with optimized root distribution in bread wheat. Theor. Appl. Genet. 2015, 128, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Fernando, K.M.C.; Mayes, S.; Sparkes, D.L. Identifying seedling root architectural traits associated with yield and yield components in wheat. Ann. Bot. 2017, 119, 1115–1129. [Google Scholar] [CrossRef]
- Goriewa-Duba, K.; Duba, A.; Wachowska, U.; Wiwart, M. An Evaluation of the Variation in the Morphometric Parameters of Grain of Six Triticum Species with the Use of Digital Image Analysis. Agronomy 2018, 8, 296. [Google Scholar] [CrossRef]
- Rovner, I.; Gyulai, F. Computer-Assisted Morphometry: A New Method for Assessing and Distinguishing Morphological Variation in Wild and Domestic Seed Populations. Econ. Bot. 2007, 61, 154–172. [Google Scholar] [CrossRef]
- Sonka, M.; Hlavac, V.; Boyle, R. Image Processing Analysis and Machine Vision, 3rd ed.; Thomson Engineering: Toronto, ON, Canada, 2008; p. 850. [Google Scholar]
- Songsri, P.; Joglloy, S.; Kesmala, T.; Vorasoot, N.; Akkasaeng, C.P.A.; Holbrook, C. Heritability of Drought Resistance Traits and Correlation of Drought Resistance and Agronomic Traits in Peanut. Crop Sci. 2008, 48, 2245–2253. [Google Scholar] [CrossRef]
- Regmi, S.; Poudel, B.; Ojha, B.R.; Kharel, R.; Joshi, P.; Khanal, S.; Kandel, B.P. Estimation of Genetic Parameters of Different Wheat Genotype Traits in Chitwan, Nepal. Int. J. Agron. 2021, 2021, 6651325. [Google Scholar] [CrossRef]
- Lukina, E.V.; Freeman, K.W.; Wynn, K.J.; Thomason, W.E.; Mullen, R.W.; Stone, M.L.; Solie, J.B.; Klatt, A.R.; Johnson, G.V.; Elliot, R.L.; et al. Nitrogen Fertilizer Optimization Algorithm Based on in Season Estimates of Yield and Plant Nitrogen Uptake. J. Plant Nutr. 2001, 24, 885–898. [Google Scholar] [CrossRef]
- Hossain, M.; Azad, A.K.; Alam, S.; Eaton, T.E.-J. Estimation of Variability, Heritability and Genetic Advance for Phenological, Physiological and Yield Contributing Attributes in Wheat Genotypes under Heat Stress Condition. Am. J. Plant Sci. 2021, 12, 586–602. [Google Scholar] [CrossRef]
- Maniee, M.; Kahrizi, D.; Mohammadi, R. Genetic Variability of Some Morpho-Physiological Traits in Durum Wheat (Triticum durum Desf.). J. Appl. Sci. 2009, 9, 1383–1387. [Google Scholar] [CrossRef]
- Khan, N.; Naqvi, F.N. Heritability of Morphological Traits in Bread Wheat Advanced Lines under Irrigated and Non-Irrigated Conditions. Asian J. Agric. Sci. 2011, 3, 215–222. [Google Scholar]
- Sharma, A.K.; Grag, D.K. Genetic Variability in Wheat (Triticum aestivum L.) Crosses under Different Normal and Saline Environments. Ann. Agric. Sci. 2002, 23, 497–499. [Google Scholar]
- Sgro, C.; Hoffmann, A. Genetic correlations, tradeoffs and environmental variation. Heredity 2004, 93, 241–248. [Google Scholar] [CrossRef]
- Arnall, B.; Raun, W.R.; Solie, J.B.; Stone, M.L.; Johnson, G.V.; Girma, K.; Freeman, K.W.; Teal, R.K.; Martin, K.L. Relationship between Coefficient of Variation Measured by Spectral Reflectance and Plant Density at Early Growth Stages in Winter Wheat. J. Plant Nutr. 2006, 29, 1983–1997. [Google Scholar] [CrossRef]
- Ahmed, A.A.M.; Mohamed, E.A.; Hussein, M.Y.; Sallam, A. Genomic regions associated with leaf wilting traits under drought stress in spring wheat at the seedling stage revealed by GWAS. Environ. Exp. Bot. 2021, 184, 104393. [Google Scholar] [CrossRef]
- Mukherjee, S.; Gupta, S.; Maji, A.; Gupta, S.; Bhowmik, N. Character association and path coefficient analysis of wheat (Triticum aestivum L.) genotypes under late sown condition. Environ. Ecol. 2008, 26, 2218–2220. [Google Scholar]
- Sarkar, M.; Naher, J.; Hasan, A.T.; Nazim, T.M.; Nath, U.K. Genetic-environment interaction and genetic advance of superior wheat mutants by Francis and Kannenberg’s method. Prog. Agric. 2019, 30, 20–31. [Google Scholar] [CrossRef]
- Mohi-Ud-Din, M.; Hossain, M.A.; Rohman, M.M.; Uddin, M.N.; Haque, M.S.; Ahmed, J.U.; Hossain, A.; Hassan, M.M.; Mostofa, M.G. Multivariate Analysis of Morpho-Physiological Traits Reveals Differential Drought Tolerance Potential of Bread Wheat Genotypes at the Seedling Stage. Plants 2021, 10, 879. [Google Scholar] [CrossRef] [PubMed]
- El-Aref, H.M.; Taghian, A.S.; El-Sayed, B.E.A.; El-Rawy, M.A.; El-Sanousy, S.A. Genetic relationships among wheat varieties based on ISSR markers. Res. J. Appl. Biotechnol. 2016, 2, 117–126. [Google Scholar] [CrossRef][Green Version]
- Carvalho, A.; Lima-Brito, J.; Maçãs, B.; Guedes-Pinto, H. Genetic diversity and variation among botanical varieties of old Portuguese wheat cultivars revealed by ISSR assays. Biochem. Genet. 2009, 47, 276–294. [Google Scholar] [CrossRef]
- Abd El-Moneim, D. Characterization of ISSR and SCoT markers and TaWRKY gene expression in some Egyptian wheat genotypes under drought Stress. JPPS 2019, 8, 31–46. [Google Scholar]
- Thudi, M.; Manthena, R.; Wani, S.P.; Tatikonda, L.; Hoisington, D.A.; Varshney, R.K. Analysis of genetic diversity in pongamia [Pongamia pinnata (L) pierrre] using AFLP markers. J. Plant Biochem. Biotechnol. 2010, 19, 209–216. [Google Scholar] [CrossRef]
- Peng, J.H.; Lapitan, N.L.V. Characterization of EST derived microsatellites in the wheat genome and development of ESSR markers. Funct. Integr. Genom. 2005, 5, 80–96. [Google Scholar] [CrossRef]
- Ramadugu, C.; Keremane, M.L.; Hu, X.; Karp, D.; Federici, C.T.; Kahn, T.; Lee, R.F. Genetic analysis of citron (Citrus medica L.) using simple sequence repeats and single nucleotide polymorphisms. Sci. Hortic. 2015, 195, 124–137. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Ahmadi, J.; Mehrabi, A.A.; Etminan, A.; Moghaddam, M. Insight into the genetic variability analysis and relationships among some Aegilops and Triticum species, as genome progenitors of bread wheat, using SCoT markers. Plant Biosyst. 2018, 152, 694–703. [Google Scholar] [CrossRef]
- Khodaee, L.; Azizinezhad, R.; Etminan, A.R.; Khosroshahi, M. Assessment of genetic diversity among Iranian Aegilops triuncialis accessions using ISSR, SCoT, and CBDP markers. J. Genet. Eng. Biotechnol. 2021, 19, 5. [Google Scholar] [CrossRef]
- Hamidi, H.; Talebi, R.; Keshavarzi, F. Comparative efficiency of functional gene-based markers, start codon targeted polymorphism (SCoT) and conserved DNA-derived polymorphism (CDDP) with ISSR markers for diagnostic fingerprinting in wheat (Triticum aestivum L.). Cereal Res. Commun. 2014, 42, 558–567. [Google Scholar] [CrossRef]
- Alshehri, M.A. Genetic variation of Saudi Wheat Genotypes through ISSR and SCoT Assays. Int. J. Pharm. Phytopharm. Res. 2019, 9, 57–65. [Google Scholar]
- Gregorczyk, A.; Smagacz, J.; Stankowski, S.; Fiejtek, A. Assessment of relations between the technological characteristics of winter wheat. Acta Sci. Pol. Agric. 2008, 7, 65–71. [Google Scholar]
- Golparvar, A.R.; Ghasemi-Pirbalouti, A.; Madani, H. Genetic control of some physiological attributes in wheat under drought stress conditions. Pak. J. Bio. Sci. 2006, 9, 1442–1446. [Google Scholar]
- Beheshtizadeh, H.; Rezaie, A.; Rezaie, A.; Ghandi, A. Principal component analysis and determination of the selection criteria in bread wheat (Triticum aestivum L.) genotypes. Int. J. Agric. Crop Sci. 2013, 5, 2024–2027. [Google Scholar]
- Adilova, S.S.; Qulmamatova, D.E.; Baboev, S.K.; Bozorov, T.A.; Morgunov, A.I. Multivariate Cluster and Principal Component Analyses of Selected Yield Traits in Uzbek Bread Wheat Cultivars. Am. J. Plant Sci. 2020, 11, 903–912. [Google Scholar] [CrossRef]
- CBOL Plant Working Group. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef] [PubMed]
- Pahlavan, A.; Solouki, M.; Fakheri, B.; Fazeli-Nasab, B. Using Morphological and Phytochemical Traits and ITS (1, 4) and rbcl DNA Barcodes in the Assessment of Different Malva sylvestris L. Genotypes. J. Med. Plants By-Prod. 2021, 1, 19–35. [Google Scholar]
- Sarhan, S.; Hamed, F.; Al-Youssef, W. The rbcL Gene Sequence Variations among and within Prunus Species. J. Agric. Sci. Technol. 2016, 18, 1105–1115. [Google Scholar]
- Hapsari, L.; Trimanto, T.; Wahyudi, D. Species diversity and phylogenetic analysis of Heliconia spp. collections of Purwodadi Botanic Garden (East Java, Indonesia) inferred by rbcL gene sequences. Biodivers. J. Biol. Divers. 2019, 20, 1266–1283. [Google Scholar] [CrossRef]
- Newmaster, S.; Fazekas, A.; Steeves, R.; Janovec, J. Testing candidate plant barcode regions in the Myristicaceae. Mol. Ecol. Resour. 2008, 8, 480–490. [Google Scholar] [CrossRef]
- Young, N.D.; de Pamphilis, C.W. Purifying selection detected in the plastid gene matK and flanking ribozyme regions within a group II intron of nonphotosynthetic plants. Mol. Biol. Evol. 2000, 17, 1933–1941. [Google Scholar] [CrossRef] [PubMed]
- Federici, S.; Galimberti, A.; Bartolucci, F.; Bruni, I.; De Mattia, F.; Cortis, P.; Labra, M. DNA barcoding to analyse taxonomically complex groups in plants: The case of Thymus (Lamiaceae). Bot. J. Linn. Soc. 2013, 171, 687–699. [Google Scholar] [CrossRef]
- Mohamed, A.H.; Omar, A.A.; Attya, A.M.; Elashtokhy, M.M.A.; Zayed, E.M.; Rizk, R.M. Morphological and Molecular Characterization of Some Egyptian Six-Rowed Barley (Hordeum vulgare L.). Plants 2021, 10, 2527. [Google Scholar] [CrossRef] [PubMed]
- Osman, S.A.; Ramadan, W.A. DNA barcoding of different Triticum species. Bull. Natl. Res. Cent. 2019, 43, 174. [Google Scholar] [CrossRef]
- Burgess, K.S.; Fazekas, A.J.; Kesanakurti, P.R.; Graham, S.W.; Husband, B.C.; Newmaster, S.G.; Percy, D.M.; Hajibabaei, M.; Barrett, S.C.H. Discriminating plant species in a local temperate flora using the rbcL + matK DNA barcode. Methods Ecol. Evol. 2011, 2, 333–340. [Google Scholar] [CrossRef]
- Awad, M.; Fahmy, R.M.; Mosa, K.A.; Helmy, M.; El-Feky, F.A. Identification of Effective DNA Barcodes for Triticum Plants through Chloroplast Genome-wide Analysis. Comput. Biol. Chem. 2017, 71, 20–31. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Name | Pedigree | Selection History | Year of Release |
|---|---|---|---|---|
| W1 | SIDS 1 | HD 2172/Pavon “S”//1158.57/Maya 74 “S” | S 46-4SD-2SD-1SD-0SD-0EGY | 1994 |
| W2 | SIDS 12 | BUC//7C/ALD/5/MAYA74/ON//1160.147/3/BB/GLL/4/CHAT”S”/6/MAYA/VUL//CMH74A.630/4*SX | SD7096-4SD-1SD-1SD-0SD-0EGY | 2009 |
| W3 | SIDS 14 | BOW “S”/VEE”S”//BOW”S”/TSI/3/BANI SEWEF 1 | SD293-1SD-2SD-4SD-0SD-0EGY | - |
| W4 | SHANDWEEL 1 | SITE/MO/4/NAC/TH.AC//3*PVN/3/MIRLO/BUC | CMSS93B00567S-72Y-010M-010Y-010M-3Y-0M-0HTY-0SH-0EGY | 2011 |
| W5 | SAKHA 94 | OPATA/RAYON//KAUZ | CMBW90Y3180-0TOPM-3Y-010M-010M-010Y-10M-015Y-0Y-0AP-0S-0EGY | 2004 |
| W6 | SAKHA 95 | PASTOR//SITE/MO/3/CHEN/AEGILOPS SQUARROSA (TAUS)//BCN/4/WBLL1. | CMA01Y00158S-040POY-040M-030ZTM-040SY-26M-0Y-0SY-0S-0EGY | - |
| W7 | GIZA 171 | SAKHA 93/GEMMIZA 9 | S.6-1GZ-4GZ-1GZ-2GZ-0EGY | 2013 |
| W8 | GIZA 168 | MRL/BUC//SERI | CM93046-8M-0Y-0M-2Y-0B-0SH-0EGY | 1999 |
| W9 | GEMMIZA 7 | CMH 74A.630/5X//SERI 82/3/AGENT | GM 4611-2GM-3GM-1GM-0GM-0EGY | 2000 |
| W10 | GEMMIZA 9 | ALD “S”/HUAC//CMH 74A. 630/5X | GM 4583-5GM-1GM-0GM-0EGY | 2000 |
| W11 | GEMMIZA 10 | MAYA 74 “S”/ON//1160-147/3/BB/GLL/4/CHAT”S”/5/CROW “S” | CGM5820-3GM-1GM-2GM-0GM-0EGY | 2004 |
| W12 | GEMMIZA 11 | BOW”S”/KVZ”S”//7C/SER182/3/GIZA168/SAKHA61 | GM7892-2GM-1GM-2GM-1GM-0GM-0EGY | 2011 |
| W13 | GEMMIZA 12 | OTUS/3/SARA/THB//VEE | CMSS97Y00227S-5Y-010M-010Y-010M-2Y-1M-0Y-0GM-0EGY | 2013 |
| W14 | MISR 1 | OASIS/SKAUZ//4*BCN/3/2*PASTOR | CMSS00Y01881T-050M-030Y-030M-030WGY-33M-0Y-0EGY | 2014 |
| W15 | MISR 2 | SKAUZ/BAV92 | CMSS96M03611S-1M-010SY-010M-010SY-8M-0Y-0EGY | 2014 |
| W16 | MISR 3 | ATTILA*2/PBW65*2/KACHU | CMSS06Y00582T-099TOPM-099Y-099ZTM-099Y-099M-10WGY-0B-0EGY | 2019 |
| Primer Code | Sequence (5′-3′) | Size (bp) |
|---|---|---|
| ISSR marker | ||
| ISSR-807 | (AG)8 T | 172–1156 |
| ISSR-810 | (GA)8 T | 159–2949 |
| ISSR-835 | (Ag)8 YC | 96–4039 |
| ISSR-841 | (GA)8 YC | 102–420 |
| ISSR-857 | (AC)8 YG | 84–244 |
| ISSR-825 | (AC)7 T | 327–2562 |
| ISSR-814 | (CT)7 CAT | 215–496 |
| ISSR-826 | (AC)8 C | 223–1808 |
| ISSR-827 | (AC)8 G | 255–1408 |
| ISSR-840 | (gA)8 TT | 123–1164 |
| SCoT marker | ||
| SCoT 1 | CAACAATGGCTACCACCC | 513–3099 |
| SCoT 2 | ACCATGGCTACCACCGGC | 296–3658 |
| SCoT 3 | CAACAATGGCTACCACGC | 183–2079 |
| SCoT 4 | CAACAATGGCTACCACCG | 177–3009 |
| SCoT 5 | ACGACATGGCGACCACGC | 142–669 |
| SCoT 6 | CCATGGCTACCACCGCAG | 331–2274 |
| Plastid rbcL and matK genes | ||
| rbcL | F: 5′-ATGTCACCACAAACAGAGACTAAAGC-3′ | 600 |
| R: 5′-TCGCATGTACCTGCAGTAGC-3′ | ||
| matK | F: 5′-CGATCTATTCATTCAATATTTC-3′ | 900 |
| R: 5′-TCTAGCACACGAAAGTCGAAGT-3′ | ||
| Morphometric and Geometric Traits | Average | Max. | Min. |
|---|---|---|---|
| Seedling parameters | |||
| PB | 0.12 | 0.20 | 0.07 |
| Shoot parameters | |||
| SL | 14.73 | 18.26 | 7.90 |
| LFI | 3.15 | 5.16 | 1.12 |
| DFI | 0.18 | 0.23 | 0.15 |
| Leaf Parameters | |||
| LA | 2.92 | 4.48 | 1.13 |
| LW | 0.32 | 0.42 | 0.24 |
| Root parameters | |||
| RN | 4.19 | 6.00 | 3.00 |
| RL | 8.60 | 11.09 | 6.83 |
| RW | 7.03 | 10.23 | 5.07 |
| TA | 35.89 | 48.63 | 27.34 |
| Seed parameters | |||
| SW | 0.05 | 0.06 | 0.03 |
| SA | 0.34 | 0.45 | 0.29 |
| SP | 2.45 | 2.81 | 2.27 |
| L | 0.89 | 1.04 | 0.82 |
| W | 0.49 | 0.61 | 0.43 |
| AR | 1.84 | 2.03 | 1.55 |
| Circ. | 0.72 | 0.75 | 0.69 |
| Round | 0.55 | 0.66 | 0.49 |
| Feret | 0.90 | 1.04 | 0.84 |
| TRAITS | R MS (2) | G MS (15) | RXG MS (30) | RCV% | GCV% | RXG CV (%) | H2% | LSD (5%) | F VALUE OF G |
|---|---|---|---|---|---|---|---|---|---|
| PB | 0.0015 | 0.0032 | 0.0011 | 0.0001 | 0.007 | 0.0003 | 65.33 | 0.06 | 2.88 ** |
| SL | 3.1912 | 24.0514 | 2.7140 | 0.0298 | 7.1125 | 0.6785 | 88.72 | 2.75 | 8.86 ** |
| LFI | 0.0889 | 2.2307 | 0.2609 | 0.0057 | 0.6566 | 0.0652 | 88.30 | 0.85 | 8.55 ** |
| DFI | 0.002 | 0.0014 | 0.0014 | 0.0000 | 0.0000 | 0.0003 | 4.93 | 0.06 | 1.05 |
| LA | 0.0777 | 2.7584 | 0.3378 | 0.0063 | 0.8069 | 0.0844 | 87.76 | 0.97 | 8.17 ** |
| LW | 0.0125 | 0.0079 | 0.0058 | 0.0004 | 0.007 | 0.0014 | 26.91 | 0.13 | 1.37 |
| RN | 0.000 | 2.4875 | 0.000 | 0.0000 | 0.8292 | 0.0000 | 100.00 | 0.00 | 0.00 |
| RL | 2.8541 | 4.8841 | 1.6033 | 0.0782 | 1.0936 | 0.4008 | 67.17 | 2.11 | 3.05 ** |
| RW | 1.2819 | 6.6247 | 4.2020 | 0.0867 | 0.8076 | 1.0505 | 36.57 | 3.42 | 1.58 |
| TA | 164.8038 | 93.6098 | 33.5567 | 8.2029 | 20.0177 | 8.3892 | 64.15 | 9.66 | 2.79 ** |
| SW | 0.000 | 0.0002 | 0.0001 | 0.0000 | 0.0001 | 0.000 | 74.02 | 0.00 | 0.00 |
| SA | 0.0006 | 0.0060 | 0.0010 | 0.0000 | 0.0017 | 0.0002 | 83.77 | 0.05 | 6.16 ** |
| SP | 0.0024 | 0.0638 | 0.0089 | 0.0002 | 0.0183 | 0.0022 | 86.07 | 0.16 | 7.18 ** |
| L | 0.0000 | 0.0082 | 0.0013 | 0.0000 | 0.0023 | 0.0003 | 84.78 | 0.06 | 6.57 ** |
| W | 0.0010 | 0.0052 | 0.0013 | 0.0000 | 0.0013 | 0.0003 | 74.08 | 0.06 | 3.86 ** |
| AR | 0.0099 | 0.0434 | 0.0178 | 0.0005 | 0.0086 | 0.0044 | 59.11 | 0.22 | 2.45 * |
| CIRC. | 0.0001 | 0.0005 | 0.0002 | 0.0000 | 0.0001 | 0.0001 | 58.82 | 0.02 | 2.43 * |
| ROUND | 0.0010 | 0.0047 | 0.0021 | 0.0001 | 0.0009 | 0.0005 | 56.11 | 0.08 | 2.28 * |
| FERET | 0.0001 | 0.0083 | 0.0011 | 0.0000 | 0.0024 | 0.0003 | 86.68 | 0.06 | 7.51 ** |
| Parameters | PB | SL | LFI | DFI | LA | LW | RN | RL | RW | TA | SW | SA | SP | L | W | AR | Circ. | Round | Feret |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PB | 0 | 0.21 | 0.41 | 0.05 | 0.47 | 0.39 | 0.14 | 0.11 | 0.4 | 0.26 | 0.57 * | 0.72 ** | 0.75 ** | 0.74 ** | 0.55 * | 0.04 | −0.07 | 0.03 | 0.76 ** |
| SL | 0.23 | 0 | 0.73 ** | 0.35 | 0.80 ** | 0.4 | −0.67 ** | 0.25 | 0.02 | 0.02 | −0.01 | 0.17 | 0.19 | 0.19 | 0.14 | 0.01 | −0.02 | 0.03 | 0.19 |
| LFI | 0.50 + | 0.78 + | 0 | 0.07 | 0.66 ** | 0.39 | −0.42 | 0.11 | −0.21 | −0.02 | 0.21 | 0.53 * | 0.48 | 0.3 | 0.59 * | −0.37 | 0.35 | 0.44 | 0.44 |
| DFI | −0.22 | 1.36 | −0.49 | 0 | 0.36 | 0.26 | −0.02 | −0.16 | 0.49 | 0.01 | −0.19 | −0.43 | −0.4 | −0.28 | −0.44 | 0.28 | −0.29 | −0.26 | −0.37 |
| LA | 0.56 ++ | 0.84 ++ | 0.75 ++ | 1.74 | 0 | 0.75 ** | −0.45 | 0.17 | 0.26 | −0.1 | −0.06 | 0.45 | 0.46 | 0.44 | 0.37 | −0.05 | 0.04 | 0.1 | 0.47 |
| LW | 0.64 + | 0.83 + | 0.85 + | 1.24 | 1.51 + | 0 | −0.12 | −0.02 | 0.35 | −0.37 | −0.06 | 0.45 | 0.41 | 0.29 | 0.45 | −0.29 | 0.28 | 0.31 | 0.38 |
| RN | 0.17 | −0.72 ++ | −0.44 ++ | −0.1 | −0.48 ++ | −0.24 | 0 | −0.61 * | 0.22 | −0.09 | 0.18 | 0.04 | 0.02 | −0.03 | 0.08 | −0.08 | 0.07 | 0.11 | 0.01 |
| RL | 0.3 | 0.30 + | 0.18 | −0.02 | 0.18 | 0.31 | −0.74 + | 0 | −0.03 | 0.41 | 0.13 | 0.14 | 0.18 | 0.26 | 0.02 | 0.21 | −0.21 | −0.19 | 0.21 |
| RW | 0.75 + | −0.09 | −0.48 + | 3.02 | 0.43 + | 0.45 | 0.37 | 0.05 | 0 | −0.14 | −0.04 | 0.03 | 0.1 | −0.27 | 0.15 | 0.39 | −0.4 | −0.37 | 0.16 |
| TA | 0.48 + | 0.05 | 0.06 | 0.31 | −0.16 | −0.82 + | −0.11 | 0.55 + | 0 | 0 | 0.17 | −0.07 | −0.03 | −0.09 | 0.19 | 0.36 | −0.37 | −0.31 | 0.01 |
| SW | 0.88 ++ | −0.04 | 0.26 | −0.9 | −0.13 | −0.25 | 0.21 | 0.19 | −0.1 | 0.18 | 0 | 0.45 | 0.45 | 0.38 | 0.41 | −0.11 | 0.11 | 0.13 | 0.44 |
| SA | 1.07 ++ | 0.24 | 0.62 ++ | −2.09 | 0.55 ++ | 0.76 + | 0.05 | 0.17 | 0.03 | −0.1 | 0.51 + | 0 | 0.99 ** | 0.82 ** | 0.92 ** | −0.35 | 0.33 | 0.4 | 0.95 ** |
| SP | 1.07 ++ | 0.25 | 0.57 ++ | −1.92 | 0.54 ++ | 0.69 + | 0.02 | 0.21 | 0.14 | −0.05 | 0.50 + | 0.99 ++ | 0 | 0.90 ** | 0.84 ** | −0.19 | 0.17 | 0.24 | 0.99 ** |
| L | 1.01 ++ | 0.23 | 0.37 + | −1.3 | 0.48 ++ | 0.51 + | −0.04 | 0.29 | 0.44 + | 0.11 | 0.40 + | 0.88 ++ | 0.93 ++ | 0 | 0.53* | 0.24 | −0.26 | −0.2 | 0.95 ** |
| W | 0.92 ++ | 0.23 | 0.72 ++ | −2.31 | 0.52 ++ | 0.79 + | 0.09 | 0.06 | −0.29 | −0.26 | 0.52 + | 0.93 ++ | 0.87 ++ | 0.64 ++ | 0 | −0.69 ** | 0.67 ** | 0.73 ** | 0.76 ** |
| AR | −0.05 | −0.03 | −0.47 | 1.65 + | −0.19 | −0.59 | −0.11 | 0.22 | 0.76 + | 0.53 + | −0.22 | 0.31 + | −0.18 | −0.18 | 0.64 ++ | 0 | −1.00 ** | −0.99 ** | −0.05 |
| Circ. | 0 | 0.02 | 0.44 + | −1.66 | 0.17 | 0.56 | 0.09 | −0.22 | −0.76 + | −0.55 + | 0.21 | 0.29 | 0.16 | −0.21 | 0.62 ++ | −1.00 ++ | 0 | 0.98 ** | 0.03 |
| Round | 0.21 | 0.08 | 0.57 ++ | −1.66 | 0.25 | 0.62 | 0.15 | −0.23 | −0.76 + | −0.48 + | 0.24 | 0.38 + | 0.25 | −0.12 | 0.69 ++ | −1.00 ++ | 1.00 ++ | 0 | 0.11 |
| Feret | 1.07 ++ | 0.25 | 0.52 ++ | −1.74 | 0.53 ++ | 0.64 + | 0.01 | 0.24 | 0.24 | 0.01 | 0.47 + | 0.97 ++ | 0.99 ++ | 0.97 ++ | 0.81 ++ | −0.06 | 0.04 | 0.13 | 0 |
| Molecular Marker | MB | Polymorphic Bands | TAB | PPB | MBF | H | PIC | E | MI | H.av | D | Rp | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| UB | NUB | NPB | ||||||||||||
| ISSR-807 | 1 | 7 | 0 | 7 | 8 | 88 | 0.5 | 0.50 | 0.37 | 3.8 | 0.02 | 0.00 | 0.78 | 2.9 |
| ISSR-810 | 2 | 6 | 0 | 6 | 8 | 75 | 0.6 | 0.49 | 0.37 | 4.5 | 0.02 | 0.00 | 0.69 | 3.3 |
| ISSR-835 | 4 | 5 | 2 | 7 | 11 | 64 | 0.7 | 0.44 | 0.34 | 7.4 | 0.02 | 0.00 | 0.55 | 3.6 |
| ISSR-841 | 3 | 3 | 1 | 4 | 7 | 57 | 0.5 | 0.49 | 0.37 | 3.9 | 0.02 | 0.00 | 0.70 | 1.7 |
| ISSR-857 | 0 | 9 | 1 | 10 | 10 | 100 | 0.3 | 0.44 | 0.35 | 3.3 | 0.01 | 0.00 | 0.89 | 4.9 |
| ISSR-825 | 1 | 5 | 0 | 5 | 6 | 83 | 0.5 | 0.50 | 0.37 | 3.1 | 0.02 | 0.01 | 0.73 | 2.7 |
| ISSR-814 | 0 | 2 | 1 | 3 | 3 | 100 | 0.3 | 0.39 | 0.31 | 0.8 | 0.01 | 0.01 | 0.93 | 1.2 |
| ISSR-826 | 3 | 7 | 0 | 7 | 10 | 70 | 0.8 | 0.35 | 0.29 | 7.7 | 0.02 | 0.00 | 0.40 | 2.4 |
| ISSR-827 | 1 | 5 | 2 | 7 | 8 | 88 | 0.5 | 0.50 | 0.37 | 3.7 | 0.02 | 0.00 | 0.78 | 2.3 |
| ISSR-840 | 0 | 7 | 1 | 8 | 8 | 100 | 0.3 | 0.39 | 0.31 | 2.1 | 0.01 | 0.00 | 0.93 | 4.1 |
| Total | 15 | 56 | 8 | 64 | 79 | - | - | 4.50 | 3.47 | 40.4 | 0.15 | 0.04 | 7.38 | 29.2 |
| Mean | 1.5 | 5.6 | 0.8 | 6.4 | 7.9 | 82.41 | 0.49 | 0.45 | 0.35 | 4.04 | 0.01 | 0.00 | 0.74 | 2.92 |
| SCoT 1 | 0 | 8 | 0 | 8 | 8 | 100 | 0.3 | 0.40 | 0.30 | 2.6 | 0.01 | 0.00 | 0.9 | 3.3 |
| SCoT 2 | 2 | 9 | 0 | 9 | 11 | 82 | 0.6 | 0.50 | 0.40 | 6.3 | 0.02 | 0.00 | 0.7 | 5.1 |
| SCoT 3 | 1 | 9 | 1 | 10 | 11 | 91 | 0.3 | 0.50 | 0.40 | 3.9 | 0.01 | 0.00 | 0.9 | 5.7 |
| SCoT 4 | 0 | 11 | 2 | 13 | 13 | 100 | 0.3 | 0.40 | 0.30 | 3.5 | 0.01 | 0.00 | 0.9 | 6.5 |
| SCoT 5 | 0 | 10 | 0 | 10 | 10 | 100 | 0.4 | 0.50 | 0.40 | 4.5 | 0.01 | 0.00 | 0.8 | 7.1 |
| SCoT 6 | 0 | 7 | 4 | 11 | 11 | 100 | 0.2 | 0.40 | 0.30 | 2.8 | 0.01 | 0.00 | 0.9 | 4.9 |
| Total | 3 | 54 | 7 | 61 | 64 | - | - | 2.70 | 2.10 | 23.6 | 0.07 | 0.02 | 5.1 | 32.7 |
| Mean | 0.5 | 9 | 1.17 | 10.17 | 10.70 | 95.46 | 0.36 | 0.40 | 0.30 | 3.9 | 0.01 | 0.00 | 0.9 | 5.4 |
| Total of All | 18 | 110 | 15 | 125 | 143 | - | - | 7.2 | 5.5 | 64.0 | 0.21 | 0.06 | 12.5 | 61.9 |
| Mean of All | 1.2 | 7.33 | 1 | 8.33 | 9.53 | 88.94 | 0.43 | 0.4 | 0.3 | 4.0 | 0.01 | 0.00 | 0.8 | 3.9 |
| Genotypes | W1 | W2 | W3 | W4 | W5 | W6 | W7 | W8 | W9 | W10 | W11 | W12 | W13 | W14 | W15 | W16 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| W1 | 1.00 | |||||||||||||||
| W2 | 0.79 | 1.00 | ||||||||||||||
| W3 | 0.87 | 0.75 | 1.00 | |||||||||||||
| W4 | 0.73 | 0.69 | 0.74 | 1.00 | ||||||||||||
| W5 | 0.81 | 0.79 | 0.82 | 0.68 | 1.00 | |||||||||||
| W6 | 0.60 | 0.63 | 0.61 | 0.51 | 0.71 | 1.00 | ||||||||||
| W7 | 0.71 | 0.67 | 0.71 | 0.67 | 0.69 | 0.71 | 1.00 | |||||||||
| W8 | 0.88 | 0.85 | 0.86 | 0.78 | 0.88 | 0.63 | 0.73 | 1.00 | ||||||||
| W9 | 0.86 | 0.77 | 0.79 | 0.69 | 0.83 | 0.64 | 0.72 | 0.83 | 1.00 | |||||||
| W10 | 0.83 | 0.79 | 0.84 | 0.66 | 0.76 | 0.64 | 0.73 | 0.83 | 0.79 | 1.00 | ||||||
| W11 | 0.79 | 0.78 | 0.80 | 0.74 | 0.82 | 0.68 | 0.79 | 0.81 | 0.77 | 0.76 | 1.00 | |||||
| W12 | 0.80 | 0.73 | 0.80 | 0.63 | 0.77 | 0.73 | 0.79 | 0.81 | 0.73 | 0.89 | 0.78 | 1.00 | ||||
| W13 | 0.71 | 0.72 | 0.67 | 0.56 | 0.72 | 0.67 | 0.65 | 0.73 | 0.72 | 0.72 | 0.72 | 0.78 | 1.00 | |||
| W14 | 0.76 | 0.66 | 0.77 | 0.71 | 0.68 | 0.56 | 0.62 | 0.78 | 0.72 | 0.66 | 0.68 | 0.71 | 0.64 | 1.00 | ||
| W15 | 0.73 | 0.69 | 0.72 | 0.64 | 0.64 | 0.69 | 0.78 | 0.71 | 0.67 | 0.74 | 0.72 | 0.76 | 0.65 | 0.67 | 1.00 | |
| W16 | 0.73 | 0.74 | 0.76 | 0.63 | 0.80 | 0.77 | 0.75 | 0.77 | 0.74 | 0.75 | 0.78 | 0.79 | 0.79 | 0.68 | 0.69 | 1.00 |
| Barcode/Genetic Variability of each Marker | rbcL | matK |
|---|---|---|
| Total alignment length (bp) | 728 | 757 |
| Total matrix cells | 4368 | 4542 |
| Missing percent | 9.501 | 0.132 |
| Number of variable sites | 13 | 24 |
| Proportion of variable sites | 0.018 | 0.032 |
| Number of parsimony informative sites (PIC) | 0.00 | 8 |
| Proportion of Parsimony informative sites | 0.000 | 0.011 |
| AT content | 0.567 | 0.684 |
| GC content | 0.433 | 0.316 |
| A | 1112 | 1429 |
| C | 821 | 795 |
| G | 891 | 641 |
| T | 1129 | 1671 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Esawi, M.A.; Elashtokhy, M.M.A.; Shamseldin, S.A.M.; El-Ballat, E.M.; Zayed, E.M.; Heikal, Y.M. Analysis of Genetic Diversity and Phylogenetic Relationships of Wheat (Triticum aestivum L.) Genotypes Using Phenological, Molecular and DNA Barcoding Markers. Genes 2023, 14, 34. https://doi.org/10.3390/genes14010034
El-Esawi MA, Elashtokhy MMA, Shamseldin SAM, El-Ballat EM, Zayed EM, Heikal YM. Analysis of Genetic Diversity and Phylogenetic Relationships of Wheat (Triticum aestivum L.) Genotypes Using Phenological, Molecular and DNA Barcoding Markers. Genes. 2023; 14(1):34. https://doi.org/10.3390/genes14010034
Chicago/Turabian StyleEl-Esawi, Mohamed A., Mohamed M. A. Elashtokhy, Sahar A. M. Shamseldin, Enas M. El-Ballat, Ehab M. Zayed, and Yasmin M. Heikal. 2023. "Analysis of Genetic Diversity and Phylogenetic Relationships of Wheat (Triticum aestivum L.) Genotypes Using Phenological, Molecular and DNA Barcoding Markers" Genes 14, no. 1: 34. https://doi.org/10.3390/genes14010034
APA StyleEl-Esawi, M. A., Elashtokhy, M. M. A., Shamseldin, S. A. M., El-Ballat, E. M., Zayed, E. M., & Heikal, Y. M. (2023). Analysis of Genetic Diversity and Phylogenetic Relationships of Wheat (Triticum aestivum L.) Genotypes Using Phenological, Molecular and DNA Barcoding Markers. Genes, 14(1), 34. https://doi.org/10.3390/genes14010034