

New Insights into the Identity of the DFNA58 Gene

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Whole Genome Sequencing
2.2. Reuse of Zebrafish and Mouse Transcriptome Data
2.3. Animals
2.3.1. Zebrafish
2.3.2. Mouse
2.4. RNA Extraction and cDNA Synthesis
2.5. Quantitative Real-Time PCR (qPCR)
2.6. Statisical Anaylsis
2.7. In Situ Hybridization Analyses
2.8. Indirect Immunofluorescence
2.9. Western Blotting Cnrip1-Inner Ear Mouse
3. Results
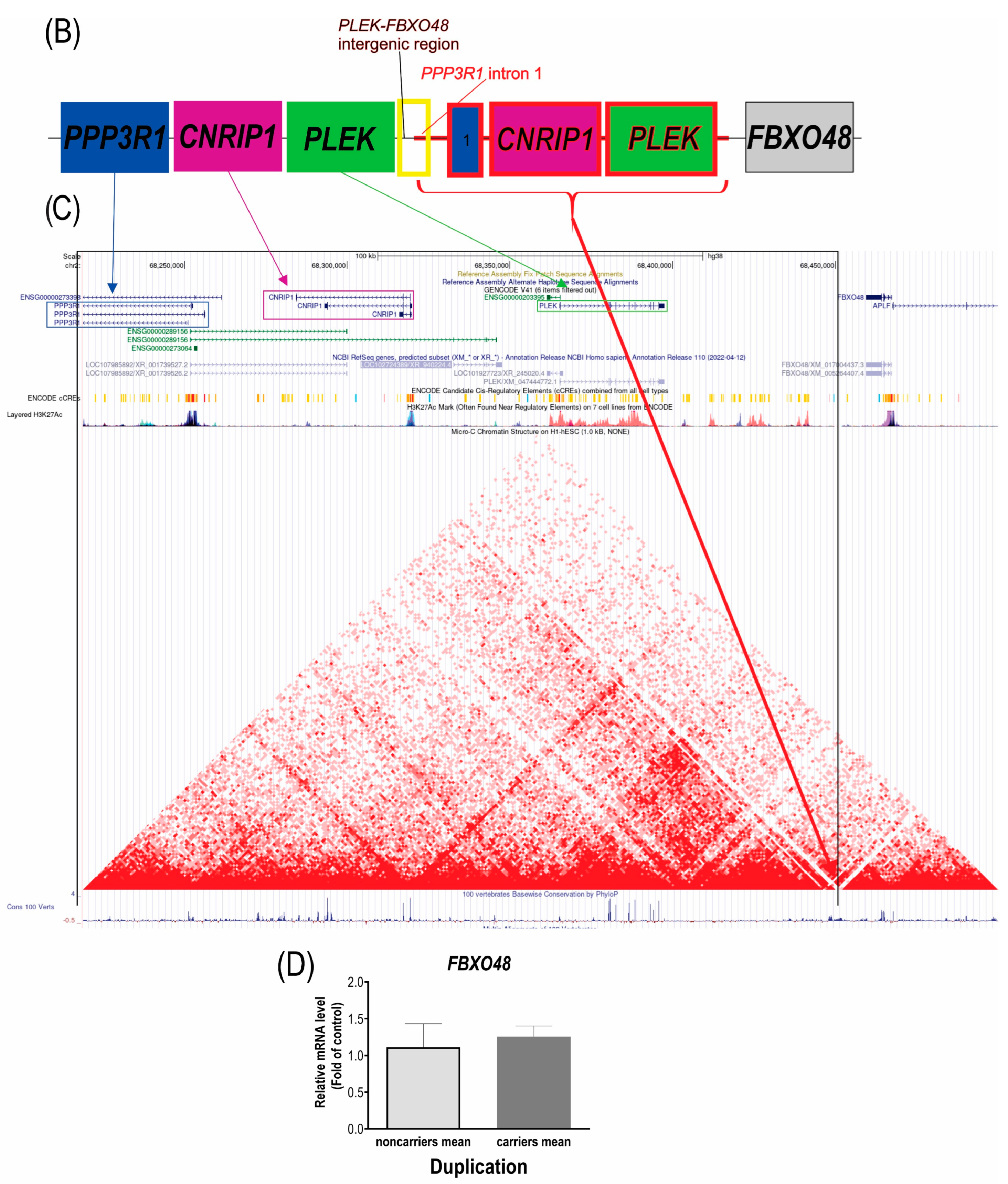
3.1. The Duplication Is in Tandem between the PLEK and FBXO48 Genes
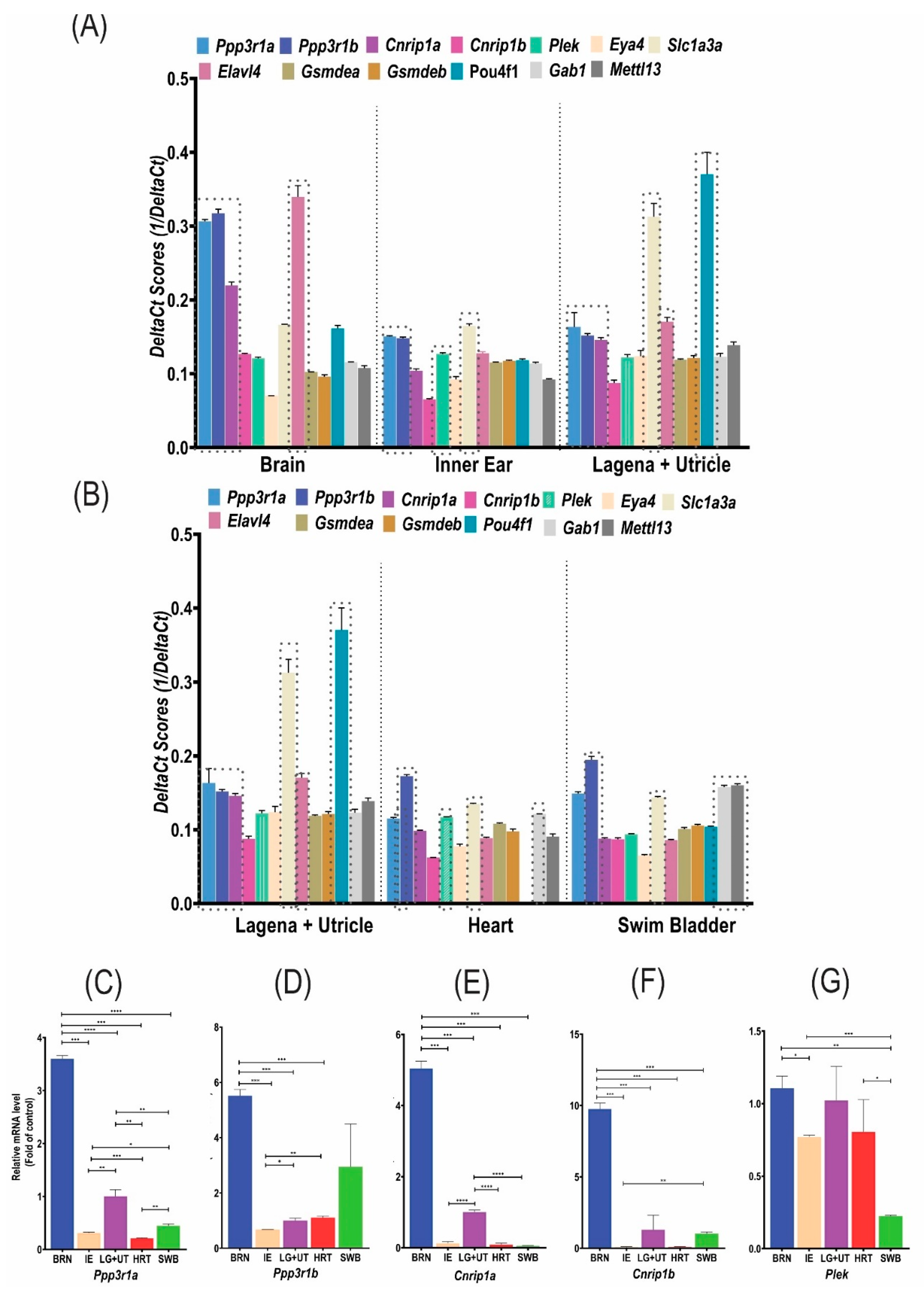
3.2. Comparison by RT-qPCR between Reference/Control and Candidate/Target Genes within Each Zebrafish Tissue
3.2.1. Inner Ear and Neuroepithelium (Lagena + Utricle) Expression
3.2.2. Other Tissues Expression
3.3. Comparison by RT-qPCR of Each Candidate/Target Gene between the Neuroepithelium (Lagena + Utricle) and the Different Zebrafish Tissues
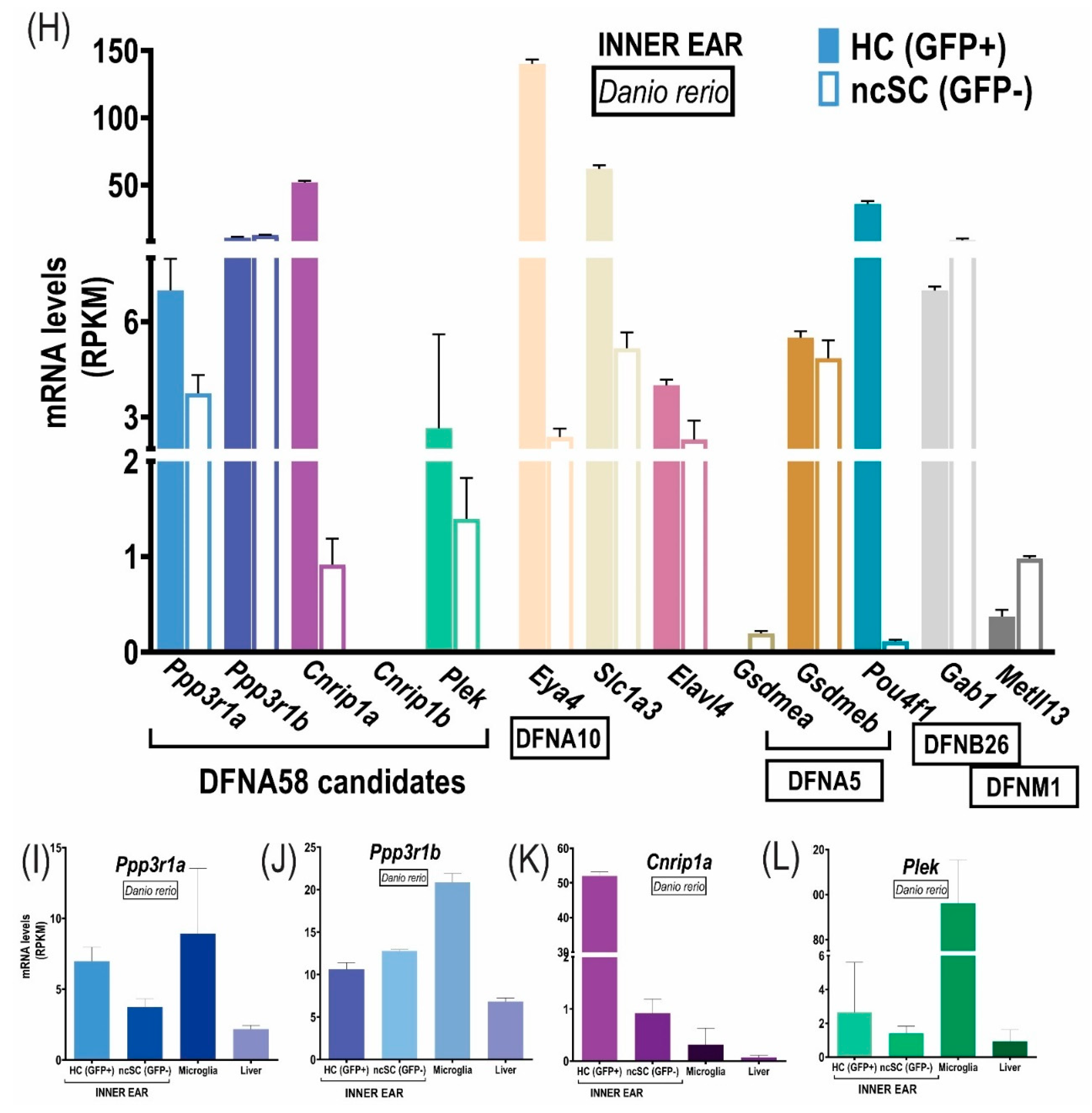
3.4. Reuse of Transcriptome Data in Zebrafish
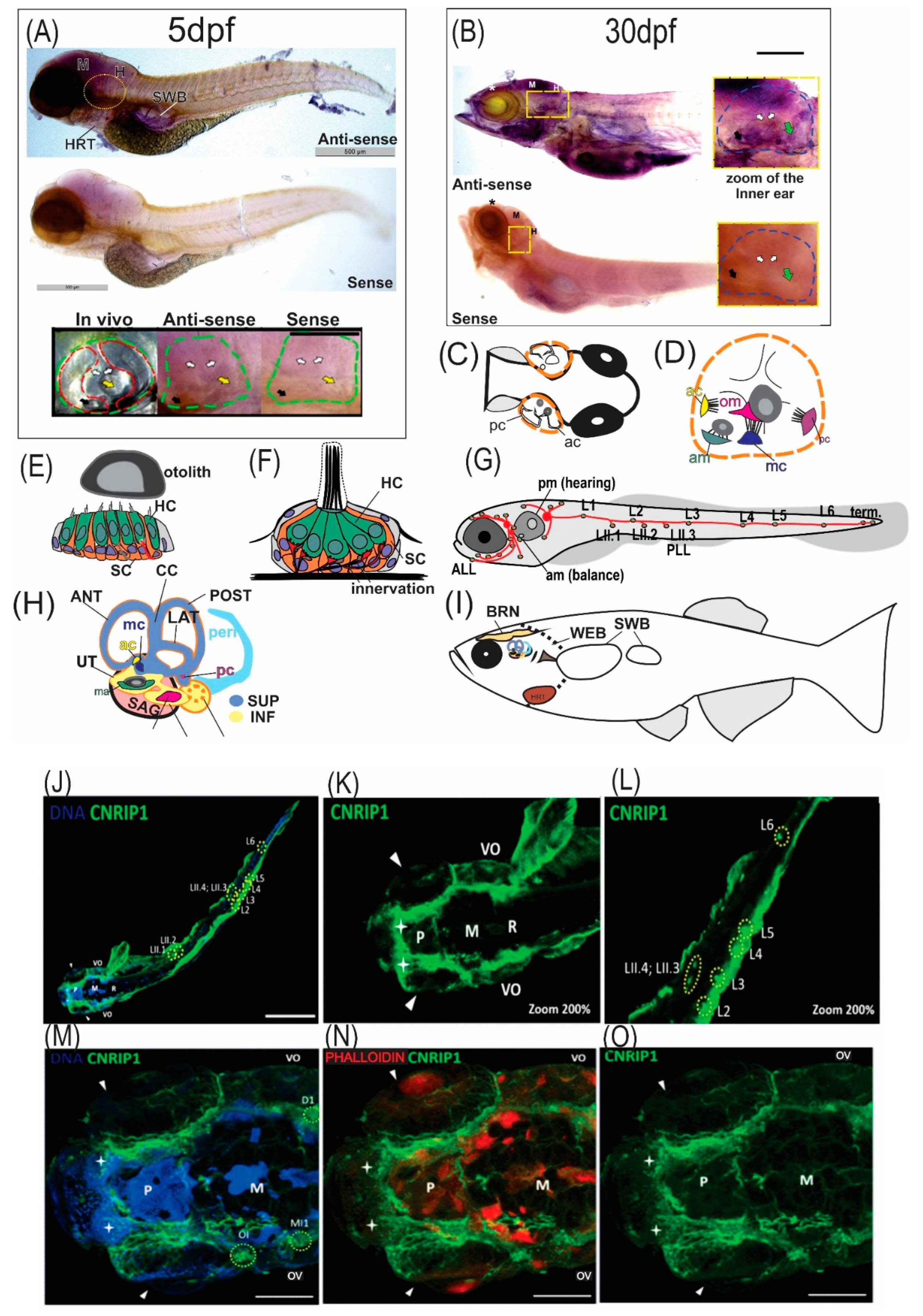
3.5. In Situ Hybridization and Immunofluorescence of Zebrafish
3.6. Expression Analyses in Mice
4. Discussion
4.1. No Evidence of Additional Genes Affected by the Duplication
4.2. CNRIP1, PPP3R1, and PLEK Function Could Potentially Be Important for Hearing
4.3. Expression Analyses in the Inner Ear
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mulrow, C.D.; Aguilar, C.; Endicott, J.E.; Velez, R.; Tuley, M.R.; Charlip, W.S.; Hill, J.A. Association Between Hearing Impairment and the Quality of Life of Elderly Individuals. J. Am. Geriatr. Soc. 1990, 38, 45–50. [Google Scholar] [CrossRef]
- Cacciatore, F.; Napoli, C.; Abete, P.; Marciano, E.; Triassi, M.; Rengo, F. Quality of life determinants and hearing function in an elderly population: Osservatorio Geriatrico Campano Study Group. Gerontology 1999, 45, 323–328. [Google Scholar] [CrossRef]
- World Health Organization. Deafness and Hearing Loss. Available online: https://www.who.int/health-topics/hearing-loss#tab=tab_1 (accessed on 28 December 2021).
- Petit, C.; Levilliers, J.; Hardelin, J.P. Molecular genetics of hearing loss. Annu. Rev. Genet. 2001, 35, 589–646. [Google Scholar] [CrossRef]
- Batissoco, A.C.; Pedroso-Campos, V.; Pardono, E.; Sampaio-Silva, J.; Sonoda, C.Y.; Vieira-Silva, G.A.; da Silva de Oliveira Longati, E.U.; Mariano, D.; Hoshino, A.C.H.; Tsuji, R.K.; et al. Molecular and genetic characterization of a large Brazilian cohort presenting hearing loss. Hum. Genet. 2022, 141, 519–538. [Google Scholar] [CrossRef]
- Shearer, E.A.; Hildebrand, M.S.; Smith, R.J. Hereditary Hearing Loss and Deafness Overview. In GeneReviews®; Adam, M.P., Everman, D.B., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Eds.; University of Washington: Seattle, WA, USA, 2017. [Google Scholar]
- Van Camp, G.; Smith, R.J.H. Hereditary Hearing Loss Homepage. Available online: https://hereditaryhearingloss.org (accessed on 3 November 2021).
- Lezirovitz, K.; Braga, M.C.; Thiele-Aguiar, R.S.; Auricchio, M.T.; Pearson, P.L.; Otto, P.A.; Mingroni-Netto, R.C. A novel autosomal dominant deafness locus (DFNA58) maps to 2p12-p21. Clin. Genet. 2009, 75, 490–493. [Google Scholar] [CrossRef]
- Nicolson, T. The genetics of hearing and balance in zebrafish. Annu. Rev. Genet. 2005, 39, 9–22. [Google Scholar] [CrossRef]
- Whitfield, T.T. Zebrafish as a model for hearing and deafness. J. Neurobiol. 2002, 53, 157–171. [Google Scholar] [CrossRef]
- He, Y.; Bao, B.; Li, H. Using zebrafish as a model to study the role of epigenetics in hearing loss. Expert Opin. Drug Discov. 2017, 12, 967–975. [Google Scholar] [CrossRef]
- Xia, W.; Hu, J.; Mam, J.; Huang, J.; Wang, X.; Jiang, N.; Zhang, J.; Ma, Z.; Ma, D. Novel TRRAP mutation causes autosomal dominant non-syndromic hearing loss. Clin. Genet. 2019, 96, 300–308. [Google Scholar] [CrossRef]
- Harris, J.A.; Cheng, A.G.; Cunningham, L.L.; MacDonald, G.; Raible, D.W.; Rubel, E.W. Neomycin-induced hair cell death and rapid regeneration in the lateral line of zebrafish (Danio rerio). JARO -J. Assoc. Res. Otolaryngol. 2003, 4, 219–234. [Google Scholar] [CrossRef]
- Ma, E.Y.; Rubel, E.W.; Raible, D.W. Notch signaling regulates the extent of hair cell regeneration in the zebrafish lateral line. J. Neurosci. 2008, 28, 2261–2273. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Liu, F.; Wang, Y.; Wang, H.; Mam, J.; Xia, W.; Zhang, J.; Jiang, N.; Sun, S.; Wang, X.; et al. Down-regulation of msrb3 and destruction of normal auditory system development through hair cell apoptosis in zebrafish. Int. J. Dev. Biol. 2015, 59, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Sielemann, K.; Hafner, A.; Pucker, B. The reuse of public datasets in the life sciences: Potential risks and rewards. PeerJ 2020, 8, e9954. [Google Scholar] [CrossRef] [PubMed]
- Curty, R.G.; Crowston, K.; Specht, A.; Grant, B.W.; Dalton, E.D. Attitudes and norms affecting scientists’ data reuse. PLoS ONE 2017, 12, e0189288. [Google Scholar] [CrossRef]
- Wade, T.D. Traits and types of health data repositories. Heal. Inf. Sci. Syst. 2014, 2, 4. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, B.; Li, R.; Lu, Z.; Huang, Y. Does comorbidity increase the risk of patients with covid-19: Evidence from meta-analysis. Aging (Albany. NY) 2020, 12, 6049–6057. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets - Update. Nucleic Acids Res. 2013, 41, 991–995. [Google Scholar] [CrossRef] [PubMed]
- Kodama, Y.; Shumway, M.; Leinonen, R. The sequence read archive: Explosive growth of sequencing data. Nucleic Acids Res. 2012, 40, 2011–2013. [Google Scholar] [CrossRef]
- Rustici, G.; Kolesnikov, N.; Brandizi, M.; Burdett, T.; Dylag, M.; Emam, I.; Farne, A.; Hastings, E.; Ison, J.; Keays, M.; et al. ArrayExpress update-trends in database growth and links to data analysis tools. Nucleic Acids Res. 2013, 41, 987–990. [Google Scholar] [CrossRef]
- Erickson, T.; Nicolson, T. Identification of sensory hair-cell transcripts by thiouracil-tagging in zebrafish. BMC Genom. 2015, 16, 842. [Google Scholar] [CrossRef]
- McDermott, B.M.; Baucom, J.M.; Hudspeth, A.J. Analysis and functional evaluation of the hair-cell transcriptome. Proc. Natl. Acad. Sci. USA 2007, 104, 11820–11825. [Google Scholar] [CrossRef] [PubMed]
- Barta, C.L.; Liu, H.; Chen, L.; Giffen, K.P.; Li, Y.; Kramer, K.L.; Beisel, K.W.; He, D.Z. RNA-seq transcriptomic analysis of adult zebrafish inner ear hair cells. Sci. Data 2018, 5, 180005. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Pecka, J.L.; Zhang, Q.; Soukup, G.A.; Beisel, K.W.; He, D.Z.Z. Characterization of transcriptomes of cochlear inner and outer hair cells. J. Neurosci. 2014, 34, 11085–11095. [Google Scholar] [CrossRef] [PubMed]
- Elkon, R.; Milon, B.; Morrison, L.; Shah, M.; Vijayakumar, S.; Racherla, M.; Leitch, C.C.; Silipino, L.; Hadi, S.; Weiss-Gayet, M.; et al. RFX transcription factors are essential for hearing in mice. Nat. Commun. 2015, 6, 8549. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Jen, H.I.; Kang, H.; Klisch, T.J.; Zoghbi, H.Y.; Groves, A.K. Characterization of the transcriptome of nascent hair cells and identification of direct targets of the atoh1 transcription factor. J. Neurosci. 2015, 35, 5870–5883. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.C.; Kelly, M.C.; Hoa, M.; Morell, R.J.; Kelley, M.W. Single-cell RNA-Seq resolves cellular complexity in sensory organs from the neonatal inner ear. Nat. Commun. 2015, 6, 8557. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Giffen, K.P.; Chen, L.; Beisel, K.W.; He, D.Z.Z. Transcriptomes of cochlear inner and outer hair cells from adult mice. Sci. Data 2018, 5, 180199. [Google Scholar] [CrossRef]
- Li, C.; Li, X.; Bi, Z.; Sugino, K.; Wang, G.; Zhu, T.; Liu, Z. Comprehensive transcriptome analysis of cochlear spiral ganglion neurons at multiple ages. Elife 2020, 9, 50491. [Google Scholar] [CrossRef]
- Sun, S.; Babola, T.; Pregernig, G.; So, K.S.; Nguyen, M.; Su, S.M.; Palermo, A.T.; Bergles, D.E.; Burns, J.C.; Müller, U. Hair Cell Mechanotransduction Regulates Spontaneous Activity and Spiral Ganglion Subtype Specification in the Auditory System. Cell 2018, 174, 1247–1263.e15. [Google Scholar] [CrossRef]
- Lezirovitz, K.; Vieira-Silva, G.A.; Batissoco, A.C.; Levy, D.; Kitajima, J.P.; Trouillet, A.; Ouyang, E.; Zebarjadi, N.; Sampaio-Silva, J.; Pedroso-Campos, V.; et al. A rare genomic duplication in 2p14 underlies autosomal dominant hearing loss DFNA58. Hum. Mol. Genet. 2020, 29, 1520–1536. [Google Scholar] [CrossRef]
- Baumgart, M.; Priebe, S.; Groth, M.; Hartmann, N.; Menzel, U.; Pandolfini, L.; Koch, P.; Felder, M.; Ristow, M.; Englert, C.; et al. Longitudinal RNA-seq analysis of vertebrate aging identifies mitochondrial complex i as a small-molecule-sensitive modifier of lifespan. Cell Syst. 2016, 2, 122–132. [Google Scholar] [CrossRef]
- Oosterhof, N.; Holtman, I.R.; Kuil, L.E.; van der Linde, H.C.; Boddeke, E.W.; Eggen, B.J.; van Ham, T.J. Identification of a conserved and acute neurodegeneration-specific microglial transcriptome in the zebrafish. Glia 2017, 65, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Hertzano, R.; Elkon, R.; Kurima, K.; Morrisson, A.; Chan, S.L.; Sallin, M.; Biedlingmaier, A.; Darling, D.S.; Griffith, A.J.; Eisenman, D.J.; et al. Cell type-specific transcriptome analysis reveals a major role for Zeb1 and miR-200b in mouse inner ear morphogenesis. PLoS Genet. 2011, 7, e1002309. [Google Scholar] [CrossRef] [PubMed]
- Einarsson, R.; Haden, M.; Diciolli, G.; Lim, A.; Mah-Ginn, K.; Aguilar, K.; Yazejian, L.; Yazejian, B. Patch clamp recordings in inner ear hair cells isolated from zebrafish. J. Vis. Exp. 2012, 68, 4281. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Hosoya, M.; Okano, H.; Fujioka, M.; Ogawa, K. Expression pattern of EYA4 in the common marmoset (Callithrix jacchus) cochlea. Neurosci Lett. 2018, 662, 185–188. [Google Scholar] [CrossRef]
- Busch-Nentwich, E.; Söllner, C.; Roehl, H.; Nicolson, T. The deafness gene dfna5 is crucial for ugdh expression and HA production in the developing ear in zebrafish. Development 2004, 131, 943–951. [Google Scholar] [CrossRef]
- Schwarzer, S.; Asokan, N.; Bludau, O.; Kuscha, V.; Kaslin, J.; Hans, S. Neurogenesis in the inner ear: The zebrafish statoacoustic ganglion provides new neurons from a Neurod/Nestin-positive progenitor pool well into adulthood. Development 2020, 147, dev176750. [Google Scholar] [CrossRef]
- Thisse, B.; Heyer, V.; Lux, A.; Alunni, V.; Degrave, A.; Seiliez, I.; Kirchner, J.; Parkhill, J.P.; Thisse, C. Spatial and temporal expression of the zebrafish genome by large-scale in situ hybridization screening. Methods Cell Biol. 2004, 77, 505–519. [Google Scholar] [CrossRef]
- Giffen, K.P.; Liu, H.; Kramer, K.L.; He, D.Z. Expression of Protein-Coding Gene Orthologs in Zebrafish and Mouse Inner Ear Non-sensory Supporting Cells. Front. Neurosci. 2019, 13, 1117. [Google Scholar] [CrossRef]
- Maskell, L.J.; Qamar, K.; Babakr, A.A.; Hawkins, T.A.; Heads, R.J.; Budhram-mahadeo, V.S. Essential but partially redundant roles for POU4F1/Brn-3a and POU4F2/Brn-3b transcription factors in the developing heart. Nat. Publ. Gr. 2017, 8, e2861-12. [Google Scholar] [CrossRef]
- Yousaf, R.; Ahmed, Z.M.; Giese, A.P.; Morell, R.J.; Lagziel, A.; Dabdoub, A.; Wilcox, E.R.; Riazuddin, S.; Friedman, T.B.; Riazuddin, S. Modifier variant of METTL13 suppresses human GAB1-associated profound deafness. J. Clin. Investig. 2018, 128, 1509–1522. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, E45. [Google Scholar] [CrossRef] [PubMed]
- Rassier, G.T.; Silveira, T.L.R.; Remião, M.H.; Daneluz, L.O.; Martins, A.W.S.; Dellagostin, E.N.; Ortiz, H.G.; Domingues, W.B.; Komninou, E.R.; Kütter, M.T.; et al. Evaluation of qPCR reference genes in GH-overexpressing transgenic zebrafish (Danio rerio). Sci. Rep. 2020, 10, 12692. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, C.; Zeng, Q.; Agrawal, I.; Zhu, X.; Gong, Z. Genome-wide identification of suitable zebrafish Danio rerio reference genes for normalization of gene expression data by RT-qPCR. J. Fish. Biol. 2016, 88, 2095–2110. [Google Scholar] [CrossRef] [PubMed]
- Krohs, C.; Bordeynik-Cohen, M.; Messika-Gold, N.; Elkon, R.; Avraham, K.B.; Nothwang, H.G. Expression pattern of cochlear microRNAs in the mammalian auditory hindbrain. Cell Tissue Res. 2021, 383, 655–666. [Google Scholar] [CrossRef]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2008, 3, 59–69. [Google Scholar] [CrossRef]
- Hu, S.S.; Arnold, A.; Hutchens, J.M.; Radicke, J.; Cravatt, B.F.; Wager-Miller, J.; Mackie, K.; Straiker, A. Architecture of cannabinoid signaling in mouse retina. J. Comp. Neurol. 2010, 518, 3848–3866. [Google Scholar] [CrossRef]
- Smith, T.H.; Blume, L.C.; Straiker, A.; Cox, J.O.; David, B.G.; McVoy, J.R.; Sayers, K.W.; Poklis, J.L.; Abdullah, R.A.; Egertová, M.; et al. Cannabinoid receptor-interacting protein 1a modulates CB1 receptor signaling and regulation. Mol. Pharmacol. 2015, 87, 747–765. [Google Scholar] [CrossRef]
- Deng, M.; Yang, H.; Xie, X.; Liang, G.; Gan, L. Comparative expression analysis of POU4F1, POU4F2 and ISL1 in developing mouse cochleovestibular ganglion neurons. Gene Expr. Patterns 2014, 15, 31–37. [Google Scholar] [CrossRef]
- Maier, E.C.; Saxena, A.; Alsina, B.; Bronner, M.E.; Whitfield, T.T. Sensational placodes: Neurogenesis in the otic and olfactory systems. Dev. Biol. 2014, 389, 50–67. [Google Scholar] [CrossRef]
- Bang, P.I.; Yelick, P.C.; Malicki, J.J.; Sewell, W.F. High-throughput behavioral screening method for detecting auditory response defects in zebrafish. J. Neurosci. Methods 2002, 118, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Kindt, K.S.; Sheets, L.; Wingert, R.A. Transmission Disrupted: Modeling Auditory Synaptopathy in Zebrafish. Front. Cell Dev. Biol. 2018, 6, 114. [Google Scholar] [CrossRef] [PubMed]
- Ghysen, A.; Dambly-chaudie, C. Development of the zebrafish lateral line. Curr. Opin. Neurobiol. 2004, 14, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Haehnel-Taguchi, M.; Fernandes, A.M.; Böhler, M.; Schmitt, I.; Tittel, L.; Driever, W. Projections of the diencephalospinal dopaminergic system to peripheral sense organs in larval zebrafish (Danio rerio). Front. Neuroanat. 2018, 12, 20. [Google Scholar] [CrossRef]
- Rabbitts, T.H. Chromosomal translocations in cancer. Biochim. Biophys. Acta - Rev. Cancer 2008, 1786, 139–152. [Google Scholar] [CrossRef]
- Guarnerio, J.; Bezzi, M.; Jeong, J.C.; Paffenholz, S.V.; Berry, K.; Naldini, M.M.; Lo-Coco, F.; Tay, Y.; Beck, A.H.; Pandolfi, P.P. Oncogenic Role of Fusion-circRNAs Derived from Cancer-Associated Chromosomal Translocations. Cell 2016, 165, 289–302. [Google Scholar] [CrossRef]
- Barrett, S.P.; Wang, P.L.; Salzman, J. Circular RNA biogenesis can proceed through an exon-containing lariat precursor. Elife 2015, 4, e07540. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef]
- Liang, D.; Wilusz, J.E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev. 2014, 28, 2233–2247. [Google Scholar] [CrossRef]
- Zhang, X.O.; Bin Wang, H.; Zhang, Y.; Lu, X.; Chen, L.L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef]
- Fin, L.; Bergamin, G.; Steiner, R.A.; Hughes, S.M. The Cannabinoid Receptor Interacting Proteins 1 of zebrafish are not required for morphological development, viability or fertility. Sci. Rep. 2017, 7, 4858. [Google Scholar] [CrossRef]
- Elphick, M.R. The evolution and comparative neurobiology of endocannabinoid signalling. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3201–3215. [Google Scholar] [CrossRef]
- Guggenhuber, S.; Romo-Parra, H.; Bindila, L.; Leschik, J.; Lomazzo, E.; Remmers, F.; Zimmermann, T.; Lerner, R.; Klugmann, M.; Pape, H.C.; et al. Impaired 2-AG signaling in hippocampal glutamatergic neurons: Aggravation of anxiety-like behavior and unaltered seizure susceptibility. Int. J. Neuropsychopharmacol. 2016, 19, pyv091. [Google Scholar] [CrossRef] [PubMed]
- Niehaus, J.L.; Liu, Y.; Wallis, K.T.; Egertová, M.; Bhartur, S.G.; Mukhopadhyay, S.; Shi, S.; He, H.; Selley, D.E.; Howlett, A.C.; et al. CB1 cannabinoid receptor activity is modulated by the cannabinoid receptor interacting protein CRIP 1a. Mol. Pharmacol. 2007, 72, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Kreitzer, A.C.; Regehr, W.G. Cerebellar depolarization-induced suppression of inhibition is mediated by endogenous cannabinoids. J. Neurosci. 2001, 21, RC174. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.I.; Kunos, G.; Nicoll, R.A.; Francisco, S. Presynaptic Specificity of Endocannabinoid Signaling in the Hippocampus. Neuron 2001, 31, 453–462. [Google Scholar] [CrossRef]
- Sjöström, P.J.; Turrigiano, G.G.; Nelson, S.B. Neocortical LTD via Coincident Activation of Presynaptic NMDA and Cannabinoid Receptors. Neuron 2003, 39, 641–654. [Google Scholar] [CrossRef]
- Galvan, A.; Frullanti, E.; Anderlini, M.; Manenti, G.; Noci, S.; Dugo, M.; Ambrogi, F.; De Cecco, L.; Spinelli, R.; Piazza, R.; et al. Gene expression signature of non-involved lung tissue associated with survival in lung adenocarcinoma patients. Carcinogenesis 2013, 34, 2767–2773. [Google Scholar] [CrossRef]
- Jayaraman, T.; Marks, A.R. Calcineurin Is Downstream of the Inositol 1, 4, 5-Trisphosphate Receptor in the Apoptotic and Cell Growth Pathways *. J. Biol. Chem. 2000, 275, 6417–6420. [Google Scholar] [CrossRef]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of cell death: The calcium-apoptosis link. Nat. Rev. Mol. Cell Biol. 2003, 4, 552–565. [Google Scholar] [CrossRef]
- Minami, S.B.; Yamashita, D.; Schacht, J.; Miller, J.M. Calcineurin activation contributes to noise-induced hearing loss. J. Neurosci. Res. 2004, 78, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Sano, K.; Takai, Y.; Yamanishi, J.; Nishizuka, Y. A Role of Calcium-activated Phospholipid-dependent Protein Kinase in Human Platelet Activation. J. Biol. Chem. 2013, 258, 2010–2013. [Google Scholar] [CrossRef]
- Tyers, M.; Rachubinski, R.A.; Stewart, M.I.; Varrichio, A.M.; Shorr, R.G.; Haslam, R.J.; Harley, C.B. Molecular cloning and expression of the major protein kinase C substrate of platelets. Nature 1988, 333, 470–473. [Google Scholar] [CrossRef]
- Winstel, R.; Freund, S.; Krasel, C.; Hoppe, E.; Lohset, M.J. Protein kinase cross-talk: Membrane targeting of the receptor kinase by protein kinase C. Proc. Natl. Acad. Sci. USA 1996, 93, 2105–2109. [Google Scholar] [CrossRef]
- García-Sáinz, J.A.; Vázquez-Prado, J.; del Carmen Medina, L. Alpha 1-adrenoceptors: Function and phosphorylation. Eur J. Pharmacol. 2000, 389, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cmarik, J.L.; Hegamyer, G.; Gerrard, B.; Dean, M.; Colburn, N.H. cDNA cloning and mapping of mouse pleckstrin (Plek), a gene upregulated in transformation-resistant cells. Genomics 2000, 66, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Kitajiri, S.; Fukumoto, K.; Hata, M.; Sasaki, H.; Katsuno, T.; Nakagawa, T.; Ito, J.; Tsukita, S.; Tsukita, S. Radixin deficiency causes deafness associated with progressive degeneration of cochlear stereocilia. J. Cell Biol. 2004, 166, 559–570. [Google Scholar] [CrossRef]
- Hilgert, N.; Smith, R.J.H.; Van Camp, G. Function and expression pattern of nonsyndromic deafness genes. Curr. Mol. Med. 2009, 9, 546–564. [Google Scholar] [CrossRef]
- Salazar-Silva, R.; Dantas, V.L.G.; Alves, L.U.; Batissoco, A.C.; Oiticica, J.; Lawrence, E.A.; Kawafi, A.; Yang, Y.; Nicastro, F.S.; Novaes, B.C.; et al. NCOA3 identified as a new candidate to explain autosomal dominant progressive hearing loss. Hum. Mol. Genet. 2021, 29, 3691–3705. [Google Scholar] [CrossRef]
- Oltrabella, F.; Melgoza, A.; Nguyen, B.; Guo, S. Role of the endocannabinoid system in vertebrates: Emphasis on the zebrafish model. Dev. Growth Differ. 2017, 59, 194–210. [Google Scholar] [CrossRef]
- BGEE Database. Available online: https://bgee.org/ (accessed on 28 December 2021).
- Blasiole, B.; Canfield, V.A.; Vollrath, M.A.; Huss, D.; Mohideen, M.-A.P.; Dickman, J.D.; Cheng, K.; Fekete, D.M.; Levenson, R. Separate Na,K-ATPase genes are required for otolith formation and semicircular canal development in zebrafish. Dev. Biol. 2006, 294, 148–160. [Google Scholar] [CrossRef]
- Breuer, M.; Guglielmi, L.; Zielonka, M.; Hemberger, V.; Kölker, S.; Okun, J.G.; Hoffmann, G.F.; Carl, M.; Sauer, S.W.; Opladen, T. QDPR homologues in Danio rerio regulate melanin synthesis, early gliogenesis, and glutamine homeostasis. PLoS ONE 2019, 14, e0215162. [Google Scholar] [CrossRef]
- Cheng, W.; Guo, L.; Zhang, Z.; Soo, H.M.; Wen, C.; Wu, W.; Peng, J. HNF factors form a network to regulate liver-enriched genes in zebrafish. Dev. Biol. 2006, 294, 482–496. [Google Scholar] [CrossRef]
- Gesemann, M.; Lesslauer, A.; Maurer, C.M.; Schönthaler, H.B.; Neuhauss, S.C.F. Phylogenetic analysis of the vertebrate Excitatory/Neutral Amino Acid Transporter (SLC1/EAAT) family reveals lineage specific subfamilies. BMC Evol. Biol. 2010, 10, 117. [Google Scholar] [CrossRef]
- Hammond, D.R.; Udvadia, A.J. Cabin1 expression suggests roles in neuronal development. Dev. Dyn. 2010, 239, 2443–2451. [Google Scholar] [CrossRef] [PubMed]
- Le, T.H.; Duy, P.Q.; An, M.; Talbot, J.; Iyer, C.C.; Wolman, M.; Beattie, C.E. HuD and the Survival Motor Neuron Protein Interact in Motoneurons and Are Essential for Motoneuron Development, Function, and mRNA Regulation. J. Neurosci. 2017, 37, 11559–11571. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, M.; Fujioka, M.; Ogawa, K.; Okano, H. Distinct Expression Patterns Of Causative Genes Responsible For Hereditary Progressive Hearing Loss In Non-Human Primate Cochlea. Sci. Rep. 2016, 6, 22250. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.-I.; Thisse, C.; Thisse, B. Identification and mechanism of regulation of the zebrafish dorsal determinant. Proc. Natl. Acad. Sci. USA 2011, 108, 15876–15880. [Google Scholar] [CrossRef]
- Rico, E.P.; de Oliveira, D.L.; Rosemberg, D.B.; Mussulini, B.H.; Bonan, C.D.; Dias, R.D.; Wofchuk, S.; Souza, D.O.; Bogo, M.R. Expression and functional analysis of Na+-dependent glutamate transporters from zebrafish brain. Brain Res. Bull. 2010, 81, 517–523. [Google Scholar] [CrossRef]
- Sato, T.; Hamaoka, T.; Aizawa, H.; Hosoya, T.; Okamoto, H. Genetic Single-Cell Mosaic Analysis Implicates ephrinB2 Reverse Signaling in Projections from the Posterior Tectum to the Hindbrain in Zebrafish. J. Neurosci. 2007, 27, 5271–5279. [Google Scholar] [CrossRef]
- Schönberger, J.; Wang, L.; Shin, J.T.; Kim, S.D.; Depreux, F.F.S.; Zhu, H.; Zon, L.; Pizard, A.; Kim, J.B.; A MacRae, C.; et al. Mutation in the transcriptional coactivator EYA4 causes dilated cardiomyopathy and sensorineural hearing loss. Nat. Genet. 2005, 37, 418–422. [Google Scholar] [CrossRef]
- Wang, L.; Sewell, W.F.; Kim, S.D.; Shin, J.T.; MacRae, C.A.; Zon, L.I.; Seidman, J.G.; Seidman, C.E. Eya4 regulation of Na+/K+-ATPase is required for sensory system development in zebrafish. Development 2008, 135, 3425–3434. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nascimento, L.R.d.; Vieira-Silva, G.A.; Kitajima, J.P.F.W.; Batissoco, A.C.; Lezirovitz, K. New Insights into the Identity of the DFNA58 Gene. Genes 2022, 13, 2274. https://doi.org/10.3390/genes13122274
Nascimento LRd, Vieira-Silva GA, Kitajima JPFW, Batissoco AC, Lezirovitz K. New Insights into the Identity of the DFNA58 Gene. Genes. 2022; 13(12):2274. https://doi.org/10.3390/genes13122274
Chicago/Turabian StyleNascimento, Larissa Reis do, Gleiciele Alice Vieira-Silva, João Paulo Fumio Whitaker Kitajima, Ana Carla Batissoco, and Karina Lezirovitz. 2022. "New Insights into the Identity of the DFNA58 Gene" Genes 13, no. 12: 2274. https://doi.org/10.3390/genes13122274
APA StyleNascimento, L. R. d., Vieira-Silva, G. A., Kitajima, J. P. F. W., Batissoco, A. C., & Lezirovitz, K. (2022). New Insights into the Identity of the DFNA58 Gene. Genes, 13(12), 2274. https://doi.org/10.3390/genes13122274