
Horizontal Transfer and Evolutionary Profiles of Two Tc1/DD34E Transposons (ZB and SB) in Vertebrates

 , , ,
, , ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Transposon Searching
2.2. Phylogenetic Analysis
2.3. Sequence Analysis
2.4. HT Analysis
3. Results
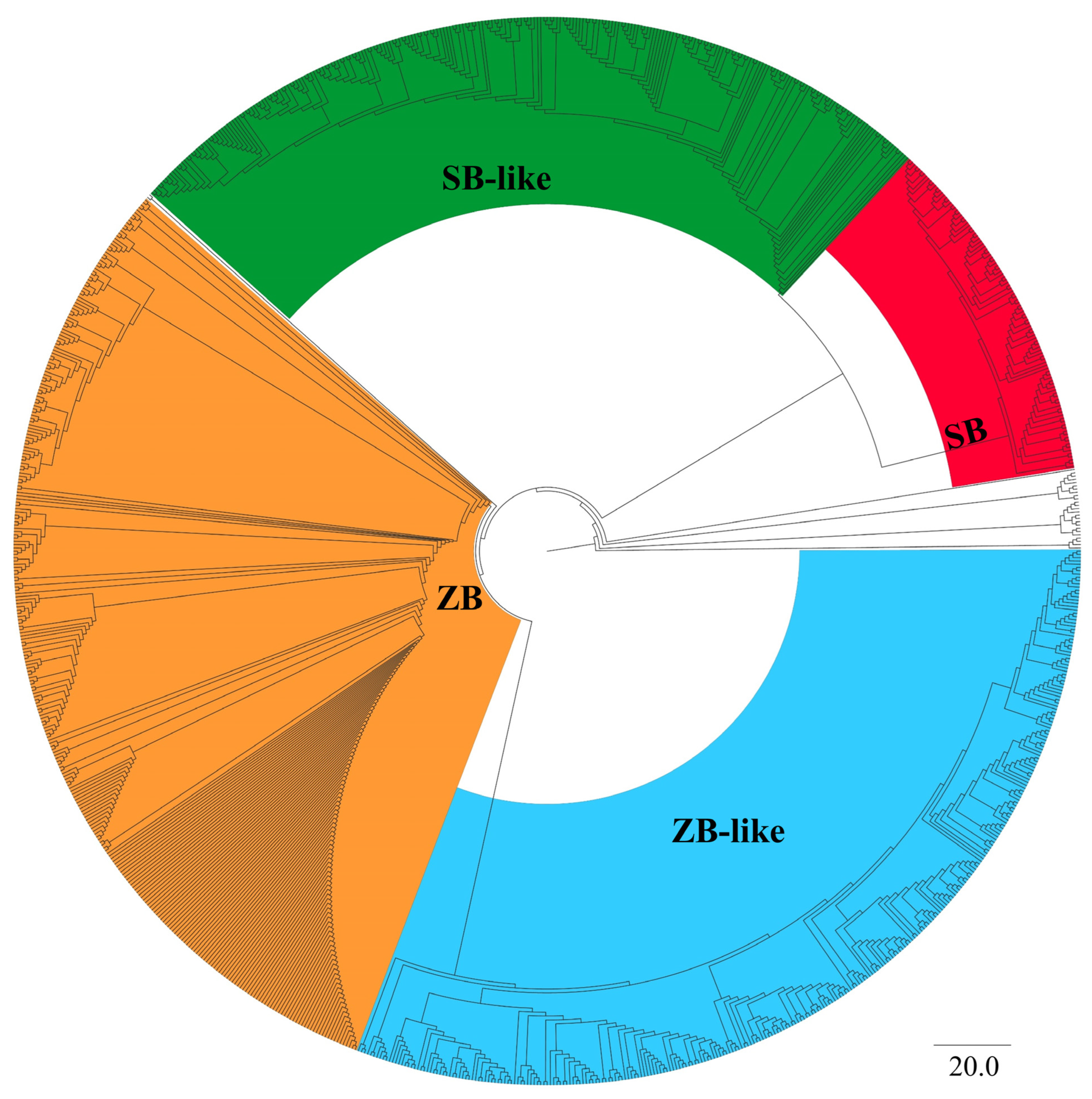
3.1. Phylogeny and Sequence Analysis of ZB and SB Transposons
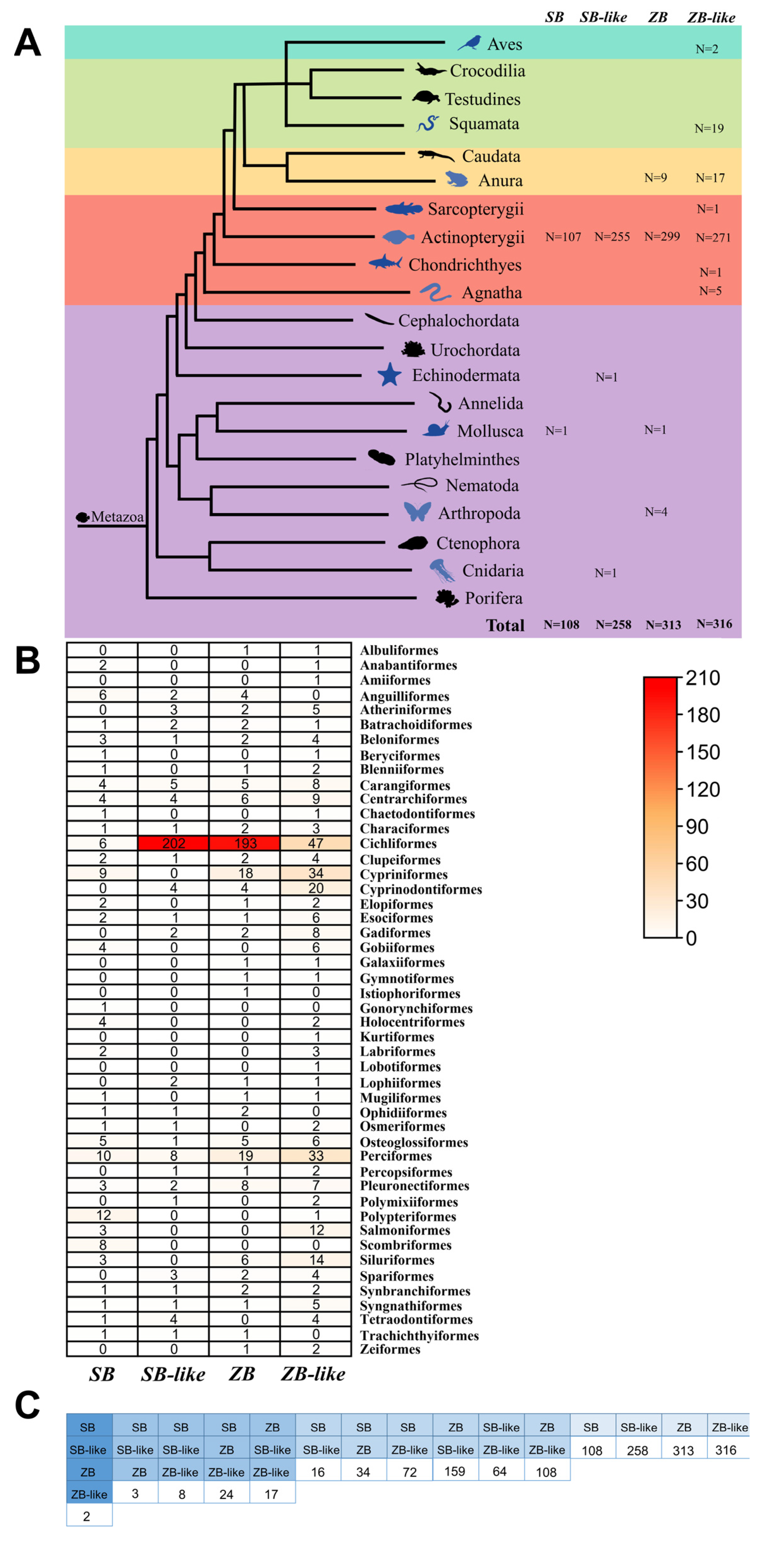
3.2. Taxonomic Distribution and Phylogenetic Analysis of ZB and SB Transposons
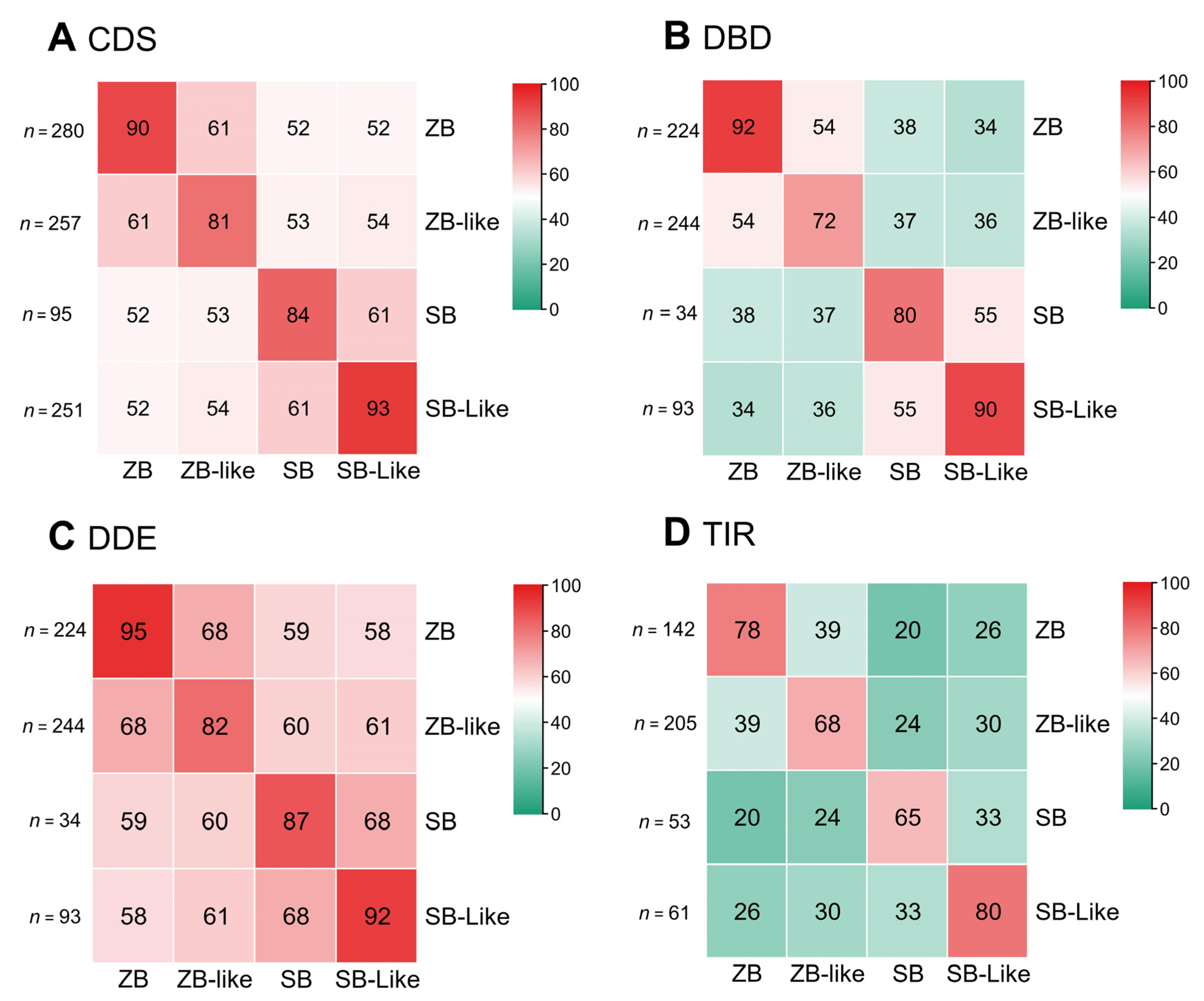
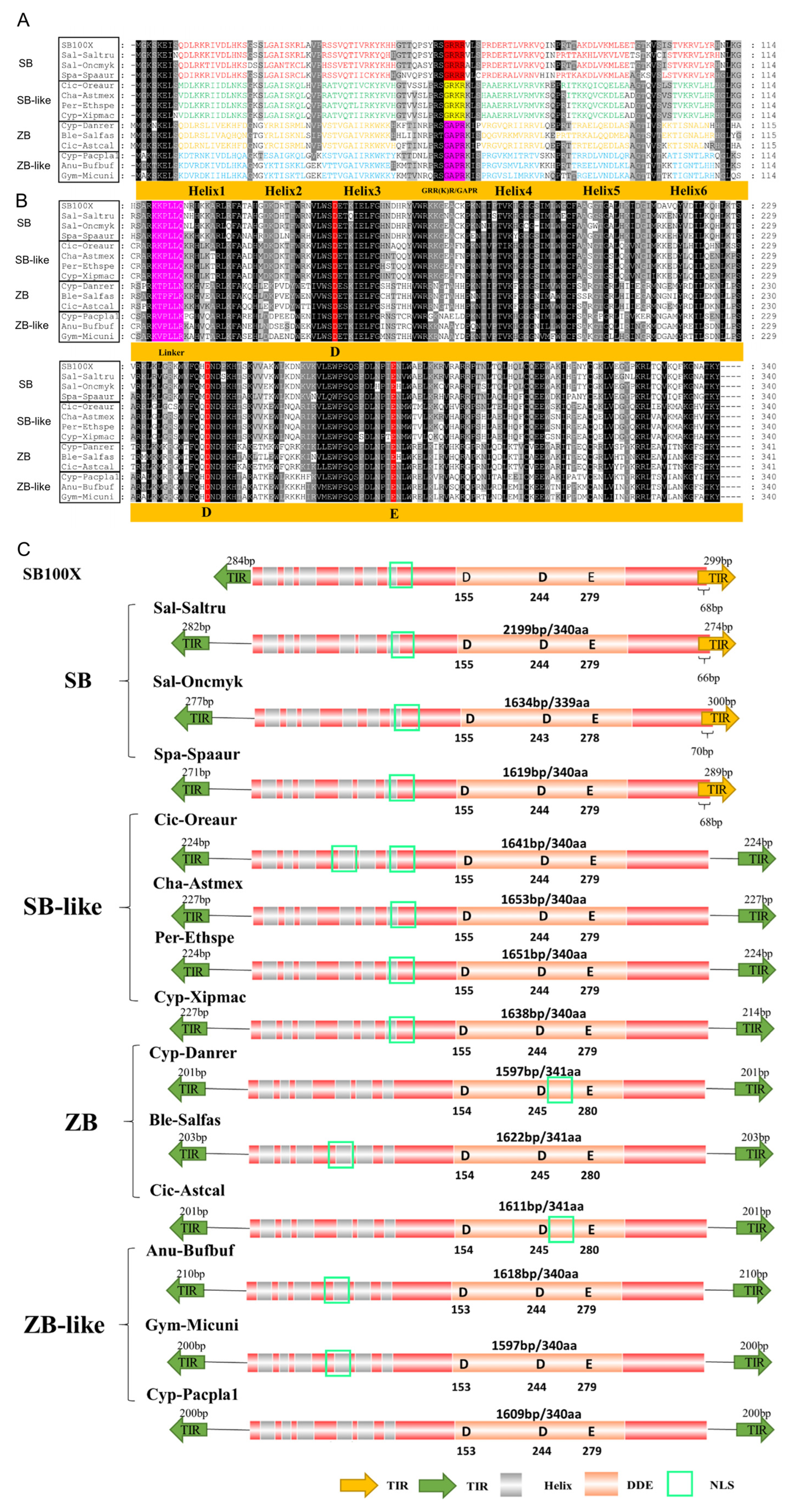
3.3. Structural Organization of ZB and SB
3.4. Evolution Dynamics of ZB and SB Transposons
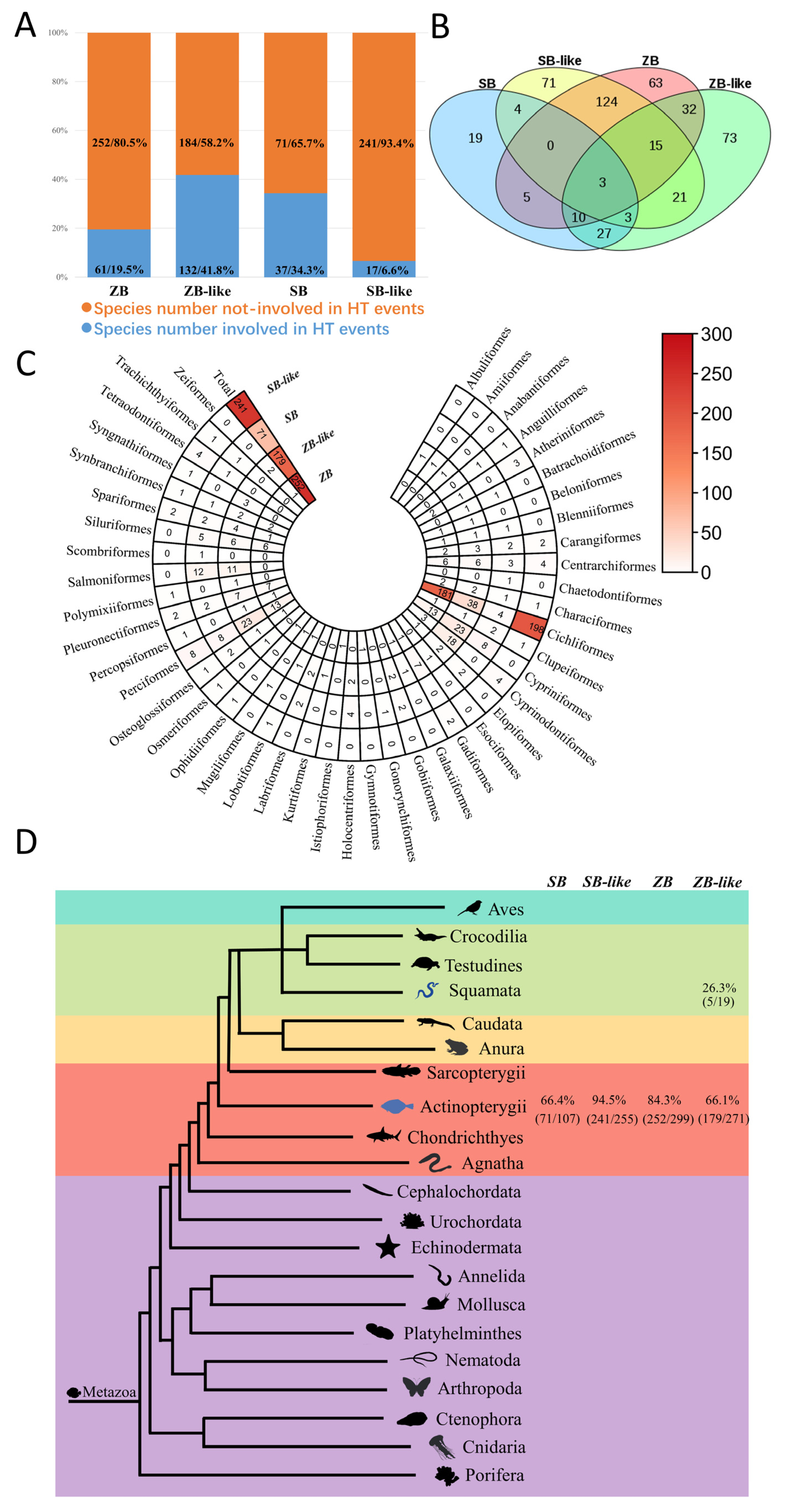
3.5. Most ZB and SB Transposons Obtained by Horizontal Transfer
4. Discussion
4.1. Recent Origins of ZB and SB Transposons
4.2. Horizontal Transfer of ZB and SB Transposons
4.3. Structure Organization of ZB and SB Transposons
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feschotte, C.; Pritham, E.J. DNA Transposons and the Evolution of Eukaryotic Genomes. Annu. Rev. Genet. 2007, 41, 331–368. [Google Scholar] [CrossRef] [PubMed]
- Sotero-Caio, C.G.; Platt, R.N.; Suh, A.; Ray, D.A. Evolution and Diversity of Transposable Elements in Vertebrate Genomes. Genome Biol. Evol. 2017, 9, 161–177. [Google Scholar] [CrossRef]
- Shao, H.; Tu, Z. Expanding the Diversity of the IS630-Tc1-Mariner Superfamily: Discovery of a Unique DD37E Transposon and Reclassification of the DD37D and DD39D Transposons. Genetics 2001, 159, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Hickman, A.B.; Chandler, M.; Dyda, F. Integrating Prokaryotes and Eukaryotes: DNA Transposases in Light of Structure. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 50–69. [Google Scholar] [CrossRef]
- Plasterk, R.H.; Izsvák, Z.; Ivics, Z. Resident Aliens: The Tc1/ Mariner Superfamily of Transposable Elements. Trends Genet. 1999, 15, 326–332. [Google Scholar] [CrossRef]
- Makałowski, W.; Gotea, V.; Pande, A.; Makałowska, I. Transposable Elements: Classification, Identification, and Their Use as a Tool for Comparative Genomics. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2019; pp. 177–207. [Google Scholar]
- Sandoval-Villegas, N.; Nurieva, W.; Amberger, M.; Ivics, Z. Contemporary Transposon Tools: A Review and Guide through Mechanisms and Applications of Sleeping Beauty, PiggyBac and Tol2 for Genome Engineering. Int. J. Mol. Sci. 2021, 22, 5084. [Google Scholar] [CrossRef]
- Gao, B.; Chen, W.; Shen, D.; Wang, S.; Chen, C.; Zhang, L.; Wang, W.; Wang, X.; Song, C. Characterization of Autonomous Families of Tc1/Mariner Transposons in Neoteleost Genomes. Mar. Genom. 2017, 34, 67–77. [Google Scholar] [CrossRef]
- Ivics, Z.; Hackett, P.B.; Plasterk, R.H.; Izsvák, Z. Molecular Reconstruction of Sleeping Beauty, a Tc1-like Transposon from Fish, and Its Transposition in Human Cells. Cell 1997, 91, 501–510. [Google Scholar] [CrossRef]
- Shen, D.; Song, C.; Miskey, C.; Chan, S.; Guan, Z.; Sang, Y.; Wang, Y.; Chen, C.; Wang, X.; Müller, F.; et al. A Native, Highly Active Tc1/Mariner Transposon from Zebrafish (ZB) Offers an Efficient Genetic Manipulation Tool for Vertebrates. Nucleic Acids Res. 2021, 49, 2126–2140. [Google Scholar] [CrossRef] [PubMed]
- Zong, W.; Gao, B.; Diaby, M.; Shen, D.; Wang, S.; Wang, Y.; Sang, Y.; Chen, C.; Wang, X.; Song, C. Traveler, a New DD35E Family of Tc1/Mariner Transposons, Invaded Vertebrates Very Recently. Genome Biol. Evol. 2020, 12, 66–76. [Google Scholar] [CrossRef]
- Sang, Y.; Gao, B.; Diaby, M.; Zong, W.; Chen, C.; Shen, D.; Wang, S.; Wang, Y.; Ivics, Z.; Song, C. Incomer, a DD36E Family of Tc1/Mariner Transposons Newly Discovered in Animals. Mobile DNA 2019, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Diaby, M.; Puzakov, M.; Ullah, N.; Wang, Y.; Danley, P.; Chen, C.; Wang, X.; Gao, B.; Song, C. Divergent Evolution Profiles of DD37D and DD39D Families of Tc1/Mariner Transposons in Eukaryotes. Mol. Phylogenet. Evol. 2021, 161, 107143. [Google Scholar] [CrossRef]
- Gao, B.; Zong, W.; Miskey, C.; Ullah, N.; Diaby, M.; Chen, C.; Wang, X.; Ivics, Z.; Song, C. Intruder (DD38E), a Recently Evolved Sibling Family of DD34E/Tc1 Transposons in Animals. Mobile DNA 2020, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, Y.; Shen, D.; Zhang, L.; Chen, W.; Chan, S.; Guan, Z.; Song, C.; Gao, B. ZB Transposon and Chicken Vasa Homologue (Cvh) Promoter Interact to Increase Transfection Efficiency of Primordial Germ Cells in Vivo. Br. Poult. Sci. 2019, 60, 724–728. [Google Scholar] [CrossRef] [PubMed]
- Mátés, L.; Chuah, M.K.L.; Belay, E.; Jerchow, B.; Manoj, N.; Acosta-Sanchez, A.; Grzela, D.P.; Schmitt, A.; Becker, K.; Matrai, J.; et al. Molecular Evolution of a Novel Hyperactive Sleeping Beauty Transposase Enables Robust Stable Gene Transfer in Vertebrates. Nat. Genet. 2009, 41, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Kowarz, E.; Löscher, D.; Marschalek, R. Optimized Sleeping Beauty Transposons Rapidly Generate Stable Transgenic Cell Lines. Biotechnol. J. 2015, 10, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Tipanee, J.; VandenDriessche, T.; Chuah, M.K. Transposons: Moving Forward from Preclinical Studies to Clinical Trials. Hum. Gene Ther. 2017, 28, 1087–1104. [Google Scholar] [CrossRef]
- Magnani, C.F.; Tettamanti, S.; Alberti, G.; Pisani, I.; Biondi, A.; Serafini, M.; Gaipa, G. Transposon-Based CAR T Cells in Acute Leukemias: Where Are We Going? Cells 2020, 9, 1337. [Google Scholar] [CrossRef]
- Prommersberger, S.; Reiser, M.; Beckmann, J.; Danhof, S.; Amberger, M.; Quade-Lyssy, P.; Einsele, H.; Hudecek, M.; Bonig, H.; Ivics, Z. Caramba: A first-in-human clinical trial with slamf7 car-t cells prepared by virus-free sleeping beauty gene transfer to treat multiple myeloma. Gene Ther. 2021, 28, 560–571. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Geurts, A.M.; Yang, Y.; Clark, K.J.; Liu, G.; Cui, Z.; Dupuy, A.J.; Bell, J.B.; Largaespada, D.A.; Hackett, P.B. Gene Transfer into Genomes of Human Cells by the Sleeping Beauty Transposon System. Mol. Ther. 2003, 8, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Craig, P.A.; Goodsell, D.; Bourne, P.E. BioEditor—Simplifying Macromolecular Structure Annotation. Bioinformatics 2003, 19, 897–898. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.D.; Tomii, K.; Katoh, K. Application of the MAFFT Sequence Alignment Program to Large Data—Reexamination of the Usefulness of Chained Guide Trees. Bioinformatics 2016, 32, 3246–3251. [Google Scholar] [CrossRef]
- Warnow, T. Revisiting Evaluation of Multiple Sequence Alignment Methods. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2021; pp. 299–317. [Google Scholar]
- Park, M.; Zaharias, P.; Warnow, T. Disjoint Tree Mergers for Large-Scale Maximum Likelihood Tree Estimation. Algorithms 2021, 14, 148. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- McGuffin, L.J.; Bryson, K.; Jones, D.T. The PSIPRED Protein Structure Prediction Server. Bioinformatics 2000, 16, 404–405. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, K.B. GeneDoc: Analysis and Visualization of Genetic Variation. EMBnet News 1997, 4, 14. [Google Scholar]
- Liu, W.; Xie, Y.; Ma, J.; Luo, X.; Nie, P.; Zuo, Z.; Lahrmann, U.; Zhao, Q.; Zheng, Y.; Zhao, Y.; et al. IBS: An Illustrator for the Presentation and Visualization of Biological Sequences. Bioinformatics 2015, 31, 3359–3361. [Google Scholar] [CrossRef]
- Guan, Z.; Shi, S.; Diaby, M.; Danley, P.; Ullah, N.; Puzakov, M.; Gao, B.; Song, C. Horizontal Transfer of Buster Transposons across Multiple Phyla and Classes of Animals. Mol. Phylogenet. Evol. 2022, 173, 107506. [Google Scholar] [CrossRef] [PubMed]
- Diaby, M.; Guan, Z.; Shi, S.; Sang, Y.; Wang, S.; Wang, Y.; Zong, W.; Ullah, N.; Gao, B.; Song, C. Revisiting the Tigger Transposon Evolution Revealing Extensive Involvement in the Shaping of Mammal Genomes. Biology 2022, 11, 921. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Tellier, M.; Bouuaert, C.C.; Chalmers, R. Mariner and the ITm Superfamily of Transposons. Microbiol. Spectr. 2015, 3, MDNA3-0033-2014. [Google Scholar] [CrossRef] [PubMed]
- Voigt, F.; Wiedemann, L.; Zuliani, C.; Querques, I.; Sebe, A.; Mátés, L.; Izsvák, Z.; Ivics, Z.; Barabas, O. Sleeping Beauty Transposase Structure Allows Rational Design of Hyperactive Variants for Genetic Engineering. Nat. Commun. 2016, 7, 11126. [Google Scholar] [CrossRef] [PubMed]
- Coy, M.R.; Tu, Z. Gambol and Tc1 Are Two Distinct Families of DD34E Transposons: Analysis of the Anopheles gambiae Genome Expands the Diversity of the IS630-Tc1-Mariner Superfamily. Insect Mol. Biol. 2005, 14, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Wang, Y.; Diaby, M.; Zong, W.; Shen, D.; Wang, S.; Chen, C.; Wang, X.; Song, C. Evolution of Pogo, a Separate Superfamily of IS630-Tc1-Mariner Transposons, Revealing Recurrent Domestication Events in Vertebrates. Mobile DNA 2020, 11, 25. [Google Scholar] [CrossRef]
- Shi, S.; Puzakov, M.; Guan, Z.; Xiang, K.; Diaby, M.; Wang, Y.; Wang, S.; Song, C.; Gao, B. Prokaryotic and Eukaryotic Horizontal Transfer of Sailor (DD82E), a New Superfamily of IS630-Tc1-Mariner DNA Transposons. Biology 2021, 10, 1005. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Shen, D.; Xue, S.; Chen, C.; Cui, H.; Song, C. The Contribution of Transposable Elements to Size Variations between Four Teleost Genomes. Mobile DNA 2016, 7, 4. [Google Scholar] [CrossRef]
- Ivics, Z.; Izsvák, Z. Sleeping Beauty Transposition. Microbiol. Spectr. 2015, 3, MDNA3-0042-2014. [Google Scholar] [CrossRef]
- Shin, J.-E.; Lin, C.; Lim, H.N. Horizontal Transfer of DNA Methylation Patterns into Bacterial Chromosomes. Nucleic Acids Res. 2016, 44, 4460–4471. [Google Scholar] [CrossRef]
- Carducci, F.; Barucca, M.; Canapa, A.; Carotti, E.; Biscotti, M.A. Mobile Elements in Ray-Finned Fish Genomes. Life 2020, 10, 221. [Google Scholar] [CrossRef]
- Gao, D.; Chu, Y.; Xia, H.; Xu, C.; Heyduk, K.; Abernathy, B.; Ozias-Akins, P.; Leebens-Mack, J.H.; Jackson, S.A. Horizontal Transfer of Non-LTR Retrotransposons from Arthropods to Flowering Plants. Mol. Biol. Evol. 2018, 35, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Ivancevic, A.M.; Kortschak, R.D.; Bertozzi, T.; Adelson, D.L. Horizontal Transfer of BovB and L1 Retrotransposons in Eukaryotes. Genome Biol. 2018, 19, 85. [Google Scholar] [CrossRef] [PubMed]
- Peccoud, J.; Loiseau, V.; Cordaux, R.; Gilbert, C. Massive Horizontal Transfer of Transposable Elements in Insects. Proc. Natl. Acad. Sci. USA 2017, 114, 4721–4726. [Google Scholar] [CrossRef]
- Schaack, S.; Gilbert, C.; Feschotte, C. Promiscuous DNA: Horizontal transfer of transposable elements and why it matters for eukaryotic evolution. Trends Ecol. Evol. 2010, 25, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.; Schaack, S.; Pace, J.K.; Brindley, P.J.; Feschotte, C. A Role for Host-Parasite Interactions in the Horizontal Transfer of Transposons Across Phyla. Nature 2010, 464, 1347–1350. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.; Feschotte, C. Horizontal acquisition of transposable elements and viral sequences: Patterns and consequences. Curr. Opin. Genet. Dev. 2018, 49, 15–24. [Google Scholar] [CrossRef]
- Pace, J.K.; Gilbert, C.; Clark, M.S.; Feschotte, C. Repeated Horizontal Transfer of a DNA Transposon in Mammals and Other Tetrapods. Proc. Natl. Acad. Sci. USA 2008, 105, 17023–17028. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-H.; Peccoud, J.; Xu, M.-R.-X.; Zhang, X.-G.; Gilbert, C. Horizontal Transfer and Evolution of Transposable Elements in Vertebrates. Nat. Commun. 2020, 11, 1362. [Google Scholar] [CrossRef]
- Zhang, H.-H.; Li, G.-Y.; Xiong, X.-M.; Han, M.-J.; Zhang, X.-G.; Dai, F.-Y. TRT, a Vertebrate and Protozoan Tc1 -like Transposon: Current Activity and Horizontal Transfer. Genome Biol. Evol. 2016, 8, 2994–3005. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Gao, B.; Miskey, C.; Chen, C.; Sang, Y.; Zong, W.; Wang, S.; Wang, Y.; Wang, X.; Ivics, Z.; et al. Multiple Invasions of Visitor, a DD41D Family of Tc1/Mariner Transposons, throughout the Evolution of Vertebrates. Genome Biol. Evol. 2020, 12, 1060–1073. [Google Scholar] [CrossRef]
- Claeys Bouuaert, C.; Walker, N.; Liu, D.; Chalmers, R. Crosstalk between Transposase Subunits during Cleavage of the Mariner Transposon. Nucleic Acids Res. 2014, 42, 5799–5808. [Google Scholar] [CrossRef]
- Dornan, J.; Grey, H.; Richardson, J.M. Structural Role of the Flanking DNA in Mariner Transposon Excision. Nucleic Acids Res. 2015, 43, 2424–2432. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, D.; Ullah, N.; Diaby, M.; Gao, B.; Song, C. Characterization and Expression Pattern of ZB and PS Transposons in Zebrafish. Gene Expr. Patterns 2021, 42, 119203. [Google Scholar] [CrossRef] [PubMed]
- Vigdal, T.J.; Kaufman, C.D.; Izsvák, Z.; Voytas, D.F.; Ivics, Z. Common Physical Properties of DNA Affecting Target Site Selection of Sleeping Beauty and Other Tc1/Mariner Transposable Elements. J. Mol. Biol. 2002, 323, 441–452. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Motif | Distribution | Species Number | Length of Full Transposon (bp) a | Length of Intact Transposon (bp) b | Transposase Length (aa) (>300) | TIR Length (bp) | TSD |
|---|---|---|---|---|---|---|---|
| ZB | Anura | 9 | / | / | / | / | / |
| Actinopterygii | 299 | 1448–2940 | 1513–1687 | 302–411 | 113–205 | TA | |
| Mollusca | 1 | 1645 | / | / | / | TA | |
| Arthropoda | 4 | 1601–1622 | 1601–1622 | 324–366 | 198–209 | TA | |
| ZB-like | Aves | 2 | / | / | / | / | / |
| Squamata | 19 | 1496–2208 | 1496–2208 | 338–366 | 126–214 | TA | |
| Anura | 17 | 1596–1618 | 1596–1618 | 340–343 | 45–211 | TA | |
| Sarcopterygii | 1 | 1597 | 1597 | 340 | 200 | TA | |
| Actinopterygii | 271 | 1331–2063 | 1391–2063 | 304–404 | 53–231 | TA | |
| Chondrichthyes | 1 | 1612 | 1612 | 340 | 201–203 | TA | |
| Agnatha | 5 | 1606–2172 | 1606–2172 | 339–340 | 183–207 | TA | |
| SB | Actinopterygii | 107 | 858–2979 | 1332–2979 | 307–360 | 44–415 | TA |
| Mollusca | 1 | 1597 | 1597 | 356 | 267–275 | TA | |
| SB-like | Aves | 1 | / | / | / | / | / |
| Actinopterygii | 255 | 1571–2111 | 1621–1753 | 306–359 | 105–228 | TA | |
| Echinodermata | 1 | 1816 | 1816 | 340 | 27 | TA | |
| Cnidaria | 1 | 1663 | / | / | / | TA |
| Family | Distribution | Genome Number | Average a | Genome Number | Average b | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1–9 Full Copy | 10–99 Full Copies | ≥100 Full Copies | Total | 1–9 Intact Copy | 10–99 Intact Copies | ≥100 Intact Copies | Total | ||||
| ZB | Actinopterygii | 18 | 108 | 12 | 138 | 44 ± 6.03 | 46 | 56 | 0 | 102 | 13 ± 1.15 |
| Arthropoda | 0 | 1 | 2 | 3 | 438 ± 240.06 | 2 | 1 | 0 | 3 | 7 ± 3.21 | |
| Mollusca | 0 | 1 | 0 | 1 | / | 0 | 0 | 0 | 0 | / | |
| Total | 18 | 110 | 14 | 142 | / | 48 | 57 | 0 | 105 | - | |
| ZB-like | Actinopterygii | 18 | 103 | 54 | 175 | 190 ± 38.21 | 95 | 41 | 19 | 155 | 144 ± 41.85 |
| Agnatha | 0 | 0 | 4 | 4 | 859 ± 389.09 | 1 | 0 | 3 | 4 | 332 ± 210.98 | |
| Anura | 0 | 5 | 4 | 9 | 180 ± 89.24 | 1 | 5 | 3 | 9 | 560 ± 332.42 | |
| Chondrichthyes | 0 | 0 | 1 | 1 | / | 0 | 1 | 0 | 1 | / | |
| Sarcopterygii | 0 | 0 | 1 | 1 | / | 0 | 0 | 1 | 1 | / | |
| Squamata | 0 | 9 | 6 | 15 | 190 ± 50.32 | 9 | 4 | 0 | 13 | 6 ± 1.73 | |
| Total | 18 | 117 | 70 | 205 | / | 106 | 51 | 26 | 183 | / | |
| SB | Actinopterygii | 15 | 25 | 13 | 53 | 329 ± 98.10 | 20 | 4 | 1 | 25 | 19 ± 7.91 |
| Mollusca | 1 | 0 | 0 | 1 | / | 1 | 0 | 0 | 1 | / | |
| Total | 16 | 25 | 13 | 54 | / | 21 | 4 | 1 | 26 | / | |
| SB-like | Actinopterygii | 9 | 45 | 7 | 61 | 78 ± 24.99 | 28 | 3 | 4 | 35 | 37 ± 17.04 |
| Cnidaria | 1 | 0 | 0 | 1 | / | 0 | 0 | 0 | 0 | / | |
| Echinodermata | 0 | 1 | 0 | 1 | / | 1 | 0 | 0 | 1 | / | |
| Total | 10 | 46 | 7 | 63 | / | 29 | 3 | 4 | 36 | / | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, W.; Asare, E.; Liu, T.; Zhang, P.; Wang, Y.; Wang, S.; Shen, D.; Miskey, C.; Gao, B.; Ivics, Z.; et al. Horizontal Transfer and Evolutionary Profiles of Two Tc1/DD34E Transposons (ZB and SB) in Vertebrates. Genes 2022, 13, 2239. https://doi.org/10.3390/genes13122239
Jia W, Asare E, Liu T, Zhang P, Wang Y, Wang S, Shen D, Miskey C, Gao B, Ivics Z, et al. Horizontal Transfer and Evolutionary Profiles of Two Tc1/DD34E Transposons (ZB and SB) in Vertebrates. Genes. 2022; 13(12):2239. https://doi.org/10.3390/genes13122239
Chicago/Turabian StyleJia, Wenzhu, Emmanuel Asare, Tao Liu, Pingjing Zhang, Yali Wang, Saisai Wang, Dan Shen, Csaba Miskey, Bo Gao, Zoltán Ivics, and et al. 2022. "Horizontal Transfer and Evolutionary Profiles of Two Tc1/DD34E Transposons (ZB and SB) in Vertebrates" Genes 13, no. 12: 2239. https://doi.org/10.3390/genes13122239
APA StyleJia, W., Asare, E., Liu, T., Zhang, P., Wang, Y., Wang, S., Shen, D., Miskey, C., Gao, B., Ivics, Z., Qian, Q., & Song, C. (2022). Horizontal Transfer and Evolutionary Profiles of Two Tc1/DD34E Transposons (ZB and SB) in Vertebrates. Genes, 13(12), 2239. https://doi.org/10.3390/genes13122239