
SINE Insertion May Act as a Repressor to Affect the Expression of Pig LEPROT and Growth Traits

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
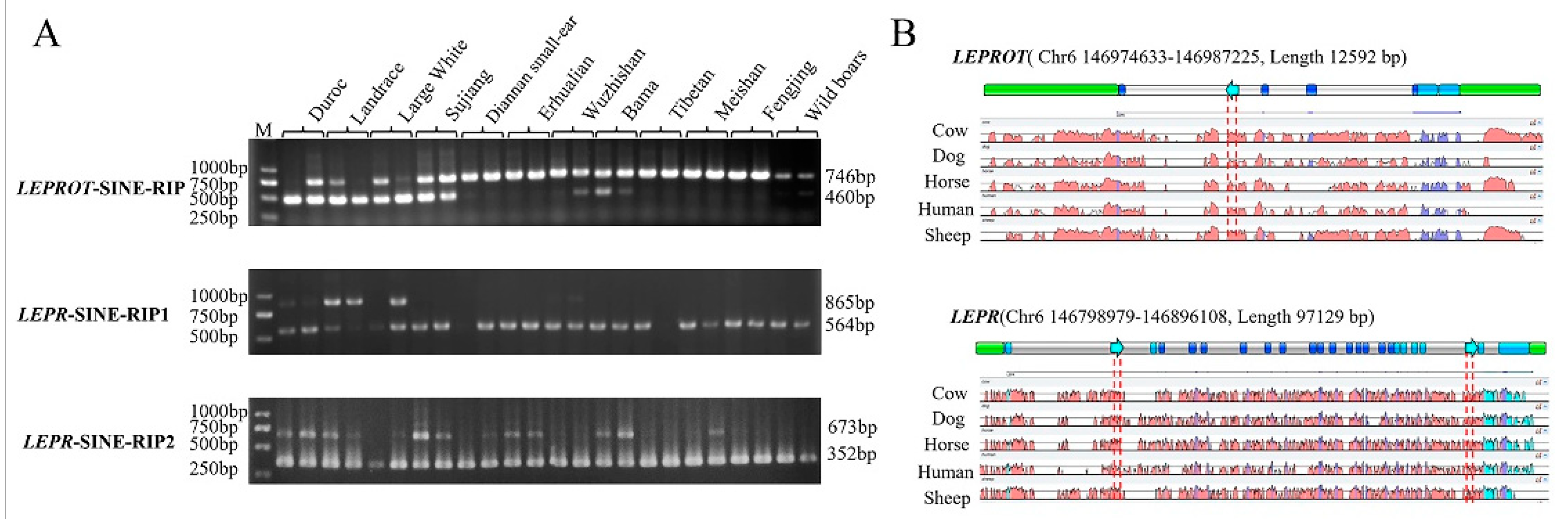
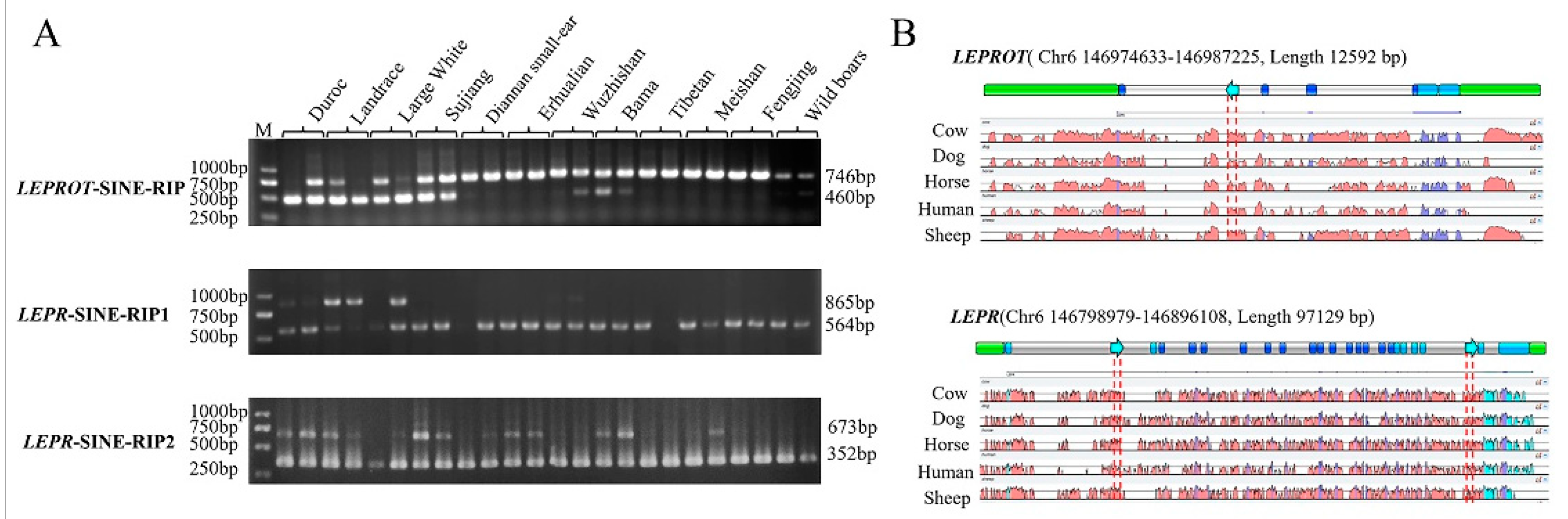
2.1. RIPs Screen and Identified
2.2. RIP Genotyping
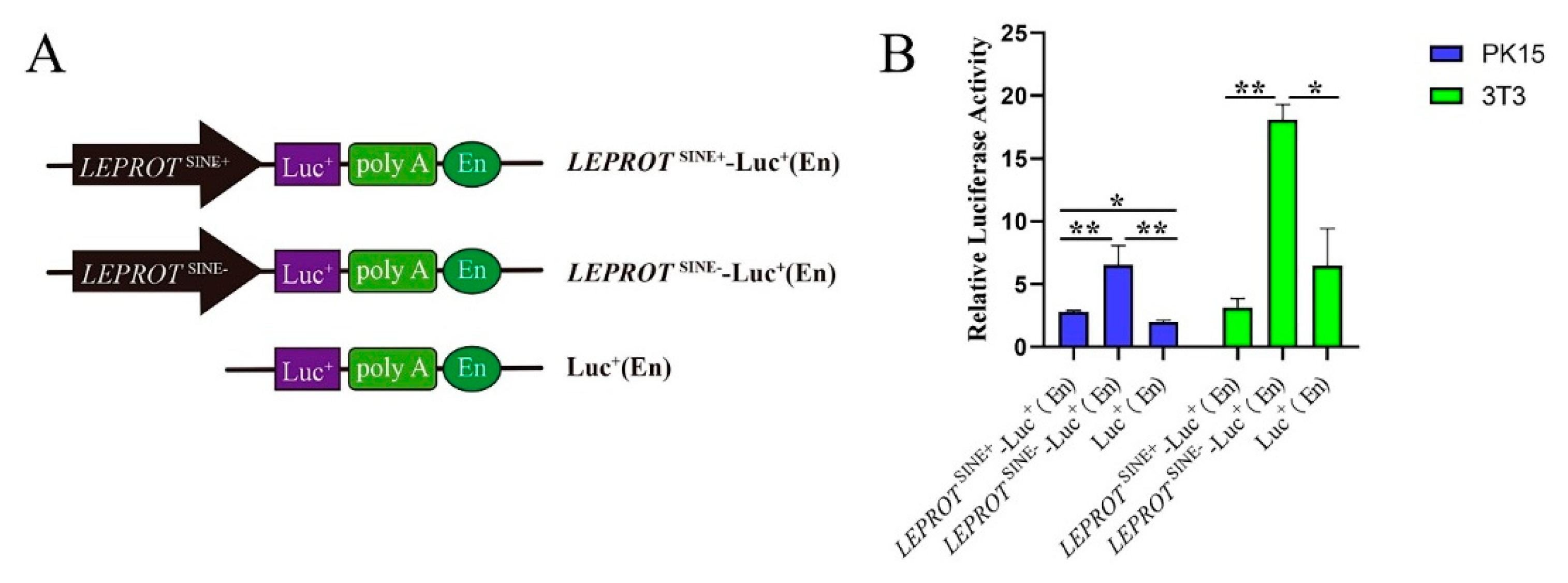
2.3. Dual-Luciferase Reporter Assay
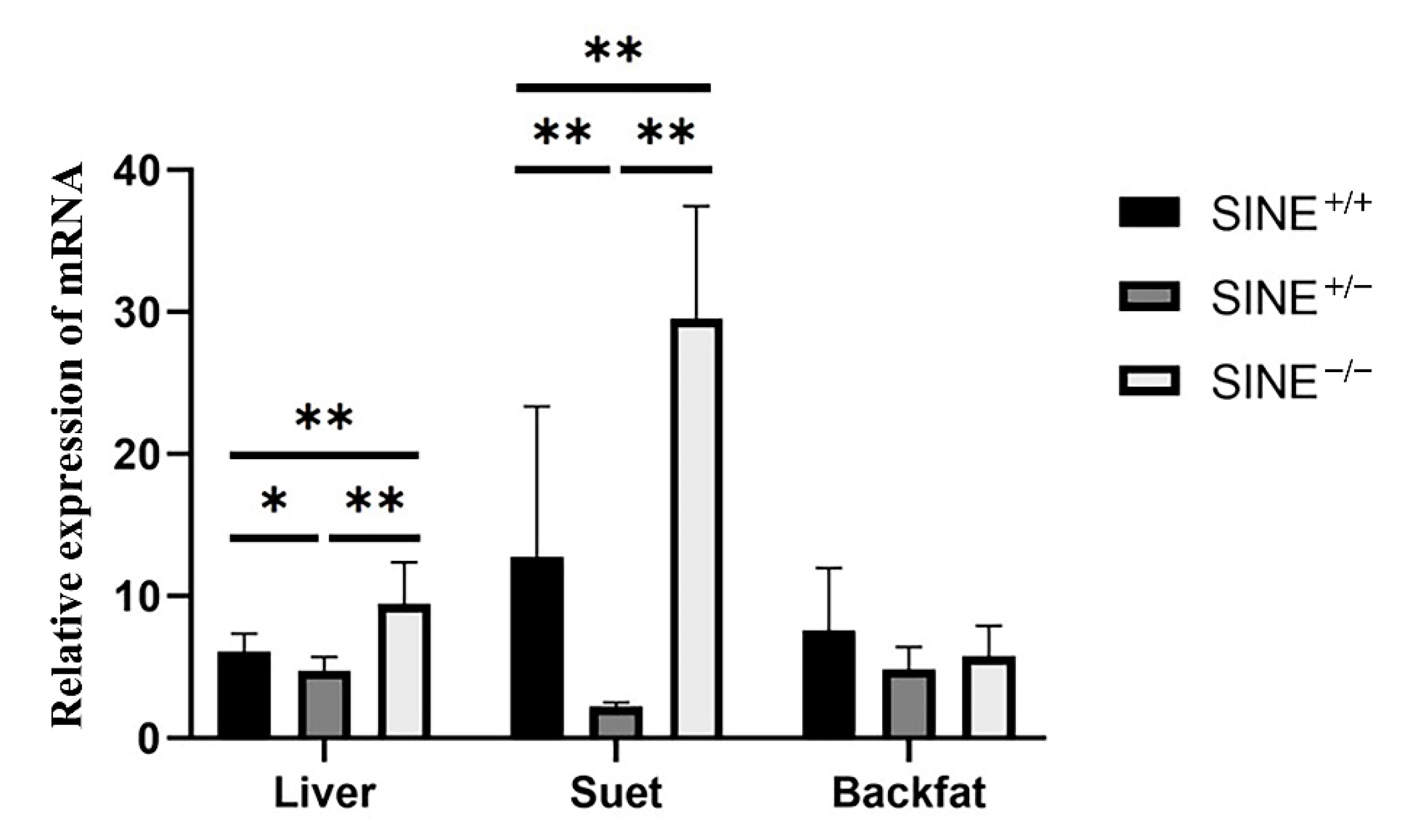
2.4. Expression Analysis
2.5. Statistical Analysis
3. Results
3.1. Three RIPs Generated by Retrotransposon Insertions in the Pig LEPR and LEPROT Gene
3.2. Three RIP Distributions in Different Pig Breeds
3.3. Correlation of LEPROT-SINE-RIP with Growth Performance of Large White
3.4. Expression Pattern of LEPROT in Tissues of 30-Day-Old Piglets
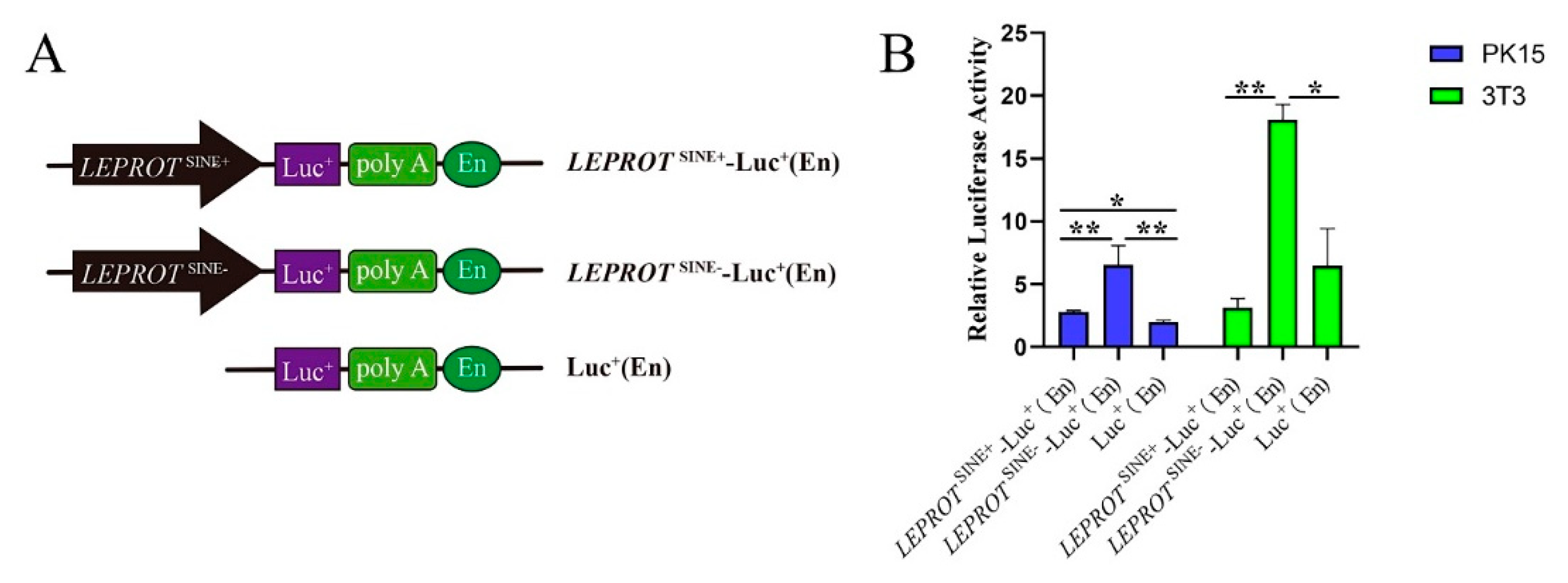
3.5. SINE Insertion May Serve as a Repressor in Regulating the Expression of LEPROT
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, F.; Chen, Y.; Heiman, M.; Dimarchi, R. Leptin: Structure, function and biology. Vitam. Horm. 2005, 71, 345–372. [Google Scholar] [PubMed]
- Obradovic, M.; Sudar-Milovanovic, E.; Soskic, S.; Essack, M.; Arya, S.; Stewart, A.J.; Gojobori, T.; Isenovic, E.R. Leptin and Obesity: Role and Clinical Implication. Front. Endocrinol. (Lausanne) 2021, 12, 585887. [Google Scholar] [CrossRef] [PubMed]
- Munzberg, H.; Singh, P.; Heymsfield, S.B.; Yu, S.; Morrison, C.D. Recent advances in understanding the role of leptin in energy homeostasis. F1000Research 2020, 9, 451. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.G.; Kusminski, C.M.; Elmquist, J.K.; Scherer, P.E. Leptin: Less Is More. Diabetes 2020, 69, 823–829. [Google Scholar] [CrossRef]
- Barb, C.R.; Hausman, G.J.; Houseknecht, K.L. Biology of leptin in the pig. Domest. Anim. Endocrinol. 2001, 21, 297–317. [Google Scholar] [CrossRef]
- Bailleul, B.; Akerblom, I.; Strosberg, A.D. The leptin receptor promoter controls expression of a second distinct protein. Nucleic Acids Res. 1997, 25, 2752–2758. [Google Scholar] [CrossRef] [Green Version]
- Seron, K.; Couturier, C.; Belouzard, S.; Bacart, J.; Monte, D.; Corset, L.; Bocquet, O.; Dam, J.; Vauthier, V.; Lecoeur, C.; et al. Endospanins Regulate a Postinternalization Step of the Leptin Receptor Endocytic Pathway. J. Biol. Chem. 2011, 286, 17968–17981. [Google Scholar] [CrossRef] [Green Version]
- Couturier, C.; Sarkis, C.; Seron, K.; Belouzard, S.; Chen, P.; Lenain, A.; Corset, L.; Dam, J.; Vauthier, V.; Dubartt, A.; et al. Silencing of OB-RGRP in mouse hypothalamic arcuate nucleus increases leptin receptor signaling and prevents diet-induced obesity. Proc. Natl. Acad. Sci. USA 2007, 104, 19476–19481. [Google Scholar] [CrossRef] [Green Version]
- Londraville, R.L.; Tuttle, M.; Liu, Q.; Andronowski, J.M. Endospanin Is a Candidate for Regulating Leptin Sensitivity. Front. Physiol. 2022, 12, 2307. [Google Scholar] [CrossRef]
- Ghalandari, H.; Hosseini-Esfahani, F.; Mirmiran, P. The Association of Polymorphisms in Leptin/Leptin Receptor Genes and Ghrelin/Ghrelin Receptor Genes with Overweight/Obesity and the Related Metabolic Disturbances: A Review. Int. J. Endocrinol. Metab. 2015, 13, e19073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasmi, A.; Mujawdiya, P.K.; Noor, S.; Piscopo, S.; Menzel, A. Lifestyle Genetics-Based Reports in the Treatment of Obesity. Arch. Razi Inst. 2021, 76, 707–719. [Google Scholar] [PubMed]
- Couto Alves, A.; De Silva, N.M.G.; Karhunen, V.; Sovio, U.; Das, S.; Taal, H.R.; Warrington, N.M.; Lewin, A.M.; Kaakinen, M.; Cousminer, D.L.; et al. Early Growth Genetics, Consortium. GWAS on longitudinal growth traits reveals different genetic factors influencing infant, child, and adult BMI. Sci. Adv. 2019, 5, eaaw3095. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Y.; Fu, Y.; Gu, P.; Zhang, Y.Y.; Tu, W.L.; Chao, Z.; Wu, H.L.; Cao, J.; Zhou, X.; Liu, B.; et al. Genome-Wide Characterization and Comparative Analyses of Simple Sequence Repeats among Four Miniature Pig Breeds. Animals 2020, 10, 1792. [Google Scholar] [CrossRef]
- Henriquez-Rodriguez, E.; Bosch, L.; Tor, M.; Pena, R.N.; Estany, J. The effect of SCD and LEPR genetic polymorphisms on fat content and composition is maintained throughout fattening in Duroc pigs. Meat Sci. 2016, 121, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadousis, C.; Somavilla, A.; Ilska, J.J.; Johnsson, M.; Batista, L.; Mellanby, R.J.; Headon, D.; Gottardo, P.; Whalen, A.; Wilson, D.; et al. A genome-wide association analysis for body weight at 35 days measured on 137,343 broiler chickens. Genet. Sel. Evol. 2021, 53, 1–14. [Google Scholar] [CrossRef]
- Wang, L.Y.; Raza, S.H.A.; Gui, L.S.; Li, S.J.; Liu, X.C.; Yang, X.R.; Wang, S.H.; Zan, L.S.; Zhao, C.P. Associations between UASMS2 polymorphism in leptin gene and growth, carcass and meat quality traits of cattle: A meta-analysis. Anim. Biotechnol. 2020, 33, 279–288. [Google Scholar] [CrossRef]
- Cai, Z.; Christensen, O.F.; Lund, M.S.; Ostersen, T.; Sahana, G. Large-scale association study on daily weight gain in pigs reveals overlap of genetic factors for growth in humans. BMC Genom. 2022, 23, 133. [Google Scholar] [CrossRef]
- Richardson, S.R.; Doucet, A.J.; Kopera, H.C.; Moldovan, J.B.; Garcia-Perez, J.L.; Moran, J.V. The Influence of LINE-1 and SINE Retrotransposons on Mammalian Genomes. Microbiol. Spectr. 2015, 3, MDNA3-0061-2014. [Google Scholar] [CrossRef] [Green Version]
- Platt, R.N.; Vandewege, M.W.; Ray, D.A. Mammalian transposable elements and their impacts on genome evolution. Chromosome Res. 2018, 26, 25–43. [Google Scholar] [CrossRef] [Green Version]
- Burns, K.H. Our Conflict with Transposable Elements and Its Implications for Human Disease. Annu. Rev. Pathol.-Mech. 2020, 15, 51–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proukakis, C. Somatic mutations in neurodegeneration: An update. Neurobiol. Dis. 2020, 144, 105021. [Google Scholar] [CrossRef] [PubMed]
- Chenais, B. Transposable Elements and Human Diseases: Mechanisms and Implication in the Response to Environmental Pollutants. Int. J. Mol. Sci. 2022, 23, 2551. [Google Scholar] [CrossRef]
- Wicker, T.; Sabot, F.; Hua-Van, A.; Bennetzen, J.L.; Capy, P.; Chalhoub, B.; Flavell, A.; Leroy, P.; Morgante, M.; Panaud, O.; et al. A unified classification system for eukaryotic transposable elements. Nat. Rev. Genet. 2007, 8, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Chuong, E.B.; Elde, N.C.; Feschotte, C. Regulatory activities of transposable elements: From conflicts to benefits. Nat. Rev. Genet. 2017, 18, 71–86. [Google Scholar] [CrossRef] [Green Version]
- Mangiavacchi, A.; Liu, P.; Della Valle, F.; Orlando, V. New insights into the functional role of retrotransposon dynamics in mammalian somatic cells. Cell Mol. Life Sci. 2021, 78, 5245–5256. [Google Scholar] [CrossRef]
- Chen, C.; D’alessandro, E.; Murani, E.; Zheng, Y.; Giosa, D.; Yang, N.S.; Wang, X.Y.; Gao, B.; Li, K.; Wimmers, K.; et al. SINE jumping contributes to large-scale polymorphisms in the pig genomes. Mob. DNA 2021, 12, 17. [Google Scholar] [CrossRef]
- Chen, C.; Wang, W.; Wang, X.; Shen, D.; Wang, S.; Wang, Y.; Gao, B.; Wimmers, K.; Mao, J.; Li, K.; et al. Retrotransposons evolution and impact on lncRNA and protein coding genes in pigs. Mob. DNA 2019, 10, 19. [Google Scholar] [CrossRef]
- Chen, C.; Wang, X.Y.; Zong, W.C.; D’alessandro, E.; Giosa, D.; Guo, Y.F.; Mao, J.D.; Song, C.Y. Genetic Diversity and Population Structures in Chinese Miniature Pigs Revealed by SINE Retrotransposon Insertion Polymorphisms, a New Type of Genetic Markers. Animals 2021, 11, 1136. [Google Scholar] [CrossRef]
- Wang, X.; D’alessandro, E.; Chi, C.; Moawad, A.S.; Zong, W.; Chen, C.; Song, C. Genetic Evaluation and Population Structure of Jiangsu Native Pigs in China Revealed by SINE Insertion Polymorphisms. Animals 2022, 12, 1345. [Google Scholar] [CrossRef]
- Wang, W.; Chen, C.; Wang, X.; Zhang, L.; Shen, D.; Wang, S.; Gao, B.; Mao, J.; Song, C. Development of Molecular Markers Based on the L1 Retrotransposon Insertion Polymorphisms in Pigs (Sus scrofa) and Their Association with Economic Traits. Russ. J. Genet. 2020, 56, 183–191. [Google Scholar] [CrossRef]
- Wang, X.Y.; Chen, Z.X.; Murani, E.; D’alessandro, E.; An, Y.L.; Chen, C.; Li, K.; Galeano, G.; Wimmers, K.; Song, C.Y. A 192 bp ERV fragment insertion in the first intron of porcine TLR6 may act as an enhancer associated with the increased expressions of TLR6 and TLR1. Mob. DNA 2021, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zheng, Y.; Wang, M.L.; Murani, E.; D’alessandro, E.; Moawad, A.S.; Wang, X.Y.; Wimmers, K.; Song, C.Y. SINE Insertion in the Intron of Pig GHR May Decrease Its Expression by Acting as a Repressor. Animals 2021, 11, 1871. [Google Scholar] [CrossRef]
- Yeh, F.C.; Yang, R.C.; Boyle, T. POPGENE version 1.32: Microsoft Windows– Based Freeware for Population Genetic Analysis, Quick User Guide. Master’s Thesis, Molecular Biology and Biotechnology Centre, University of Alberta, Edmonton, AB, Canada, 1999. [Google Scholar]
- Chan, S.; Shen, D.; Sang, Y.; Wang, S.; Wang, Y.; Chen, C.; Gao, B.; Song, C. Development of enhancer-trapping and -detection vectors mediated by the Tol2 transposon in zebrafish. PeerJ 2019, 7, e6862. [Google Scholar] [CrossRef]
- Triantafyllou, G.A.; Paschou, S.A.; Mantzoros, C.S. Leptin and Hormones: Energy Homeostasis. Endocrinol. Metab. Clin. N. Am. 2016, 45, 633–645. [Google Scholar] [CrossRef]
- Macedo, T.J.S.; Menezes, V.G.; Barberino, R.S.; Silva, R.L.S.; Gouveia, B.B.; Monte, A.P.O.; Lins, T.L.B.G.; Santos, J.M.S.; Bezerra, M.E.S.; Wischral, A.; et al. Leptin decreases apoptosis and promotes the activation of primordial follicles through the phosphatidylinositol-3-kinase/protein kinase B pathway in cultured ovine ovarian tissue. Zygote 2021, 29, 445–451. [Google Scholar] [CrossRef]
- Han, Y.C.; Ma, B.; Guo, S.; Yang, M.; Li, L.J.; Wang, S.J.; Tan, J. Leptin regulates disc cartilage endplate degeneration and ossification through activation of the MAPK-ERK signalling pathway in vivo and in vitro. J. Cell Mol. Med. 2018, 22, 2098–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Y.H.; Mckeith, F.K.; Novakofski, J.; Carr, T.R. Carcass and muscle characteristics of Yorkshire, Meishan, Yorkshire × Meishan, Meishan × Yorkshire, Fengjing × Yorkshire, and Minzhu × Yorkshire pigs. J. Anim. Sci. 1993, 71, 3344–3349. [Google Scholar] [CrossRef]
- Song, B.; Zheng, C.; Zheng, J.; Zhang, S.; Zhong, Y.; Guo, Q.; Li, F.; Long, C.; Xu, K.; Duan, Y.; et al. Comparisons of carcass traits, meat quality, and serum metabolome between Shaziling and Yorkshire pigs. Anim. Nutr. 2022, 8, 125–134. [Google Scholar] [CrossRef]
- Liu, J.; Nong, Q.; Wang, J.; Chen, W.; Xu, Z.; You, W.; Xie, J.; Wang, Y.; Shan, T. Breed difference and regulatory role of CRTC3 in porcine intramuscular adipocyte. Anim. Genet. 2020, 51, 521–530. [Google Scholar] [CrossRef]
- Lei, L.; Wang, Z.; Li, J.; Yang, H.; Yin, Y.; Tan, B.; Chen, J. Comparative Microbial Profiles of Colonic Digesta between Ningxiang Pig and Large White Pig. Animals 2021, 11, 1862. [Google Scholar] [CrossRef]
- Poklukar, K.; Candek-Potokar, M.; Batorek Lukac, N.; Tomazin, U.; Skrlep, M. Lipid Deposition and Metabolism in Local and Modern Pig Breeds: A Review. Animals 2020, 10, 424. [Google Scholar] [CrossRef] [Green Version]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvak, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten things you should know about transposable elements. Genome Biol. 2018, 19, 199. [Google Scholar] [CrossRef] [PubMed]
- Lanciano, S.; Mirouze, M. Transposable elements: All mobile, all different, some stress responsive, some adaptive? Curr. Opin. Genet. Dev. 2018, 49, 106–114. [Google Scholar] [CrossRef]
- Roujeau, C.; Jockers, R.; Dam, J. Endospanin 1 Determines the Balance of Leptin-Regulated Hypothalamic Functions. Neuroendocrinology 2019, 108, 132–141. [Google Scholar] [CrossRef]
- Ramsay, T.G.; Yan, X.; Morrison, C. The obesity gene in swine: Sequence and expression of porcine leptin. J. Anim. Sci. 1998, 76, 484–490. [Google Scholar] [CrossRef]
- Barbe, A.; Kurowska, P.; Mlyczynska, E.; Rame, C.; Staub, C.; Venturi, E.; Billon, Y.; Rak, A.; Dupont, J. Adipokines expression profiles in both plasma and peri renal adipose tissue in Large White and Meishan sows: A possible involvement in the fattening and the onset of puberty. Gen. Comp. Endocrinol. 2020, 299, 113584. [Google Scholar] [CrossRef]
- Yang, X.F.; Qiu, Y.Q.; Wang, L.; Gao, K.G.; Jiang, Z.Y. A high-fat diet increases body fat mass and up-regulates expression of genes related to adipogenesis and inflammation in a genetically lean pig. J. Zhejiang Univ. Sci. B 2018, 19, 884–894. [Google Scholar] [CrossRef]
- Nakanishi, A.; Kobayashi, N.; Suzuki-Hirano, A.; Nishihara, H.; Sasaki, T.; Hirakawa, M.; Sumiyama, K.; Shimogori, T.; Okada, N. A SINE-derived element constitutes a unique modular enhancer for mammalian diencephalic Fgf8. PLoS ONE 2012, 7, e43785. [Google Scholar] [CrossRef] [Green Version]
- Ichiyanagi, T.; Katoh, H.; Mori, Y.; Hirafuku, K.; Boyboy, B.A.; Kawase, M.; Ichiyanagi, K. B2 SINE Copies Serve as a Transposable Boundary of DNA Methylation and histone Modifications in the Mouse. Mol. Biol. Evol. 2021, 38, 2380–2395. [Google Scholar] [CrossRef]
- Sundaram, V.; Wysocka, J. Transposable elements as a potent source of diverse cis-regulatory sequences in mammalian genomes. Philos Trans. R Soc. Lond. B Biol. Sci. 2020, 375, 20190347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbez-Masson, L.; Rowe, H.M. Retrotransposons shape species-specific embryonic stem cell gene expression. Retrovirology 2015, 12, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Loci | Insertion in Exon/Intron/UTR | Length (bp) | Direction | Type | Chromosome (Sus Scrofa11.1) |
|---|---|---|---|---|---|
| LEPROT-SINE-RIP | Intron1 | 286 | - | SINEA1 | 6:146978072-146978073 |
| LEPR-SINE-RIP1 | Intron2 | 301 | + | SINEA1 | 6:146873751-146874052 |
| LEPR-SINE-RIP2 | Intron21 | 321 | + | SINEA1 | 6:146808226-146808547 |
| Polymorphic Site | Breeds | Number | Genotype Frequency | Allele Frequency | Hardy–Weinberg Equilibrium | Polymorphic Information Content | |||
|---|---|---|---|---|---|---|---|---|---|
| +/+ | +/− | −/− | + | − | |||||
| LEPROT-SINE-RIP | Large White | 450 | 14.00 | 66.00 | 20.00 | 47.00 | 53.00 | <0.01 | 0.374 |
| Duroc | 24 | 12.50 | 66.67 | 20.83 | 45.83 | 54.17 | 0.09 | 0.373 | |
| Landrace | 24 | 0 | 37.50 | 62.50 | 18.75 | 81.25 | 0.26 | 0.258 | |
| Sujiang | 24 | 29.17 | 37.50 | 33.33 | 47.92 | 52.08 | 0.22 | 0.375 | |
| Jiangquhai | 24 | 62.50 | 20.83 | 16.67 | 72.92 | 27.08 | 0.02 | 0.317 | |
| Jinhua | 24 | 75.00 | 0 | 25.00 | 75.00 | 25.00 | <0.01 | 0.305 | |
| Rongchang | 24 | 29.17 | 70.83 | 0 | 64.58 | 35.42 | 0.01 | 0.353 | |
| LEPR-SINE-RIP1 | Large White | 429 | 4.90 | 33.57 | 61.54 | 21.68 | 78.32 | 0.81 | 0.282 |
| Duroc | 24 | 0 | 45.83 | 54.17 | 22.92 | 77.08 | 0.15 | 0.291 | |
| Sujiang | 24 | 0 | 62.50 | 37.50 | 31.25 | 58.75 | 0.03 | 0.490 | |
| Jiangquhai | 18 | 0 | 16.67 | 83.33 | 8.33 | 91.67 | 0.66 | 0.141 | |
| Jinhua | 24 | 4.17 | 29.17 | 66.67 | 18.75 | 81.25 | 0.83 | 0.258 | |
| LEPR-SINE-RIP2 | Large White | 429 | 0 | 62.70 | 37.30 | 31.35 | 68.65 | <0.01 | 0.338 |
| Duroc | 24 | 0 | 75.00 | 25.00 | 37.50 | 62.50 | 0.003 | 0.359 | |
| Landrace | 24 | 0 | 95.83 | 4.17 | 47.92 | 52.08 | <0.01 | 0.375 | |
| Sujiang | 24 | 0 | 66.67 | 33.33 | 33.33 | 66.67 | 0.01 | 0.346 | |
| Bama | 24 | 0 | 87.50 | 12.50 | 43.75 | 56.25 | 0.0001 | 0.371 | |
| Jiangquhai | 24 | 0 | 33.33 | 66.67 | 16.67 | 83.33 | 0.33 | 0.239 | |
| Erhualian | 24 | 0 | 8.33 | 91.67 | 4.17 | 95.83 | 0.83 | 0.077 | |
| Jinhua | 24 | 0 | 37.50 | 62.50 | 18.75 | 81.25 | 0.26 | 0.258 | |
| Wuzhishan | 24 | 25.00 | 58.33 | 16.67 | 54.17 | 45.83 | 0.39 | 0.373 | |
| Genotype | Number | Body Weight before Slaughter | Thickness of Backfat | Age at 100 kg Body Weight | 100 kg Day-Old Backfat Thickness |
|---|---|---|---|---|---|
| SINE+/+ | 63 | 99.23 ± 1.26 A | 11.17 ± 0.37 | 164.81 ± 1.12 a | 11.20 ± 0.32 |
| SINE+/− | 297 | 105.10 ± 0.66 B | 11.53 ± 0.18 | 160.49 ± 0.59 B | 11.02 ± 0.15 |
| SINE−/− | 90 | 104.04 ± 1.20 B | 11.64 ± 0.34 | 160.59 ± 1.08 b | 11.22 ± 0.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Chi, C.; He, J.; Du, Z.; Zheng, Y.; D’Alessandro, E.; Chen, C.; Moawad, A.S.; Asare, E.; Song, C. SINE Insertion May Act as a Repressor to Affect the Expression of Pig LEPROT and Growth Traits. Genes 2022, 13, 1422. https://doi.org/10.3390/genes13081422
Wang X, Chi C, He J, Du Z, Zheng Y, D’Alessandro E, Chen C, Moawad AS, Asare E, Song C. SINE Insertion May Act as a Repressor to Affect the Expression of Pig LEPROT and Growth Traits. Genes. 2022; 13(8):1422. https://doi.org/10.3390/genes13081422
Chicago/Turabian StyleWang, Xiaoyan, Chengling Chi, Jia He, Zhanyu Du, Yao Zheng, Enrico D’Alessandro, Cai Chen, Ali Shoaib Moawad, Emmanuel Asare, and Chengyi Song. 2022. "SINE Insertion May Act as a Repressor to Affect the Expression of Pig LEPROT and Growth Traits" Genes 13, no. 8: 1422. https://doi.org/10.3390/genes13081422
APA StyleWang, X., Chi, C., He, J., Du, Z., Zheng, Y., D’Alessandro, E., Chen, C., Moawad, A. S., Asare, E., & Song, C. (2022). SINE Insertion May Act as a Repressor to Affect the Expression of Pig LEPROT and Growth Traits. Genes, 13(8), 1422. https://doi.org/10.3390/genes13081422