
Genetic Dissection of Growth and Eco-Physiological Traits Associated with Altitudinal Adaptation in Sakhalin Fir (Abies sachalinensis) Based on QTL Mapping

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Phenotyping
2.3. Characterization of Morphological Traits
2.4. Evaluation of Bud Phenology
2.5. Evaluation of Freezing Tolerance
2.6. Measurement of Chlorophyll Fluorescence
Determination of Relationships between Phenotypic Traits
2.7. Construction of Double-Digest Restriction Site-Associated DNA Sequencing (ddRAD-seq) Library
2.8. Genotyping
2.9. Linkage Map Construction
2.10. QTL Analysis
2.11. Candidate Gene Prediction
3. Results
3.1. Phenotypic Traits
3.2. Linkage Map Construction and QTL Detection
3.3. Candidate Gene Prediction
4. Discussion
4.1. Segregated Population and Linkage Maps
4.2. Growth Traits
4.3. Eco-Physiological Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef] [Green Version]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Leimu, R.; Fischer, M. A meta-analysis of local adaptation in plants. PLoS ONE 2008, 3, e4010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene flow and local adaptation in trees. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Climate warming will reduce growth and survival of Scots pine except in the far north. Ecol. Lett. 2008, 11, 588–597. [Google Scholar] [CrossRef]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life-Functional Plant Ecology of High Mountain Ecosystems; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Oleksyn, J.; Modrzýnski, J.; Tjoelker, M.; Zytkowiak, R.; Reich, P.B.; Karolewski, P. Growth and physiology of Picea abies populations from elevational transects: Common garden evidence for altitudinal ecotypes and cold adaptation. Funct. Ecol. 1998, 12, 573–590. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Ying, C.C.; Spittlehouse, D.L.; Hamilton, D.A., Jr. Genetic responses to climate in Pinus contorta: Niche breadth, climate change, and reforestation. Ecol. Monog. 1999, 69, 375–407. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Guzmán-Reyna, R.R.; Rehfeldt, G.E. Altitudinal genetic variation among Pinus oocarpa populations in Michoacán, Mexico: Implications for seed zoning, conservation, tree breeding and global warming. For. Ecol. Manag. 2006, 229, 340–350. [Google Scholar] [CrossRef]
- White, T.; Adams, W.; Neale, D. Forest Genetics; CABI Pub.: Wallingford, Oxfordshire, UK; Cambridge, MA, USA, 2007; p. 682. [Google Scholar]
- Hufford, K.M.; Mazer, S.J. Plant ecotypes: Genetic differentiation in the age of ecological restoration. Trends Ecol. Evol. 2003, 18, 147–155. [Google Scholar] [CrossRef]
- Clausen, J.; Keck, D.D.; Hiesey, W.M. Regional differentiation in plant species. Am. Nat. 1941, 75, 231–250. [Google Scholar] [CrossRef]
- Clausen, J.; Keck, D.D.; Hiesey, W.M. Heredity of geographically and ecologically isolated races. Amer. Nat. 1947, 81, 114–133. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, O.; Lascoux, M.; Merilä, J. Ecological genomics of local adaptation. Nat. Rev. Genet. 2013, 14, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Kurahashi, A.; Hamaya, T. Variation of morphological characters and growth response of Sakhalin fir (Abies sachalinensis) in different altitude. Bull. Tokyo Univ. For. 1981, 71, 101–151. [Google Scholar]
- Taneda, H.; Funayama-Noguchi, S.; Mayr, S.; Goto, S. Elevational adaptation of morphological and anatomical traits by Sakhalin fir (Abies sachalinensis). Trees 2020, 34, 507–520. [Google Scholar] [CrossRef]
- Ishizuka, W.; Goto, S. Modeling intraspecific adaptation of Abies sachalinensis to local altitude and responses to global warming, based on a 36-year reciprocal transplant experiment. Evol. Appl. 2012, 5, 229–244. [Google Scholar] [CrossRef]
- Goto, S.; Kajiya-Kanegae, H.; Ishizuka, W.; Kitamura, K.; Ueno, S.; Hisamoto, Y.; Kudoh, H.; Yasugi, M.; Nagano, A.J.; Iwata, H. Genetic mapping of local adaptation along the altitudinal gradient in Abies sachalinensis. Tree Genet. Gen. 2017, 13, 104. [Google Scholar] [CrossRef]
- Ueno, S.; Nakamura, Y.; Kobayashi, M.; Terashima, S.; Ishizuka, W.; Uchiyama, K.; Tsumura, Y.; Yano, K.; Goto, S. TodoFirGene: Developing transcriptome resources for genetic analysis of Abies sachalinensis. Plant Cell Physiol. 2018, 59, 1276–1284. [Google Scholar] [CrossRef]
- Yamazaki, T. Pinaceae. In Flora of Japan: Pteridophyta and Gymnospermae; Kodansha: Tokyo, Japan, 1995; pp. 266–277. [Google Scholar]
- Kitamura, K.; Uchiyama, K.; Ueno, S.; Ishizuka, W.; Tsuyama, I.; Goto, S. Geographical gradients of genetic diversity and differentiation among the southernmost marginal populations of Abies sachalinensis revealed by EST-SSR polymorphism. Forests 2020, 11, 233. [Google Scholar] [CrossRef] [Green Version]
- Tsuyama, I.; Ishizuka, W.; Kitamura, K.; Taneda, H.; Goto, S. Ten Years of provenance trials and application of Multivariate Random Forests predicted the most preferable seed source for silviculture of Abies sachalinensis in Hokkaido, Japan. Forests 2020, 11, 1058. [Google Scholar] [CrossRef]
- Ishizuka, W.; Kon, H.; Kita, K.M.K.; Goto, S. Local adaptation to contrasting climatic conditions in Sakhalin fir (Abies sachalinensis) revealed by long-term provenance trials. Ecol. Res. 2021. [Google Scholar] [CrossRef]
- Ishizuka, W.; Ono, K.; Hara, T.; Goto, S. Use of intraspecific variation in thermal responses for estimating an elevational cline in the timing of cold hardening in a sub-boreal conifer. Plant. Biol. 2015, 17, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, K.; Hällgren, J.-E. Cold acclimation of Pinus contorta and Pinus sylvestris assessed by chlorophyll fluorescence. Tree Physiol. 1993, 13, 97–106. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of Q A redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209. [Google Scholar] [CrossRef]
- Hendrickson, L.; Furbank, R.T.; Chow, W.S. A simple alternative approach to assessing the fate of absorbed light energy using chlorophyll fluorescence. Photosynth. Res. 2004, 82, 73. [Google Scholar] [CrossRef] [PubMed]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 201–247. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 15 July 2021).
- Kuhn, M.; Jackson, S.; Cimentada, J. Corrr: Correlations in R. R Package Version 0.4.3. 2020. Available online: https://CRAN.R-project.org/package=corrr (accessed on 15 June 2021).
- Peterson, B.K.; Weber, J.N.; Kay, E.H.; Fisher, H.S.; Hoekstra, H.E. Double digest RADseq: An inexpensive method for de novo SNP discovery and genotyping in model and non-model species. PLoS ONE 2012, 7, e37135. [Google Scholar]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef]
- Parchman, T.L.; Jahner, J.P.; Uckele, K.A.; Galland, L.M.; Eckert, A.J. RAD-seq approaches and applications for forest tree genetics. Tree Genet. Gen. 2018, 14, 1–25. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Rochette, N.C.; Rivera-Colón, A.G.; Catchen, J.M. Stacks 2: Analytical methods for paired-end sequencing improve RADseq-based population genomics. Mol. Ecol. 2019, 28, 4737–4754. [Google Scholar] [CrossRef] [PubMed]
- Van Ooijen, J. Joinmap 5, Software for the Calculation of Genetic Linkage Maps in Experimental Populations of Diploid Species; Kyazma, B.V.: Wageningen, The Neitherland.
- Piironen, J.; Vehtari, A. Sparsity information and regularization in the horseshoe and other shrinkage priors. Ele. J. Stat. 2017, 11, 5018–5051. [Google Scholar] [CrossRef]
- Goodrich, B.; Gabry, J.; Ali, I.; Brilleman, S. Rstanarm: Bayesian applied regression modeling via Stan. R Package Version 2018, 2, 1758. [Google Scholar]
- Neale, D.B.; Kremer, A. Forest tree genomics: Growing resources and applications. Nat. Rev. Genet. 2011, 12, 111–122. [Google Scholar] [CrossRef]
- Piironen, J.; Paasiniemi, M.; Vehtari, A. Projective inference in high- dimensional problems: Prediction and feature selection. Ele. J. Stat. 2020, 14, 2155–2197. [Google Scholar]
- Vehtari, A.; Gelman, A.; Gabry, J. Practical Bayesian model evaluation using leave-one-out cross-validation and WAIC. Stat. Comput. 2017, 27, 1413–1432. [Google Scholar] [CrossRef] [Green Version]
- Vehtari, A.; Gelman, A.; Gabry, J. Loo: Efficient leave-one-out cross-validation and WAIC for Bayesian models. R Package Version 2018, 2, 1003. [Google Scholar]
- Brown, G.R.; Bassoni, D.L.; Gill, G.P.; Fontana, J.R.; Wheeler, N.C.; Megraw, R.A.; Mark, F.; Davis, M.F.; Sewell, M.M.; Tuskan, G.A.; et al. Identification of quantitative trait loci influencing wood property traits in loblolly pine (Pinus taeda L.). III. QTL verification and candidate gene mapping. Genetics 2003, 164, 1537–1546. [Google Scholar] [CrossRef]
- Yang, Y.; Xuan, L.; Yu, C.; Wang, Z.; Xu, J.; Fan, W.; Guo, J.; Yin, Y. High-density genetic map construction and quantitative trait loci identification for growth traits in (Taxodium distichum var. distichum × T. mucronatum) × T. mucronatum. BMC Plant. Biol. 2018, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, T.; Xu, B.; Liu, C.; Zhao, F.; Huang, S. QTL detection for growth and form traits in three full-sib pedigrees of Pinus elliottii var. elliottii × P. caribaea var. hondurensis hybrids. Tree Genet. Gen. 2015, 11, 1–9. [Google Scholar] [CrossRef]
- Mori, H.; Ueno, S.; Ujino-Ihara, T.; Fujiwara, T.; Yamashita, K.; Kanetani, S.; Endo, R.; Matsumoto, A.; Uchiyama, K.; Matsui, Y. Mapping quantitative trait loci for growth and wood property traits in Cryptomeria japonica across multiple environments. Tree Genet. Gen. 2019, 15, 1–15. [Google Scholar] [CrossRef]
- Pelgas, B.; Bousquet, J.; Meirmans, P.G.; Ritland, K.; Isabel, N. QTL mapping in white spruce: Gene maps and genomic regions underlying adaptive traits across pedigrees, years and environments. BMC Genom. 2011, 12, 145. [Google Scholar] [CrossRef] [Green Version]
- De Miguel, M.; Cabezas, J.A.; de María, N.; Sánchez-Gómez, D.; Guevara, M.Á.; Vélez, M.D.; Sáez-Laguna, E.; Díaz, L.-M.; Mancha, J.-A.; Barbero, M.-C.; et al. Genetic control of functional traits related to photosynthesis and water use efficiency in Pinus pinaster Ait. drought response: Integration of genome annotation, allele association and QTL detection for candidate gene identification. BMC Genom. 2014, 15, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Ukrainetz, N.K.; Ritland, K.; Mansfield, S.D. Identification of quantitative trait loci for wood quality and growth across eight full-sib coastal Douglas-fir families. Tree Genet. Gen. 2008, 4, 159–170. [Google Scholar] [CrossRef]
- Lind, M.; Källman, T.; Chen, J.; Ma, X.F.; Bousquet, J.; Morgante, M.; Zaina, G.; Karlsson, B.; Elfstrand, M.; Lascoux, M.; et al. A Picea abies linkage map based on SNP markers identifies QTLs for four aspects of resistance to Heterobasidion parviporum infection. PLoS ONE 2014, 9, e101049. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Andersen, S.U.; Ljung, K.; Dolezal, K.; Miotk, A.; Schultheiss, S.J.; Lohmann, J.U. Hormonal control of the shoot stem-cell niche. Nature 2010, 465, 1089–1092. [Google Scholar] [CrossRef]
- Cortizo, M.; Alvarez, J.M.; Rodríguez, A.; Fernández, B.; Ordás, R.J. Cloning and characterization of a type-A response regulator differentially expressed during adventitious shoot formation in Pinus pinea L. J. Plant. Physiol. 2010, 167, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Drost, D.R.; Puranik, S.; Novaes, E.; Novaes, C.R.; Dervinis, C.; Gailing, O.; Kirst, M. Genetical genomics of Populus leaf shape variation. BMC Plant. Biol. 2015, 15, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalfi, M.; Troggio, M.; Piovani, P.; Leonardi, S.; Magnaschi, G.; Vendramin, G.; Menozzi, P. A RAPD, AFLP and SSR linkage map, and QTL analysis in European beech (Fagus sylvatica L.). Theor. Appl. Genet. 2004, 108, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, W.; Ono, K.; Hara, T.; Goto, S. Influence of low-and high-elevation plant genomes on the regulation of autumn cold acclimation in Abies sachalinensis. Front. Plant. Sci. 2015, 6, 890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grattapaglia, D.; Bertolucci, F.L.; Penchel, R.; Sederoff, R.R. Genetic mapping of quantitative trait loci controlling growth and wood quality traits in Eucalyptus grandis using a maternal half-sib family and RAPD markers. Genetics 1996, 144, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Robakowski, P. Susceptibility to low-temperature photoinhibition in three conifers differing in successional status. Tree Physiol. 2005, 25, 1151–1160. [Google Scholar] [CrossRef] [Green Version]
- Chapin, F.S., III. Morphological and physiological mechanisms of temperature compensation in phosphate absorption along a latitudinal gradient. Ecology 1974, 55, 1180–1198. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Pedigree | Female × Male Parent | Female × Male Parent |
|---|---|---|
| Grandparents | Clone A33 × Clone C1-1 † | Clone A39 × Clone C1-2 † |
| Parents | Clone P336 × Clone P236, Clone P236 × Clone P336 | |
| Parent size | P336 (height: 15.5 m, d.b.h.: 21.1 cm) | |
| P236 (height: 16.4 m, d.b.h.: 23.4 cm) | ||
| Crossing | May 2011 | |
| Seed collection | September 2011 | |
| Progeny | 252 | |
| Category | Trait Abberviations | Trait Explanation | Correlation with Altitude | Reference | Mean (SD) | CV |
|---|---|---|---|---|---|---|
| Growth | D16 | Stem diameter in 2016 | Negative | [16] | 35.01 (8.595) | 0.246 |
| H16 | Height in 2016 | Negative | [16] | 11.39 (2.298) | 0.202 | |
| H17 | Height in 2017 | Negative | [16] | 45.15 (10.94) | 0.242 | |
| CR17 | Crown area in 2017 | Unknown | - | 106.3 (39.43) | 0.371 | |
| Phenology | Bud_fl | Bud flush in 2016 | Unknown | - | 20.05 (2.538) | 0.127 |
| Freez_tol | Freezing tolerance in Nov 2016 | Positive | [25] | 0.387 (0.209) | 0.541 | |
| Morphology | Lw_ratio | Needle length/width ratio | Negative | [17] | 19.21 (3.277) | 0.171 |
| LMA | Leaf mass per area | Positive† | [17] (Thickness) | 0.112 (0.024) | 0.211 | |
| Bark_xy | Bark-xylem length ratio | Positive | [17] | 0.452 (0.042) | 0.093 | |
| Norm_reac | Normal/reaction wood ratio | Negative | [17] | 0.604 (0.113) | 0.188 | |
| SD | Stoma density | Unknown | - | 289.8 (54.34) | 0.188 | |
| SRN | Number of stoma row | Positive | [17] | 11.80 (1.583) | 0.134 | |
| Photosynthesis | ΦII | Effective quantum yield of PSII | Unknown | - | 0.308 (0.043) | 0.138 |
| NPQ | Non-photochemical quenching | Unknown | - | 2.483 (0.513) | 0.207 | |
| ΦNO | Non-regulated energy dissipation at PSII centers | Unknown | - | 0.202 (0.024) | 0.117 |
| Map | Linkage Group | Marker | Length (cM) | Average Distance between Markers (cM) | Gap (Max.) |
|---|---|---|---|---|---|
| P336 | 1 | 42 | 154.4 | 3.7 | 17.3 |
| 2 | 30 | 174.9 | 5.8 | 21.8 | |
| 3 | 31 | 145.6 | 4.7 | 17.6 | |
| 4 | 50 | 192.9 | 3.9 | 22.2 | |
| 5 | 28 | 128.6 | 4.6 | 15.8 | |
| 6 | 40 | 147.5 | 3.7 | 29.8 | |
| 7 | 44 | 208.1 | 4.7 | 18.0 | |
| 8 | 45 | 162.5 | 3.6 | 14.1 | |
| 9 | 51 | 151.3 | 3.0 | 10.1 | |
| 10 | 36 | 192.6 | 5.4 | 26.3 | |
| 11 | 45 | 151.8 | 3.4 | 20.9 | |
| 12 | 44 | 176.0 | 4.0 | 27.8 | |
| Total | 486 | 1986.2 | 4.1 | 29.8 | |
| P236 | 1 | 51 | 185.0 | 3.6 | 18.7 |
| 2 | 45 | 189.5 | 4.2 | 27.3 | |
| 3 | 35 | 131.7 | 3.8 | 13.7 | |
| 4 | 44 | 180.7 | 4.1 | 19.9 | |
| 5 | 31 | 126.6 | 4.1 | 17.4 | |
| 6 | 39 | 141.3 | 3.6 | 22.2 | |
| 7 | 54 | 180.1 | 3.3 | 18.5 | |
| 8 | 43 | 185.8 | 4.3 | 18.5 | |
| 9 | 43 | 155.2 | 3.6 | 19.0 | |
| 10 | 44 | 166.1 | 3.8 | 18.3 | |
| 11 | 43 | 136.4 | 3.2 | 12.5 | |
| 12 | 44 | 154.7 | 3.5 | 13.7 | |
| Total | 516 | 1932.8 | 3.7 | 27.3 |
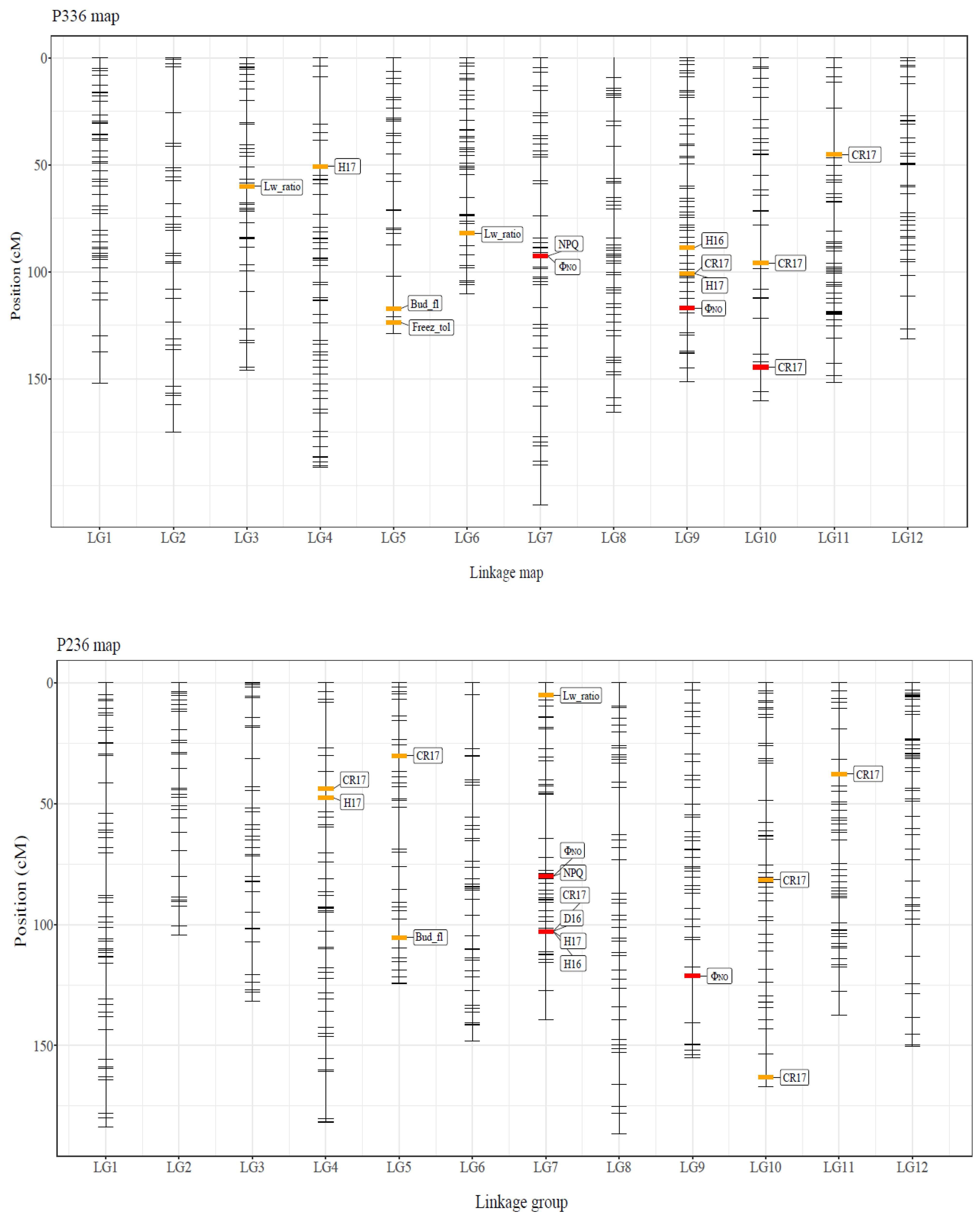
| Category | Trait | Locus | Map-LG | Pos (cM) | Sig. | PVE (%) |
|---|---|---|---|---|---|---|
| Growth | D16 | #12865 | P236-LG7 | 102.8 | * | 6.70 |
| H16 | #10164 | P336-LG9 | 88.7 | * | 4.27 | |
| H16 | #12865 | P236-LG7 | 102.8 | ** | 9.17 | |
| H17 | #10758 | P336-LG4 | 50.7 | * | 4.16 | |
| H17 | #10541 | P336-LG9 | 100.9 | * | 4.57 | |
| H17 | #10758 | P236-LG4 | 47.5 | * | 4.16 | |
| H17 | #12865 | P236-LG7 | 102.8 | ** | 7.81 | |
| CR17 | #6809 | P336-LG10 | 95.9 | * | 2.62 | |
| CR17 | #10829 | P336-LG10 | 144.6 | ** | 5.66 | |
| CR17 | #1970 | P336-LG11 | 45.2 | * | 5.45 | |
| CR17 | #10541 | P336-LG9 | 100.9 | * | 4.28 | |
| CR17 | #6809 | P236-LG10 | 81.5 | * | 2.62 | |
| Phenology | Bud_fl | #6899 | P336-LG5 | 117.2 | * | 6.07 |
| Bud_fl | #6899 | P236-LG5 | 105.4 | * | 6.07 | |
| Freez_tol | #25432 | P336-LG5 | 123.8 | * | 7.40 | |
| Morphology | Lw_ratio | #24342 | P336-LG3 | 60.0 | * | 5.96 |
| Lw_ratio | #7510 | P336-LG6 | 82.0 | * | 2.97 | |
| Lw_ratio | #30964 | P236-LG7 | 5.2 | * | 5.28 | |
| Photosynthesis | NPQ | #2055 | P336-LG7 | 92.6 | * | 6.12 |
| NPQ | #2055 | P236-LG7 | 80.1 | * | 6.12 | |
| ΦNO | #2055 | P336-LG7 | 92.6 | ** | 9.32 | |
| ΦNO | #27288 | P336-LG9 | 117.1 | ** | 7.24 | |
| ΦNO | #2055 | P236-LG7 | 80.1 | ** | 9.32 | |
| ΦNO | #27288 | P236-LG9 | 121.3 | ** | 7.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goto, S.; Mori, H.; Uchiyama, K.; Ishizuka, W.; Taneda, H.; Kono, M.; Kajiya-Kanegae, H.; Iwata, H. Genetic Dissection of Growth and Eco-Physiological Traits Associated with Altitudinal Adaptation in Sakhalin Fir (Abies sachalinensis) Based on QTL Mapping. Genes 2021, 12, 1110. https://doi.org/10.3390/genes12081110
Goto S, Mori H, Uchiyama K, Ishizuka W, Taneda H, Kono M, Kajiya-Kanegae H, Iwata H. Genetic Dissection of Growth and Eco-Physiological Traits Associated with Altitudinal Adaptation in Sakhalin Fir (Abies sachalinensis) Based on QTL Mapping. Genes. 2021; 12(8):1110. https://doi.org/10.3390/genes12081110
Chicago/Turabian StyleGoto, Susumu, Hideki Mori, Kentaro Uchiyama, Wataru Ishizuka, Haruhiko Taneda, Masaru Kono, Hiromi Kajiya-Kanegae, and Hiroyoshi Iwata. 2021. "Genetic Dissection of Growth and Eco-Physiological Traits Associated with Altitudinal Adaptation in Sakhalin Fir (Abies sachalinensis) Based on QTL Mapping" Genes 12, no. 8: 1110. https://doi.org/10.3390/genes12081110
APA StyleGoto, S., Mori, H., Uchiyama, K., Ishizuka, W., Taneda, H., Kono, M., Kajiya-Kanegae, H., & Iwata, H. (2021). Genetic Dissection of Growth and Eco-Physiological Traits Associated with Altitudinal Adaptation in Sakhalin Fir (Abies sachalinensis) Based on QTL Mapping. Genes, 12(8), 1110. https://doi.org/10.3390/genes12081110