
Flow Cytometry-Based Determination of Ploidy from Dried Leaf Specimens in Genomically Complex Collections of the Tropical Forage Grass Urochloa s. l.

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Collection and Preservation of Plant Material for Flow Cytometry
- Leaf fragments of approximately 1 g fresh weight were harvested in the field, folded into permeable manila seed storage envelopes (80 gsm) and kept in a sealed plastic bag on wet ice. Young leaves from typical vigorous specimens, representative of the population in each plot, were selected. Insect damaged and discolored plants were avoided.
- The envelopes were then stored in a sealed desiccator at ambient pressure, or an airtight plastic box (as used for sandwiches or larger sizes), at room temperature with a thick layer of self-indicating silica gel (a granular material with c. 3–5 mm irregular beads; Type III Sigma-Aldrich, S7625; or self-indicating mixed with non-indicating silica gel, cheaply available from online marketplaces). The silica gel was changed daily until it did not change color, which was after approximately 4–5 days. In total, 250 g of silica gel was used for 30 leaf samples.
- Multiple samples in the paper envelopes were then transferred to sealed plastic bags with a small amount of silica gel.
- The plastic bags with envelopes of dried leaves and silica gel were shipped under ambient conditions to the University of Leicester, UK (with appropriate export and import documentation, here under “Section IV: Cut flowers, foliage and vegetables” and “Section III: Seeds for planting” of the UK “Import requirements for plants, plant produce and products”). The sealed bags, after inspection and replacement of silica gel if required, were then stored in 4 °C in plastic boxes containing silica gel until flow cytometry analysis.
- The seeds received from VIR, USDA and CIAT were germinated in a tropical greenhouse (25 °C), and leaf samples were collected from plants, and dried and preserved in the same way as those collected in the field in Colombia, and then used as standards for flow cytometry analysis.
2.3. Sample Preparation for Flow Cytometry
- In total, 500 mg of dried leaf of each accession were chopped with a sharp razor blade in a 55 × 15 mm polystyrene Petri dish with ice cold 1 mL nucleus-isolation buffer. Much smaller amounts of leaf material (100 mg) did not give suitable nuclear suspensions. We used double edge stainless razor blades, allocating one razor edge per one studied accession. For safe holding of the razor blade while chopping, a rubber grip was used.
- Three different standard buffers were evaluated, as shown in Table 2. Buffers were supplemented with 15 mM β-mercaptoethanol (Calbiochem CAS 60-24-2) and 1% PVP-40 (polyvinylpyrrolidone-40; Sigma PVP40) and the effect of these chemicals on reducing the negative effect of cytosolic and phenolic compounds was tested.
- After finely chopping the material in the buffer, the nuclei suspension was passed through a 50 μm mesh nylon filter (CellTrics, Sysmex 04-004-2327) into the 12 × 75 mm round-bottom polystyrene flow cytometry tubes (Falcon® 352052, with caps preventing cross-contamination), and placed on ice.
- The nuclei suspension was then supplemented with propidium iodide (PI, Sigma P4170; 50 µg mL−1; solution in deionized water, passed through a 0.22 mm filter), and ribonuclease A (Sigma R6513; 50 µg mL−1) to prevent staining of double-stranded RNA, and mixed gently using vortex.
- Samples were incubated for at least 10 min on ice in darkness, and then were analyzed in an Accuri C6 Flow Cytometer (Becton Dickinson), equipped with a 20 mW laser illumination operating at 488 nm.
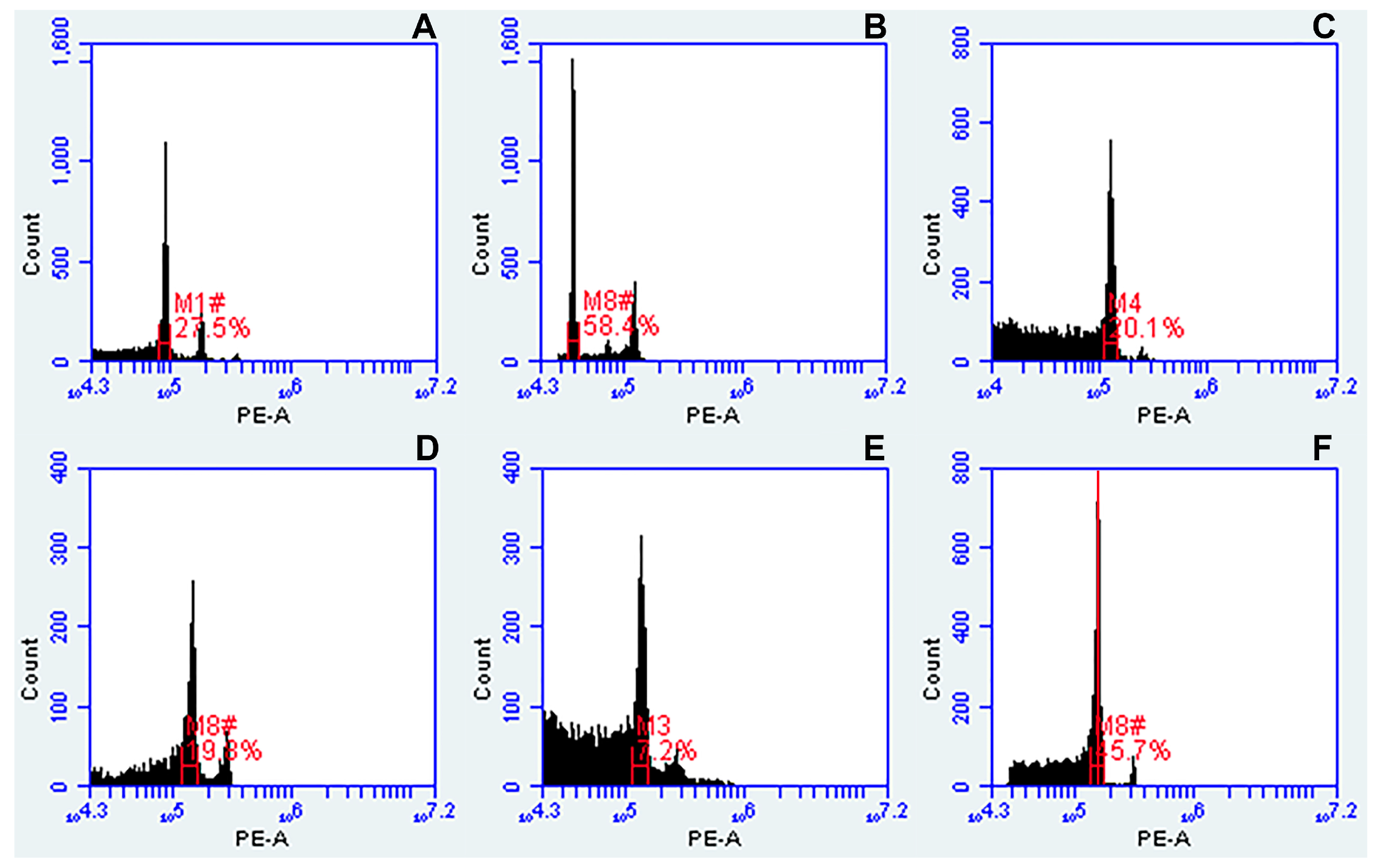
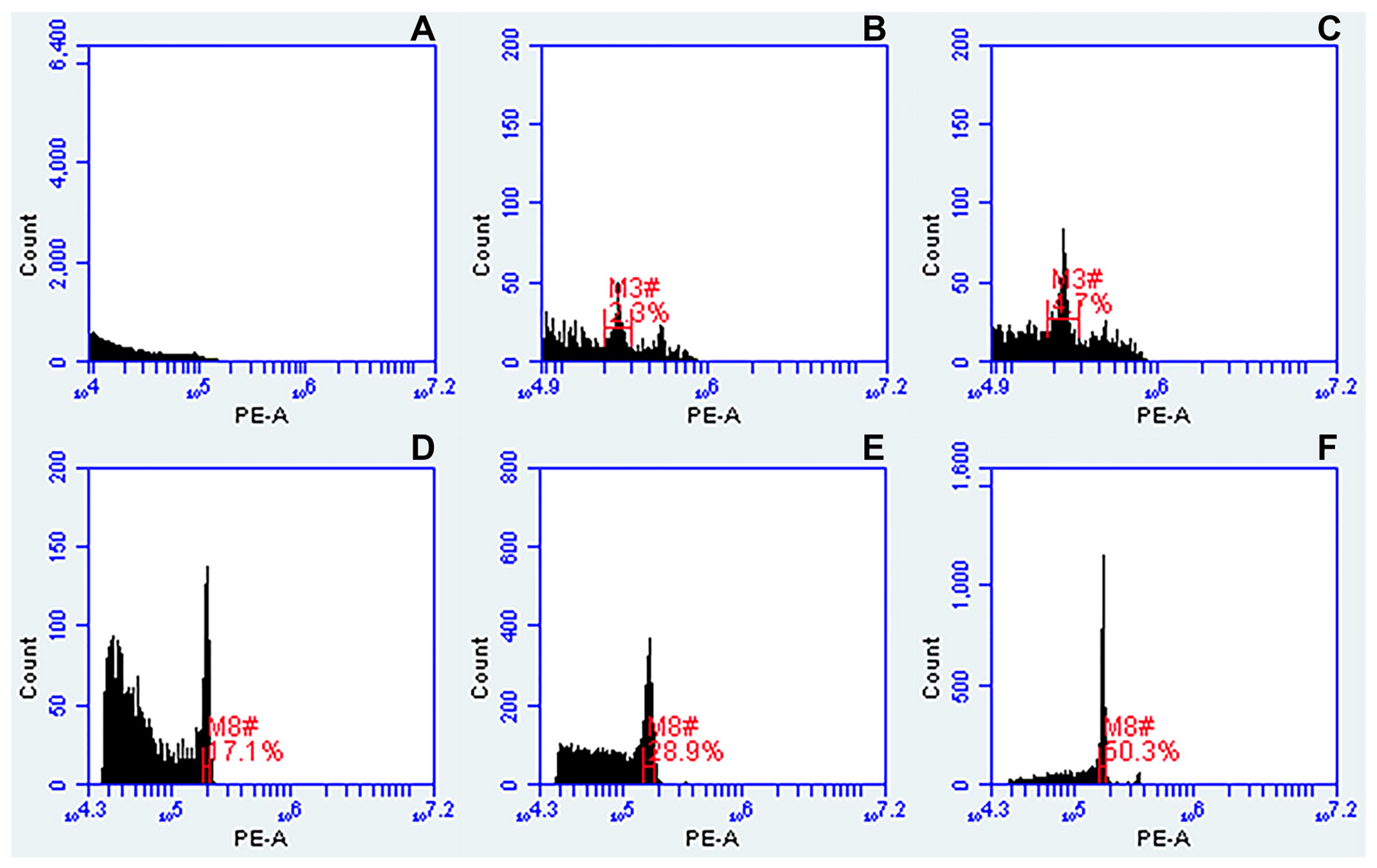
2.4. Flow Cytometry Analysis
- Prior to measuring the ploidy of samples from a given species, the flow cytometer was calibrated using the specified external standard (Table 1). For calibration, we used VirtualGain tool. In the external standard plot, the Peak Definition Marker (see red line in Figure 2F) was manually moved to the center of the external standard peak, becoming the reference point. Samples to be measured were aligned to this position.
- The histograms (FSC-A vs. SSC-A, FL1-A vs. FL2-A, FL3-A vs. FL2-A, and a univariate histogram of FL2-A) were acquired using the CFlow® Plus software set up according to Galbraith and Lambert [46]. Here, the following filter configurations were used: FL-1-a 530/14-nm bandpass filter; FL-2-a 585/20-nm bandpass filter; FL-3-a 670-nm longpass filter. The primary threshold was set to channel 10,000 on FSC-A to gate out debris and noise from nuclei suspension. The secondary threshold was set at 1000 for FL-2. Polygonal gating tool was used to draw a region on the FSC-A vs. SSC-A plot, and a line-shaped cluster of dots showing PI-stained nuclei in the biparametric dot plot of FL2-A vs. FL3-A. Based on this gating, G0/G1 and G2 peaks appeared in a univariate histogram of FL2-A.
- The relative fluorescence values of the peak positions of PI-stained nuclei (FL) and the coefficient of variation (CV) of the G0/G1 peak to estimate nuclei integrity and variation in DNA staining were evaluated in each sample by manually placing regions of identification across the peak to export values.
- Ploidy of studied plants was determined by comparing the fluorescence values of the peak positions of samples to that of external standards.
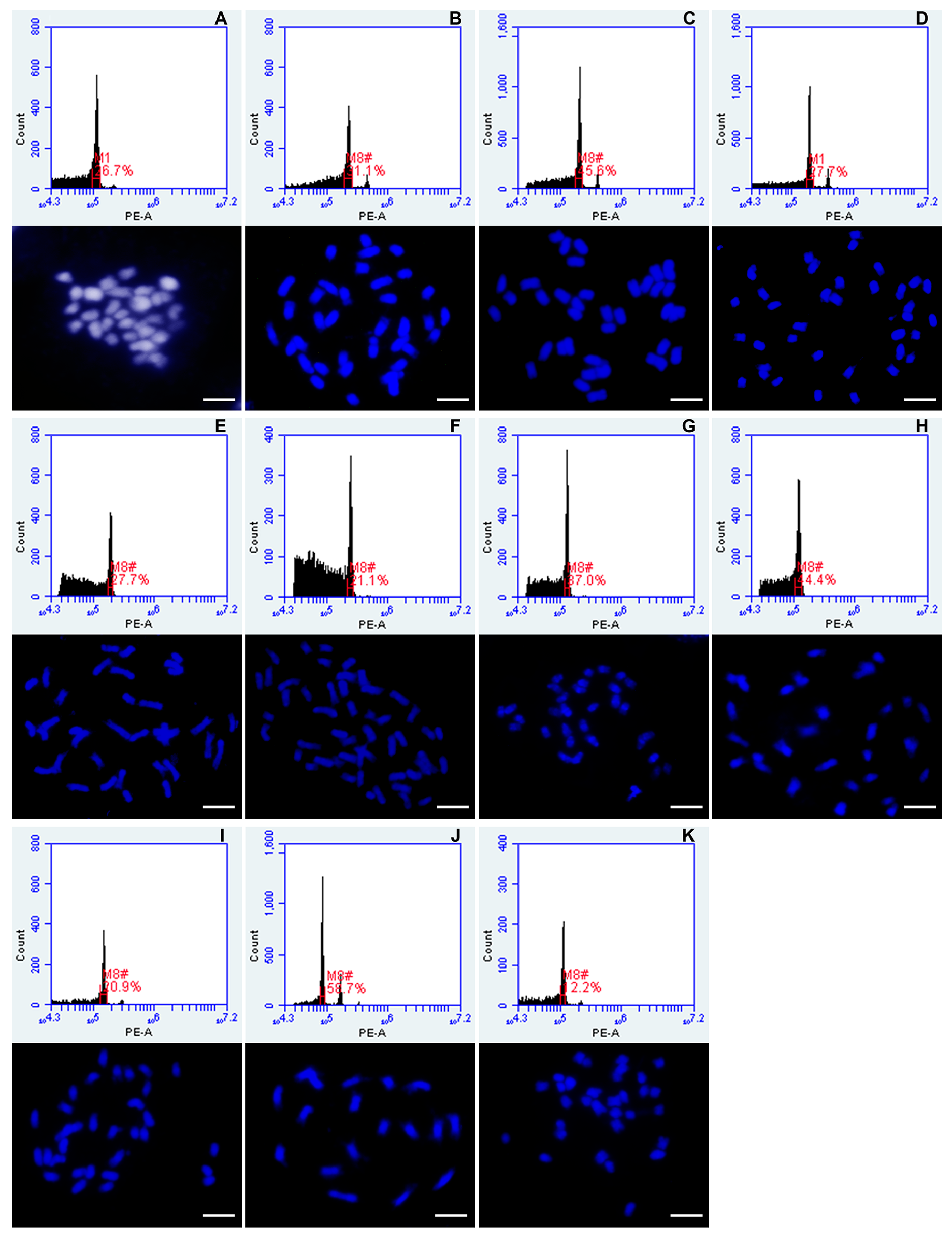
2.5. Microscopy and Validation of Chromosome Numbers
- Urochloa seeds, like many other tropical grasses, did not germinate in Petri dishes. The seeds were germinated in a 25 °C greenhouse, in 15 × 15 cm plastic pots containing Levington F2+S soil.
- Root tips were collected from plants cultivated in a greenhouse, treated with α-bromonaphthalene (Sigma B73104) at room temperature for 2 h, and 4 °C for 4 h, and fixed in absolute ethyl alcohol:acetic acid solution, 3:1.
- The root tips were washed in enzyme buffer (10 mM citric acid/sodium citrate) for 15 min, and then they underwent enzymatic maceration in 20 U/mL cellulase (Sigma C1184), 10 U/mL ‘Onozuka’ RS cellulase (RPI C32400) and 20 U/mL pectinase (Sigma P4716 from Aspergillus niger; solution in 40% glycerol) in 10 mM enzyme buffer for 60 min at 37 °C.
- Digested root tips were squashed in 60% acetic acid. Cover slips were removed after freezing with dry ice.
- Air-dried slides were counterstained with DAPI (4′,6-diamidino-2-phenylindole, Sigma D9542; 2 µg mL) in antifade solution (Citifluor, Vectashield, Slowfade or any other commercial antifading reagents for fluorescence microscopy), which prevents the permanent loss of fluorescence due to prolonged exposure to high intensity light sources.
- Slides were analyzed with an epifluorescence microscope with appropriate UV illumination, filters and camera (Nikon Eclipse 80i; DS-QiMc monochromatic camera, and NIS-Elements v.2.34 software, Nikon, Tokyo, Japan). The number of chromosomes was counted for approximately 50 metaphases derived from 5 plants of each accession used as potential external standard for flow cytometry.
3. Results
3.1. Optimization of Flow Cytometry Assay for Dried Leaves of Urochloa
3.1.1. Flow Cytometry Troubleshooting
3.1.2. Buffers
3.1.3. External Standards Used for Flow Cytometry Analysis
3.2. Ploidy Measurement of Urochloa Species
3.2.1. ‘Brizantha’ Agamic Complex
3.2.2. ‘Humidicola’ Agamic Complex
3.2.3. Urochloa Maxima
3.2.4. Related Species
4. Discussion
4.1. Flow Cytometry as a Useful Tool for Measuring the Ploidy of Large Germplasm Pool
4.2. Needs for Screening Urochloa Germplasm Collection
4.3. Choice of Flow Cytometry Approaches to Determine Ploidy from Dried Leaf Specimens
4.4. Urochloa Germplasm Findings
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- González, A.T.; Morton, C. Molecular and morphological phylogenetic analysis of Brachiaria and Urochloa (Poaceae). Mol. Phylogenetics Evol. 2005, 37, 36–44. [Google Scholar] [CrossRef]
- Renvoize, S.A.; Clayton, W.D.; Kabuye, C.H.S. Morphology, taxonomy, and natural distribution of Brachiaria (Trin.) Griseb. In Brachiaria: Biology, Agronomy, and Improvement; Miles, J.W., Maass, B.L., do Valle, C.B., Kumble, V., Eds.; Centro Internacional de Agricultura Tropical (CIAT): Rome, Italy, 1996; pp. 1–15. [Google Scholar]
- Keller-Grein, G.; Maass, B.L.; Hanson, J. Natural variation in Brachiaria and existing germplasm collections. In Brachiaria: Biology, Agronomy, and Improvement; Miles, J.W., Maass, B.L., do Valle, C.B., Kumble, V., Eds.; Centro Internacional de Agricultura Tropical (CIAT): Rome, Italy, 1996; pp. 16–39. [Google Scholar]
- Capstaff, N.M.; Miller, A.J. Improving the yield and nutritional quality of forage crops. Front. Plant Sci. 2018, 9, 535. [Google Scholar] [CrossRef] [PubMed]
- Barrios, S.C.; Valle, C.B.D.; Alves, G.F.; Simeão, R.M.; Jank, L. Reciprocal recurrent selection in the breeding of Brachiaria decumbens. Trop. Grasslands-Forrajes Trop. 2013, 1, 52. [Google Scholar] [CrossRef]
- Hanley, S.J.; Pellny, T.K.; de Vega, J.J.; Castiblanco, V.; Arango, J.; Eastmond, P.J.; Heslop-Harrison, J.S.; Mitchell, R.A.C. Allele mining in diverse accessions of Urochloa and Megathyrsus spp. tropical grasses to improve forage quality and reduce environmental impact. bioRxiv Prepr. 2020. [Google Scholar] [CrossRef]
- Baptistella, J.L.C.; De Andrade, S.A.L.; Favarin, J.L.; Mazzafera, P. Urochloa in tropical agroecosystems. Front. Sustain. Food Syst. 2020, 4, 119. [Google Scholar] [CrossRef]
- Ochatt, S.J. Flow cytometry in plant breeding. Cytom. Part A 2008, 73, 581–598. [Google Scholar] [CrossRef]
- Tomaszewska, P.; Vorontsova, M.S.; Renvoize, S.A.; Ficinski, S.Z.; Tohme, J.; Schwarzacher, T.; Castiblanco, V.; de Vega, J.J.; Mitchell, R.A.C.; Heslop-Harrison, J.S. Complex polyploid and hybrid species in an apomictic and sexual tropical forage grass group: Genomic composition and evolution in Urochloa (Brachiaria) species. bioRxiv Prepr. 2021. [Google Scholar] [CrossRef]
- Heslop-Harrison, J.S.; Schwarzacher, T. Flow cytometry and chromosome sorting. In Plant Chromosomes: Laboratory Methods; Fukui, K., Nakayama, S., Eds.; CRC: Boca Raton, FL, USA, 1996; pp. 85–108. [Google Scholar]
- Doležel, J.; Lysák, M.A.; van Den Houwe, I.; Dolezelová, M.; Roux, N. Use of flow cytometry for rapid ploidy determination in Musa species. Infomusa 1997, 6, 6–9. [Google Scholar]
- Schwarzacher, T.; Wang, M.L.; Leitch, A.R.; Moore, G.; Heslop-Harrison, J.; Miller, N. Flow cytometric analysis of the chromosomes and stability of a wheat cell-culture line. Theor. Appl. Genet. 1997, 94, 91–97. [Google Scholar] [CrossRef]
- Bennett, M.D.; Bhandol, P.; Leitch, I.J. Nuclear DNA amounts in Angiosperms and their modern uses—807 new estimates. Ann. Bot. 2000, 86, 859–909. [Google Scholar] [CrossRef]
- Śliwińska, E. Flow cytometry—A modern method for exploring genome size and nuclear DNA synthesis in horticultural and medicinal plant species. Folia Hortic. 2018, 30, 103–128. [Google Scholar] [CrossRef]
- Loureiro, J.; Rodriguez, E.; Doležel, J.; Santos, C. Flow cytometric and microscopic analysis of the effect of tannic acid on plant nuclei and estimation of DNA content. Ann. Bot. 2006, 98, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.D.; Price, H.J.; Johnston, J.S. Anthocyanin inhibits propidium iodide DNA fluorescence in Euphorbia pulcherrima: Implications for genome size variation and flow cytometry. Ann. Bot. 2007, 101, 777–790. [Google Scholar] [CrossRef] [PubMed]
- Greilhuber, J. Cytochemistry and C-values: The less-well-known world of nuclear DNA amounts. Ann. Bot. 2007, 101, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Price, H.J.; Hodnett, G.; Johnston, J.S. Sunflower (Helianthus annuus) leaves contain compounds that reduce nuclear propidium iodide fluorescence. Ann. Bot. 2000, 86, 929–934. [Google Scholar] [CrossRef]
- Jędrzejczyk, I.; Śliwińska, E. Leaves and seeds as materials for flow cytometric estimation of the genome size of 11 Rosaceae woody species containing DNA-staining inhibitors. J. Bot. 2010, 2010, 1–9. [Google Scholar] [CrossRef]
- Ooka, J.K.; Owens, D.K. Allelopathy in tropical and subtropical species. Phytochem. Rev. 2018, 17, 1225–1237. [Google Scholar] [CrossRef]
- Ribeiro, R.C.; de Carvalho, M.G.; Lopes, H.M.; Rossiello, R.O.P.; Junior, É.B. Allelopathic activity of the hydrolate and water decoction of Brachiaria humidicola (Rendle) plant parts on the germination of four tropical leguminous species. ISRN Agron. 2012, 2012, 1–6. [Google Scholar] [CrossRef]
- Ribeiro, R.C.; de Carvalho, M.G.; Moraes, M.D.L.; Rossiello, R.O.P.; de Oliveira, D.R.; de Amorim, R.M.Q.; Junior, E.B. Chemical screening of Urochloa humidicola: Methods for characterizing secondary metabolites and allelopathic activity on forage legumes. Am. J. Plant Sci. 2018, 09, 1260–1278. [Google Scholar] [CrossRef]
- De Oliveira, D.R.; Nepomuceno, D.D.; Castro, R.N.; Filho, R.B.; De Carvalho, M.G. Special metabolites isolated from Urochloa humidicola (Poaceae). An. Acad. Bras. Ciências 2017, 89, 789–797. [Google Scholar] [CrossRef]
- Feitoza, R.B.B.; Varela, R.M.; Torres, A.; Molinillo, J.M.G.; Lima, H.R.P.; Moraes, L.F.D.; Da Cunha, M.; Macías, F.A. Evaluation of the phytotoxicity of Urochloa humidicola roots by bioassays and microscopic analysis. Characterization of new compounds. J. Agric. Food Chem. 2020, 68, 4851–4864. [Google Scholar] [CrossRef] [PubMed]
- Penteado, M.I.O.; Santos, A.C.M.; Rodrigues, I.F.; do Valle, C.B.; Seixas, M.A.C.; Esteves, A. Determinação de Ploidia e Avaliação da Quantidade de DNA Total Emdiferentesespécies do Gênero Brachiaria; Embrapa Gado de Corte: Campo Grande, Brasil, 2000. [Google Scholar]
- Roche, D.; Hanna, W.W.; Ozias-Akins, P. Is supernumerary chromatin involved in gametophytic apomixis of polyploid plants? Sex. Plant Reprod. 2001, 13, 343–349. [Google Scholar] [CrossRef]
- Timbó, A.L.D.O.; Pereira, R.C.; Sobrinho, F.S.; Davide, L.C. Nuclear DNA content and chromosome number in Brachiaria spp. genotypes. Rev. Ciência Agronômica 2014, 45, 62–67. [Google Scholar] [CrossRef]
- Moraes, I.D.C.; Rume, G.D.C.; Sobrinho, F.S.; Techio, V.H. Characterization of aneuploidy in interspecific hybrid between Urochloa ruziziensis (R. Germ. & Evrard) Crins and Urochloa decumbens (Stapf) R. D. Webster. Mol. Biol. Rep. 2019, 46, 1931–1940. [Google Scholar] [CrossRef]
- Da Rocha, M.J.; Chiavegatto, R.B.; Damasceno, A.G.; Rocha, L.C.; Sobrinho, F.S.; Techio, V.H. Comparative meiosis and cytogenomic analysis in euploid and aneuploid hybrids of Urochloa P. Beauv. Chromosome Res. 2019, 27, 333–344. [Google Scholar] [CrossRef]
- de Wet, J.M.J. Hybridization and polyploidy in the Poaceae. In Grass Systematics and Evolution; Soderstrom, T.R., Hilu, W.H., Campbell, W., Barkworth, M.E., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1986; pp. 179–187. [Google Scholar]
- Basappa, G.P.; Muniyamma, M.; Chinnappa, C.C. An investigation of chromosome numbers in the genus Brachiaria (Poaceae: Paniceae) in relation to morphology and taxonomy. Can. J. Bot. 1987, 65, 2297–2309. [Google Scholar] [CrossRef]
- Bernini, C.; Marin-Morales, M. Karyotype analysis in Brachiaria (Poaceae) species. Cytobios 2001, 104, 157–171. [Google Scholar]
- Risso-Pascotto, C.; Pagliarini, M.S.; Valle, C.B.D. A new basic chromosome number for the genus Brachiaria (Trin.) Griseb. (Poaceae: Panicoideae: Paniceae). Genet. Resour. Crop. Evol. 2006, 53, 7–10. [Google Scholar] [CrossRef]
- Boldrini, K.; Pagliarini, M.; Valle, C. Meiotic behavior of a nonaploid accession endorses x = 6 for Brachiaria humidicola (Poaceae). Genet. Mol. Res. 2009, 8, 1444–1450. [Google Scholar] [CrossRef]
- Worthington, M.; Ebina, M.; Yamanaka, N.; Heffelfinger, C.; Quintero, C.; Zapata, Y.P.; Perez, J.G.; Selvaraj, M.; Ishitani, M.; Duitama, J.; et al. Translocation of a parthenogenesis gene candidate to an alternate carrier chromosome in apomictic Brachiaria humidicola. BMC Genom. 2019, 20, 41. [Google Scholar] [CrossRef]
- Suda, J.; Trávníček, P. Reliable DNA ploidy determination in dehydrated tissues of vascular plants by DAPI flow cytometry—New prospects for plant research. Cytom. Part A 2006, 69, 273–280. [Google Scholar] [CrossRef]
- Bennett, M.D.; Smith, J.B. Nuclear DNA amounts in angiosperms. Philos. Trans. R. Soc. B Biol. Sci. 1976, 274, 227–274. [Google Scholar] [CrossRef]
- Bennett, M.D.; Smith, J.B.; Heslop-Harrison, J.S. Nuclear DNA amounts in angiosperms. Proc. R. Soc. Lond. Ser. B Boil. Sci. 1982, 216, 179–199. [Google Scholar] [CrossRef]
- Arumuganathan, K.; Earle, E.D. Estimation of nuclear DNA content of plants by flow cytometry. Plant Mol. Biol. Rep. 1991, 9, 229–241. [Google Scholar] [CrossRef]
- Doležel, J.; Bartoš, J. Plant DNA flow cytometry and estimation of nuclear genome size. Ann. Bot. 2005, 95, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.D.; Leitch, I.J. Nuclear DNA amounts in angiosperms: Targets, trends and tomorrow. Ann. Bot. 2011, 107, 467–590. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, J.; Rodriguez, E.; Dolezel, J.; Santos, C. Comparison of four nuclear isolation buffers for plant DNA flow cytometry. Ann. Bot. 2006, 98, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.V. The use of bead beating to prepare suspensions of nuclei for flow cytometry from fresh leaves, herbarium leaves, petals and pollen. Cytom. Part A 2007, 71, 1039–1044. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef]
- Hunt, H.V.; Badakshi, F.; Romanova, O.; Howe, C.J.; Jones, M.K.; Heslop-Harrison, J. (Pat) Reticulate evolution in Panicum (Poaceae): The origin of tetraploid broomcorn millet, P. miliaceum. J. Exp. Bot. 2014, 65, 3165–3175. [Google Scholar] [CrossRef]
- Galbraith, D.W.; Lambert, G.M. Using the BD AccuriTM C6 cytometer for rapid and accurate analysis of the nuclear DNA contents of flowering plants. BD Bioscience 2021, 1, 1–10. [Google Scholar]
- Galbraith, D.W.; Harkins, K.R.; Maddox, J.R.; Ayres, N.M.; Sharma, D.P.; Firoozabady, E. Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 1983, 220, 1049–1051. [Google Scholar] [CrossRef] [PubMed]
- Otto, F. Preparation and staining of cells for high-resolution DNA analysis. In Flow Cytometry and Cell Sorting; Radbruch, A., Ed.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 101–104. [Google Scholar]
- de Laat, A.M.M.; Göhde, W.; Vogelzang, J.D.C. Determination of ploidy of single plants and plant populations by flow cytometry. Plant Breed. 1987, 99, 303–307. [Google Scholar] [CrossRef]
- Schwarzacher, T.; Heslop-Harrison, J.S. Practical In Situ Hybridization; Bios: Oxford, UK, 2000. [Google Scholar]
- Salariato, D.L.; Zuloaga, F.O.; Giussani, L.M.; Morrone, O. Molecular phylogeny of the subtribe Melinidinae (Poaceae: Panicoideae: Paniceae) and evolutionary trends in the homogenization of inflorescences. Mol. Phylogenetics Evol. 2010, 56, 355–369. [Google Scholar] [CrossRef]
- Salariato, D.L.; Morrone, O.; Zuloaga, F.O. Mayariochloa, a new monotypic genus segregated from Scutachne (Poaceae, Panicoideae, Paniceae). Syst. Bot. 2012, 37, 105–116. [Google Scholar] [CrossRef]
- Šmarda, P.; Stančík, D. Ploidy level variability in South American Fescues (Festuca L., Poaceae): Use of flow cytometry in up to 5 ½-year-old caryopses and herbarium specimens. Plant Biol. 2008, 8, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Yang, Y. The effects of fresh and rapid desiccated tissue on estimates o Ophiopogoneae genome size. Plant Divers. 2016, 38, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Alves, G.F.; De Figueiredo, U.J.; Filho, A.D.P.; Barrios, S.C.; Valle, C.B.D. Breeding strategies for Brachiaria spp. to improve productivity—An ongoing project. Trop. Grasslands-Forrajes Trop. 2014, 2, 1. [Google Scholar] [CrossRef]
- Matias, F.I.; Barrios, S.C.L.; Valle, C.B.D.; Mateus, R.G.; Martins, L.B.; Moro, G.V. Estimate of genetic parameters in Brachiaria decumbens hybrids. Crop. Breed. Appl. Biotechnol. 2016, 16, 115–122. [Google Scholar] [CrossRef]
- Soltis, D.E.; Soltis, P.S. The dynamic nature of polyploid genomes. Proc. Natl. Acad. Sci. USA 1995, 92, 8089–8091. [Google Scholar] [CrossRef]
- Francis, D.; Davies, M.S.; Barlow, P.W. A strong nucleotypic effect on the cell cycle regardless of ploidy level. Ann. Bot. 2008, 101, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, R.; Boudolf, V.; Cuevas, F.; Lu, R.; Eekhout, T.; Hu, Z.; Van Isterdael, G.; Lambert, G.M.; Xu, F.; Nowack, M.K.; et al. A spatiotemporal DNA endoploidy map of the Arabidopsis root reveals roles for the endocycle in root development and stress adaptation. Plant Cell 2018, 30, 2330–2351. [Google Scholar] [CrossRef] [PubMed]
- Mgwatyu, Y.; Stander, A.A.; Ferreira, S.; Williams, W.; Hesse, U. Rooibos (Aspalathus linearis) genome size estimation using flow cytometry and k-mer analyses. Plants 2020, 9, 270. [Google Scholar] [CrossRef] [PubMed]
- Samanta, A.; Paul, B.K.; Guchhait, N. Photophysics of DNA staining dye propidium iodide encapsulated in bio-mimetic micelle and genomic fish sperm DNA. J. Photochem. Photobiol. B Biol. 2012, 109, 58–67. [Google Scholar] [CrossRef]
- Krahulcová, A.; Rotreklová, O. Use of flow cytometry in research on apomictic plants. Preslia 2010, 82, 23–39. [Google Scholar]
- Ishigaki, G.; Gondo, T.; Ebina, M.; Suenaga, K.; Akashi, R. Estimation of genome size in Brachiaria species. Grassl. Sci. 2010, 56, 240–242. [Google Scholar] [CrossRef]
- Boldrini, K.; Micheletti, P.; Gallo, P.; Mendes-Bonato, A.; Pagliarini, M.; Valle, C. Origin of a polyploid accession of Brachiaria humidicola (Poaceae: Panicoideae: Paniceae). Genet. Mol. Res. 2009, 8, 888–895. [Google Scholar] [CrossRef]
- Triviño, N.J.; Perez, J.G.; Recio, M.E.; Ebina, M.; Yamanaka, N.; Tsuruta, S.-I.; Ishitani, M.; Worthington, M. Genetic diversity and population structure of Brachiaria species and breeding populations. Crop. Sci. 2017, 57, 2633–2644. [Google Scholar] [CrossRef]
- Jank, L.; Valle, C.B.; Resende, R. Breeding tropical forages. Crop. Breed. Appl. Biotechnol. 2011, 11, 27–34. [Google Scholar] [CrossRef]
- Jungmann, L. Caracterização da Diversidadegenética Molecular Emgermoplasma de Brachiaria spp. Ph.D. Thesis, Universidade Estadual de Campinas, São Paulo, Brasil, 2009. [Google Scholar]
- Jungmann, L.; Vigna, B.B.; Boldrini, K.R.; Sousa, A.C.; Valle, C.B.D.; Resende, R.M.; Pagliarini, M.S.; Zucchi, M.I.; De Souza, A.P. Genetic diversity and population structure analysis of the tropical pasture grass Brachiaria humidicola based on microsatellites, cytogenetics, morphological traits, and geographical origin. Genome 2010, 53, 698–709. [Google Scholar] [CrossRef]
- Nitthaisong, P.; Ishigaki, G.; Tanaka, H.; Akashi, R. Chromosome number, genomic variation, and molecular markers to assess genetic diversity of Brachiaria species. Crop. Sci. 2016, 56, 312–321. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| External Standard | Number of Chromosomes | Mean Fluorescence Values of the Peak Positions | Sample Species |
|---|---|---|---|
| Panicum miliaceum VIR, Mil69 | 2n = 4x = 36 | 112 | - |
| Urochloa brizantha, USDA, PI 292187 | 2n = 4x = 36 | 225 | U. brizantha, U. dura, U. platynota, U. ruziziensis × U. decumbens × U. brizantha hybrid |
| Urochloa decumbens, CIAT, 664 | 2n = 4x = 36 | 205 | U. arrecta, U. decumbens, U. jubata, U. nigropedata, U. plantaginea, U. ruziziensis |
| Urochloa decumbens, CIAT, 6370 | 2n = 4x = 36 | 194 | U. arrecta, U. decumbens, U. jubata, U. nigropedata, U. plantaginea, U. ruziziensis |
| Urochloa humidicola, CIAT, 26151 | 2n = 6x = 36 | 197 | U. humidicola, U. dictyoneura |
| Urochloa maxima, CIAT, 6171 | 2n = 4x = 32 | 131 | U. maxima, U. nigropedata |
| Urochloa maxima, CIAT, 16004 | 2n = 4x = 32 | 120 | U. maxima, U. nigropedata |
| Urochloa maxima, USDA, PI 284156 | 2n = 4x = 32 | 149 | U. maxima, U. nigropedata |
| Urochloa ruziziensis, CIAT, 6419 | 2n = 2x = 18 | 83 | U. arrecta, U. decumbens, U. jubata, U. nigropedata, U. plantaginea, U. ruziziensis |
| Urochloa sp., USDA, PI 657653 | 2n = 4x = 32 | 111 | U. maxima |
| Buffer | Composition |
|---|---|
| Galbraith [47] | 45 mM MgCl2, 20 mM MOPS, 30 mM sodium citrate, 0.1% (v/v) Triton X-100 (pH 7) |
| Otto [48] | Otto I: 100 mM citric acid, 0.5 % (v/v) Tween 20 (pH 2–3)Otto II: 400 mM Na2PO4 · 12H2O (pH 8–9) |
| Modified Otto (this paper) | Otto I: 100 mM citric acid, 0.5 % (v/v) Tween 20 (pH 2–3)Otto II: 400 mM Na2PO4 · 12H2O (pH 8–9), 15 mM β-mercaptoethanol, 1% PVP-40 |
| Partec [49] | 100 mM Tris, 2.5 mM MgCl2 · 6H2O, 85 mM NaCl, 0.1% (v/v) Triton X-100 (pH 7,0) |
| Modified Partec (this paper) | 100 mM Tris, 2.5 mM MgCl2 · 6H2O, 85 mM NaCl, 0.1% (v/v) Triton X-100 (pH 7.0), 15 mM β-mercaptoethanol, 1% PVP-40 |
| Species | Ploidy | Number of Studied Accessions | Number of Studied Plants | Range of Fluorescence Values of the Peak Positions | Mean Fluorescence Values of the Peak Positions | CV (%) Range | CV (%) Average |
|---|---|---|---|---|---|---|---|
| Urochloa arrecta | 2n = 4x | 1 | 1 | 93 | 93 | 7.53 | 7.53 |
| Urochloa brizantha | 2n = 2x | 6 | 9 | 82–110 | 96 | 5.46–9.14 | 7.32 |
| 2n = 4x | 59 | 70 | 111–225 | 172 | 2.9–9.89 | 5.65 | |
| 2n = 5x | 25 | 37 | 215–291 | 247 | 3.4–8.17 | 5.34 | |
| 2n = 6x | 1 | 1 | 303 | 303 | 3.83 | 3.83 | |
| Urochloa decumbens | 2n = 2x | 18 | 19 | 72–104 | 87 | 2.68–6.87 | 4.76 |
| 2n = 4x | 25 | 28 | 152–211 | 183 | 3.25–5.66 | 4.52 | |
| 2n = 6x | 1 | 1 | 270 | 270 | 4.02 | 4.02 | |
| Urochloa dictyoneura | 2n = 7x | 1 | 1 | 220 | 220 | 5.91 | 5.91 |
| Urochloa dura | 2n = 5x | 1 | 2 | 255–282 | 268 | 4.82–5.18 | 5 |
| Urochloa humidicola | 2n = 6x | 16 | 21 | 108–205 | 174 | 3.69–6.24 | 4.65 |
| 2n = 7x | 33 | 45 | 215–298 | 259 | 2.84–6.4 | 4.31 | |
| 2n = 8x + 2 or 9x − 4 | 1 | 2 | 253–259 | 256 | 3.04–3.49 | 3.27 | |
| 2n = 9x | 3 | 4 | 320–338 | 330 | 3.39–5.33 | 4.7 | |
| Urochloa jubata | 2n = 2x | 1 | 1 | 87 | 87 | 5.89 | 5.89 |
| 2n = 4x | 1 | 1 | 123 | 123 | 4.73 | 4.73 | |
| Urochloa maxima | 2n = 2x | 25 | 31 | 74–104 | 94 | 4.81–9.23 | 7.02 |
| 2n = 4x | 99 | 102 | 104–190 | 128 | 3.73–8.81 | 5.5 | |
| Urochloa nigropedata | 2n = 4x | 1 | 2 | 142–146 | 144 | 3.91–6.53 | 5.22 |
| Urochloa plantaginea | 2n = 2x | 1 | 1 | 90 | 90 | 6.62 | 6.62 |
| Urochloa platynota | 2n = 2x | 1 | 1 | 98 | 98 | 5.96 | 5.96 |
| Urochloa ruziziensis | 2n = 2x | 26 | 33 | 75–103 | 86 | 2.42–6.92 | 4.32 |
| Urochloa ruziziensis × Urochloa decumbens × Urochloa brizantha | 2n = 4x | 1 | 1 | 190 | 190 | 2.67 | 2.67 |
| Urochloa sp. PI657653 | 2n = 4x | 1 | 1 | 111 | 111 | 4.42 | 4.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomaszewska, P.; Pellny, T.K.; Hernández, L.M.; Mitchell, R.A.C.; Castiblanco, V.; de Vega, J.J.; Schwarzacher, T.; Heslop-Harrison, P. Flow Cytometry-Based Determination of Ploidy from Dried Leaf Specimens in Genomically Complex Collections of the Tropical Forage Grass Urochloa s. l. Genes 2021, 12, 957. https://doi.org/10.3390/genes12070957
Tomaszewska P, Pellny TK, Hernández LM, Mitchell RAC, Castiblanco V, de Vega JJ, Schwarzacher T, Heslop-Harrison P. Flow Cytometry-Based Determination of Ploidy from Dried Leaf Specimens in Genomically Complex Collections of the Tropical Forage Grass Urochloa s. l. Genes. 2021; 12(7):957. https://doi.org/10.3390/genes12070957
Chicago/Turabian StyleTomaszewska, Paulina, Till K. Pellny, Luis M. Hernández, Rowan A. C. Mitchell, Valheria Castiblanco, José J. de Vega, Trude Schwarzacher, and Pat (J.S.) Heslop-Harrison. 2021. "Flow Cytometry-Based Determination of Ploidy from Dried Leaf Specimens in Genomically Complex Collections of the Tropical Forage Grass Urochloa s. l." Genes 12, no. 7: 957. https://doi.org/10.3390/genes12070957
APA StyleTomaszewska, P., Pellny, T. K., Hernández, L. M., Mitchell, R. A. C., Castiblanco, V., de Vega, J. J., Schwarzacher, T., & Heslop-Harrison, P. (2021). Flow Cytometry-Based Determination of Ploidy from Dried Leaf Specimens in Genomically Complex Collections of the Tropical Forage Grass Urochloa s. l. Genes, 12(7), 957. https://doi.org/10.3390/genes12070957