
Maternal One-Carbon Metabolism during the Periconceptional Period and Human Foetal Brain Growth: A Systematic Review

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
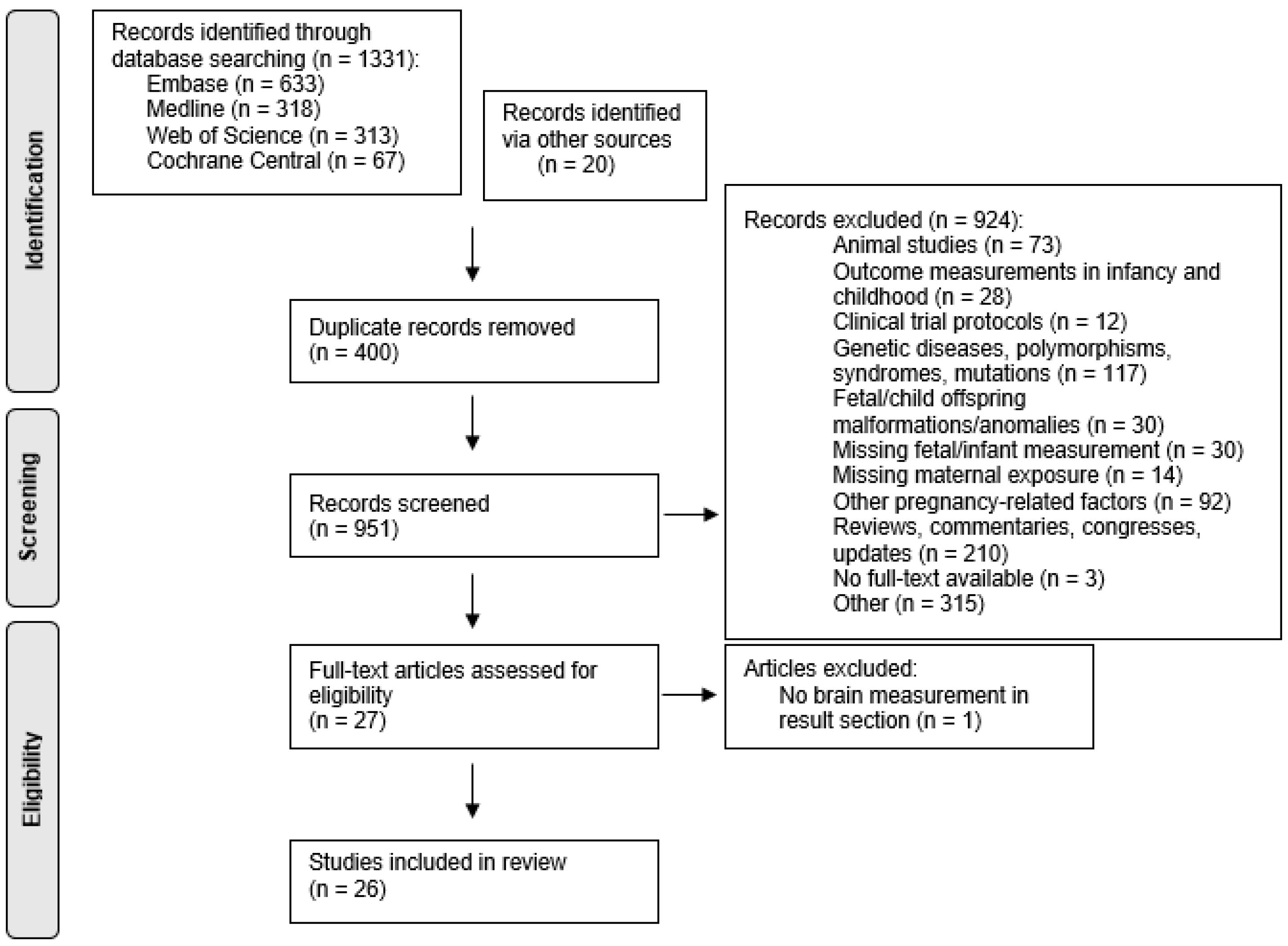
2. Materials and Methods
2.1. Search Strategy
2.2. Eligibility Criteria
2.3. Quality Assessment
3. Results
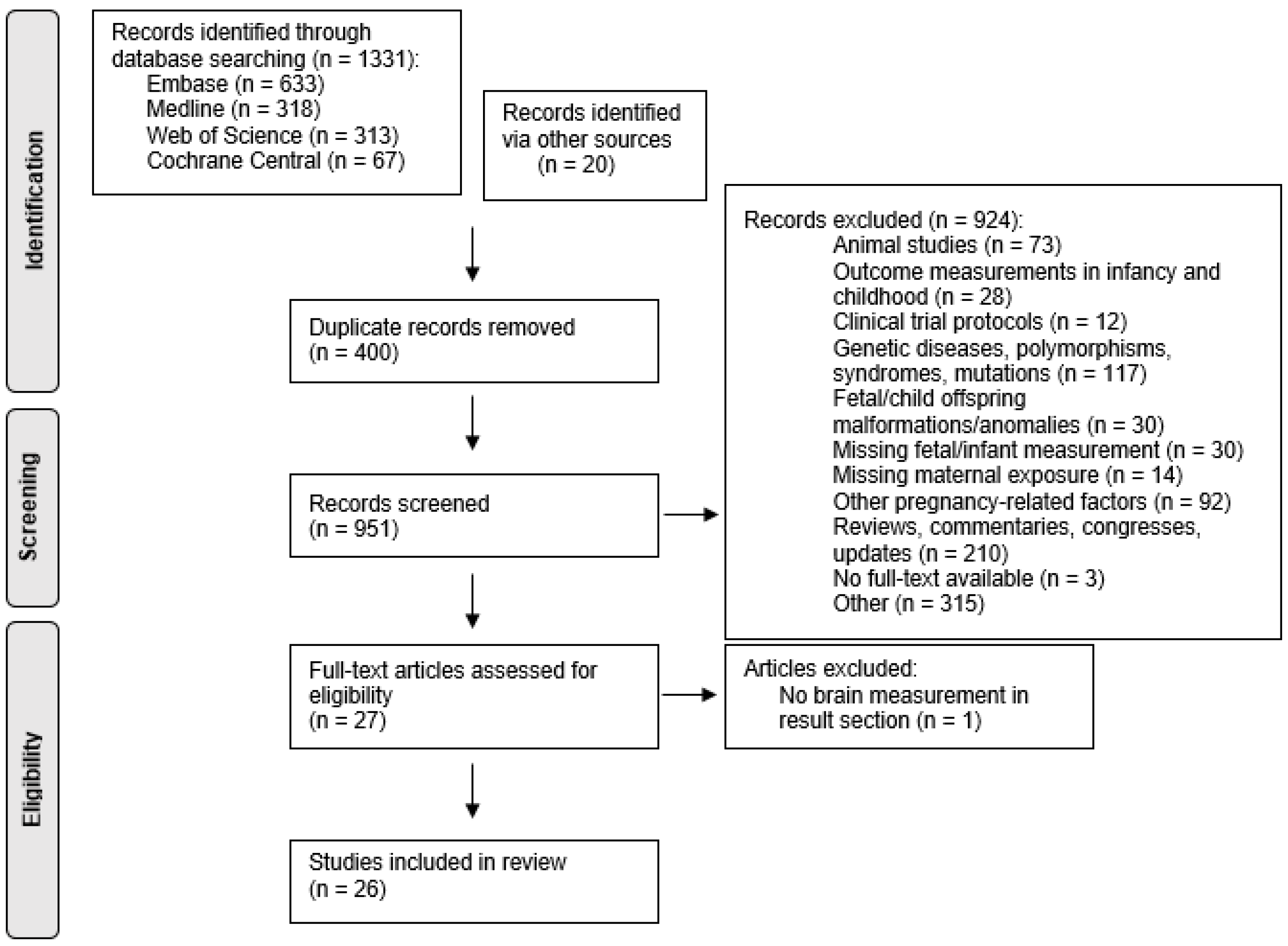
3.1. Study Selection
3.2. Vitamin B6
3.3. Vitamin B9/11 (Folate)
3.3.1. Erythrocyte, Serum and Plasma Folate Concentrations
3.3.2. Dietary Intake of Folate
3.3.3. Folic Acid Supplement Use
3.4. Vitamin B12
3.5. Choline
3.6. Total Homocysteine
3.7. Maternal Dietary Patterns
4. Discussion
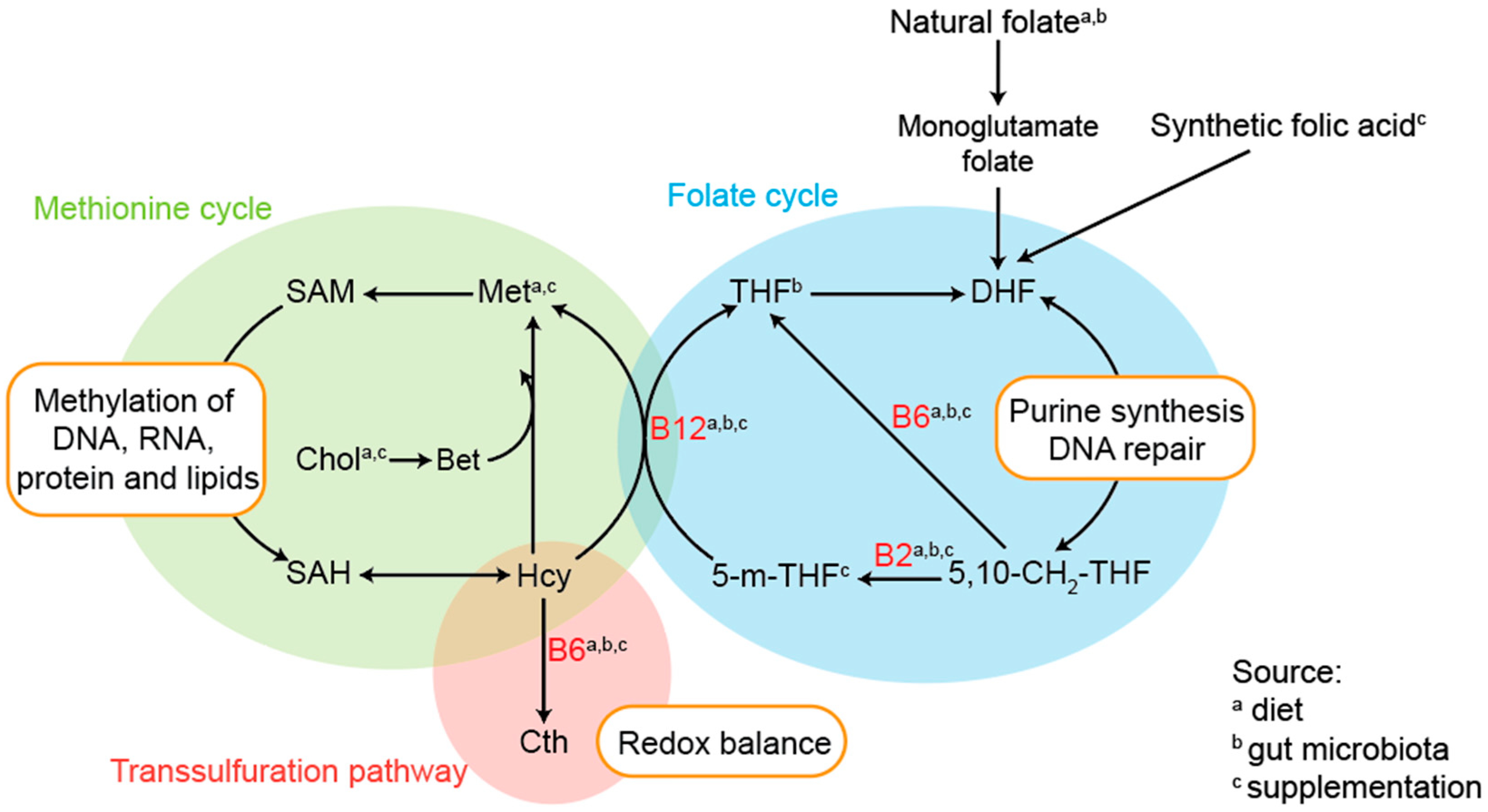
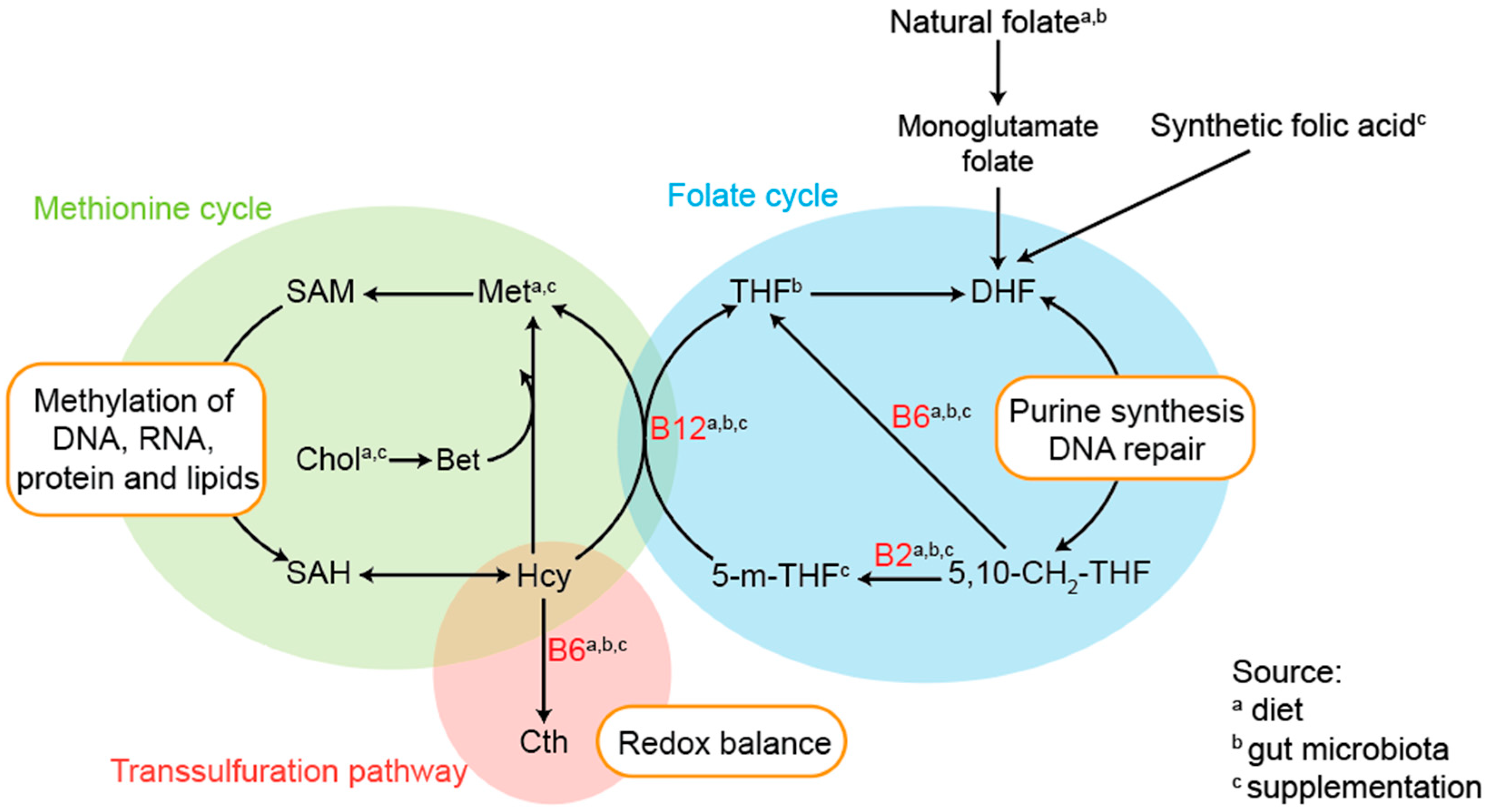
4.1. One-Carbon Metabolism
4.1.1. Folate
4.1.2. Folic Acid Supplement Use
4.1.3. Dietary Patterns
4.1.4. Vitamin B6
4.1.5. Vitamin B12
4.1.6. Homocysteine
4.1.7. Choline
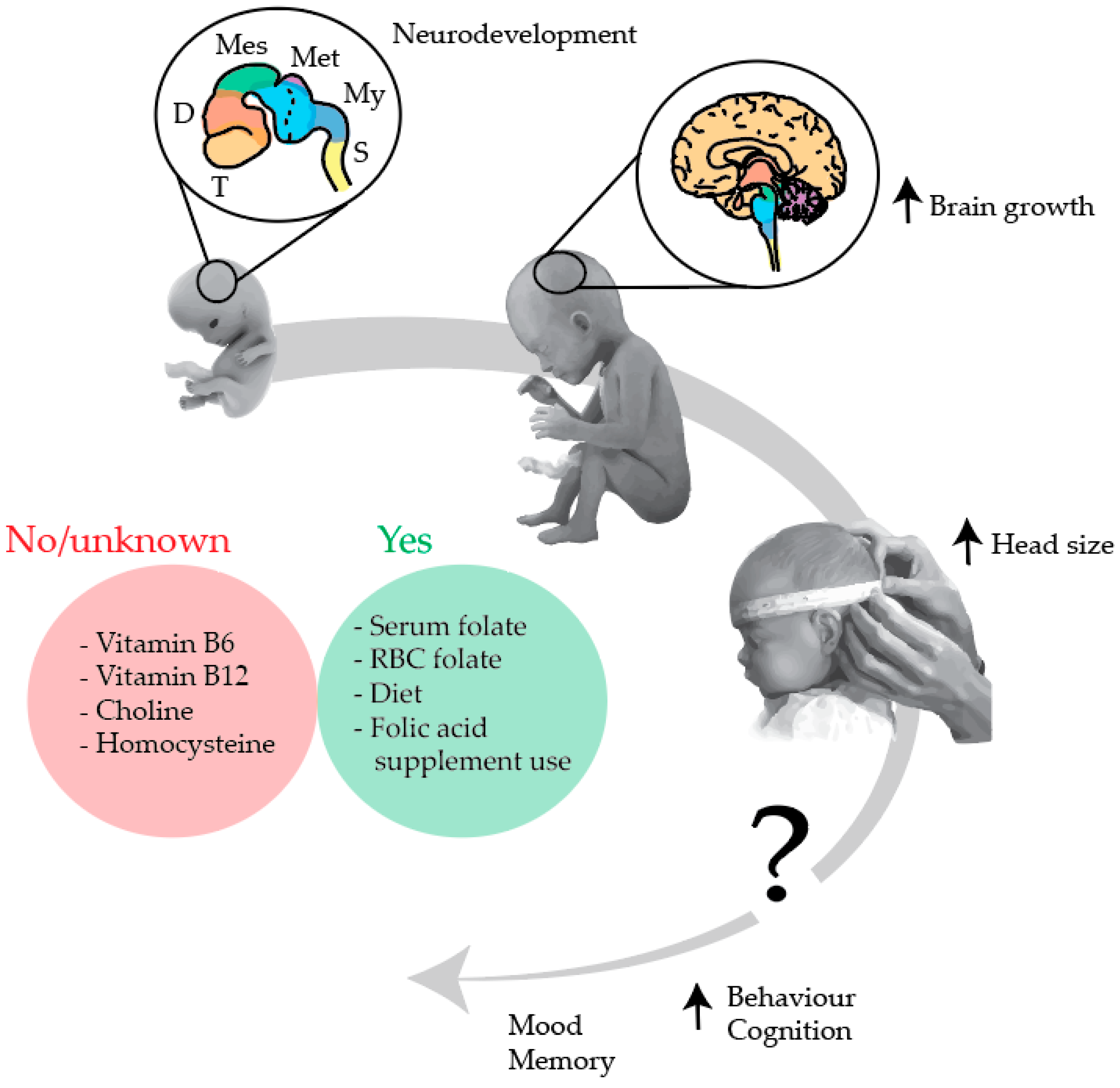
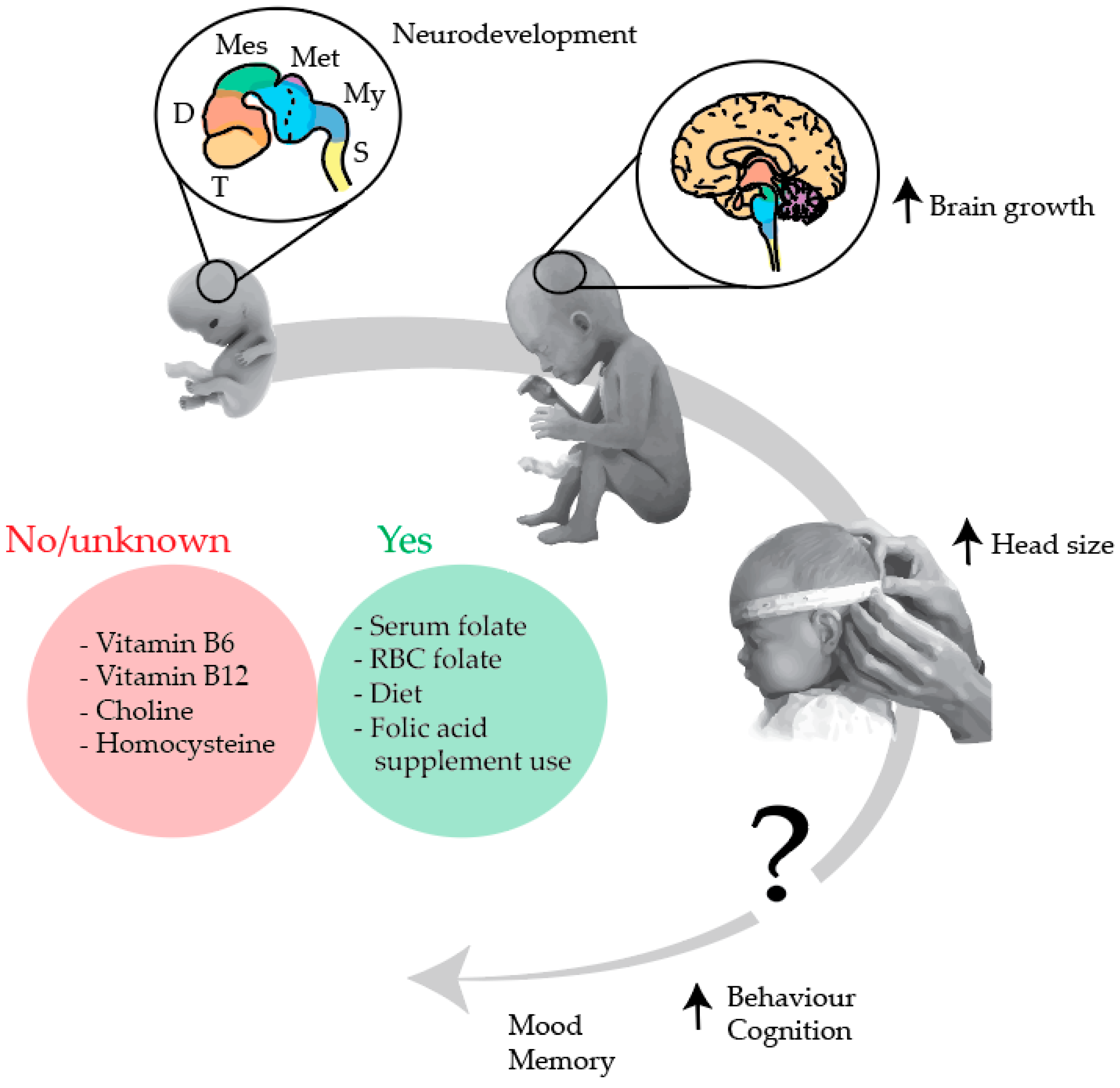
4.2. Main Interpretation
4.3. Clinical Relevance and Future Research
4.4. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barker, D.J. The origins of the developmental origins theory. J. Intern. Med. 2007, 261, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Fleming, T.P.; Watkins, A.J.; Velazquez, M.A.; Mathers, J.C.; Prentice, A.M.; Stephenson, J.; Barker, M.; Saffery, R.; Yajnik, C.S.; Eckert, J.J.; et al. Origins of lifetime health around the time of conception: Causes and consequences. Lancet 2018, 391, 1842–1852. [Google Scholar] [CrossRef]
- Hanson, M.A.; Gluckman, P.D. Early developmental conditioning of later health and disease: Physiology or pathophysiology? Physiol. Rev. 2014, 94, 1027–1076. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Quan, S.; Yang, G.; Chen, M.; Ye, Q.; Wang, G.; Yu, H.; Wang, Y.; Qiao, S.; Zeng, X. Nutritional Status Impacts Epigenetic Regulation in Early Embryo Development: A Scoping Review. Adv. Nutr. 2021, 12, 6238544. [Google Scholar] [CrossRef]
- Li, Y. Epigenetic Mechanisms Link Maternal Diets and Gut Microbiome to Obesity in the Offspring. Front. Genet. 2018, 9, 342. [Google Scholar] [CrossRef]
- Wiklund, P.; Karhunen, V.; Richmond, R.C.; Parmar, P.; Rodriguez, A.; De Silva, M.; Wielscher, M.; Rezwan, F.I.; Richardson, T.G.; Veijola, J.; et al. DNA methylation links prenatal smoking exposure to later life health outcomes in offspring. Clin. Epigenet. 2019, 11, 97. [Google Scholar] [CrossRef] [Green Version]
- Ungerer, M.; Knezovich, J.; Ramsay, M. In utero alcohol exposure, epigenetic changes, and their consequences. Alcohol. Res. 2013, 35, 37–46. [Google Scholar]
- Ornoy, A.; Reece, E.A.; Pavlinkova, G.; Kappen, C.; Miller, R.K. Effect of maternal diabetes on the embryo, fetus, and children: Congenital anomalies, genetic and epigenetic changes and developmental outcomes. Birth Defects Res. C Embryo Today 2015, 105, 53–72. [Google Scholar] [CrossRef]
- DeBaun, M.R.; Niemitz, E.L.; Feinberg, A.P. Association of in vitro fertilization with Beckwith-Wiedemann syndrome and epigenetic alterations of LIT1 and H19. Am. J. Hum. Genet. 2003, 72, 156–160. [Google Scholar] [CrossRef] [Green Version]
- Lehnen, H.; Zechner, U.; Haaf, T. Epigenetics of gestational diabetes mellitus and offspring health: The time for action is in early stages of life. Mol. Hum. Reprod 2013, 19, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Francis, E.; Hinkle, S.N.; Ajjarapu, A.S.; Zhang, C. Preconception and Prenatal Nutrition and Neurodevelopmental Disorders: A Systematic Review and Meta-Analysis. Nutrients 2019, 11, 1628. [Google Scholar] [CrossRef] [Green Version]
- Steegers-Theunissen, R.P.; Twigt, J.; Pestinger, V.; Sinclair, K.D. The periconceptional period, reproduction and long-term health of offspring: The importance of one-carbon metabolism. Hum. Reprod. Update 2013, 19, 640–655. [Google Scholar] [CrossRef] [Green Version]
- Clare, C.E.; Pestinger, V.; Kwong, W.Y.; Tutt, D.A.R.; Xu, J.; Byrne, H.M.; Barrett, D.A.; Emes, R.D.; Sinclair, K.D. Interspecific Variation in One-Carbon Metabolism within the Ovarian Follicle, Oocyte, and Preimplantation Embryo: Consequences for Epigenetic Programming of DNA Methylation. Int. J. Mol. Sci. 2021, 22, 1838. [Google Scholar] [CrossRef] [PubMed]
- Gurwara, S.; Ajami, N.J.; Jang, A.; Hessel, F.C.; Chen, L.; Plew, S.; Wang, Z.; Graham, D.Y.; Hair, C.; White, D.L.; et al. Dietary Nutrients Involved in One-Carbon Metabolism and Colonic Mucosa-Associated Gut Microbiome in Individuals with an Endoscopically Normal Colon. Nutrients 2019, 11, 613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, M.M. Effects of vitamin B12 and folate deficiency on brain development in children. Food Nutr. Bull. 2008, 29, S126–S131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaevitz, L.; Berger-Sweeney, J.; Ricceri, L. One-carbon metabolism in neurodevelopmental disorders: Using broad-based nutraceutics to treat cognitive deficits in complex spectrum disorders. Neurosci. Biobehav. Rev. 2014, 46, 270–284. [Google Scholar] [CrossRef] [PubMed]
- Keverne, E.B.; Pfaff, D.W.; Tabansky, I. Epigenetic changes in the developing brain: Effects on behavior. Proc. Natl. Acad. Sci. USA 2015, 112, 6789–6795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrototaro, G.; Zaghi, M.; Sessa, A. Epigenetic Mistakes in Neurodevelopmental Disorders. J. Mol. Neurosci. 2017, 61, 590–602. [Google Scholar] [CrossRef]
- Imbard, A.; Benoist, J.F.; Blom, H.J. Neural tube defects, folic acid and methylation. Int. J. Environ. Res. Public Health 2013, 10, 4352–4389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steegers-Theunissen, R.P.; Boers, G.H.; Blom, H.J.; Nijhuis, J.G.; Thomas, C.M.; Borm, G.F.; Eskes, T.K. Neural tube defects and elevated homocysteine levels in amniotic fluid. Am. J. Obstet. Gynecol. 1995, 172, 1436–1441. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.L.; McPartlin, J.M.; Kirke, P.N.; Lee, Y.J.; Conley, M.R.; Weir, D.G.; Scott, J.M. Homocysteine metabolism in pregnancies complicated by neural-tube defects. Lancet 1995, 345, 149–151. [Google Scholar] [CrossRef]
- Freedman, R.; Hunter, S.K.; Law, A.J.; Clark, A.M.; Roberts, A.; Hoffman, M.C. Choline, folic acid, Vitamin D, and fetal brain development in the psychosis spectrum. Schizophr. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Ars, C.L.; Nijs, I.M.; Marroun, H.E.; Muetzel, R.; Schmidt, M.; Steenweg-De Graaff, J.; Van Der Lugt, A.; Jaddoe, V.W.; Hofman, A.; Steegers, E.A.; et al. Prenatal folate, homocysteine and Vitamin B12 levels and child brain volumes, cognitive development and psychological functioning: The Generation R Study. Br. J. Nutr. 2019, 122, S1–S9. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, R.; Riley, A.W.; Volk, H.; Caruso, D.; Hironaka, L.; Sices, L.; Hong, X.; Wang, G.; Ji, Y.; Brucato, M.; et al. Maternal Multivitamin Intake, Plasma Folate and Vitamin B12 Levels and Autism Spectrum Disorder Risk in Offspring. Paediatr. Perinat. Epidemiol. 2018, 32, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Naninck, E.F.G.; Stijger, P.C.; Brouwer-Brolsma, E.M. The Importance of Maternal Folate Status for Brain Development and Function of Offspring. Adv. Nutr. 2019, 10, 502–519. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, E.; Campbell, K.; Solso, S. Brain growth across the life span in autism: Age-specific changes in anatomical pathology. Brain Res. 2011, 1380, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Sacco, R.; Gabriele, S.; Persico, A.M. Head circumference and brain size in autism spectrum disorder: A systematic review and meta-analysis. Psychiatry Res. 2015, 234, 239–251. [Google Scholar] [CrossRef]
- Brambilla, P.; Hardan, A.; di Nemi, S.U.; Perez, J.; Soares, J.C.; Barale, F. Brain anatomy and development in autism: Review of structural MRI studies. Brain Res. Bull. 2003, 61, 557–569. [Google Scholar] [CrossRef]
- Page, M.J.; Moher, D.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. PRISMA 2020 explanation and elaboration: Updated guidance and exemplars for reporting systematic reviews. BMJ 2021, 372, n160. [Google Scholar] [CrossRef]
- Hamilton, O. Quality Assessment Tool for Quantitative Studies; National Collaborating Centre for Methods and Tools: Hamilton, ON, Canada, 2008. [Google Scholar]
- Takimoto, H.; Hayashi, F.; Kusama, K.; Kato, N.; Yoshiike, N.; Toba, M.; Ishibashi, T.; Miyasaka, N.; Kubota, T. Elevated maternal serum folate in the third trimester and reduced fetal growth: A longitudinal study. J. Nutr. Sci. Vitaminol. 2011, 57, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Brough, L.; Rees, G.A.; Crawford, M.A.; Morton, R.H.; Dorman, E.K. Effect of multiple-micronutrient supplementation on maternal nutrient status, infant birth weight and gestational age at birth in a low-income, multi-ethnic population. Br. J. Nutr. 2010, 104, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Schlotz, W.; Jones, A.; Phillips, D.I.; Gale, C.R.; Robinson, S.M.; Godfrey, K.M. Lower maternal folate status in early pregnancy is associated with childhood hyperactivity and peer problems in offspring. J. Child. Psychol. Psychiatry 2010, 51, 594–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steenweg-de Graaff, J.; Roza, S.J.; Walstra, A.N.; El Marroun, H.; Steegers, E.A.P.; Jaddoe, V.W.V.; Hofman, A.; Verhulst, F.C.; Tiemeier, H.; White, T. Associations of maternal folic acid supplementation and folate concentrations during pregnancy with foetal and child head growth: The Generation R Study. Eur. J. Nutr. 2017, 56, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, R.; El Marroun, H.; Cecil, C.; Jaddoe, V.W.V.; Hillegers, M.; Tiemeier, H.; White, T. Maternal folate levels during pregnancy and offspring brain development in late childhood. Clin. Nutr. 2020, 3391–3400. [Google Scholar] [CrossRef]
- Koning, I.V.; Groenenberg, I.A.L.; Gotink, A.W.; Willemsen, S.P.; Gijtenbeek, M.; Dudink, J.; Go, A.T.J.I.; Reiss, I.K.M.; Steegers, E.A.P.; Steegers-Theunissen, R.P.M. Periconception maternal folate status and human embryonic cerebellum growth trajectories: The Rotterdam predict study. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsen, R.M.; Vollset, S.E.; Monsen, A.L.B.; Ulvik, A.; Haugen, M.; Meltzer, H.M.; Magnus, P.; Ueland, P.M. Infant birth size is not associated with maternal intake and status of folate during the second trimester in Norwegian pregnant women. J. Nutr. 2010, 140, 572–579. [Google Scholar] [CrossRef]
- Takimoto, H.; Mito, N.; Umegaki, K.; Ishiwaki, A.; Kusama, K.; Abe, S.; Yamawaki, M.; Fukuoka, H.; Ohta, C.; Yoshiike, N. Relationship between dietary folate intakes, maternal plasma total homocysteine and B-vitamins during pregnancy and fetal growth in Japan. Eur J. Nutr. 2007, 46, 300–306. [Google Scholar] [CrossRef]
- Jiang, H.L.; Cao, L.Q.; Chen, H.Y. Blood folic acid, vitamin B12, and homocysteine levels in pregnant women with fetal growth restriction. Genet. Mol. Res. 2016, 15, gmr15048890. [Google Scholar] [CrossRef]
- Bergen, N.E.; Schalekamp-Timmermans, S.; Jaddoe, V.W.V.; Hofman, A.; Lindemans, J.; Russcher, H.; Tiemeier, H.; Steegers-Theunissen, R.P.M.; Steegers, E.A.P. Maternal and Neonatal Markers of the Homocysteine Pathway and Fetal Growth: The Generation R Study. Paediatr. Perinat. Epidemiol. 2016, 30, 386–396. [Google Scholar] [CrossRef]
- Gadgil, M.; Joshi, K.; Pandit, A.; Otiv, S.; Joshi, R.; Brenna, J.T.; Patwardhan, B. Imbalance of folic acid and vitamin B12 is associated with birth outcome: An Indian pregnant women study. Eur J. Clin. Nutr. 2014, 68, 726–729. [Google Scholar] [CrossRef]
- WHO. Serum and Red Blood Cell Folate Concentrations for Assessing Folate Status in Populations; WHO: Geneva, Switzerland, 2012. [Google Scholar]
- Timmermans, S.; Jaddoe, V.W.V.; Hofman, A.; Steegers-Theunissen, R.P.M.; Steegers, E.A.P. Periconception folic acid supplementation, fetal growth and the risks of low birth weight and preterm birth: The Generation R Study. Br. J. Nutr. 2009, 102, 777–785. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, K.K.; Salihu, H.M.; Wilson, R.; Mbah, A.; Sappenfield, W.; Bruder, K.; Wudil, U.J.; Aliyu, M.H. Folic Acid Intake, Fetal Brain Growth, and Maternal Smoking in Pregnancy: A Randomized Controlled Trial. Curr. Dev. Nutr. 2019, 3, nzz025. [Google Scholar] [CrossRef]
- Christian, P.; Khatry, S.K.; Katz, J.; Pradhan, E.K.; LeClerq, S.C.; Shrestha, S.R.; Adhikari, R.K.; Sommer, A.; West, K.P., Jr. Effects of alternative maternal micronutrient supplements on low birth weight in rural nepal: Double blind randomised community trial. Br. Med. J. 2003, 326, 571–574. [Google Scholar] [CrossRef] [Green Version]
- Bulloch, R.E.; Wall, C.R.; Thompson, J.M.D.; Taylor, R.S.; Poston, L.; Roberts, C.T.; Dekker, G.A.; Kenny, L.C.; Simpson, N.A.B.; Myers, J.E.; et al. Folic acid supplementation is associated with size at birth in the Screening for Pregnancy Endpoints (SCOPE) international prospective cohort study. Early Hum. Dev. 2020, 147. [Google Scholar] [CrossRef] [PubMed]
- Catena, A.; Martínez-Zaldívar, C.; Diaz-Piedra, C.; Torres-Espínola, F.J.; Brandi, P.; Pérez-García, M.; Decsi, T.; Koletzko, B.; Campoy, C. On the relationship between head circumference, brain size, prenatal long-chain PUFA/5-methyltetrahydrofolate supplementation and cognitive abilities during childhood. Br. J. Nutr. 2019, 122, S40. [Google Scholar] [CrossRef]
- Husen, S.C.; Koning, I.V.; Go, A.T.J.I.; Groenenberg, I.A.L.; Willemsen, S.P.; Rousian, M.; Steegers-Theunissen, R.P.M. IVF with or without ICSI and the impact on human embryonic brain development: The Rotterdam Periconceptional Cohort. Hum. Reprod. 2021, 36, 596–604. [Google Scholar] [CrossRef]
- Koning, I.V.; Dudink, J.; Groenenberg, I.A.L.; Willemsen, S.P.; Reiss, I.K.M.; Steegers-Theunissen, R.P.M. Prenatal cerebellar growth trajectories and the impact of periconceptional maternal and fetal factors. Hum. Reprod. 2017, 32, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Nemescu, D.; Adam, A.M.; Tanasa, I.A.; Socolov, D.; Bohiltea, R.E.; Navolan, D.B.; Zvanca, M.E. Reference ranges for the fetal mesencephalon to occiput measurement at 11 to 13+6 weeks of gestation. Exp. Ther. Med. 2020, 20, 2475–2480. [Google Scholar] [CrossRef] [PubMed]
- Hossein-Nezhad, A.; Mirzaei, K.; Maghbooli, Z.; Najmafshar, A.; Larijani, B. The influence of folic acid supplementation on maternal and fetal bone turnover. J. Bone Miner. Metab. 2011, 29, 186–192. [Google Scholar] [CrossRef]
- Lecorguillé, M.; Lioret, S.; de Lauzon-Guillain, B.; de Gavelle, E.; Forhan, A.; Mariotti, F.; Charles, M.A.; Heude, B. Association between dietary intake of one-carbon metabolism nutrients in the year before pregnancy and birth anthropometry. Nutrients 2020, 12, 838. [Google Scholar] [CrossRef] [Green Version]
- Tan, A.; Sinclair, G.; Mattman, A.; Vallance, H.D.; Lamers, Y. Maternal Vitamin B12Status in Early Pregnancy and Its Association with Birth Outcomes in Canadian Mother-Newborn Dyads. Br. J. Nutr. 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Neumann, C.G.; Oace, S.M.; Chaparro, M.P.; Herman, D.; Drorbaugh, N.; Bwibo, N.O. Low vitamin B12 intake during pregnancy and lactation and low breastmilk vitamin 12 content in rural Kenyan women consuming predominantly maize diets. Food Nutr. Bull. 2013, 34, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, M.; Funahashi, N.; Fukuoka, H.; Nammo, T.; Sato, Y.; Yoshihara, H.; Oishi, H.; Tanaka, M.; Yano, T.; Minoura, S.; et al. Effects of maternal and fetal choline concentrations on the fetal growth and placental DNA methylation of 12 target genes related to fetal growth, adipogenesis, and energy metabolism. J. Obstet. Gynaecol. Res. 2021, 47, 734–744. [Google Scholar] [CrossRef]
- Parisi, F.; Rousian, M.; Koning, I.V.; Willemsen, S.P.; de Vries, J.H.M.; Steegers, E.A.P.; Steegers-Theunissen, R.P.M. Periconceptional maternal dairy-rich dietary pattern is associated with prenatal cerebellar growth. PLoS ONE 2018, 13, e0197901. [Google Scholar] [CrossRef] [Green Version]
- Timmermans, S.; Steegers-Theunissen, R.P.; Vujkovic, M.; Den Breeijen, H.; Russcher, H.; Lindemans, J.; MacKenbach, J.; Hofman, A.; Lesaffre, E.E.; Jaddoe, V.V.; et al. The Mediterranean diet and fetal size parameters: The Generation R Study. Br. J. Nutr. 2012, 108, 1399–1409. [Google Scholar] [CrossRef] [Green Version]
- Clare, C.E.; Brassington, A.H.; Kwong, W.Y.; Sinclair, K.D. One-Carbon Metabolism: Linking Nutritional Biochemistry to Epigenetic Programming of Long-Term Development. Annu. Rev. Anim. Biosci. 2019, 7, 263–287. [Google Scholar] [CrossRef]
- Lawton, A.; Morgan, C.R.; Schreiner, C.R.; Schreiner, C.G.; Baumann, J.; Upchurch, B.; Xu, F.; Price, M.S.; Isaacs, G.D. Folate-Dependent Cognitive Impairment Associated With Specific Gene Networks in the Adult Mouse Hippocampus. Front. Nutr. 2020, 7, 574730. [Google Scholar] [CrossRef]
- Regev, O.; Cohen, G.; Hadar, A.; Schuster, J.; Flusser, H.; Michaelovski, A.; Meiri, G.; Dinstein, I.; Hershkovitch, R.; Menashe, I. Association Between Abnormal Fetal Head Growth and Autism Spectrum Disorder. J. Am. Acad. Child. Adolesc. Psychiatry 2020. [Google Scholar] [CrossRef]
- Elder, L.M.; Dawson, G.; Toth, K.; Fein, D.; Munson, J. Head circumference as an early predictor of autism symptoms in younger siblings of children with autism spectrum disorder. J. Autism Dev. Disord. 2008, 38, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, A.J.; Zubrick, S.R.; Blair, E.; Newnham, J.P.; Hickey, M. Fetal head circumference growth in children with specific language impairment. Arch. Dis. Child. 2012, 97, 49–51. [Google Scholar] [CrossRef] [Green Version]
- Lainhart, J.E.; Bigler, E.D.; Bocian, M.; Coon, H.; Dinh, E.; Dawson, G.; Deutsch, C.K.; Dunn, M.; Estes, A.; Tager-Flusberg, H.; et al. Head circumference and height in autism: A study by the Collaborative Program of Excellence in Autism. Am. J. Med. Genet. A 2006, 140, 2257–2274. [Google Scholar] [CrossRef] [Green Version]
- Bartholomeusz, H.H.; Courchesne, E.; Karns, C.M. Relationship between head circumference and brain volume in healthy normal toddlers, children, and adults. Neuropediatrics 2002, 33, 239–241. [Google Scholar] [CrossRef]
- Rousian, M.; Koster, M.P.H.; Mulders, A.; Koning, A.H.J.; Steegers-Theunissen, R.P.M.; Steegers, E.A.P. Virtual reality imaging techniques in the study of embryonic and early placental health. Placenta 2018, 64, S29–S35. [Google Scholar] [CrossRef]
- WHO. Guideline: Oprimal Serum and Red Blood Cell Folate Concentrations in Women of Reproductive Age for Prevention of Neural Tube Defects; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- de Benoist, B. Conclusions of a WHO Technical Consultation on folate and vitamin B12 deficiencies. Food Nutr. Bull. 2008, 29, S238–S244. [Google Scholar] [CrossRef]
- Hooijkaas, H.; Mohrmann, K.; Smeets, L.C.; Souverijn, J.H.M.; Taks, G.H.M. Handboek Medische Laboratoriumdiagnostiek; Prelum Uitgevers: Houten, The Netherlands, 2013. [Google Scholar]
- Head Circumference. In A.D.A.M. Medical Encyclopedia; U.S. National Library of Medicine; Posted Online 2 February 2021. Available online: https://medlineplus.gov/ency/imagepages/17206.htm (accessed on 13 July 2021).
- Friso, S.; Udali, S.; De Santis, D.; Choi, S.W. One-carbon metabolism and epigenetics. Mol. Aspects Med. 2017, 54, 28–36. [Google Scholar] [CrossRef]
- Volpe, J.J. Cerebellum of the premature infant: Rapidly developing, vulnerable, clinically important. J. Child. Neurol. 2009, 24, 1085–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolinoy, D.C.; Weidman, J.R.; Waterland, R.A.; Jirtle, R.L. Maternal genistein alters coat color and protects Avy mouse offspring from obesity by modifying the fetal epigenome. Environ. Health Perspect. 2006, 114, 567–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Ye, Y.; Li, Y.; Zhang, Y.; Zhang, Y.; Jiang, Y.; Chen, X.; Wang, L.; Yan, W. Maternal folate levels during pregnancy and children’s neuropsychological development at 2 years of age. Eur. J. Clin. Nutr. 2020, 74, 1585–1593. [Google Scholar] [CrossRef] [PubMed]
- Veena, S.R.; Krishnaveni, G.V.; Srinivasan, K.; Wills, A.K.; Muthayya, S.; Kurpad, A.V.; Yajnik, C.S.; Fall, C.H.D. Higher maternal plasma folate but not vitamin B-12 concentrations during pregnancy are associated with better cognitive function scores in 9- to 10- year-old children in South India. J. Nutr. 2010, 140, 1014–1022. [Google Scholar] [CrossRef] [Green Version]
- Tamura, T.; Goldenberg, R.L.; Chapman, V.R.; Johnston, K.E.; Ramey, S.L.; Nelson, K.G. Folate status of mothers during pregnancy and mental and psychomotor development of their children at five years of age. Pediatrics 2005, 116, 703–708. [Google Scholar] [CrossRef]
- Boeke, C.E.; Gillman, M.W.; Hughes, M.D.; Rifas-Shiman, S.L.; Villamor, E.; Oken, E. Choline intake during pregnancy and child cognition at age 7 years. Am. J. Epidemiol. 2013, 177, 1338–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, Z.; Zhu, Y.; Yan, J.; Liu, H.; Huang, G.; Li, W. Maternal folic acid impacts DNA methylation profile in male rat offspring implicated in neurodevelopment and learning/memory abilities. Genes Nutr. 2021, 16, 1. [Google Scholar] [CrossRef]
- Julvez, J.; Fortuny, J.; Mendez, M.; Torrent, M.; Ribas-Fitó, N.; Sunyer, J. Maternal use of folic acid supplements during pregnancy and four-year-old neurodevelopment in a population-based birth cohort. Paediatr. Perinat. Epidemiol. 2009, 23, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Roza, S.J.; Van Batenburg-Eddes, T.; Steegers, E.A.P.; Jaddoe, V.W.V.; MacKenbach, J.P.; Hofman, A.; Verhulst, F.C.; Tiemeier, H. Maternal folic acid supplement use in early pregnancy and child behavioural problems: The Generation R Study. Br. J. Nutr. 2010, 103, 445–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meador, K.J.; Pennell, P.B.; May, R.C.; Brown, C.A.; Baker, G.; Bromley, R.; Loring, D.W.; Cohen, M.J. Effects of periconceptional folate on cognition in children of women with epilepsy: NEAD study. Neurology 2020, 94, e729–e740. [Google Scholar] [CrossRef] [PubMed]
- Eryilmaz, H.; Dowling, K.F.; Huntington, F.C.; Rodriguez-Thompson, A.; Soare, T.W.; Beard, L.M.; Lee, H.; Blossom, J.C.; Gollub, R.L.; Susser, E.; et al. Association of prenatal exposure to population-wide folic acid fortification with altered cerebral cortex maturation in youths. JAMA Psychiatry 2018, 75, 918–928. [Google Scholar] [CrossRef]
- Caffrey, A.; McNulty, B.; Irwin, B.; Cassidy, T.; McLaughlin, M.; Lees-Murdock, D.; Walsh, C.P.; Pentieva, K. Impact of folic acid supplementation during pregnancy on cognitive performance of children at age 11 years: Preliminary results from the FASSTT Offspring study. Proc. Nutr. Soc. 2018, 77. [Google Scholar] [CrossRef] [Green Version]
- Freitas-Vilela, A.A.; Pearson, R.M.; Emmett, P.; Heron, J.; Smith, A.; Emond, A.; Hibbeln, J.R.; Castro, M.B.T.; Kac, G. Maternal dietary patterns during pregnancy and intelligence quotients in the offspring at 8 years of age: Findings from the ALSPAC cohort. Matern. Child. Nutr. 2018, 14. [Google Scholar] [CrossRef]
- Amati, F.; Hassounah, S.; Swaka, A. The Impact of Mediterranean Dietary Patterns During Pregnancy on Maternal and Offspring Health. Nutrients 2019, 11, 1098. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, K.; Thomas, T.; Kapanee, A.R.M.; Ramthal, A.; Bellinger, D.C.; Bosch, R.J.; Kurpad, A.V.; Duggan, C. Effects of maternal vitamin B12 supplementation on early infant neurocognitive outcomes: A randomized controlled clinical trial. Matern Child. Nutr. 2017, 13. [Google Scholar] [CrossRef]
- Golding, J.; Gregory, S.; Clark, R.; Iles-Caven, Y.; Ellis, G.; Taylor, C.M.; Hibbeln, J. Maternal prenatal vitamin B12 intake is associated with speech development and mathematical abilities in childhood. Nutr. Res. 2021, 86, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Bhate, V.K.; Joshi, S.M.; Ladkat, R.S.; Deshmukh, U.S.; Lubree, H.G.; Katre, P.A.; Bhat, D.S.; Rush, E.C.; Yajnik, C.S. Vitamin B12 and folate during pregnancy and offspring motor, mental and social development at 2 years of age. J. Dev. Orig. Health Dis. 2012, 3, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.S.; Ayob, M.N.M.; Cai, S.; Quah, P.L.; Gluckman, P.D.; Shek, L.P.; Yap, F.; Tan, K.H.; Chong, Y.S.; Godfrey, K.M.; et al. Maternal plasma vitamin B-12 concentrations during pregnancy and infant cognitive outcomes at 2 years of age. Br. J. Nutr. 2019, 121, 1303–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonilla, C.; Lawlor, D.A.; Taylor, A.E.; Gunnell, D.J.; Ben-Shlomo, Y.; Ness, A.R.; Timpson, N.J.; Pourcain, B.S.; Ring, S.M.; Emmett, P.M.; et al. Vitamin B-12 Status during Pregnancy and Child’s IQ at Age 8: A Mendelian Randomization Study in the Avon Longitudinal Study of Parents and Children. PLoS ONE 2012, 7, e51084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhate, V.; Deshpande, S.; Bhat, D.; Joshi, N.; Ladkat, R.; Watve, S.; Fall, C.; de Jager, C.A.; Refsum, H.; Yajnik, C. Vitamin B12 status of pregnant Indian women and cognitive function in their 9-year-old children. Food Nutr. Bull. 2008, 29, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Bottiglieri, T. Folate, vitamin B12, and S-adenosylmethionine. Psychiatr. Clin. N. Am. 2013, 36, 1–13. [Google Scholar] [CrossRef]
- Setien-Suero, E.; Suarez-Pinilla, M.; Suarez-Pinilla, P.; Crespo-Facorro, B.; Ayesa-Arriola, R. Homocysteine and cognition: A systematic review of 111 studies. Neurosci. Biobehav. Rev. 2016, 69, 280–298. [Google Scholar] [CrossRef] [Green Version]
- Blusztajn, J.K.; Mellott, T.J. Choline nutrition programs brain development via DNA and histone methylation. Cent. Nerv. Syst. Agents Med. Chem. 2012, 12, 82–94. [Google Scholar] [CrossRef]
- Signore, C.; Ueland, P.M.; Troendle, J.; Mills, J.L. Choline concentrations in human maternal and cord blood and intelligence at 5 y of age. Am. J. Clin. Nutr. 2008, 87, 896–902. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Author (Year) | Exposure (Range) | Exposure Timing | Outcome(s) | Association | Statistical Value | Quality Score |
|---|---|---|---|---|---|---|
| Bergen et al. (2016) [40] | Plasma folate (6.2–34.3 nmol/L) | Early pregnancy (median 13.5 weeks GA) | Trend for folate in low quintiles (<13.10 nnmol/L) associated with reduced foetal HC in 2nd and 3rd trimester and birth | = | Q1 (≤9.10 nmol/L) B = −2.0 Q2 (9.11–13.10) B = −1.2 Q3 (13.11–18.90) B = −0.2 Q4 (18.91–25.80) B = −1.3 P = 0.14 | 8 |
| Steenweg-de Graaff et al. (2017) [34] | Plasma folate (1.8–45.3 nmol/L) | Early pregnancy (13.2 weeks GA) | Higher folate associated with larger foetal HC 30 weeks GA but not at 20 weeks and at birthHigher folate associated with increased foetal head growth between 2nd and 3rd trimester | + | B = 0.47, P ≤ 0.001 f B = 0.004, P = 0.02 g | 8 |
| Nilsen et al. (2010) [37] | Plasma folate (<5.9->14.8 nmol/L) | Mid-pregnancy (median 18 weeks GA) | No linear association with neonatal HC at birth | = | P = 0.53 | 7 |
| Zou et al. (2020) [35] | Plasma folate (1.8–45.3 nmol/L) | Early pregnancy (mean 13.3 weeks GA) | Folate deficiency associated with reduced foetal brain volume from 3rd trimester to childhood | − | B = −0.04, P = 0.02 | 7 |
| Koning et al. (2015) [36] | RBC folate (814–2936 nmol/L) | 1st trimester (≤ 8+0 weeks GA) | Highest foetal proportional cerebellar growth and TCD, RCD, and LCD was highest in the third quartile of RBC folate (1538–1813 nmol/L) in 1st trimester | + | Q1 (−0.0721 mm/day) P < 0.01 c Q2 (−0.0438) P < 0.01 c Q4 (−0.0459) P = 0.05 c Q1 (−0.0364 mm/mm) P < 0.05 d Q2 (−0.0280) P > 0.05 d Q4 (−0.0232) P > 0.05 d Q1 (−0.0017 mm/day) P >0.05 e Q2 (−0.0010) P > 0.05 e Q4 (−0.0459) P < 0.05 e | 6 |
| Brough et al. (2010) [32] | RBC folate (Not reported) | Early pregnancy (>13 weeks GA) | Positively correlated with neonatal HC at birth | + | R = 0.111, P = 0.046 | 5 |
| Schlotz et al. (2010) [33] | RBC folate (373.5–588.5 µg/L) | Median 95 days GA | Association with neonatal HC at birth | + | Β = 0.17, P = 0.031 | 5 |
| Gadgil et al. (2014) [41] | Plasma folate (15.1–18.95 ng/mL) | Late pregnancy (36 weeks GA) | No correlation with neonatal HC at birth Higher folate-to-vitamin B12 ratio correlated with decreased neonatal HC at birth | = (- folate-to-vitamin B12) | R = −0.089, P = 0.672 a R = −0.469, P = 0.018 b | 4 |
| Jiang et al. (2016) [39] | Serum folate (Not reported) | Not reported | Positively correlated with neonatal HC at birth | + | R = 0.394, P = 0.044 | 3 |
| Takimoto et al. (2007) [38] | Serum and RBC folate (Not reported) | 1st, 2nd, and 3rd trimester | No associations with neonatal HC at birth apart from RBC folate in the 2nd trimester | = (+ RBC folate 2nd trimester) | Serum folate: ES = 0.08 P = 0.09 h ES = −0.03 P = 0.07 i ES = 0.004 P = 0.57 j RBC folate: ES = −0.0003 P = 0.56 h ES = 0.0005 P = 0.01 i ES = −0.0003 P = 0.90 j | 3 |
| Author (Year) | Exposure | Exposure Timing | Outcome(s) | Association | Statistical Value | Quality Score |
|---|---|---|---|---|---|---|
| Dietary intake | ||||||
| Nilsen et al. (2010) [37] | Daily mean intake of dietary folate from FFQs | Throughout pregnancy | No association with neonatal HC at birth | = | P = 0.27 | 7 |
| Schlotz et al. (2010) [33] | Dietary folate intake based on FFQs | Early (median gestational day 101) and late pregnancy (median gestational day 199) | Trend for association with neonatal HC at birth | = | B = 0.15, P = 0.083 | 5 |
| Takimoto et al. (2007) [38] | Daily intake of dietary folate calculated from the Standard Food Consumption Table | Prenatal | No relation with neonatal HC at birth | = | Not reported | 3 |
| Supplement use | ||||||
| Steenweg-de-Graaff et al. (2017) [34] | Folic acid supplement use via self-administered questionnaires | Early pregnancy (median 13.2 weeks GA) | Preconceptional supplement use slightly increased foetal head size and circumference at 20 weeks of gestation No association with foetal head growth | + (= foetal head growth) | B = 0.112, P = 0.01 g B = 0.120, P = 0.01 h | 8 |
| Timmermans et al. (2009) [43] | Folic acid supplement use via self-administered questionnaires | Mid-pregnancy (median 15.4 weeks GA) | Periconceptional supplement use was associated with trends towards larger foetal HC at 20 and 30 weeks of pregnancy | + | B = 0.61, P > 0.05 i B = 1.34, P > 0.05 j | 8 |
| Yusuf et al. (2019) [44] | 0.8 mg (control) or 4 mg (high dose, intervention) folic acid supplement per day | 1st trimester (mean 12.3 weeks GA) | Higher dose users had a 1.88 mm larger neonatal HC at birth Higher dose had no effect on neonatal brain weight at birth | + (= brain weight at birth) | P = 0.28 h 6.90 ± 5.85 g, P = 0.24 k | 8 |
| Christian et al. (2003) [45] | Folic acid (400 µg/day), folic acid-iron (60 mg ferrous fumarate), folic acid-iron-zinc (30 mg zinc sulphate) | Preconception until birth | Folic acid-iron supplement use increased neonatal HC at birth by 0.16 cm | + | B = 0.16, P = 0.012 | 7 |
| Nilsen et al. (2010) [37] | Folic acid supplement use via self-administered questionnaires | From start to mid-pregnancy (median 18 weeks GA) | No association with neonatal HC at birth | = | P = 0.44 | 7 |
| Bulloch et al. (2020) [46] | Folic acid supplement use via lifestyle questionnaires | Preconception and at 15 + 1 weeks GA | No association with neonatal HC z-score at birth | = | B = 0.04, P = 0.197 a B = 0.01, P = 0.662 b | 6 |
| Catena et al. (2019) [47] | 5-m-THF supplement use | From 22 weeks GA to delivery | No effect on neonatal HC at birth | P > 0.13 | 6 | |
| Husen et al. (2021) [48] | Folic acid supplement use via self-reported questionnaires | Early pregnancy (<10 weeks GA) | Preconceptional initiation was not associated with either 9 or 11 weeks foetal DTD, MTD, TTL, or TTR measurements | = | DTD: B = 0.093, P = 0.422 c B = −0.068, P = 0.609 d MTD: B = −0.025, P = 0.814 c B = −0.004, P = 0.968 d TTL: B = 0.047, P = 0.141 c B = 0.027, P = 0.570 d TTR: B = 0.028, P = 0.531 c B = 0.025, P = 0.599 d | 6 |
| Koning et al. (2015) [36] | Folic acid supplement use via self-administered questionnaires | 1st trimester (≤8 + 0 weeks GA) | Preconceptional supplement use increased proportional foetal cerebellar growth for TCD, RCD, and LCD in 1st trimester | + | TCD: B = 0.257, P = 0.032 e B = −0.078, P = 0.616 f RCD: B = 0.156, P = 0.015 e B = 0.008, P = 0.918 f LCD: B = 0.171, P = 0.013 e B = 0.041, P = 0.601 f | 6 |
| Koning et al. (2017) [49] | Folic acid supplement use via self-administered questionnaires | 1st trimester (≤12 weeks GA) | No associations with foetal cerebellum growth trajectories in the 1st trimester | = | B = 0.22, P = 0.19 | 5 |
| Nemescu et al. (2020) [50] | Folic acid supplement use | 1st trimester (11–13 weeks GA) | No supplement use increased foetal MO in the 1st trimester | − | P = 0.014 | 5 |
| Hossein-nezhad et al. (2011) [51] | Group 1: 1 mg/day folic acid in 1st and 2nd trimesters and Group 2: 1 mg/day folic acid until birth | Throughout pregnancy | No effect of timing of supplement use on neonatal HC at birth | = | P = 0.5 | 4 |
| Takimoto et al. (2011) [31] | Folic acid supplement use via 24 h dietary recall survey | Throughout pregnancy | Decreased HC at birth in female neonates | − | B = 0.112, P = 0.01 | 4 |
| Takimoto et al. (2007) [38] | Folic acid use from self-administered questionnaires | 1st, 2nd, and 3rd trimesters | Not related to neonatal HC at birth | = | Not reported | 3 |
| Author (Year) | Exposure (Range) | Exposure Timing | Outcome(s) | Association | Statistical Value | Quality Score |
|---|---|---|---|---|---|---|
| Serum/plasma | ||||||
| Bergen et al. (2016) [40] | Non-fasting serum total and active vitamin B12 (83–315 and 20–83 pmol/L) | Early pregnancy (median 13.5 GA) | No associations with neonatal HC at birth | = | Q1 (≤119.0 pmol/L) B = −0.6 Q2 (119.0–153.0) B = −0.4 Q3 (153.01–189.0) B = 0.5 Q4 (189.01–244.0) B = −0.3, P = 0.28 a Q1 (≤30 pmol/L) B = −0.7 Q2 (31–38) B = 0.1 Q3 (39–46) B = −0.4 Q4 (47–59) B = 0.4, P = 0.30 b | 8 |
| Tan et al. (2021) [52] | Non-fasting serum vitamin B12 (147–297 pmol/L) | 1st and 2nd trimester | No linear association with neonatal HC z-score at birth | = | B = −0.00278, P = 0.52 c B = −0.00715, P = 0.10 d | 6 |
| Gadgil et al. (2014) [41] | Plasma vitamin B12 (138.6–261.4 pg/mL) | Late pregnancy (mean 36 weeks GA) | No correlation with neonatal HC at birth | = | R = 0.22, P = 0.28 | 4 |
| Jiang et al. (2016) [39] | Serum vitamin B12 (Not reported) | Not reported | Positively correlated with neonatal HC at birth | + | R = 0.511, P = 0.029 | 3 |
| Takimoto et al. (2007) [38] | Non-fasting serum vitamin B12 (Not reported) | 1st, 2nd, and 3rd trimesters | Association with neonatal HC at birth but effect size too small for physiological significance | = | Not reported | 3 |
| Dietary intake | ||||||
| Neumann et al. (2013) [53] | Dietary vitamin B12 intake from quantitative food weighing and dietary recall | From 1st/2nd trimester to term | No associations with neonatal HC at birth | = | Not reported | 3 |
| Takimoto et al. (2007) [38] | Daily dietary vitamin B12 intake | 1st, 2nd, and 3rd trimesters | Not related to neonatal HC at birth | = | ES = −0.003, P = 0.20 c ES = 0.02, P = 0.08 d ES = 0.01, P = 0.10 e | 3 |
| Author (Year) | Exposure (Range) | Exposure Timing | Outcome(s) | Association | Statistical Value | Quality Score |
|---|---|---|---|---|---|---|
| Bergen et al. (2016) [40] | Plasma tHcy (4.9–10.5 µmol/L) | Early pregnancy (median 13.5 weeks GA) | ≥8.31 µmol/L tHcy associated with reduced foetal HC (−1.6 mm) from late pregnancy (median 30.4 weeks GA) to birth | = | Β = −1.6, P = 0.06 | 8 |
| Nilsen et al. (2010) [37] | Plasma tHcy (<4.5->5.8 µmol/L) | Mid-pregnancy (median 18 weeks GA) | No linear association with neonatal HC at birth | = | P = 0.48 | 7 |
| Tan et al. (2021) [52] | Non-fasting serum tHcy (4.5–5.7 µmol/L) | 1st and 2nd trimester | No linear association with neonatal HC z-score at birth | = | B = 0.0202, P = 0.65 a B = 0.0762, P = 0.06 b | 6 |
| Takimoto et al. (2011) [31] | Non-fasting plasma tHcy (Not reported) | 3rd trimester | Positively correlated with neonatal HC at birth | + | R = 0.53, P < 0.01 | 4 |
| Jiang et al. (2016) [39] | Serum tHcy (Not reported) | Not reported | Negative correlation with neonatal HC at birth | − | R = −0.401, P = 0.034 | 3 |
| Takimoto et al. (2007) [38] | Non-fasting plasma tHcy (Not reported) | 1st, 2nd, and 3rd trimesters | No relation with neonatal HC at birth except for the second trimester | = (+ second trimester tHcy) | ES = 0.02, P = 0.92 a ES = 1.54, P = 0.03 b ES = −0.24, P = 0.53 c | 3 |
| Author (Year) | Exposure | Exposure Timing | Association with One-Carbon Metabolism | Outcome(s) | Association | Statistical Value | Quality Score |
|---|---|---|---|---|---|---|---|
| Timmermans et al. (2012) [57] | Mediterranean diet | Early pregnancy (median 13.5 weeks GA) | Low adherence was associated with high tHcy, low serum vitamin B12, and folate | Low adherence associated with a smaller foetal HC in late pregnancy | + | Difference in SDS = −0.08, P= 0.01 | 7 |
| Lecorguillé et al. (2020) [52] | Varied and balanced diet | 2nd trimester (average 15 weeks GA) | High positive coefficients for B vitamins, choline, and methionine | Not associated with neonatal HC at birth | = | B = 0.01, P = 0.43 | 6 |
| Parisi et al. (2018) [56] | Dairy-rich diet | 1st trimester (≤8 + 0 weeks GA) | Associated with lower plasma tHcy and correlated to vitamin B2, B6, and B12 | Associated with increased foetal TCD measurements in 1st and 3rd trimester | + | B = 0.02, P < 0.01 | 6 |
| Reference Values | References | ||
|---|---|---|---|
| General Population | Pregnancy | ||
| Serum/plasma folate (nmol/L) | 13.5–45.3 | >10 * | [42] |
| RBC folate (nmol/L) | >340 ** | >906 | [42,66] |
| Serum/plasma vitamin B12 (pmol/L) | 130–700 | >150 *** | [67,68] |
| Homocysteine (µmol/L) | <15 | The lower, the most optimal for pregnancy health | [68] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubini, E.; Baijens, I.M.M.; Horánszky, A.; Schoenmakers, S.; Sinclair, K.D.; Zana, M.; Dinnyés, A.; Steegers-Theunissen, R.P.M.; Rousian, M. Maternal One-Carbon Metabolism during the Periconceptional Period and Human Foetal Brain Growth: A Systematic Review. Genes 2021, 12, 1634. https://doi.org/10.3390/genes12101634
Rubini E, Baijens IMM, Horánszky A, Schoenmakers S, Sinclair KD, Zana M, Dinnyés A, Steegers-Theunissen RPM, Rousian M. Maternal One-Carbon Metabolism during the Periconceptional Period and Human Foetal Brain Growth: A Systematic Review. Genes. 2021; 12(10):1634. https://doi.org/10.3390/genes12101634
Chicago/Turabian StyleRubini, Eleonora, Inge M. M. Baijens, Alex Horánszky, Sam Schoenmakers, Kevin D. Sinclair, Melinda Zana, András Dinnyés, Régine P. M. Steegers-Theunissen, and Melek Rousian. 2021. "Maternal One-Carbon Metabolism during the Periconceptional Period and Human Foetal Brain Growth: A Systematic Review" Genes 12, no. 10: 1634. https://doi.org/10.3390/genes12101634
APA StyleRubini, E., Baijens, I. M. M., Horánszky, A., Schoenmakers, S., Sinclair, K. D., Zana, M., Dinnyés, A., Steegers-Theunissen, R. P. M., & Rousian, M. (2021). Maternal One-Carbon Metabolism during the Periconceptional Period and Human Foetal Brain Growth: A Systematic Review. Genes, 12(10), 1634. https://doi.org/10.3390/genes12101634