Abstract
Telomeres are nucleoprotein complexes protecting chromosome ends in most eukaryotic organisms. In addition to chromosome ends, telomeric-like motifs can be accumulated in centromeric, pericentromeric and intermediate (i.e., between centromeres and telomeres) positions as so-called interstitial telomeric repeats (ITRs). We mapped the distribution of (TTAGGG)n repeats in the karyotypes of 30 species from nine families of turtles using fluorescence in situ hybridization. All examined species showed the expected terminal topology of telomeric motifs at the edges of chromosomes. We detected ITRs in only five species from three families. Combining our and literature data, we inferred seven independent origins of ITRs among turtles. ITRs occurred in turtles in centromeric positions, often in several chromosomal pairs, in a given species. Their distribution does not correspond directly to interchromosomal rearrangements. Our findings support that centromeres and non-recombining parts of sex chromosomes are very dynamic genomic regions, even in turtles, a group generally thought to be slowly evolving. However, in contrast to squamate reptiles (lizards and snakes), where ITRs were found in more than half of the examined species, and birds, the presence of ITRs is generally rare in turtles, which agrees with the expected low rates of chromosomal rearrangements and rather slow karyotype evolution in this group.
1. Introduction
Telomeres are regions of repetitive DNA motifs and associated proteins localized at the edges of chromosomes. They play a crucial role in maintaining the chromosome structure as a single unit, preventing fusions of free “sticky” chromosome ends, or degeneration and loss of genetic information during the replication events [1]. In vertebrates, telomeric regions consist of long tandem repeats of the TTAGGG motif, which was first characterized in humans [2] and seems to be extremely conserved across all studied vertebrates [3]. Telomeres are formed and preserved by telomerase, a reverse transcriptase able to add new TTAGGG repeats based on an RNA template and to compensate for the normal shortening of telomeres after each replication event [4,5,6,7]. However, the presence and, therefore, activity of telomerase varies across cell types and developmental stages [8]. Notably, the shortening of the telomeres is associated with cell aging and cell death [9,10,11,12]. Interestingly, (TTAGGG)n motifs can be found also in non-terminal positions of the chromosomes [3,13,14] in the form of interstitial telomeric repeats (ITRs) also referred to as interstitial telomeric sequences (ITSs). Some authors classified ITRs into two main groups: the short ITRs (s-ITRs) and the heterochromatic ITRs (Het-ITRs) [15]. Other authors report a more detailed classification including s-ITRs, subtelomeric ITRs, fusion ITRs and pericentromeric ITRs [14,15,16,17,18]. s-ITRs are characterized by a low copy number of (TTAGGG)n tandem repeats, located in interstitial positions of the chromosomes. The bloom of genome sequencing projects revealed that s-ITRs are common in vertebrate genomes [19,20,21,22,23,24,25,26]. They are often not detectable by molecular cytogenetic methods, for example by fluorescence in situ hybridization (FISH), due to the low number of repeats [15]. In contrast, Het-ITRs consist of an extensive accumulation of (TTAGGG)n tandem repeats mainly located in heterochromatic regions, particularly in centromeric and pericentromeric areas, but occasionally also interspersed within chromosome arms. Het-ITRs are less common than s-ITRs in vertebrate genomes and are often detectable by molecular cytogenetic methods [15,27,28,29,30]. In our study, we classify ITRs into three categories: centromeric, pericentromeric and intermediate ITRs, according to their position following our previous classification for squamate reptiles [28].
The origin and function of ITRs still remain unclear. According to the most prevalent hypothesis, ITRs are remnants of former terminal telomeric sequences that have been repositioned to interstitial position as a consequence of chromosomal fusions [3], fissions and inversions. Especially Het-ITRs are linked to chromosomal rearrangements [15], and their presence in centromeric and pericentromeric regions are often taken as a marker of former chromosomal fusions [22,31,32]. Alternatively, ITRs can emerge in hotspots for DNA breakage and recombination [14,17], produced by the cell repair machinery after unequal crossing over and double-strand breaks [33,34,35,36]. This last hypothesis explains the origin of s-ITRs, and it is supported by the observation that telomerase is able to insert telomeric repeats into double strand break sites during chromosome healing [37,38]. Notably, long regions of ITRs can also emerge via amplification of pre-existing short (TTAGGG)n repeats present in the genomes as latent telomeres [3].
Cytogenetic studies have detected ITRs in numerous species across vertebrate lineages, including mammals, fishes, birds, non-avian reptiles and amphibians [3,27,28,29,30,39,40,41,42,43,44,45,46]. In non-avian reptiles, distribution of telomeric sequences have been extensively studied in squamates, i.e., lizards and snakes, where ITRs were detected in approximately 100 species, despite the generally conserved chromosome morphology in this group [28,45,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64]. It was proposed that intrachromosomal rearrangements might have a crucial role in the formation of ITRs in squamate reptiles [28,63]. A telomere-only pattern was instead reported in Crocodylus siamensis [65], and as far as we know, no other representative of Crocodylia has been studied. In birds, ITRs are relatively common and mostly found in centromeric and pericentromeric positions [39,44,66,67]. Notably, in some cases, the telomeric motif TTAGGG is part of a larger satellite DNA sequence, and its presence and amplification in the genome is not directly connected to chromosomal rearrangements [68,69]. In turtles, the distribution of telomeric motifs have been studied up to our knowledge in 27 out of the 353 extant species [70,71], but ITRs have been previously reported in only six species [72,73,74,75,76,77,78,79,80,81,82], reviewed further in this study, but more systematic survey is needed before drawing solid conclusions on the frequency of ITRs in turtles.
Turtles have diploid chromosome numbers ranging from 2n = 26 to 2n = 68 [76,83]; however, diploid numbers and chromosome morphology seem to be conserved and rather stable among species within a family [76,84,85]. Phylogenetic reconstruction suggested that the ancestral turtle karyotype was composed of 2n = 52 chromosomes [76]. The variability in chromosome numbers across turtles is driven mainly by fusions involving microchromosomes leading to reduction of diploid chromosome numbers (e.g., in the ancestor of the families Pelomedusidae and Podocnemididae), and by chromosome fissions leading to increase of diploid chromosome numbers (e.g., in the ancestor of the families Carettochelyidae and Trionychidae) [76].
In this study, we explored the distribution of telomeric repeats from representative species across the turtle phylogeny by analyzing 30 species from nine families using fluorescence in situ hybridization with a probe specific for the telomeric repeats. We combine our results with previously published data, and present an overview of the distribution of ITRs across turtles and compare their frequency to other reptile lineages.
2. Materials and Methods
2.1. Sampling
We studied the distribution of telomeric (TTAGGG)n repeats in the karyotypes of 30 turtle species representing nine families (Table 1). Blood samples were collected from captive-bred or legally imported turtles from the Münster Zoo (Germany), Plzeň Zoo, Prague Zoo (both Czech Republic), the Museum of Zoology, Senckenberg Dresden (Germany), Turtle Island (Austria) and private keepers. Experiments with animals were performed by accredited researchers (LK: CZ02535, MR: CZ03540). When needed, turtles were temporarily kept in the animal facility of the Faculty of Science, Charles University (accreditation No. 37428/2019-MZE-18134). The species identity of studied turtles was verified in morphologically challenging taxa using mtDNA sequences that were compared to the sequences published in the revisionary works by Fritz et al. [86] for the genus Cyclemys, Petzold et al. [87] for Pelomedusa, and Ihlow et al. [88] for Malayemys. The Galápagos giant tortoises used in the present study from the Prague Zoo are of known provenance and species identity. All experimental procedures were carried out under the supervision and with the approval of the Ethics Committee of the Faculty of Science, Charles University, approved by the Committee for Animal Welfare of the Ministry of Agriculture of the Czech Republic (permit No. MSMT-34426/2019-7).

Table 1.
Summary of species analyzed in this study.
2.2. Chromosome Preparation and Staining
Chromosome suspensions were prepared from whole blood cell cultures. Briefly, the cultivation medium consisted of 90 mL of D-MEM medium (Sigma-Aldrich, St. Louis, MO, USA), enriched with 10 mL of fetal bovine serum (GIBCO, Carlsbad, CA, USA), 3 mL of phytohemagglutinin M (GIBCO, Carlsbad, CA, USA), 1 mL of penicillin/streptomycin solution (10,000 units/mL; GIBCO, Carlsbad, CA, USA), 1 mL L-glutamine solution (200 mM; Sigma-Aldrich, St. Louis, MO, USA) and 1 mL lipopolysaccharide solution (10 mg/mL; Sigma-Aldrich, St. Louis, MO, USA). Then, 100–300 μL of blood samples were added to the 5 mL of cultivation medium and incubated for one week at 30 °C, without CO2 supplementation. After the incubation period, the cultures were first treated with 35 μL of colcemid (10 μg/mL; Roche, Basel, Switzerland) and incubated for 3 h and 30 min at 30 °C. The cells were treated with a pre-warmed 0.075 M KCl hypotonic solution for 30 min at 37 °C, washed and fixed four times with cold 3:1 methanol/acetic acid solution. The chromosome suspensions were later stored at −20 °C for further use.
2.3. Giemsa Staining and Karyotype Reconstruction
Metaphases from species with undescribed karyotypes were stained with 8% Giemsa solution. Selected metaphases were captured using a Zeiss Axio Imager Z2 (Zeiss, Oberkochen, Germany), equipped with a Metafer-MSearch automatic scanning platform (MetaSystems, Altlussheim, Germany) and CoolCube 1 b/w digital camera (MetaSystems, Altlussheim, Germany). Karyograms were prepared using the Ikaros karyotyping platform (MetaSystems, Altlussheim, Germany). At least 10 metaphases per individual were studied.
2.4. Fluorescence In Situ Hybridization with Probes for Telomeric Repeats
The (TTAGGG)n probe for telomeric sequences was prepared and labelled with dUTP-biotin by PCR, using the (TTAGGG)5 and (CCCTAA)5 primers, without DNA template following the protocol of Ijdo et al. [89] and Rovatsos et al. [27]. The probe was diluted in a hybridization buffer (50% formamide in 2 × SSC, pH 7) and stored in the freezer for further use.
The slides with chromosome spreads were aged either overnight at 37 °C or for 1 h at 60 °C. The FISH experiment was conducted in two days. During the first day, the chromosome spreads were treated with RNase (100 μg/mL) for 1 h at 37 °C, 0.01% pepsin for 10 min at 37 °C, post-fixed in 1% formaldehyde solution for 10 min and dehydrated in a 70–85–95% ethanol series. Once dried, the slides were denatured in 70% formamide for 4 min at 75 °C and dehydrated once more with the ethanol series. Meanwhile, the probe was denatured for 6 min at 73 °C and kept in ice for at least 10 min. To each slide, 10 μL of probe was added, covered with a coverslide and incubated overnight at 37 °C. During the second day, the slides were washed 3 times with 50% formamide/2 × SSC solution for 5 min at 37 °C, two times with 2 × SSC for 5 min and once with 4 × SSC/0.05% Tween 20 (Sigma-Aldrich, St. Louis, MO, USA) for 5 min. The slides were incubated in 4 × SSC/5% blocking reagent (Roche) for 45 min at 37 °C and then in 4 × SSC/5% blocking reagent containing avidin-FITC (Vector laboratories, Burlingame, CA, USA) for 30 min at 37 °C. The fluorescence signal was twice amplified by the fluorescein–avidin D/biotinylated anti-avidin system (Vector Laboratories, Burlingame, CA, USA). After this treatment, the slides were dehydrated in ethanol series, air-dried and stained with Fluoroshield with DAPI (Sigma-Aldrich, St. Louis, MO, USA).
For each specimen, at least 20 images were obtained using a Provis AX70 (Olympus, Tokyo, Japan) fluorescence microscope equipped with a DP30BW digital camera (Olympus, Tokyo, Japan). The pictures were obtained in BW and later superimposed in color with DP Manager imaging software (Olympus, Tokyo, Japan).
2.5. Distribution of ITRs across the Turtle Phylogeny
Data for the topology on the karyotype of telomeric sequences from the species studied here was supplemented by previously published records in order to reconstruct the phylogenetic history of the presence/absence of ITRs in turtles (Figure 1). The phylogenetic trees by Pereira et al. [90] and Kehlmaier et al. [91] were used for this phylogenetic reconstruction in the software Mesquite v3.61 [92].
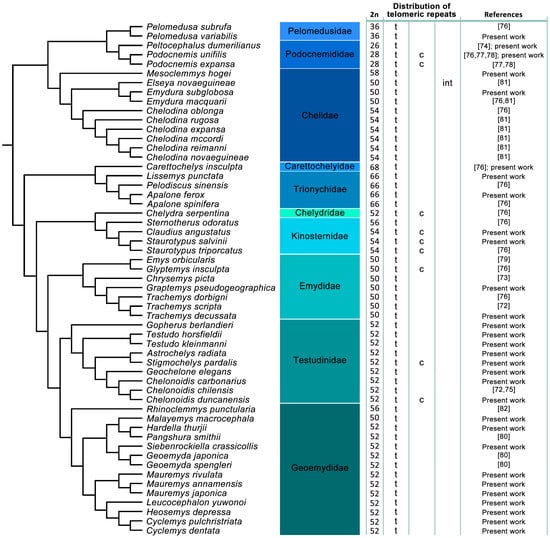
Figure 1.
Distribution of the presence of telomeric repeats across turtles. Telomeric repeats were detected in terminal topology (t) in all studied species of turtles, and additionally, interstitial telomeric repeats (ITRs) were identified in centromeric (c) (9 species) and intermediate (int) (1 species) positions. ITRs identification follows our previous classification for squamate reptiles by Rovatsos et al. [28]. Phylogeny follows Pereira et al. [90] and Kehlmaier et al. [91].
3. Results
3.1. Karyotype Description
We analyzed 30 species with karyotypes ranging from 2n = 26 in Peltocephalus dumerilianus to 2n = 66 in the softshell turtles Apalone ferox and Lissemys punctata. To the best of our knowledge, the karyotypes of the following 12 species are presented here for the first time (Figure 2): Trachemys decussata 2n = 50 (Emydidae), Cyclemys pulchristriata (2n = 52), Hardella thurjii (2n = 52), Heosemys depressa (2n = 52), Leucocephalon yuwonoi (2n = 52), Mauremys annamensis (2n = 52) (all Geoemydidae), Pelomedusa variabilis (2n = 36) (Pelomedusidae), Astrochelys radiata (2n = 52), Chelonoidis duncanensis (2n = 52), Geochelone elegans (2n = 52), Testudo horsfieldii (2n = 52) and Testudo kleinmanni (2n = 52) (all Testudinidae).
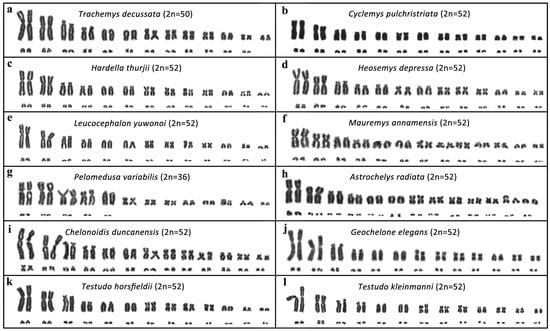
Figure 2.
Karyograms of (a) Trachemys decussata (Emydidae), (b) Cyclemys pulchristriata, (c) Hardella thurjii, (d) Heosemys depressa, (e) Leucocephalon yuwonoi, (f) Mauremys annamensis (all Geoemydidae), (g) Pelomedusa variabilis (Pelomedusidae), (h) Astrochelys radiata, (i) Chelonoidis duncanensis, (j) Geochelone elegans, (k) Testudo horsfieldii, (l) Testudo kleinmanni (all Testudinidae).
3.2. Presence of ITRs
We analyzed the topology of TTAGGG repeats across a wide range of species, covering nine out of the 14 extant families of turtles. All examined species possessed multiple TTAGGG repeats in the expected terminal telomeric positions (Figure 3 and Figure 4). The telomeric signals looked brighter on microchromosomes than on macrochromosomes in almost all species.
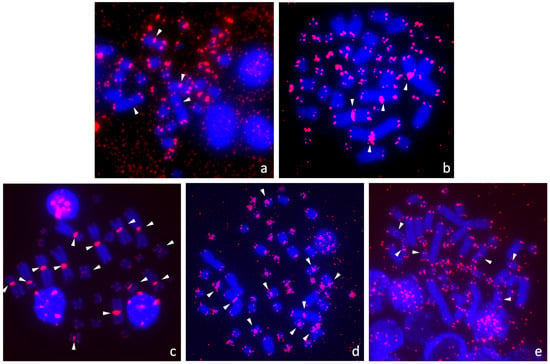
Figure 3.
Interstitial telomeric repeats (ITRs) were detected in (a) Claudius angustatus, (b) Staurotypus salvinii (both Kinosternidae), (c) Podocnemis unifilis (Podocnemididae), (d) Chelonoidis duncanensis, (e) Stigmochelys pardalis (both Testudinidae). White arrows indicate ITRs.
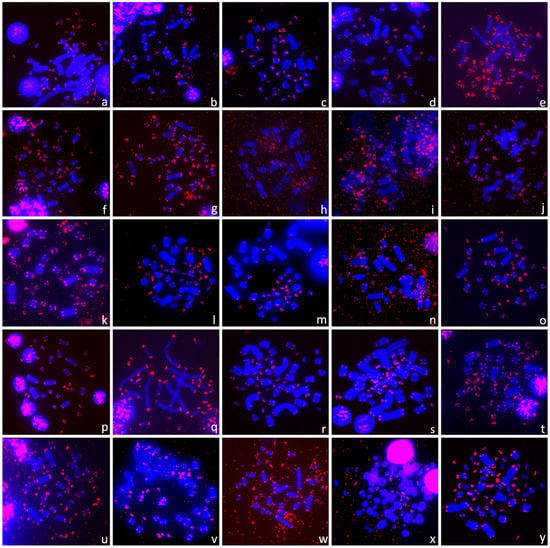
Figure 4.
The telomeric (TTAGGG)n repeats were detected only in terminal positions in (a) Carettochelys insculpta (Carettochelyidae), (b) Emydura subglobosa, (c) Mesoclemmys hogei (both Chelidae), (d) Graptemys pseudogeographica, (e) Trachemys decussata (both Emydidae), (f) Cyclemys dentata, (g) Cyclemys pulchristriata, (h) Hardella thurjii, (i) Heosemys depressa, (j) Leucocephalon yuwonoi, (k) Malayemys macrocephala, (l) Mauremys annamensis, (m) Mauremys japonica, (n) Mauremys rivulata, (o) Siebenrockiella crassicollis (all Geoemydidae), (p) Pelomedusa variabilis (Pelomedusidae), (q) Peltocephalus dumerilianus (Podocnemididae), (r) Astrochelys radiata, (s) Chelonoidis carbonarius, (t) Geochelone elegans, (u) Gopherus berlandieri, (v) Testudo horsfieldii, (w) Testudo kleinmanni (all Testudinidae), (x) Apalone ferox, (y) Lissemys punctata (both Trionychidae).
We detected ITRs in only five out of the 30 examined species and they were located exclusively in the centromeric positions (Figure 3). ITRs were detected at the centromeres of chromosomes 1 and 2 in Claudius angustatus and Staurotypus salvinii (Figure 3a,b). In Podocnemis unifilis, ITRs accumulated at the centromeres of chromosomes from the seven largest pairs (Figure 3c), confirming reports from previous studies [76,78]. In Chelonoidis duncanensis, ITRs were detected in the centromeric regions of chromosome pairs 1, 4, 8 and 9 (Figure 3d). In Stigmochelys pardalis, ITRs were present in the centromeres of chromosome pairs 6, 8 and 9 (Figure 3e).
3.3. Distribution of ITRs across the Turtle Phylogeny
Pooling together our results with data from previous studies, ITRs were identified in 10 out of 55 examined turtle species. The phylogenetic pattern of the topology of TTAGGG motifs across turtle genomes suggested that ITRs evolved independently at least seven times (Figure 1).
4. Discussion
All examined turtle species possessed multiple telomeric repeats in terminal positions. We detected ITRs in five species, increasing the total number of known turtle species with ITRs to 10. ITRs seem to be restricted to the centromeres, except for the Y chromosome of Elseya novaeguineae, where telomeric-like sequences were amplified in the intermediate position of the p-arm (Figure 1). Interestingly, as in birds [93,94,95,96,97] and squamate reptiles [28,51,98], in almost all examined turtle species the telomeric signals look brighter on microchromosomes than on macrochromosomes. This stronger signal brightness has been previously reported in other vertebrate lineages, including birds [39] and squamates [28,48,98], and it can be caused by a higher number of telomeric repeats on microchromosomes [94]. The species analyzed in the present study cover a large part of turtle phylogenetic diversity from both suborders (Pleurodira and Cryptodira) and nearly the whole variability in the diploid chromosome numbers known in turtles. Despite this variability, only 18.2% of the studied turtle species have ITRs detectable with molecular cytogenetic methods (Figure 1). Phylogenetic distribution suggests that ITRs evolved independently within turtles at least seven times.
Kinosternidae are reported to have undergone minimal chromosomal changes [99,100], where the main difference between the subfamilies Staurotypinae (2n = 54) and Kinosterninae (2n = 56) is found in a pair of microchromosomes. Centromeric ITRs have been described for the two species of the genus Staurotypus and their sister taxon Claudius angustatus (Figure 3a,b and Figure 1). The only other species from the family Kinosternidae analyzed up to now, Sternotherus odoratus (2n = 56), does not have any ITRs (Figure 1). It seems that the centromeric ITRs are an apomorphy of the subfamily Staurotypinae and cannot be directly connected to chromosome rearrangements leading to formation of the derived karyotype of the family Kinosternidae or its subfamily Staurotypinae.
A burst of ITRs emerged in Chelydra serpentina (Chelydridae), where they are present in seven chromosome pairs, although this species possesses with 2n = 52 the putatively ancestral diploid chromosome number for turtles [76] (Figure 1).
The presence of ITRs in centromeres in Podocnemis unifilis (Figure 3c) [76,77,78] and P. expansa [78] was suggested to be a remnant of ancestral fusions leading to the lower diploid chromosome number in the family Podocnemididae [76,77,78]. Nevertheless, the situation is more complicated. In P. expansa, Noronha et al. [77] detected telomeric sequences only in the terminal positions, while Cavalcante et al. [78] reported ITRs with a similar pattern as in the closely related P. unifilis. ITRs can contain a different number of telomeric-like repeats, which might be reflected in the intensity of the probe signal, and there might be interindividual variability in the number of repeats. Nevertheless, such a strong difference within the same species is surprising. None of the two studies reported a genetic identification of the examined specimens (e.g., mtDNA sequence per specimen), leaving open the possibility that perhaps due to taxonomic misidentification, different taxa were examined in each study. Regardless of this questionable report, the lack of ITRs in Peltocephalus dumerilianus, another member of family Podocnemididae, suggests that the presence of ITRs is not directly connected to fusion events. Supporting this view, ITRs have not been found in Pelomedusa variabilis (Figure 1), as well as in Pelomedusa subrufa [76] (Figure 1), even though it is possible that a different taxon was studied because formerly all ten currently recognized species [87,101] were lumped together as Pelomedusa subrufa. In any case, the genus Pelomedusa belongs to the family Pelomedusidae, the sister taxon of Podocnemididae. Pelomedusids possess a similar or even lower diploid number compared to podocnemidids, and the reduction in chromosome number can thus be a synapomorphy of Pelomedusidae and Podocnemididae. If in the common ancestor of these two families the fusions responsible for the reduction of the chromosome number would have led to ITRs in the centromeric region, the ITRs had to be retained for a long evolutionary time in some species, while they were lost in others.
A connection between ITRs and chromosomal fusions is also not supported for the family Testudinidae. Previous studies did not detect ITRs in Chelonoidis carbonarius and C. chilensis [72,75]. In Chelonoidis duncanensis, we detected centromeric ITRs in four chromosome pairs (Figure 3d), although all three species of the genus share the same chromosome number 2n = 52. We also detected ITRs in centromeres of three chromosome pairs in Stigmochelys pardalis, another species sharing the same chromosome number (2n = 52) typical for the family Testudinidae (Figure 3e). The phylogenetic position and the different topology of the accumulation of telomeric-like repeats suggest that ITRs evolved within testudinidids independently in C. duncanensis and S. pardalis (Figure 1).
Most turtles have environmental sex determination (ESD) [76]. As far as we know, genotypic sex determination (GSD), and hence sex chromosomes, evolved independently at least five times within turtles: ZZ/ZW sex chromosomes in the common ancestor of softshell turtles (family Trionychidae [102]), XX/XY sex chromosomes in the genus Glyptemys (family Emydidae [103], in the genus Staurotypus (family Kinosternidae [104,105]), in Siebenrockiella crassicollis (Geoemydidae [106]) and in chelid turtles (reviewed in [81]).
Non-recombining parts of the unpaired W and Y chromosomes tend to degenerate, i.e., to lose functional genes [107,108,109], and to accumulate repeats such as microsatellites [49,53,56,62,110,111], rDNA-derived repeats [50,73,81,112] and telomeric-like repeats [27,28,61,113,114,115]. In turtles, the Y chromosome is not enriched in telomeric-like sequences in Glyptemys insculpta [76], Staurotypus salvinii and S. triporcatus [76]. Similarly, a degenerated W chromosome in trionychid turtles is full of rDNA-derived repeats, but no accumulation of telomeric-like repeats was detected in Apalone ferox, Lissemys punctata (Figure 4x,y), Apalone spinifera or Pelodiscus sinensis [76]. Within chelids, ITRs are present on the Y chromosomes in Elseya novaeguineae [81] but not on the putatively homologous Y chromosomes of the genera Chelodina and Emydura [76,81]. The comparison of repeat content on unpaired sex chromosomes across turtles suggests that it can be very variable and can reflect historical contingency rather than a functional aspect of a given repeat.
ITRs occur only rarely in turtles, especially in comparison to squamates [28]. Wherever present in turtles, ITRs are restricted to centromeric regions and to the non-recombining parts of sex chromosomes (Figure 1). The distribution of ITRs agrees with the observation that turtle karyotypes are very conserved [85] and that centromeres and non-recombining parts of sex chromosomes are the most dynamic parts of genomes [116,117]. Centromeric ITRs do not seem to be directly related to interchromosomal rearrangements in turtles. If ITRs were formed during fusions, they had to be subsequently lost in several cases. Such a scenario was supported, for instance, by Nanda et al. [39] for birds, possibly due to selection for smaller genome size counteracting the accumulation of repetitions and resulting in the disappearance of ITRs. That the presence of centromeric ITRs is not related to rearrangements is further supported by the fact that in species possessing ITRs, the number of chromosomes with ITRs does not agree with the estimated number of interchromosomal rearrangements for the respective karyotype. However, our conclusions on the homology of chromosomes with ITRs are based on the comparison of chromosome morphology, and should be further tested using more informative approaches, such as comparative BAC-FISH or chromosome painting.
Interestingly, although ITRs are generally rare in turtles, whenever they occur, they tend to be present in centromeres in more than one chromosome pair. ITRs are present in two chromosome pairs in the kinosternids Claudius angustatus and Staurotypus salvinii, in three chromosome pairs in Stigmochelys pardalis, in four chromosome pairs in Chelonoidis duncanensis, and even in seven chromosome pairs in Podocnemis unifilis and Chelydra serpentina (Figure 1; [76]). This observation can be attributed to a phenomenon known as “centromere homogenization” potentially caused by the involvement of telomeric-like repeats in retrotransposons colonizing centromeres [118]. Another interesting observation is that only centromeric ITRs were detected in turtle autosomes. One reason can be that centromeric ITRs are the most common type, at least in squamates [28]. It is therefore likely that they would be overrepresented also in the few cases of ITRs found in turtles. However, it is also possible that in turtles, events leading to ITRs within chromosome arms are rarer. Intermediate ITRs have been found in squamates [28] and in birds [39], and it was speculated that they were formed due to chromosome inversions, which are common in these groups. Turtles, thus, may be less prone to intrachromosomal inversions than squamates [119] and birds [97,120]. As these inversions are important for the evolution of postzygotic reproductive isolation mechanisms, and thus speciation [120,121,122,123,124], this feature may contribute to the generally low species diversity in turtles [70] and their slow molecular evolution [125,126,127]. The low rate of inversions in the karyotype evolution of turtles could explain why they exhibit a high incidence of genomic introgression and why they extensively hybridize (and are often able to produce fertile offspring), even after many million years of divergence, as documented in chelid, cheloniid and geoemydid turtles [81,91,128,129,130,131,132,133,134,135,136].
In turtles, the TTAGGG motif is restricted only to the centromeric regions. The lack of a strong phylogenetic signal in the distribution of ITRs across all studied species (Figure 1) indicates that the telomeric-like sequences in the centromeres of turtles are not connected to chromosomal rearrangements. Nevertheless, the identification of ITRs is a more complicated process. Genome sequencing projects revealed that short arrays of ITRs, below the detection efficiency of in situ hybridization methods, are common in vertebrate genomes [19,20,21,22,23,24,25,26]. Such s-ITRs are expected to exist in turtle genomes as well. Furthermore, long ITRs, e.g., derived from chromosome fusions, are non-essential, non-coding regions, and like other microsatellite motifs, they can degenerate over time by reduction of copies and/or by accumulation of mutations, which will complicate their detection by in situ hybridization or bioinformatic methods. Previous studies in rodents [27] and birds [68,69] documented that the TTAGGG motif can be part of a longer satellite sequence occurring in centromeres and other heterochromatic regions. Notably, a telomeric-like TCATGGG motif forming long tandem repeats has been identified in the pericentromeric regions of Drosophila hydei, and this motif does not seem to have a telomere-related function [137], suggesting that this could also be the case in turtles with ITRs.
In conclusion, the centromere organization and sequence composition in turtles are currently not well known. This calls for further research to understand whether or not the TTAGGG motif in the centromeres of turtles has a non-telomeric origin, e.g., using long-read throughput sequencing of the centromeric regions of phylogenetically informative species.
Author Contributions
L.C. and S.M. performed the cytogenetic analyses and prepared the figures; E.P.B., B.A., L.K. and M.R. contributed to the experimental part; T.P., P.P., M.A., P.V., P.W. and U.F. provided part of the material and useful consultations; M.R. conceived and led the project; L.C., S.M., U.F., L.K. and M.R. drafted the first version of the manuscript. All authors edited and approved the final manuscript.
Funding
This study was supported by Czech Science Foundation project No. 20-27236J, Charles University Grant Agency (project 1191619), Charles University projects PRIMUS/SCI/46 and Research Centre program (204069).
Acknowledgments
We would like to express our gratitude to Tomáš Jirásek (Zoo Plzeň) for providing material for the study, Petr Ráb and members of his laboratory for their support, Jana Thomayerová and Nuria Viñuela Rodríguez for technical support.
Conflicts of Interest
The authors declare no conflict of interest
References
- Blackburn, E.H. Switching and signaling at the telomere. Cell 2001, 106, 661–673. [Google Scholar] [CrossRef]
- Moyzis, R.K.; Buckingham, J.M.; Crams, L.S.; Dani, M.; Larry, L.; Jones, M.D.; Meyne, J.; Ratliff, R.L.; Wu, J.R. A highly conserved repetitive DNA sequence, (TTAGGG)n, present at the telomeres of human chromosomes. Proc. Natl. Acad. Sci. USA 1988, 85, 6622–6626. [Google Scholar] [CrossRef]
- Meyne, J.; Baker, R.J.; Hobart, H.H.; Hsu, T.C.; Ryder, O.A.; Ward, O.G.; Wiley, J.E.; Wurster-Hill, D.H.; Yates, T.L.; Moyzis, R.K. Distribution of non-telomeric sites of the (TTAGGG)n telomeric sequence in vertebrate chromosomes. Chromosoma 1990, 99, 3–10. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, R.J.; Karlseder, J. Telomeres: Protecting chromosomes against genome instability. Nat. Rev. Mol. Cell Biol. 2010, 11, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Chiodi, I.; Belgiovine, C.; Mondello, C. Telomerase and telomeric proteins: A life beyond telomeres. In Telomerase: Composition, Functions and Clinical Implications; Gagnon, A.N., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2010; pp. 35–58. [Google Scholar]
- Zhao, Z.; Pan, X.; Liu, L.; Liu, N. Telomere length maintenance, shortening, and lengthening. J. Cell. Physiol. 2014, 229, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Jafri, M.A.; Ansari, S.A.; Alqahtani, M.H.; Shay, J.W. Roles of telomeres and telomerase in cancer, and advances in telomerase-targeted therapies. Genome Med. 2016, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Cong, Y.S.; Wright, W.E.; Shay, J.W. Human telomerase and its regulation. Microbiol. Mol. Biol. Rev. 2002, 66, 407–425. [Google Scholar] [CrossRef]
- Harley, B.C.; Futcher, B.A.; Greider, W.C. Telomeres shorten during ageing of human fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef]
- Hastie, N.D.; Dempster, M.; Dunlop, M.G.; Thompson, A.M.; Green, D.K.; Allshire, R.C. Telomere reduction in human colorectal carcinoma and with ageing. Nature 1990, 346, 866–868. [Google Scholar] [CrossRef]
- Lindsey, J.; McGill, N.I.; Lindsey, L.A.; Green, D.K.; Cooke, H.J. In vivo loss of telomeric repeats with age in humans. Mutat. Res. 1991, 256, 45–48. [Google Scholar] [CrossRef]
- Hayashi, M.T.; Cesare, A.J.; Rivera, T.; Karlseder, J. Cell death during crisis is mediated by mitotic telomere deprotection. Nature 2015, 522, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Bolzán, A.D.; Bianchi, M.S. Telomeres, interstitial telomeric repeat sequences, and chromosomal aberrations. Mutat. Res. 2006, 612, 189–214. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.W.; Yan, J. Endings in the middle: Current knowledge of interstitial telomeric sequences. Mutat. Res. 2008, 658, 95–110. [Google Scholar] [CrossRef]
- Ruiz-Herrera, A.; Nergadze, S.G.; Santagostino, M.; Giulotto, E. Telomeric repeats far from the ends: Mechanisms of origin and role in evolution. Cytogenet. Genome Res. 2008, 122, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Mucciolo, E.; Bertoni, L.; Giuliotto, E. Fluorescence in situ hybridization with a synthetic (T2AG3)n polynucleotide detects several intrachromosomal telomere-like repeats on human chromosomes. Cytogenet. Genome Res. 1997, 78, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Bolzán, A.D. Interstitial telomeric sequences in vertebrate chromosomes: Origin, function, instability and evolution. Mutat. Res. 2017, 773, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Santagostino, M.; Piras, F.M.; Cappelletti, E.; Del Giudice, S.; Semino, O.; Nergadze, S.G.; Giulotto, E. Insertion of telomeric repeats in the human and horse genomes: An evolutionary perspective. Int. J. Mol. Sci. 2020, 21, 2838. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Nergadze, S.G.; Giulotto, E. Human intrachromosomal telomeric-like repeats: Sequence organization and mechanisms of origin. Chromosoma 2001, 110, 75–82. [Google Scholar] [CrossRef]
- Faravelli, M.; Moralli, D.; Bertoni, L.; Attolini, C.; Chernova, O.; Raimondi, E.; Giulotto, E. Two extended arrays of a satellite DNA sequence at the centromere and at the short-arm telomere of Chinese hamster chromosome. Cytogenet. Genome Res. 1998, 83, 281–286. [Google Scholar] [CrossRef]
- Faravelli, M.; Azzalin, C.M.; Bertoni, L.; Chernova, O.; Attolini, C.; Mondello, C.; Giulotto, E. Molecular organization of internal telomeric sequences in Chinese hamster chromosomes. Gene 2002, 283, 11–16. [Google Scholar] [CrossRef]
- Ruiz-Herrera, A.; García, F.; Azzalin, C.; Giulotto, E.; Egozcue, J.; Ponsà, M.; Garcia, M. Distribution of intrachromosomal telomeric sequences (ITS) on Macaca fascicularis (Primates) chromosomes and their implication for chromosome evolution. Hum. Genet. 2002, 110, 578–586. [Google Scholar] [CrossRef]
- Ruiz-Herrera, A.; García, F.; Giulotto, E.; Attolini, C.; Egozcue, J.; Ponsà, M.; Garcia, M. Evolutionary breakpoints are co-localized with fragile sites and intrachromosomal telomeric sequences in primates. Cytogenet. Genome Res. 2005, 108, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Nergadze, S.G.; Rocchi, M.; Azzalin, C.M.; Mondello, C.; Giulotto, E. Insertion of telomeric repeats at intrachromosomal break sites during primate evolution. Genome Res. 2004, 14, 1704–1710. [Google Scholar] [CrossRef] [PubMed]
- Nergadze, S.G.; Santagostino, M.A.; Salzano, A.; Mondello, C.; Giulotto, E. Contribution of telomerase RNA retrotranscription to DNA double-strand break repair during mammalian genome evolution. Genome Biol. 2007, 8, R260. [Google Scholar] [CrossRef] [PubMed]
- Camats, N.; Ruiz-Herrera, A.; Parrilla, J.J.; Acien, M.; Payá, P.; Giulotto, E.; Egozcue, J.; García, F.; Garcia, M. Genomic instability in rat: Breakpoints induced by ionising radiation and interstitial telomeric-like sequences. Mutat. Res. 2006, 595, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.T.; Marchal, J.A.; Romero-Fernández, I.; Fernández, F.J.; Giagia-Athanosopoulou, E.B.; Sánchez, A. Rapid, independent, and extensive amplification of telomeric repeats in pericentromeric regions in karyotypes of arvicoline rodents. Chromosome Res. 2011, 19, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Kratochvíl, L.; Altmanová, M.; Johnson Pokorná, M. Interstitial telomeric motifs in squamate reptiles: When the exceptions outnumber the rule. PLoS ONE 2015, 10, e0134985. [Google Scholar] [CrossRef]
- Mazzoleni, S.; Schillaci, O.; Sineo, L.; Dumas, F. Distribution of interstitial telomeric sequences in primates and the pygmy tree shrew (Scandentia). Cytogenet. Genome Res. 2017, 151, 141–150. [Google Scholar] [CrossRef]
- Milioto, V.; Vlah, S.; Mazzoleni, S.; Rovatsos, M.; Dumas, F. Chromosomal localization of 18S-28S rDNA and (TTAGGG)n sequences in two South African dormice of the genus Graphiurus (Rodentia: Gliridae). Cytogenet. Genome Res. 2019, 158, 145–151. [Google Scholar] [CrossRef]
- Swier, V.J.; Anwarali Khan, F.A.; Baker, R.J. Do time, heterochromatin, NORs, or chromosomal rearrangements correlate with distribution of interstitial telomeric repeats in Sigmodon (cotton rats)? J. Hered. 2012, 103, 493–502. [Google Scholar] [CrossRef]
- Nagamachi, C.Y.; Pieczarka, J.C.; O’Brien, P.C.M.; Pinto, J.A.; Malcher, S.M.; Pereira, A.L.; das Dores Rissino, J.; Mendes-Oliveira, A.C.; Rossi, R.V.; Ferguson-Smith, M.A. FISH with whole chromosome and telomeric probes demonstrates huge karyotypic reorganization with ITS between two species of Oryzomyini (Sigmodontinae, Rodentia): Hylaeamys megacephalus probes on Cerradomys langguthi karyotype. Chromosome Res. 2013, 21, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Wiley, J.E.; Meyne, J.; Little, M.L.; Stout, J.C. Interstitial hybridization sites of the (TTAGGG)n telomeric sequence on the chromosomes of some North American hylid frogs. Cytogenet. Cell Genet. 1992, 61, 55–57. [Google Scholar] [CrossRef]
- Park, V.M.; Gustashaw, K.M.; Wathen, T.M. The presence of interstitial telomeric sequences in constitutional chromosome abnormalities. Am. J. Hum. Genet. 1992, 50, 914–923. [Google Scholar] [PubMed]
- Rossi, E.; Floridia, G.; Casali, M.; Danesino, C.; Chiumello, G.; Bernardi, F.; Magnani, I.; Papi, L.; Mura, M.; Zuffardi, O. Types, stability, and phenotypic consequences of chromosome rearrangements leading to interstitial telomeric sequences. J. Med. Genet. 1993, 30, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Vermeesch, J.R.; Petit, P.; Speleman, F.; Devriendt, K.; Fryns, J.P.; Marynen, P. Interstitial telomeric sequences at the junction site of a jumping translocation. Hum. Genet. 1997, 99, 735–737. [Google Scholar] [CrossRef] [PubMed]
- Melek, M.; Shippen, D.E. Chromosome healing: Spontaneous and programmed de novo telomere formation by telomerase. BioEssays 1996, 18, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.R.; Wright, W.E. Telomeres and telomerase: Three decades of progress. Nat. Rev. Genet. 2019, 20, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Nanda, I.; Schrama, D.; Feichtinger, W.; Haaf, T.; Schartl, M.; Schmid, M. Distribution of telomeric (TTAGGG)n sequences in avian chromosomes. Chromosoma 2002, 111, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Marchal, J.A.; Romero-Fernández, I.; Cano-Linares, M.; Fernández, F.J.; Giagia-Athanasopoulou, E.B.; Sánchez, A. Molecular and physical characterization of the complex pericentromeric heterochromatin of the vole species Microtus thomasi. Cytogenet. Genome Res. 2014, 144, 131–141. [Google Scholar] [CrossRef]
- Ocalewicz, K. Telomeres in fishes. Cytogenet. Genome Res. 2013, 141, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Bruschi, D.P.; Rivera, M.; Lima, A.P.; Zúñiga, A.B.; Recco-Pimentel, S.M. Interstitial telomeric sequences (ITS) and major rDNA mapping reveal insights into the karyotypical evolution of Neotropical leaf frogs species (Phyllomedusa, Hylidae, Anura). Mol. Cytogenet. 2014, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Steinlein, C. Chromosome Banding in Amphibia. XXXIV. Intrachromosomal telomeric DNA sequences in Anura. Cytogenet. Genome Res. 2016, 148, 211–226. [Google Scholar] [CrossRef]
- De Oliveira, T.D.; Kretschmer, R.; Bertocchi, N.A.; Degrandi, T.M.; de Oliveira, E.H.C.; Cioffi, M.B.; Garnero, A.D.V.; Gunski, R.J. Genomic organization of repetitive DNA in woodpeckers (Aves, Piciformes): Implications for karyotype and ZW sex chromosome differentiation. PLoS ONE 2017, 12, e0169987. [Google Scholar] [CrossRef] [PubMed]
- Srikulnath, K.; Azad, B.; Singchat, W.; Ezaz, T. Distribution and amplification of interstitial telomeric sequences (ITSs) in Australian dragon lizards support frequent chromosome fusions in Iguania. PLoS ONE 2019, 14, e0212683. [Google Scholar] [CrossRef] [PubMed]
- Zattera, M.L.; Lima, L.; Duarte, I.; de Sousa, D.Y. Cytogenetics chromosome spreading of the (TTAGGG)n repeats in the Pipa carvalhoi Miranda-Ribeiro, 1937 (Pipidae, Anura) karyotype. Comp. Cytogenet. 2019, 13, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Johnson Pokorná, M.; Altmanová, M.; Kratochvíl, L. Female heterogamety in Madagascar chameleons (Squamata: Chamaeleonidae: Furcifer): Differentiation of sex and neo-sex chromosomes. Sci. Rep. 2015, 5, 13196. [Google Scholar] [CrossRef]
- Rovatsos, M.; Johnson Pokorná, M.; Kratochvíl, L. Differentiation of sex chromosomes and karyotype characterisation in the dragonsnake Xenodermus javanicus (Squamata: Xenodermatidae). Cytogenet. Genome Res. 2015, 147, 48–54. [Google Scholar] [CrossRef]
- Rovatsos, M.; Johnson Pokorná, M.; Altmanová, M.; Kratochvíl, L. Mixed-up sex chromosomes: Identification of sex chromosomes in the X1X1X2X2/X1X2Y system of the legless lizards of the genus Lialis (Squamata: Gekkota: Pygopodidae). Cytogenet. Genome Res. 2016, 149, 282–289. [Google Scholar] [CrossRef]
- Rovatsos, M.; Altmanová, M.; Johnson Pokorná, M.; Velenský, P.; Sánchez Baca, A.; Kratochvíl, L. Evolution of karyotypes in chameleons. Genes 2017, 8, 382. [Google Scholar] [CrossRef]
- Rovatsos, M.; Altmanová, M.; Johnson Pokorná, M.; Augstenová, B.; Kratochvíl, L. Cytogenetics of the Javan file snake (Acrochordus javanicus) and the evolution of snake sex chromosomes. J. Zool. Syst. Evol. Res. 2018, 56, 117–125. [Google Scholar] [CrossRef]
- Rovatsos, M.; Altmanová, M.; Augstenová, B.; Mazzoleni, S.; Velenský, P.; Kratochvíl, L. ZZ/ZW sex determination with multiple neo-sex chromosomes is common in Madagascan chameleons of the genus Furcifer. Genes 2019, 10, 1020. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Uno, Y.; Srikulnath, K.; Matsuda, Y.; Miller, E.; Olsson, M. No interstitial telomeres on autosomes but remarkable amplification of telomeric repeats on the W sex chromosome in the sand lizard (Lacerta agilis). J. Hered. 2015, 106, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Srikulnath, K.; Uno, Y.; Nishida, C.; Ota, H.; Matsuda, Y. Karyotype reorganization in the Hokou gecko (Gekko hokouensis, Gekkonidae): The process of microchromosome disappearance in Gekkota. PloS ONE 2015, 10, e0134829. [Google Scholar] [CrossRef]
- Viana, P.F.; Ribeiro, L.B.; Souza, G.M.; Chalkidis, H.D.M.; Gross, M.C.; Feldberg, E. Is the karyotype of neotropical boid snakes really conserved? Cytotaxonomy, chromosomal rearrangements and karyotype organization in the Boidae family. PLoS ONE 2016, 11, e0160274. [Google Scholar] [CrossRef]
- Viana, P.F.; Ezaz, T.; Cioffi, M.D.B.; Almeida, B.J.; Feldberg, E. Evolutionary insights of the ZW sex chromosomes in snakes: A new chapter added by the Amazonian puffing snakes of the genus Spilotes. Genes 2019, 10, 288. [Google Scholar] [CrossRef]
- Altmanová, M.; Rovatsos, M.; Kratochvíl, L.; Johnson Pokorná, M. Minute Y chromosomes and karyotype evolution in Madagascan iguanas (Squamata: Iguania: Opluridae). Biol. J. Linn. Soc. 2016, 118, 618–633. [Google Scholar] [CrossRef]
- Giovannotti, M.; Nisi Cerioni, P.; Slimani, T.; Splendiani, A.; Paoletti, A.; Fawzi, A.; Olmo, E.; Caputo Barucchi, V. Cytogenetic characterization of a population of Acanthodactylus lineomaculatus Duméril and Bibron, 1839 (Reptilia, Lacertidae), from Southwestern Morocco and insights into sex chromosome evolution. Cytogenet. Genome Res. 2017, 153, 86–95. [Google Scholar] [CrossRef]
- Giovannotti, M.; Nisi Cerioni, P.; Rojo, V.; Olmo, E.; Slimani, T.; Splendiani, A.; Caputo Barucchi, V. Characterization of a satellite DNA in the genera Lacerta and Timon (Reptilia, Lacertidae) and its role in the differentiation of the W chromosome. J. Exp. Zool. B Mol. Dev. Evol. 2018, 330, 83–95. [Google Scholar] [CrossRef]
- Augstenová, B.; Mazzoleni, S.; Kratochvíl, L.; Rovatsos, M. Evolutionary dynamics of the W chromosome in caenophidian snakes. Genes 2018, 9, 5. [Google Scholar] [CrossRef]
- Augstenová, B.; Mazzoleni, S.; Kostmann, A.; Altmanová, M.; Frynta, D.; Kratochvíl, L.; Rovatsos, M. Cytogenetic analysis did not reveal differentiated sex chromosomes in ten species of boas and pythons (Reptilia: Serpentes). Genes 2019, 10, 934. [Google Scholar] [CrossRef]
- Singchat, W.; O’Connor, R.E.; Tawichasri, P.; Suntronpong, A.; Sillapaprayoon, S.; Suntrarachun, S.; Muangmai, N.; Baicharoen, S.; Peyachoknagul, S.; Chanhome, L.; et al. Chromosome map of the Siamese cobra: Did partial synteny of sex chromosomes in the amniote represent “a hypothetical ancestral super-sex chromosome” or random distribution? BMC Genom. 2018, 19, 939. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.J.; de Araújo Vieira, A.P.; Galvão Cipriano, F.M.; dos Santos Cândido, M.R.; de Oliveira, E.H.C.; Gimenez Pinheiro, T.; da Silva, E.L. The karyotype of Salvator merianae (Squamata, Teiidae): Analyses by classical and molecular cytogenetic techniques. Cytogenet. Genome Res. 2020, 160, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Sidhom, M.; Said, K.; Chatti, N.; Guarino, F.M.; Odierna, G.; Petraccioli, A.; Picariello, O.; Mezzasalma, M. Karyological characterization of the common chameleon (Chamaeleo chamaeleon) provides insights on the evolution and diversification of sex chromosomes in Chamaeleonidae. Zoology 2020, 125738, in press. [Google Scholar] [CrossRef] [PubMed]
- Kawagoshi, T.; Nishida, C.; Ota, H.; Kumazawa, Y.; Endo, H.; Matsuda, Y. Molecular structures of centromeric heterochromatin and karyotypic evolution in the Siamese crocodile (Crocodylus siamensis) (Crocodylidae, Crocodylia). Chromosome Res. 2008, 16, 1119–1132. [Google Scholar] [CrossRef] [PubMed]
- Nishida, C.; Ishijima, J.; Kosaka, A.; Tanabe, H.; Habermann, F.A.; Griffin, D.K.; Matsuda, Y. Characterization of chromosome structures of Falconinae (Falconidae, Falconiformes, Aves) by chromosome painting and delineation of chromosome rearrangements during their differentiation. Chromosome Res. 2008, 16, 171–181. [Google Scholar] [CrossRef]
- Seibold-Torres, C.; Owens, E.; Chowdhary, R.; Ferguson-Smith, M.A.; Tizard, I.; Raudsepp, T. Comparative cytogenetics of the Congo African grey parrot (Psittacus erithacus). Cytogenet. Genome Res. 2015, 147, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Liangouzov, I.A.; Derjusheva, S.E.; Saifitdinova, A.F.; Malykh, A.G.; Gaginskaya, E.R. Monomers of a satellite DNA sequence of chaffinch (Fringilla coelebs L., Aves: Passeriformes) contain short clusters of the TTAGGG repeat. Russ. J. Genet. 2002, 38, 1359–1364. [Google Scholar] [CrossRef]
- Derjusheva, S.; Kurganova, A.; Habermann, F.; Gaginskaya, E. High chromosome conservation detected by comparative chromosome painting in chicken, pigeon and passerine birds. Chromosome Res. 2004, 12, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Uetz, P.; Freed, P.; Hošek, J. The Reptile Database. Available online: http://www.reptile-database.org (accessed on 13 April 2020).
- Vargas-Ramírez, M.; Caballero, S.; Morales-Betancourt, M.A.; Lasso, C.A.; Amaya, L.; Martínez, J.G.; das Neves Silvia Viana, M.; Vogt, R.C.; Farias, I.P.; Hrbek, T.; et al. Genomic analyses reveal two species of the matamata (Testudines: Chelidae: Chelus spp.) and clarify their phylogeography. Mol. Phylogenet. Evol. 2020, 148, 106823. [Google Scholar]
- Martinez, P.A.; Boeris, J.M.; Sánchez, J.; Pastori, M.C.; Bolzán, A.D.; Ledesma, M.A. Karyotypic characterization of Trachemys dorbigni (Testudines: Emydidae) and Chelonoidis (Geochelone) donosobarrosi (Testudines: Testudinidae), two species of cryptodiran turtles from Argentina. Genetica 2009, 137, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Badenhorst, D.; Hillier, L.D.; Literman, R.; Montiel, E.E.; Radhakrishnan, S.; Shen, Y.; Minx, P.; Janes, D.E.; Warren, W.C.; Edwards, S.V.; et al. Physical mapping and refinement of the painted turtle genome (Chrysemys picta) inform amniote genome evolution and challenge turtle-bird chromosomal conservation. Genome Biol. Evol. 2013, 7, 2038–2050. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ventura, K.; Moreira, C.N.; Moretti, R.; Yonenaga-Yassuda, Y.; Rodrigues, M.T. The lowest diploid number in Testudines: Banding patterns, telomeric and 45s rDNA FISH in Peltocephalus dumerilianus, 2n = 26 and FN = 52 (Pleurodira, Podocnemididae). Genet. Mol. Biol. 2014, 37, 61–63. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, J.; Alcalde, L.; Bolzán, A.D. First evidence of chromosomal variation within Chelonoidis chilensis (Testudines: Testudinidae). Herpetol. J. 2015, 25, 83–89. [Google Scholar]
- Montiel, E.E.; Badenhorst, D.; Lee, L.S.; Literman, R.; Trifonov, V.; Valenzuela, N. Cytogenetic insights into the evolution of chromosomes and sex determination reveal striking homology of turtle sex chromosomes to amphibian autosomes. Cytogenet. Genome Res. 2016, 148, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Noronha, R.C.R.; Barros, L.M.R.; Araújo, R.E.F.; Marques, D.F.; Nagamachi, C.Y.; Martins, C.; Pieczarka, J.C. New insights of karyoevolution in the Amazonian turtles Podocnemis expansa and Podocnemis unifilis (Testudines, Podocnemidae). Mol. Cytogenet. 2016, 9, 73. [Google Scholar] [CrossRef]
- Cavalcante, M.G.; Bastos, C.E.M.C.; Nagamachi, C.Y.; Pieczarka, J.C.; Vicari, M.R.; Noronha, R.C.R. Physical mapping of repetitive DNA suggests 2n reduction in Amazon turtles Podocnemis (Testudines: Podocnemididae). PLoS ONE 2018, 13, e0197536. [Google Scholar] [CrossRef] [PubMed]
- Iannucci, A.; Svartman, M.; Bellavita, M.; Chelazzi, G.; Stanyon, R.; Ciofi, C. Insights into Emydid turtle cytogenetics: The European pond turtle as a model species. Cytogenet. Genome Res. 2019, 157, 166–171. [Google Scholar] [CrossRef]
- Mazzoleni, S.; Augstenová, B.; Clemente, L.; Auer, M.; Fritz, U.; Praschag, P.; Protiva, T.; Velenský, P.; Kratochvíl, L.; Rovatsos, M. Turtles of the genera Geoemyda and Pangshura (Testudines: Geoemydidae) lack differentiated sex chromosomes: The end of a 40-year error cascade for Pangshura. PeerJ 2019, 7, e6241. [Google Scholar] [CrossRef]
- Mazzoleni, S.; Augstenová, B.; Clemente, L.; Auer, M.; Fritz, U.; Praschag, P.; Protiva, T.; Velenský, P.; Kratochvíl, L.; Rovatsos, M. Sex is determined by XX/XY sex chromosomes in Australasian side-necked turtles (Testudines: Chelidae). Sci. Rep. 2020, 10, 4276. [Google Scholar] [CrossRef]
- Cavalcante, M.G.; Souza, L.F.; Vicari, M.R.; de Bastos, C.E.M.; de Sousa, J.V.; Nagamachi, C.Y.; Pieczarka, J.C.; Martins, C.; Noronha, R.C.R. Molecular cytogenetics characterization of Rhinoclemmys punctularia (Testudines, Geoemydidae) and description of a Gypsy-H3 association in its genome. Gene 2020, 738, 144477. [Google Scholar] [CrossRef]
- Olmo, E.; Signorino, G.G. Chromorep: A reptile chromosomes database. Available online: http://chromorep.univpm.it (accessed on 14 April 2020).
- Bickham, J.W. Two hundred million year old chromosomes: Deceleration of the rate of karyotypic evolution in turtles. Science 1981, 212, 1291–1293. [Google Scholar] [CrossRef] [PubMed]
- Olmo, E. Trends in the evolution of reptilian chromosomes. Integr. Comp. Biol. 2008, 48, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Fritz, U.; Guicking, D.; Auer, M.; Sommer, R.S.; Wink, M.; Hundsdörfer, A.K. Diversity of the Southeast Asian leaf turtle genus Cyclemys: How many leaves on its tree of life? Zool. Scr. 2008, 37, 367–390. [Google Scholar] [CrossRef]
- Petzold, A.; Vargas-Ramírez, M.; Kehlmaier, C.; Vamberger, M.; Branch, W.R.; Du Preez, L.; Hofmeyr, M.D.; Schleicher, A.; Široký, P.; Fritz, U. A revision of African helmeted terrapins (Testudines: Pelomedusidae: Pelomedusa), with descriptions of six new species. Zootaxa 2014, 3795, 523–548. [Google Scholar] [CrossRef]
- Ihlow, F.; Vamberger, M.; Flecks, M.; Hartmann, T.; Cota, M.; Makchai, S.; Meewattana, P.; Dawson, J.E.; Kheng, L.; Rödder, D. Integrative taxonomy of Southeast Asian snail-eating turtles (Geoemydidae: Malayemys) reveals a new species and mitochondrial introgression. PLoS ONE 2016, 11, e0153108. [Google Scholar] [CrossRef]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved telomere detection using a telomere repeat probe (TTAGGG)n generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef]
- Pereira, A.G.; Sterli, J.; Moreira, F.R.R.; Schrago, C.G. Multilocus phylogeny and statistical biogeography clarify the evolutionary history of major lineages of turtles. Mol. Phylogenet. Evol. 2017, 113, 59–66. [Google Scholar] [CrossRef]
- Kehlmaier, C.; Zhang, X.; Georges, A.; Campbell, P.D.; Thomson, S.; Fritz, U. Mitogenomics of historical type specimens of Australasian turtles: Clarification of taxonomic confusion and old mitochondrial introgression. Sci. Rep. 2019, 9, 5841. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A modular system for evolutionary analysis. Version 3.61. Available online: http://mesquiteproject.org (accessed on 15 March 2020).
- Delany, M.E.; Krupkin, A.B.; Miller, M.M. Organization of telomere sequences in birds: Evidence for arrays of extreme length and for in vivo shortening. Cytogenet. Cell Genet. 2000, 90, 139–145. [Google Scholar] [CrossRef]
- Raudsepp, T.; Houck, M.L.; O’Brien, P.C.; Ferguson-Smith, M.A.; Ryder, O.A.; Chowdhary, B.P. Cytogenetic analysis of California condor (Gymnogyps californianus) chromosomes: Comparison with chicken (Gallus gallus) macrochromosomes. Cytogenet. Genome Res. 2002, 98, 54–60. [Google Scholar] [CrossRef]
- Swanberg, S.E.; Delany, M.E. Telomeres in aging: Birds. In Handbook of Models for Human Aging; Conn, P.M., Ed.; Elsevier Academic Press: Cambridge, MA, USA, 2006; pp. 339–349. [Google Scholar]
- Dos Santos, M.S.; Kretschmer, R.; Silva, F.A.O.; Ledesma, M.A.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Del Valle Garnero, A.; de Oliveira, E.H.C.; Gunski, R.J. Intrachromosomal rearrangements in two representatives of the genus Saltator (Thraupidae, Passeriformes) and the occurrence of heteromorphic Z chromosomes. Genetica 2015, 143, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Karyotype evolution in birds: From conventional staining to chromosome painting. Genes 2018, 9, 181. [Google Scholar] [CrossRef]
- Johnson Pokorná, M.; Rovatsos, M.; Kratochvíl, L. Sex chromosomes and karyotype of the (nearly) mythical creature, the Gila monster, Heloderma suspectum (Squamata: Helodermatidae). PLoS ONE 2014, 9, e104716. [Google Scholar]
- Stock, A.D. Karyological relationships in turtles (Reptilia: Chelonia). Can. J. Genet. Cytol. 1972, 14, 859–868. [Google Scholar] [CrossRef]
- Killebrew, F.C. Mitotic chromosomes of turtles. III. The Kinosternidae. Herpetologica 1975, 31, 398–403. [Google Scholar]
- Rhodin, A.G.J.; Iverson, J.B.; Bour, R.; Fritz, U.; Georges, A.; Shaffer, H.B. Turtles of the World: Annotated Checklist and Atlas of Taxonomy. Synon. Distrib. Conserv. Status 2017, 8, 9–14. [Google Scholar]
- Rovatsos, M.; Praschag, P.; Fritz, U.; Kratochvíl, L. Stable Cretaceous sex chromosomes enable molecular sexing in softshell turtles (Testudines: Trionychidae). Sci. Rep. 2017, 7, 42150. [Google Scholar] [CrossRef] [PubMed]
- Literman, R.; Radhakrishnan, S.; Tamplin, J.; Burke, R.; Dresser, C.; Valenzuela, N. Development of sexing primers in Glyptemys insculpta and Apalone spinifera turtles uncovers an XX/XY sex-determining system in the critically-endangered bog turtle Glyptemys muhlenbergii. Conserv. Genet. Res. 2017, 9, 651–658. [Google Scholar] [CrossRef]
- Bull, J.J.; Moon, R.G.; Legler, J.M. Male heterogamety in kinosternid turtles (genus Staurotypus). Cytogenet. Genome Res. 1974, 13, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Kawagoshi, T.; Uno, Y.; Nishida, C.; Matsuda, Y. The Staurotypus turtles and Aves share the same origin of sex chromosomes but evolved different types of heterogametic sex determination. PLoS ONE 2014, 9, e105315. [Google Scholar] [CrossRef] [PubMed]
- Kawagoshi, T.; Nishida, C.; Matsuda, Y. The origin and differentiation process of X and Y chromosomes of the black marsh turtle (Siebenrockiella crassicollis, Geoemydidae, Testudines). Chromosome Res. 2012, 20, 95–110. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ohno, S. Sex Chromosomes and Sex-Linked Genes; Springer: Berlin, Germany, 1967; Volume 1. [Google Scholar]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity 2005, 95, 118–128. [Google Scholar] [CrossRef]
- Vicoso, B. Molecular and evolutionary dynamics of animal sex-chromosome turnover. Nat. Ecol. Evol. 2019, 3, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Kejnovsky, E.; Bertollo, L.A.C. The chromosomal distribution of microsatellite repeats in the genome of the wolf fish Hoplias malabaricus, focusing on the sex chromosomes. Cytogenet. Genome Res. 2011, 132, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Knopp, T.; Sarre, S.D.; Georges, A.; Ezaz, T. Karyotypic analysis and FISH mapping of microsatellite motifs reveal highly differentiated XX/XY sex chromosomes in the pink-tailed worm-lizard (Aprasia parapulchella, Pygopodidae, Squamata). Mol. Cytogenet. 2013, 6, 60. [Google Scholar] [CrossRef] [PubMed]
- Scacchetti, P.C.; Utsunomia, R.; Pansonato-Alves, J.C.; da Costa Silva, G.J.; Vicari, M.R.; Artoni, F.R.; Oliveira, C.; Foresti, F. Repetitive DNA sequences and evolution of ZZ/ZW sex chromosomes in Characidium (Teleostei: Characiformes). PLoS ONE 2015, 10, e0137231. [Google Scholar] [CrossRef] [PubMed]
- Pokorná, M.; Rens, W.; Rovatsos, M.; Kratochvíl, L. A ZZ/ZW sex chromosome system in the thick-tailed gecko (Underwoodisaurus milii; Squamata: Gekkota: Carphodactylidae), a member of the ancient gecko lineage. Cytogenet. Genome Res. 2014, 142, 190–196. [Google Scholar] [CrossRef]
- Meles, S.; Adega, F.; Guedes-Pinto, H.; Chaves, R. The karyotype and sex chromosomes of Praomys tullbergi (Muridae, Rodentia): A detailed characterization. Micron 2008, 39, 559–568. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Bertollo, L.A.C. Initial steps in XY chromosome differentiation in Hoplias malabaricus and the origin of an X1X2Y sex chromosome system in this fish group. Heredity 2010, 105, 554–561. [Google Scholar] [CrossRef]
- Mank, J.E. Small but mighty: The evolutionary dynamics of W and Y sex chromosomes. Chromosome Res. 2012, 20, 21–33. [Google Scholar] [CrossRef]
- Deakin, J.E.; Potter, S.; O’ Neill, R.; Ruiz-Herrera, A.; Cioffi, M.B.; Eldridge, M.D.B.; Fukui, K.; Marshall Graves, J.A.; Griffin, D.; Grutzner, F. Chromosomics: Bridging the gap between genomes and chromosomes. Genes 2019, 10, 627. [Google Scholar] [CrossRef] [PubMed]
- Birchler, J.A.; Presting, G.G. Retrotransposon insertion targeting: A mechanism for homogenization of centromere sequences on nonhomologous chromosomes. Genes. Dev. 2012, 26, 638–640. [Google Scholar] [CrossRef] [PubMed]
- Alföldi, J.; Di Palma, F.; Grabherr, M.; Williams, C.; Kong, L.; Mauceli, E.; Russell, P.; Lowe, C.B.; Glor, R.E.; Jaffe, J.D. The genome of the green anole lizard and a comparative analysis with birds and mammals. Nature 2011, 477, 587–591. [Google Scholar] [CrossRef]
- Hooper, D.M.; Price, T.D. Chromosomal inversion differences correlate with range overlap in passerine birds. Nat. Ecol. Evol. 2017, 1, 1526–1534. [Google Scholar] [CrossRef]
- Hooper, D.M.; Griffith, S.C.; Price, T.D. Sex chromosome inversions enforce reproductive isolation across an avian hybrid zone. Mol. Ecol. 2018, 28, 1246–1262. [Google Scholar] [CrossRef]
- Fuller, Z.L.; Leonard, C.J.; Young, R.E.; Schaeffer, W.; Phadnis, N. Ancestral polymorphisms explain the role of chromosomal inversions in speciation. PLoS Genet. 2018, 14, e1007526. [Google Scholar] [CrossRef]
- Kirkpatrick, M.; Barton, N. Chromosome inversion, local adaptation and speciation. Genetics 2006, 173, 419–434. [Google Scholar] [CrossRef] [PubMed]
- Wellenreuther, M.; Bernatchez, L. Eco-evolutionary genomics of chromosomal inversions. Trends Ecol. Evol. 2018, 33, 427–440. [Google Scholar] [CrossRef]
- Avise, J.C.; Bowen, B.W.; Lamb, T.; Meylan, A.B.; Bermingham, E. Mitochondrial DNA evolution at a turtle’s place: Evidence for low genetic variability and reduced microevolutionary rate in the Testudines. Mol. Biol. Evol. 1992, 9, 457–473. [Google Scholar]
- Lourenço, J.M.; Glémin, S.; Chiari, Y.; Galtier, N. The determinants of molecular substitution process in turtles. J. Evol. Biol. 2013, 26, 38–50. [Google Scholar] [CrossRef]
- Shaffer, B.H.; Minx, P.; Warren, D.E.; Shedlock, A.M.; Thomson, R.C.; Valenzuela, N.; Abramyan, J.; Amemiya, C.T.; Badenhorst, D.; Biggar, K.K. The western painted turtle genome, a model for evolution of extreme physiological adaptations in a slowly evolving lineage. Genome Biol. 2013, 14, R28. [Google Scholar] [CrossRef] [PubMed]
- Fritz, U. Schildkröten-Hybriden, 2. Halsberger-Schildkröten (Cryptodira). Herpetofauna 1995, 95, 19–34. [Google Scholar]
- Karl, S.A.; Bowen, B.W.; Avise, J.C. Hybridization among the ancient mariners: Characterization of marine turtle hybrids with molecular genetic assays. J. Hered. 1995, 86, 262–268. [Google Scholar] [CrossRef]
- Parham, J.F.; Simison, W.B.; Kozak, K.H.; Feldman, C.R.; Shi, H. New Chinese turtles: Endangered or invalid? A reassessment of two species using mitochondrial DNA, allozyme electrophoresis and known-locality specimens. Anim. Conserv. 2001, 4, 357–367. [Google Scholar] [CrossRef]
- Fritz, U.; Mendau, D. Ein Gattungsbastard zweier südostasiatischer Schildkröten: Cuora amboinensis kamaroma Rummler & Fritz, 1991 x Mauremys annamensis (Siebenrock, 1903). Salamandra 2002, 38, 129–134. [Google Scholar]
- Seminoff, J.A.; Karl, S.A.; Schwartz, T.; Resendiz, A. Hybridization of the green turtle (Chelonia mydas) and hawksbill turtle (Eretmochelys imbricata) in the Pacific Ocean: Indication of an absence of gender bias in the directionality of crosses. Bull. Mar. Sci. 2003, 73, 643–652. [Google Scholar]
- James, M.C.; Martin, K.; Dutton, P.H. Hybridization between a green turtle, Chelonia mydas, and loggerhead turtle, Caretta caretta, and the first record of a green turtle in Atlantic Canada. Can. Field-Nat. 2004, 118, 579–582. [Google Scholar] [CrossRef]
- Lara-Ruiz, P.; Lopez, G.G.; Santos, F.R.; Soares, L.S. Extensive hybridization in hawksbill turtles (Eretmochelys imbricata) nesting in Brazil revealed by mtDNA analyses. Conserv. Genet. 2006, 7, 773–781. [Google Scholar] [CrossRef]
- Stuart, B.L.; Parham, J.F. Recent hybrid origin of three rare Chinese turtles. Conserv. Genet. 2006, 8, 169–175. [Google Scholar] [CrossRef]
- Lee, Y.; Lin, J.W.; Tseng, S.P.; Chen, T.S.; Lin, S.M. Human disturbance as a possible cause of genetic introgression from exotic into native Mauremys turtles. Anim. Conserv. 2019, 22, 556–567. [Google Scholar] [CrossRef]
- Burgtorf, C.; Bünemann, H. A telomere-like satellite (GGGTCAT)n comprises 4% of genomic DNA of Drosophila hydei and is located mainly in centromeric heterochromatin of all large acrocentric autosomes. Gene 1993, 137, 287–291. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).