Genome-Wide Identification and Expression Analysis of the Protease Inhibitor Gene Families in Tomato



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Real Time PCR
2.2. Identification of Tomato PI Genes
2.3. Phylogenetic Analysis
2.4. Chromosomal Location, Gene Structure, and Sequence Alignment
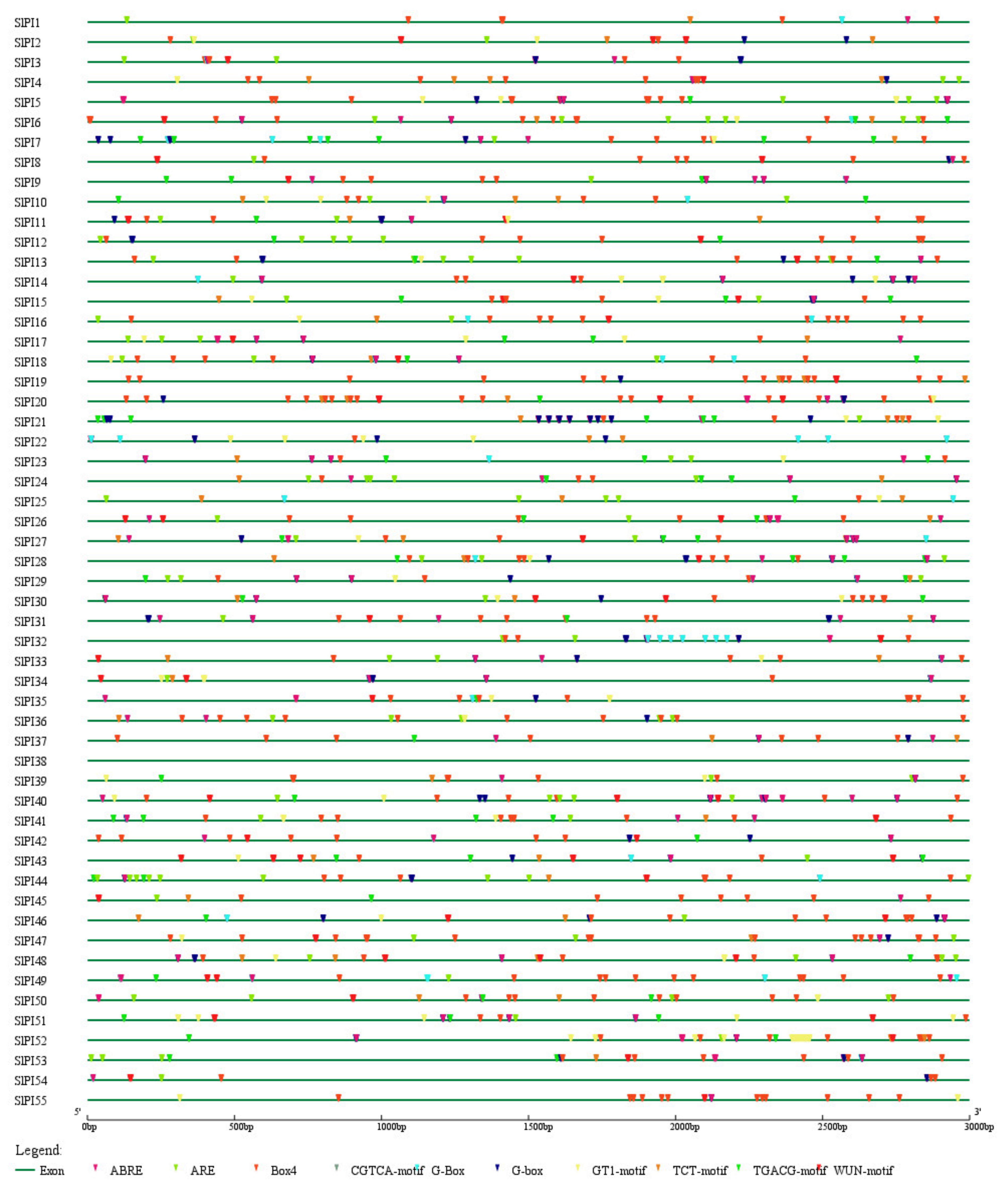
2.5. Cis-Element Prediction for PI Gene Promoter
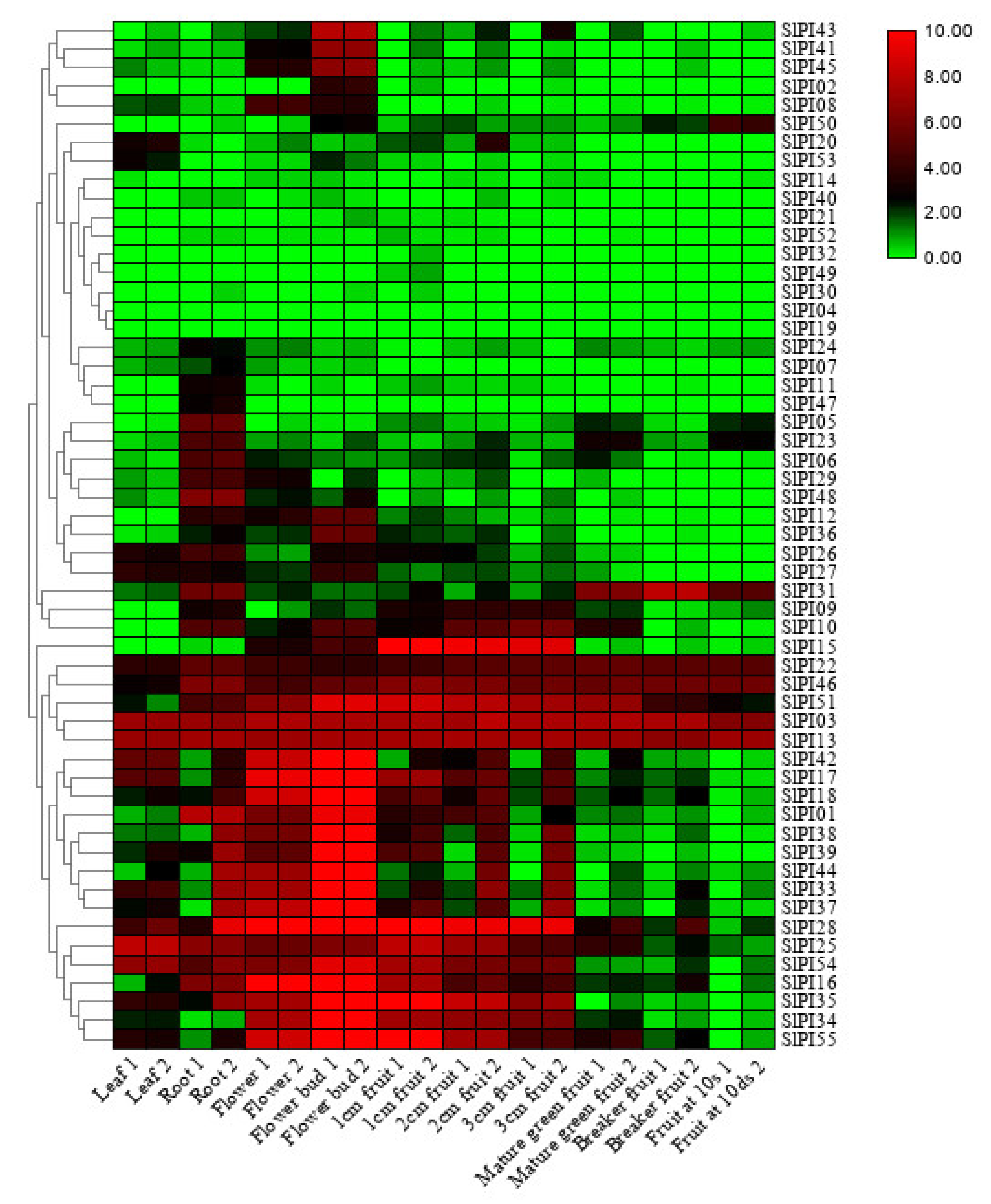
2.6. Tissue Expression Analysis of SlPIs of Tomato
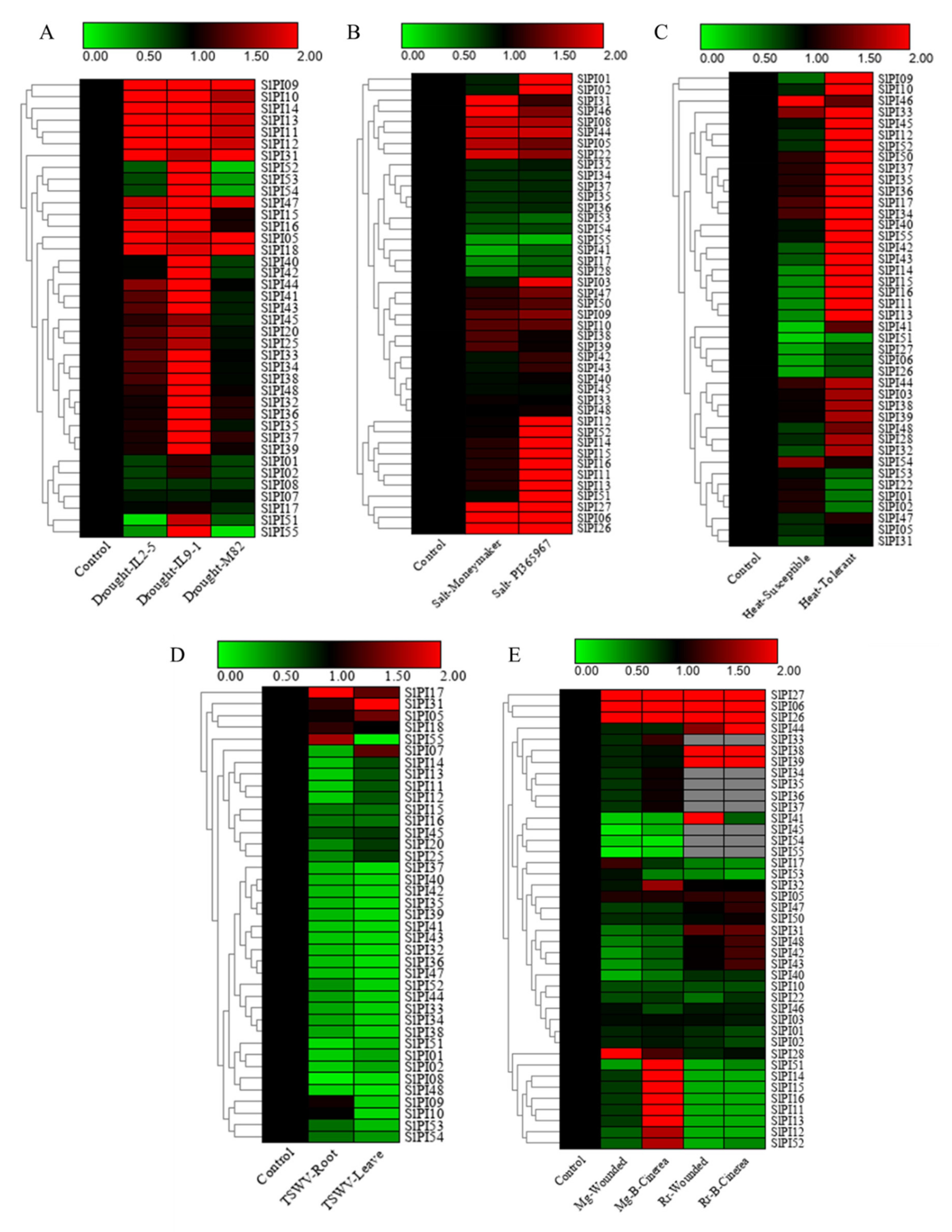
2.7. Expression Pattern of SlPI Genes Under Different Stress Conditions
3. Results
3.1. Identification of PI Genes in Tomato
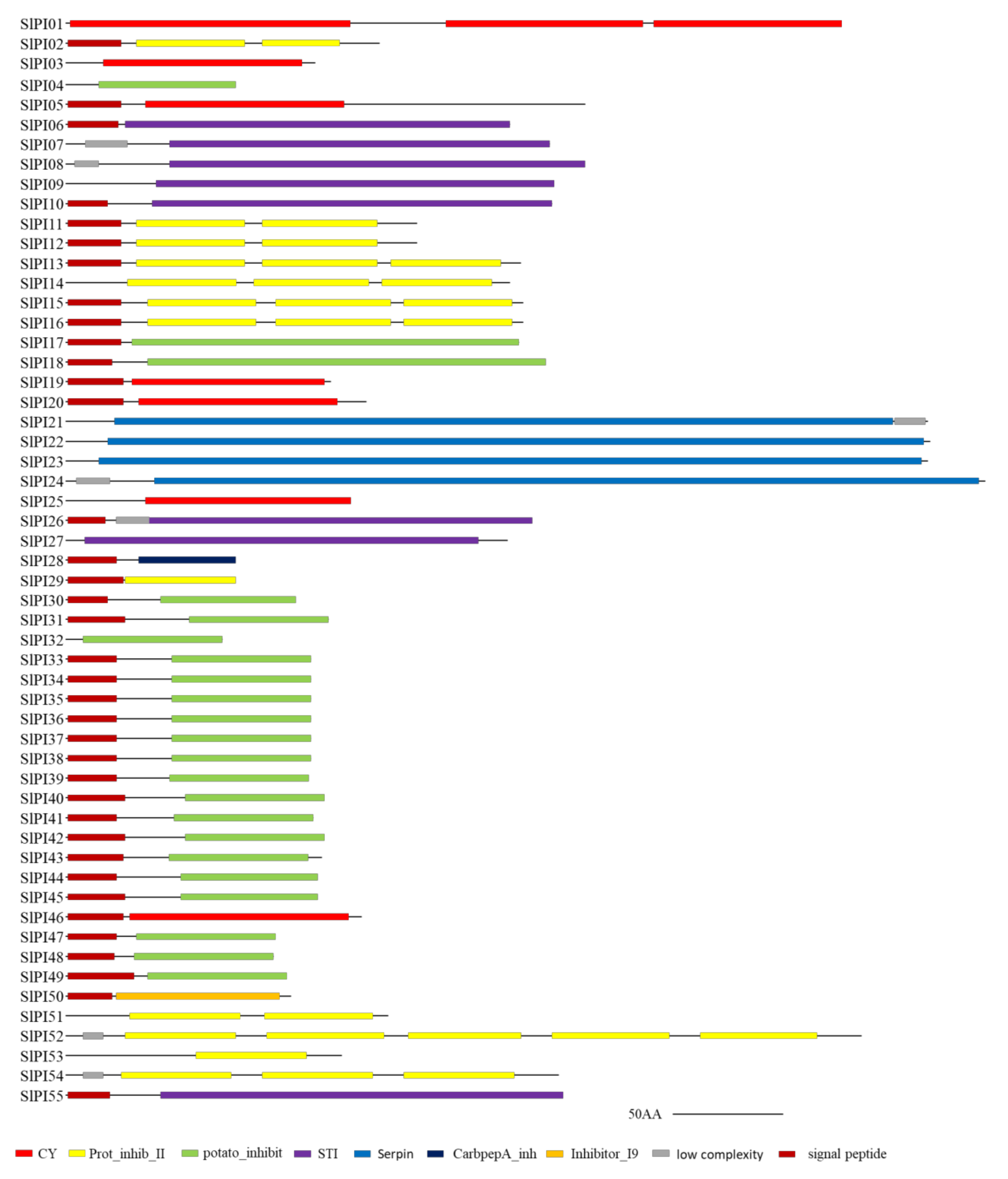
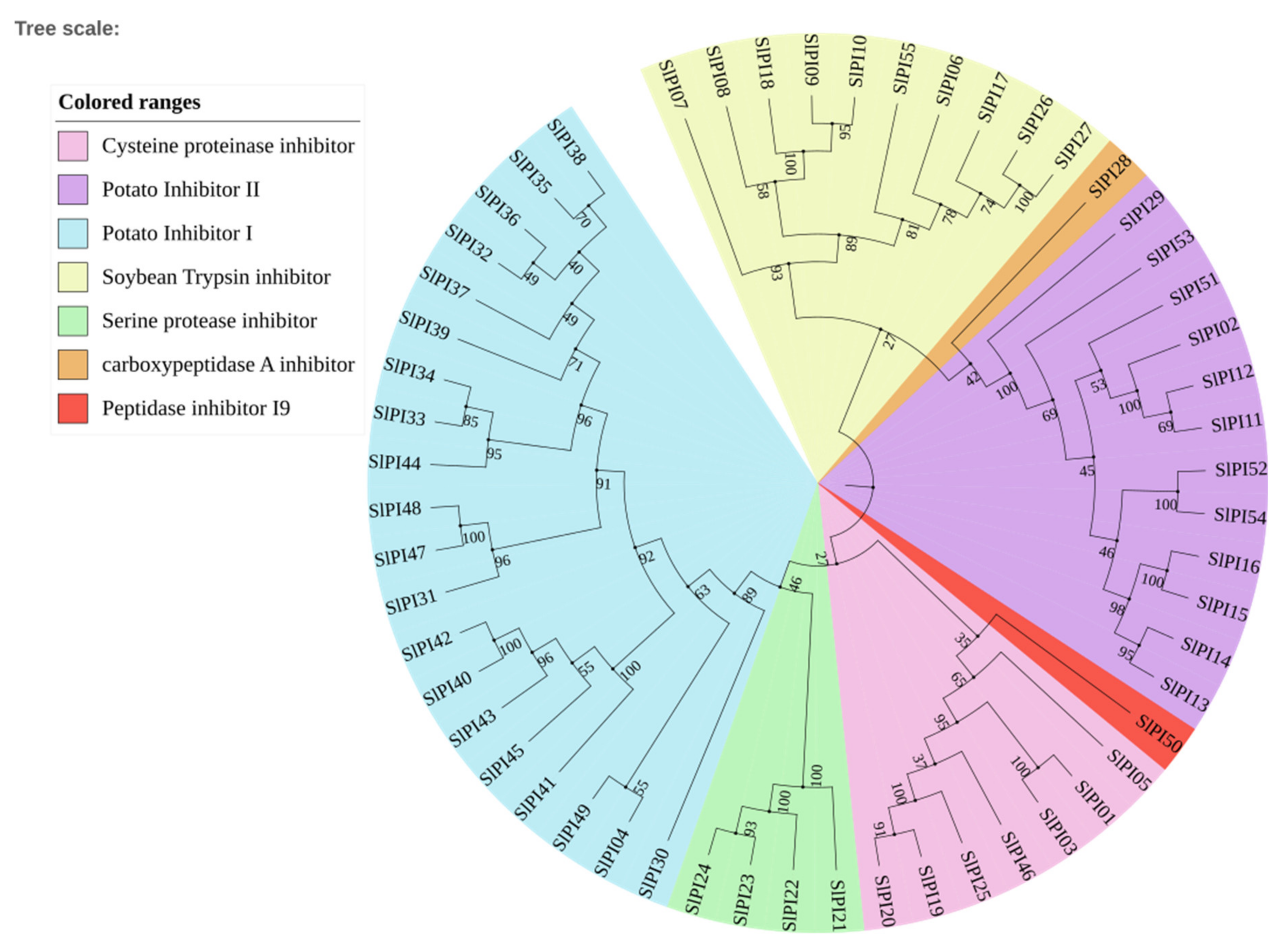
3.2. Conserved Domain and Phylogenetic Analyses of the Tomato PI Families
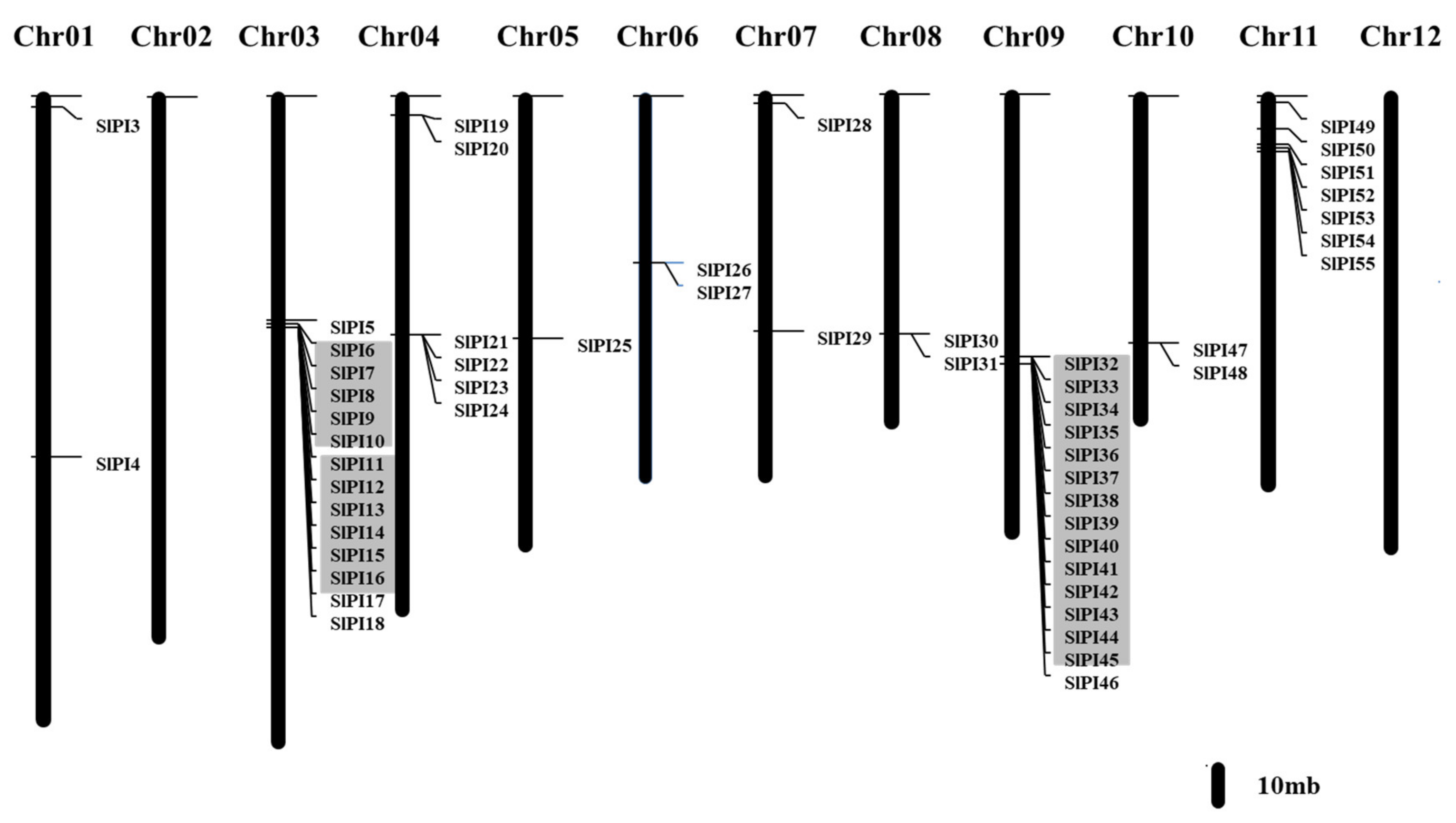
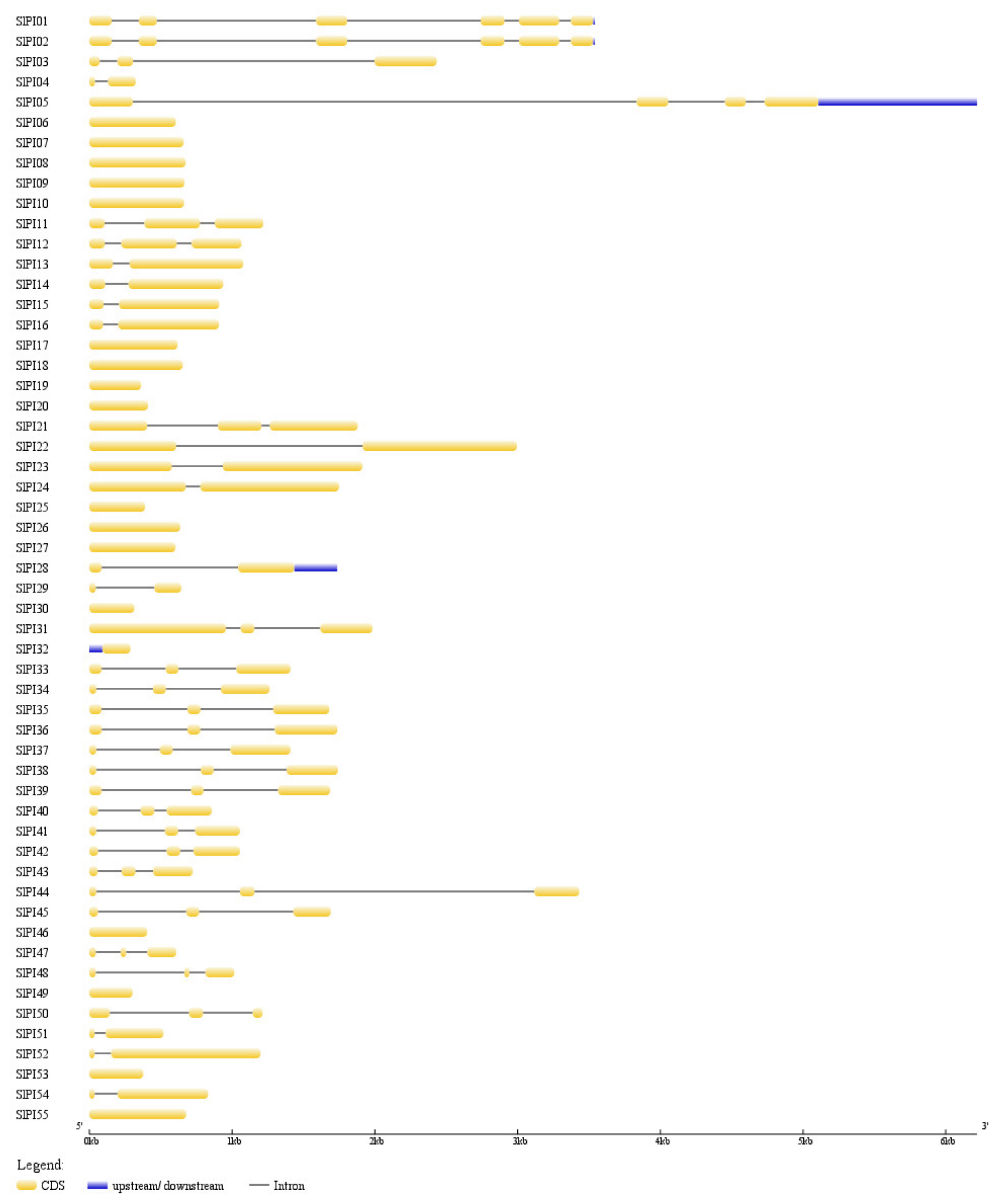
3.3. Chromosomal Localization, Gene Structure, and Duplication of SlPIs
3.4. Cis-Elements in the Promoters of Tomato PI Genes
3.5. Expression Patterns of SlPI Genes Induced by Different Abiotic and Biotic Stresses
3.6. Expression of SlPI Genes in Different Tomato Tissues
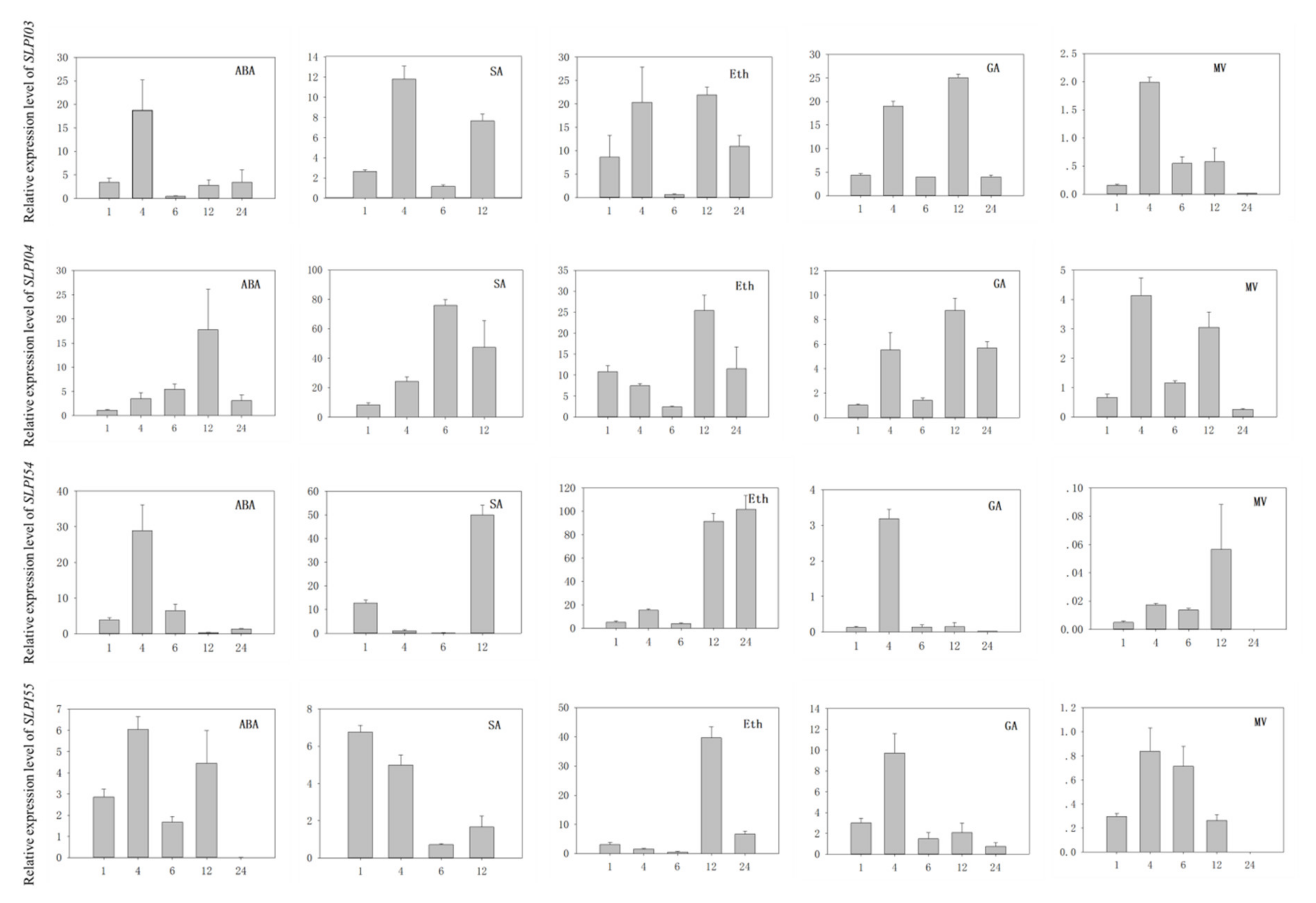
3.7. Analysis of PI Gene Expression in Tomato Under Various Hormone Induction and Oxidation
4. Discussion
4.1. Evolution of the SlPI Gene Families
4.2. Expression of the SlPI Gene Families
4.3. Classification and Function of PI Genes in Tomato
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Brix, K.; Stöcker, W. Proteases: Structure and Function; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Di Cera, E. Serine proteases. IUBMB Life 2009, 61, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Lingaraju, M.; Gowda, L.R. A Kunitz trypsin inhibitor of Entada scandens seeds: Another member with single disulfide bridge. BBA Proteins Proteom. 2008, 1784, 850–855. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.; Aziz, E.; Akhtar, W.; Ilyas, M.; Mahmood, T. Structural and functional characteristics of plant proteinase inhibitor-II (PI-II) family. Biotechnol. Lett. 2017, 39, 647–666. [Google Scholar] [CrossRef] [PubMed]
- De Leo, F.; Volpicella, M.; Licciulli, F.; Liuni, S.; Gallerani, R.; Ceci, L.R. PLANT-PIs: A database for plant protease inhibitors and their genes. Nucl. Acids Res. 2002, 30, 347–348. [Google Scholar] [CrossRef]
- Tamhane, V.A.; Giri, A.P.; Kumar, P.; Gupta, V.S. Spatial and temporal expression patterns of diverse Pin-II proteinase inhibitor genes in Capsicum annuum Linn. Gene 2009, 442, 88–98. [Google Scholar] [CrossRef]
- Lawrence, P.K.; Koundal, K.R. Plant protease inhibitors in control of phytophagous insects. Electron. J. Biotechnol. 2002, 5, 5–6. [Google Scholar] [CrossRef]
- Birk, Y. Plant Protease Inhibitors: Significance in Nutrition, Plant Protection, Cancer Prevention and Genetic Engineering; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Rawlings, N.D.; Tolle, D.P.; Barrett, A.J. Evolutionary families of peptidase inhibitors. Biochem. J. 2004, 378, 705–716. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucl. Acids Res. 2017, 46, D624–D632. [Google Scholar] [CrossRef]
- Ryan, C.A. Protease inhibitors in plants: Genes for improving defenses against insects and pathogens. Annu. Rev. Phytopathol. 1990, 28, 425–449. [Google Scholar] [CrossRef]
- Broadway, R.M. Are insects resistant to plant proteinase inhibitors? J. Insect Physiol. 1995, 41, 107–116. [Google Scholar] [CrossRef]
- Jongsma, M.A.; Bolter, C. The adaptation of insects to plant protease inhibitors. J. Insect Physiol. 1997, 43, 885–895. [Google Scholar] [CrossRef]
- Zavala, J.A.; Patankar, A.G.; Gase, K.; Hui, D.; Baldwin, I.T. Manipulation of endogenous trypsin proteinase inhibitor production in Nicotiana attenuata demonstrates their function as antiherbivore defenses. Plant Physiol. 2004, 134, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Green, T.; Ryan, C.A. Wound-induced proteinase inhibitor in plant leaves: A possible defense mechanism against insects. Science 1972, 175, 776–777. [Google Scholar] [CrossRef] [PubMed]
- Hilder, V.A.; Gatehouse, A.M.; Sheerman, S.E.; Barker, R.F.; Boulter, D. A novel mechanism of insect resistance engineered into tobacco. Nature 1987, 330, 160. [Google Scholar] [CrossRef]
- Chen, P.J.; Senthilkumar, R.; Jane, W.N.; He, Y.; Tian, Z.; Yeh, K.W. Transplastomic Nicotiana benthamiana plants expressing multiple defence genes encoding protease inhibitors and chitinase display broad-spectrum resistance against insects, pathogens and abiotic stresses. Plant Biotech. J. 2014, 12, 503–515. [Google Scholar] [CrossRef]
- Telang, M.A.; Giri, A.P.; Pyati, P.S.; Gupta, V.S.; Tegeder, M.; Franceschi, V.R. Winged bean chymotrypsin inhibitors retard growth of Helicoverpa armigera. Gene 2009, 431, 80–85. [Google Scholar] [CrossRef]
- Abdeen, A.; Virgós, A.; Olivella, E.; Villanueva, J.; Avilés, X.; Gabarra, R.; Prat, S. Multiple insect resistance in transgenic tomato plants over-expressing two families of plant proteinase inhibitors. Plant Mol. Biol. 2005, 57, 189–202. [Google Scholar] [CrossRef]
- Broadway, R.M.; Duffey, S.S. Plant proteinase inhibitors: Mechanism of action and effect on the growth and digestive physiology of larval Heliothis zea and Spodoptera exiqua. J. Insect Physiol. 1986, 32, 827–833. [Google Scholar] [CrossRef]
- Duan, X.; Li, X.; Xue, Q.; Abo-EI-Saad, M.; Xu, D.; Wu, R. Transgenic rice plants harboring an introduced potato proteinase inhibitor II gene are insect resistant. Nat. Biotech. 1996, 14, 494. [Google Scholar] [CrossRef]
- Gatehouse, A.M.; Davison, G.M.; Newell, C.A.; Merryweather, A.; Hamilton, W.D.; Burgess, E.P.; Gilbert, R.J.; Gatehouse, J.A. Transgenic potato plants with enhanced resistance to the tomato moth, Lacanobia oleracea: Growth room trials. Mol. Breed. 1997, 3, 49–63. [Google Scholar] [CrossRef]
- Srinivasan, A.; Giri, A.P.; Harsulkar, A.M.; Gatehouse, J.A.; Gupta, V.S. A Kunitz trypsin inhibitor from chickpea (Cicer arietinum L.) that exerts anti-metabolic effect on podborer (Helicoverpa armigera) larvae. Plant Mol. Biol. 2005, 57, 359–374. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Narvaez, J.; An, G.; Ryan, C. Expression of proteinase inhibitors I and II in transgenic tobacco plants: Effects on natural defense against Manduca sexta larvae. Proc. Natl. Acad. Sci. USA 1989, 86, 9871–9875. [Google Scholar] [CrossRef] [PubMed]
- Urwin, P.E.; Atkinson, H.J.; Waller, D.A.; McPherson, M.J. Engineered oryzacystatin-I expressed in transgenic hairy roots confers resistance to Globodera pallida. Plant J. 1995, 8, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Gatehouse, J.A. Prospects for using proteinase inhibitors to protect transgenic plants against attack by herbivorous insects. Curr. Protein Pept. Sci. 2011, 12, 409–416. [Google Scholar] [CrossRef]
- Cloutier, C.; Jean, C.; Fournier, M.; Yelle, S.; Michaud, D. Adult Colorado potato beetles, Leptinotarsa decemlineata compensate for nutritional stress on oryzacystatin I-transgenic potato plants by hypertrophic behavior and over-production of insensitive proteases. Arch. Insect Biochem. Physiol. 2000, 44, 69–81. [Google Scholar] [CrossRef]
- De Leo, F.; Bonadé-Bottino, M.A.; Ceci, L.R.; Gallerani, R.; Jouanin, L. Opposite effects on Spodoptera littoralis larvae of high expression level of a trypsin proteinase inhibitor in transgenic plants. Plant Physiol. 1998, 118, 997–1004. [Google Scholar] [CrossRef]
- Gosti, F.; Bertauche, N.; Vartanian, N.; Giraudat, J. Abscisic acid-dependent and-independent regulation of gene expression by progressive drought in Arabidopsis thaliana. Mol. Gen. Genet. MGG 1995, 246, 10–18. [Google Scholar] [CrossRef]
- Kim, S.; Hong, Y.-N.; An, C.S.; Lee, K.-W. Expression characteristics of serine proteinase inhibitor II under variable environmental stresses in hot pepper (Capsicum annuum L.). Plant Sci. 2001, 161, 27–33. [Google Scholar] [CrossRef]
- Pernas, M.; Sánchez-Monge, R.; Salcedo, G. Biotic and abiotic stress can induce cystatin expression in chestnut. Febs Lett. 2000, 467, 206–210. [Google Scholar] [CrossRef]
- Gaddour, K.; Vicente-Carbajosa, J.; Lara, P.; Isabel-Lamoneda, I.; Díaz, I.; Carbonero, P. A constitutive cystatin-encoding gene from barley (Icy) responds differentially to abiotic stimuli. Plant Mol. Biol. 2001, 45, 599–608. [Google Scholar] [CrossRef]
- Huang, Y.; Xiao, B.; Xiong, L. Characterization of a stress responsive proteinase inhibitor gene with positive effect in improving drought resistance in rice. Planta 2007, 226, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, T.; Kumar, K.R.R.; Kirti, P.B. Constitutive expression of a trypsin protease inhibitor confers multiple stress tolerance in transgenic tobacco. Plant Cell Physiol. 2009, 50, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-Q.; Zhang, D.; Shen, X.-H. Cloning and characterization of ApCystatin, a plant cystatin gene from Agapanthus praecox ssp. orientalis responds to abiotic stress. Protein Expr. Purif. 2018, 149, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Titarenko, E.; Rojo, E.; Leon, J.; Sanchez-Serrano, J.J. Jasmonic acid-dependent and-independent signaling pathways control wound-induced gene activation in Arabidopsis thaliana. Plant Physiol. 1997, 115, 817–826. [Google Scholar] [CrossRef]
- Koiwa, H.; Bressan, R.A.; Hasegawa, P.M. Regulation of protease inhibitors and plant defense. Trends Plant Sci. 1997, 2, 379–384. [Google Scholar] [CrossRef]
- Morris, K.; Mackerness, S.A.H.; Page, T.; John, C.F.; Murphy, A.M.; Carr, J.P.; Buchanan-Wollaston, V. Salicylic acid has a role in regulating gene expression during leaf senescence. Plant J. 2000, 23, 677–685. [Google Scholar] [CrossRef]
- Li, J.; Su, X.; Wang, Y.; Yang, W.; Pan, Y.; Su, C.; Zhang, X. Genome-wide identification and expression analysis of the BTB domain-containing protein gene family in tomato. Genes Genom. 2018, 40, 1–15. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Bostan, H.; Chiusano, M.L. NexGenEx-Tom: A gene expression platform to investigate the functionalities of the tomato genome. BMC Plant Biol. 2015, 15, 48. [Google Scholar] [CrossRef]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Oberschall, A.; Deák, M.; Török, K.; Sass, L.; Vass, I.; Kovács, I.; Fehér, A.; Dudits, D.; Horváth, G.V. A novel aldose/aldehyde reductase protects transgenic plants against lipid peroxidation under chemical and drought stresses. Plant J. 2000, 24, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [PubMed]
- Joshi, B.N.; Sainani, M.N.; Bastawade, K.B.; Gupta, V.S.; Ranjekar, P.K. Cysteine protease inhibitor from pearl millet: A new class of antifungal protein. Biochem. Biophys. Res. Commun. 1998, 246, 382–387. [Google Scholar] [CrossRef]
- Dunaevsky, Y.E.; Gladysheva, I.P.; Pavlukova, E.B.; Beliakova, G.A.; Gladyshev, D.P.; Papisova, A.I.; Larionova, N.I.; Belozersky, M.A. The anionic protease inhibitor BWI-1 from buckwheat seeds. Kinetic properties and possible biological role. Physiol. Plant. 1997, 101, 483–488. [Google Scholar] [CrossRef]
- Broadway, R.M. Dietary proteinase inhibitors alter complement of midgut proteases. Arch. Insect Biochem. Physiol. 1996, 32, 39–53. [Google Scholar] [CrossRef]
- Singh, S.; Singh, A.; Kumar, S.; Mittal, P.; Singh, I.K. Protease inhibitors: Recent advancement in its usage as a potential biocontrol agent for insect pest management. Insect Sci. 2018. [Google Scholar] [CrossRef]
- Anderson, J.P.; Badruzsaufari, E.; Schenk, P.M.; Manners, J.M.; Desmond, O.J.; Ehlert, C.; Maclean, D.J.; Ebert, P.R.; Kazan, K. Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell 2004, 16, 3460–3479. [Google Scholar] [CrossRef]
- Llorente, F.; Alonso-Blanco, C.; Sánchez-Rodriguez, C.; Jorda, L.; Molina, A. ERECTA receptor-like kinase and heterotrimeric G protein from Arabidopsis are required for resistance to the necrotrophic fungus Plectosphaerella cucumerina. Plant J. 2005, 43, 165–180. [Google Scholar] [CrossRef]
- Sánchez-Vallet, A.; López, G.; Ramos, B.; Delgado-Cerezo, M.; Riviere, M.-P.; Llorente, F.; Fernández, P.V.; Miedes, E.; Estevez, J.M.; Grant, M. Disruption of abscisic acid signaling constitutively activates Arabidopsis resistance to the necrotrophic fungus Plectosphaerella cucumerina. Plant Physiol. 2012, 160, 2109–2124. [Google Scholar] [CrossRef]
- Melville, J.C.; Ryan, C.A. Chymotrypsin Inhibitor I from Potatoes Large Scale Preparation and Characterization of its Subunit Components. J. Biol. Chem. 1972, 247, 3445–3453. [Google Scholar] [PubMed]
- Bryant, J.; Green, T.R.; Gurusaddaiah, T.; Ryan, C.A. Proteinase inhibitor II from potatoes: Isolation and characterization of its protomer components. Biochemistry 1976, 15, 3418–3424. [Google Scholar] [CrossRef]
- Bergey, D.R.; Howe, G.A.; Ryan, C.A. Polypeptide signaling for plant defensive genes exhibits analogies to defense signaling in animals. Proc. Natl. Acad. Sci. USA 1996, 93, 12053–12058. [Google Scholar] [CrossRef] [PubMed]
- Conconi, A.; Smerdon, M.J.; Howe, G.A.; Ryan, C.A. The octadecanoid signalling pathway in plants mediates a response to ultraviolet radiation. Nature 1996, 383, 826. [Google Scholar] [CrossRef] [PubMed]
- Oliva, M.L.V.; Silva, M.C.; Sallai, R.C.; Brito, M.V.; Sampaio, M.U. A novel subclassification for Kunitz proteinase inhibitors from leguminous seeds. Biochimie 2010, 92, 1667–1673. [Google Scholar] [CrossRef] [PubMed]
- Rustgi, S.; Boex-Fontvieille, E.; Reinbothe, C.; von Wettstein, D.; Reinbothe, S. The complex world of plant protease inhibitors: Insights into a Kunitz-type cysteine protease inhibitor of Arabidopsis thaliana. Commun. Integr. Biol. 2018, 11, e1368599. [Google Scholar] [CrossRef]
- Mello, G.C.; Desouza, I.A.; Marangoni, S.; Novello, J.C.; Antunes, E.; Macedo, M.L.R. Oedematogenic activity induced by Kunitz-type inhibitors from Dimorphandra mollis seeds. Toxicon 2006, 47, 150–155. [Google Scholar] [CrossRef]
- Oliva, M.L.V.; Sampaio, M.U. Bauhinia Kunitz-type proteinase inhibitors: Structural characteristics and biological properties. Biol. Chem. 2008, 389, 1007–1013. [Google Scholar]
- Cheung, A.H.; Wong, J.H.; Ng, T. Trypsin-chymotrypsin inhibitors from Vigna mungo seeds. Protein Pept. Lett. 2009, 16, 277–284. [Google Scholar] [CrossRef]
- Fang, E.F.; Wong, J.H.; Ng, T.B. Thermostable Kunitz trypsin inhibitor with cytokine inducing, antitumor and HIV-1 reverse transcriptase inhibitory activities from Korean large black soybeans. J. Biosci. Bioeng. 2010, 109, 211–217. [Google Scholar] [CrossRef]
- Asztalos, B.F.; Schaefer, E.J.; Horvath, K.V.; Cox, C.E.; Skinner, S.; Gerrior, J.; Gorbach, S.L.; Wanke, C. Protease inhibitor-based HAART, HDL, and CHD-risk in HIV-infected patients. Atherosclerosis 2006, 184, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.; Babu, C.R. Purification and biochemical characterization of a serine proteinase inhibitor from Derris trifoliata Lour. seeds: Insight into structural and antimalarial features. Phytochemistry 2009, 70, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Arnaiz, A.; Talavera-Mateo, L.; González-Melendi De León, P.; Martinez, M.; Diaz, I.; Santamaria, M.E. Arabidopsis Kunitz Trypsin Inhibitors in Defence Against Spider Mites. Front. Plant Sci. 2018, 9, 986. [Google Scholar] [CrossRef] [PubMed]
- Confalonieri, M.; Allegro, G.; Balestrazzi, A.; Fogher, C.; Delledonne, M. Regeneration of Populus nigra transgenic plants expressing a Kunitz proteinase inhibitor (KTi 3) gene. Mol. Breed. 1998, 4, 137–145. [Google Scholar] [CrossRef]
- Lee, S.I.; Lee, S.-H.; Koo, J.C.; Chun, H.J.; Lim, C.O.; Mun, J.H.; Song, Y.H.; Cho, M.J. Soybean Kunitz trypsin inhibitor (SKTI) confers resistance to the brown planthopper (Nilaparvata lugens Stål) in transgenic rice. Mol. Breed. 1999, 5, 1–9. [Google Scholar] [CrossRef]
- Gatehouse, A.M.; Norton, E.; Davison, G.M.; Babbé, S.M.; Newell, C.A.; Gatehouse, J.A. Digestive proteolytic activity in larvae of tomato moth, Lacanobia oleracea; effects of plant protease inhibitors in vitro and in vivo. J. Insect Physiol. 1999, 45, 545–558. [Google Scholar] [CrossRef]
- Marchetti, S.; Delledonne, M.; Fogher, C.; Chiaba, C.; Chiesa, F.; Savazzini, F.; Giordano, A. Soybean Kunitz, C-II and PI-IV inhibitor genes confer different levels of insect resistance to tobacco and potato transgenic plants. Theor. Appl. Genet. 2000, 101, 519–526. [Google Scholar] [CrossRef]
- Aceituno-Valenzuela, U.; Covarrubias, M.P.; Aguayo, M.F.; Valenzuela-Riffo, F.; Espinoza, A.; Gaete-Eastman, C.; Herrera, R.; Handford, M.; Norambuena, L. Identification of a type II cystatin in Fragaria chiloensis: A proteinase inhibitor differentially regulated during achene development and in response to biotic stress-related stimuli. Plant Physiol. Biochem. 2018, 129, 158–167. [Google Scholar] [CrossRef]
- Arai, S.; Matsumoto, I.; Emori, Y.; Abe, K. Plant seed cystatins and their target enzymes of endogenous and exogenous origin. J. Agric. Food Chem. 2002, 50, 6612–6617. [Google Scholar] [CrossRef]
- Diaz-Mendoza, M.; Dominguez-Figueroa, J.D.; Velasco-Arroyo, B.; Cambra, I.; Gonzalez-Melendi, P.; Lopez-Gonzalvez, A.; Garcia, A.; Hensel, G.; Kumlehn, J.; Diaz, I.; et al. HvPap-1 C1A Protease and HvCPI-2 Cystatin Contribute to Barley Grain Filling and Germination. Plant Physiol. 2016, 170, 2511–2524. [Google Scholar] [CrossRef]
- Abe, K.; Emori, Y.; Kondo, H.; Suzuki, K.; Arai, S. Molecular-Cloning of a Cysteine Proteinase-Inhibitor of Rice (Oryzacystatin)—Homology with Animal Cystatins and Transient Expression in the Ripening Process of Rice Seeds. J. Biol. Chem. 1987, 262, 16793–16797. [Google Scholar] [PubMed]
- Belenghi, B.; Acconcia, F.; Trovato, M.; Perazzolli, M.; Bocedi, A.; Polticelli, F.; Ascenzi, P.; Delledonne, M. AtCYS1, a cystatin from Arabidopsis thaliana, suppresses hypersensitive cell death. Eur. J. Biochem. 2003, 270, 2593–2604. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, L.; Martinez, M.; Ramessar, K.; Cambra, I.; Castañera, P.; Ortego, F.; Díaz, I. Expression of a barley cystatin gene in maize enhances resistance against phytophagous mites by altering their cysteine-proteases. Plant Cell Rep. 2011, 30, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.M.; Kumar, S.; Cheng, Y.S.; Venkatagiri, S.; Yang, A.H.; Yeh, K.W. Characterization of inhibitory mechanism and antifungal activity between group-1 and group-2 phytocystatins from taro (Colocasia esculenta). FEBS J. 2008, 275, 4980–4989. [Google Scholar] [CrossRef]
- Lilley, C.; Urwin, P.; McPherson, M.; Atkinson, H. Characterization of intestinally active proteinases of cystnematodes. Parasitology 1996, 113, 415–424. [Google Scholar] [CrossRef]
- Urwin, P.E.; Lilley, C.J.; McPherson, M.J.; Atkinson, H.J. Resistance to both cyst and root-knot nematodes conferred by transgenic Arabidopsis expressing a modified plant cystatin. Plant J. 1997, 12, 455–461. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, G.; Li, Z.; Cheng, Y.; Gao, C.; Zeng, L.; Chen, J.; Yan, L.; Sun, X.; Guo, L.; et al. Molecular Cloning, Recombinant Expression and Antifungal Activity of BnCPI, a Cystatin in Ramie (Boehmeria nivea L.). Genes 2017, 8, 265. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, S.; Takano, T. Two cysteine proteinase inhibitors from Arabidopsis thaliana, AtCYSa and AtCYSb, increasing the salt, drought, oxidation and cold tolerance. Plant Mol. Biol. 2008, 68, 131–143. [Google Scholar] [CrossRef]
- Irving, J.A.; Pike, R.N.; Lesk, A.M.; Whisstock, J.C. Phylogeny of the serpin superfamily: Implications of patterns of amino acid conservation for structure and function. Genome Res. 2000, 10, 1845–1864. [Google Scholar] [CrossRef]
- Law, R.H.; Zhang, Q.; McGowan, S.; Buckle, A.M.; Silverman, G.A.; Wong, W.; Rosado, C.J.; Langendorf, C.G.; Pike, R.N.; Bird, P.I. An overview of the serpin superfamily. Genome Biol. 2006, 7, 216. [Google Scholar] [CrossRef][Green Version]
- Gettins, P.G. Serpin structure, mechanism, and function. Chem. Rev. 2002, 102, 4751–4804. [Google Scholar] [CrossRef] [PubMed]
- Silverman, G.A.; Whisstock, J.C.; Bottomley, S.P.; Huntington, J.A.; Kaiserman, D.; Luke, C.J.; Pak, S.C.; Reichhart, J.-M.; Bird, P.I. Serpins flex their muscle I. Putting the clamps on proteolysis in diverse biological systems. J. Biol. Chem. 2010, 285, 24299–24305. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Davydov, O.; Fluhr, R. Plant serpin protease inhibitors: Specificity and duality of function. J. Exp. Bot. 2019, 70, 2077–2085. [Google Scholar] [CrossRef]
- Grosse-Holz, F.M.; van der Hoorn, R.A. Juggling jobs: Roles and mechanisms of multifunctional protease inhibitors in plants. New Phytol. 2016, 210, 794–807. [Google Scholar] [CrossRef]
- Fluhr, R.; Lampl, N.; Roberts, T.H. Serpin protease inhibitors in plant biology. Physiol. Plant. 2012, 145, 95–102. [Google Scholar] [CrossRef]
- Jamal, F.; Pandey, P.K.; Singh, D.; Khan, M. Serine protease inhibitors in plants: Nature’s arsenal crafted for insect predators. Phytochem. Rev. 2013, 12, 1–34. [Google Scholar] [CrossRef]
- Roberts, T.H.; Hejgaard, J. Serpins in plants and green algae. Funct. Integr. Genom. 2008, 8, 1–27. [Google Scholar] [CrossRef]
- Ryan, C.A. The systemin signaling pathway: Differential activation of plant defensive genes. Biochim. Biophys. Acta 2000, 1477, 112–121. [Google Scholar] [CrossRef]
- Hass, G.M.; Ryan, C.A. Carboxypeptidase Inhibitor from Potatoes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1981; Volume 80, pp. 778–791. [Google Scholar]
- Díez-Díaz, M.; Conejero, V.; Rodrigo, I.; Pearce, G.; Ryan, C.A. Isolation and characterization of wound-inducible carboxypeptidase inhibitor from tomato leaves. Phytochemistry 2004, 65, 1919–1924. [Google Scholar] [CrossRef]
- Molesini, B.; Rotino, G.; Dusi, V.; Chignola, R.; Sala, T.; Mennella, G.; Francese, G.; Pandolfini, T. Two metallocarboxypeptidase inhibitors are implicated in tomato fruit development and regulated by the Inner No Outer transcription factor. Plant Sci. 2018, 266, 19–26. [Google Scholar] [CrossRef]
- Harish, B.; Uppuluri, K.B. Microbial serine protease inhibitors and their therapeutic applications. Int. J. Biol. Macromol. 2018, 107, 1373–1387. [Google Scholar] [CrossRef] [PubMed]
- Maier, K.; Müller, H.; Holzer, H. Purification and molecular characterization of two inhibitors of yeast proteinase B. J. Biol. Chem. 1979, 254, 8491–8497. [Google Scholar] [PubMed]
- Dohmae, N.; Takio, K.; Tsumuraya, Y.; Hashimoto, Y. The complete amino acid sequences of two serine proteinase inhibitors from the fruiting bodies of a basidiomycete, Pleurotus ostreatus. Arch. Biochem. Biophys. 1995, 316, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Hohl, M.; Stintzi, A.; Schaller, A. A novel subtilase inhibitor in plants shows structural and functional similarities to protease propeptides. J. Biol. Chem. 2017, 292, 6389–6401. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identifier | Chromosome Location | AA | PIs/MW | Subcellular Localization | |
|---|---|---|---|---|---|
| SlPI01 | Solyc00g071180.2.1 | chr00:14605548..14605198 | 351 | 5.34/40127.40 | cyto: 7, plas: 2, vacu: 2, mito: 1, E.R.: 1, pero: 1 |
| SlPI02 | Solyc00g145170.1.1 | chr00:17251047..17251697 | 142 | 6.48/15408.6 | extr: 8, nucl: 4, chlo: 1, cyto: 1 |
| SlPI03 | Solyc01g009020.2.1 | chr01:3021936..3024369 | 113 | 7.86/12647.4 | chlo: 5, cyto: 3, cysk_nucl: 2.5, nucl: 2, mito: 2, extr: 1 |
| SlPI04 | Solyc01g108380.1.1 | chr01:95700909..95701233 | 77 | 8.45/8071.5 | chlo: 11, extr: 2, nucl: 1 |
| SlPI05 | Solyc03g097270.2.1 | chr03:59587548..59593766 | 235 | 6.39/26249.79 | vacu: 6, extr: 3, chlo: 2, cyto: 1, mito: 1, golg: 1 |
| SlPI06 | Solyc03g098670.1.1 | chr03:60963114..60963719 | 113 | 7.86/12647.4 | extr: 9, vacu: 3, chlo: 1, cyto: 1 |
| SlPI07 | Solyc03g098700.1.1 | chr03:60981171..60981830 | 219 | 5.07/24278.1 | extr: 10, chlo: 3, E.R.: 1 |
| SlPI08 | Solyc03g098710.1.1 | chr03:60989342..60990016 | 224 | 6.50/24726.1 | extr: 5, vacu: 3, E.R.: 2, chlo: 1, cyto: 1, mito: 1, plas: 1 |
| SlPI09 | Solyc03g098780.1.1 | chr03:61061419..61062084 | 221 | 7.48/24500.2 | extr: 7, vacu: 3, cyto: 1, mito: 1, plas: 1, golg: 1 |
| SlPI10 | Solyc03g098790.1.1 | chr03:61068935..61069598 | 220 | 8.84/24188.08 | extr: 7, vacu: 3, chlo: 2, cyto: 1, mito: 1 |
| SlPI11 | Solyc03g020080.2.1 | chr03:61158441..61159659 | 159 | 7.85/17304.9 | extr: 8, chlo: 2, nucl: 2, cyto: 1, vacu: 1v |
| SlPI12 | Solyc03g020070.2.1 | chr03:61164764..61165829 | 159 | 8.12/17144.7 | extr: 11, chlo: 2, cyto: 1 |
| SlPI13 | Solyc03g020060.2.1 | chr03:61171088..61172165 | 206 | 7.31/22018.4 | extr: 6, nucl: 4, chlo: 2, cyto: 1, vacu: 1 |
| SlPI14 | Solyc03g020050.2.1 | chr03:61174085..61175022 | 201 | 4.90/21370.2 | extr: 12, golg: 2 |
| SlPI15 | Solyc03g020040.2.1 | chr03:61180880..61181788 | 207 | 8.53/22682.2 | extr: 11, cyto: 1, vacu: 1, E.R.: 1 |
| SlPI16 | Solyc03g020030.2.1 | chr03:61189044..61189951 | 207 | 8.60/22653.2 | extr: 11, cyto: 1, vacu: 1, E.R.: 1 |
| SlPI17 | Solyc03g020010.1.1 | chr03:61222484..61223101 | 205 | 4.83/22945.4 | extr:7, vacu: 2, golg: 2, chlo: 1, cyto: 1, plas: 1 |
| SlPI18 | Solyc03g019690.1.1 | chr03:61452670..61453323 | 217 | 6.80/23718.9 | extr: 8, chlo: 4, cyto: 1, vacu: 1 |
| SlPI19 | Solyc04g014780.1.1 | chr04:5029049..5029411 | 120 | 6.28/13556.5 | extr: 9, vacu: 2, chlo: 1, cyto: 1, plas: 1 |
| SlPI20 | Solyc04g014790.1.1 | chr04:5030604..5031014 | 136 | 8.56/15121.3 | cyto: 5, extr: 5, chlo: 2, nucl: 1, E.R._vacu: 1 |
| SlPI21 | Solyc04g079370.2.1 | chr04:63894231..63896110 | 390 | 5.25/44417.73 | cyto: 6, cysk: 5, chlo: 2, nucl: 1 |
| SlPI22 | Solyc04g079440.2.1 | chr04:63943215..63945697 | 391 | 5.41/42832.81 | cyto: 2, vacu: 2, plas: 1.5, chlo: 1, nucl: 1, mito: 1, golg: 1 |
| SlPI23 | Solyc04g079470.2.1 | chr04:63948225..63950136 | 390 | 5.53/43200.55 | cyto: 4, vacu: 3, E.R.: 3, nucl: 2, chlo: 1, plas: 1 |
| SlPI24 | Solyc04g079480.2.1 | chr04:63950580..63952328 | 416 | 8.19/46153.11 | chlo: 9, mito: 2, extr: 2, nucl: 1 |
| SlPI25 | Solyc05g054120.1.1 | chr05:64086541..64086930 | 603 | 6.23/68743.2 | chlo: 12, extr: 1, E.R._vacu: 1 |
| SlPI26 | Solyc06g072220.1.1 | chr06:44506328..44506963 | 211 | 9.31/23333.3 | extr: 6, vacu: 4, mito: 2, chlo: 1, golg: 1 |
| SlPI27 | Solyc06g072230.1.1 | chr06:44509088..44509690 | 200 | 8.85/22259.8 | extr: 7, vacu: 3, mito: 2, chlo: 1, cyto: 1 |
| SlPI28 | Solyc07g007250.2.1 | chr07:1989153..1990888 | 77 | 6.49/8353.7 | extr: 13, vacu: 1 |
| SlPI29 | Solyc07g054720.1.1 | chr07:62945748..62946391 | 77 | 7.46/8343.6 | extr: 10, vacu: 3, cyto: 1 |
| SlPI30 | Solyc08g080020.1.1 | chr08:63378916..63379230 | 104 | 4.64/11425.3 | extr: 10, vacu: 2, chlo: 1, cyto: 1 |
| SlPI31 | Solyc08g080630.2.1 | chr08:63880194..63882176 | 119 | 5.33/13160.3 | cyto: 5, extr: 3, E.R.: 2, golg: 2, nucl: 1, vacu: 1 |
| SlPI32 | Solyc09g083430.1.1 | chr09:69089700..69089987 | 71 | 5.17/8066.5 | cyto_nucl: 5.5, cyto: 5, chlo: 2, extr: 2, golg: 2, E.R.: 1 |
| SlPI33 | Solyc09g083440.2.1 | chr09:69093760..69095168 | 111 | 6.58/12359.7 | extr: 10, cyto: 2, chlo: 1, cysk: 1 |
| SlPI34 | Solyc09g084440.2.1 | chr09:69112518..69113780 | 111 | 5.80/12483.7 | extr: 7, cyto: 3, chlo: 2, vacu: 1, E.R.: 1 |
| SlPI35 | Solyc09g084450.2.1 | chr09:69123240..69124920 | 111 | 7.76/12580.8 | extr: 8, vacu: 2, chlo: 1, cyto: 1, mito: 1, E.R.: 1 |
| SlPI36 | Solyc09g084460.2.1 | chr09:69127703..69129440 | 111 | 5.82/12656.9 | extr: 6, vacu: 4, chlo: 3, nucl: 1 |
| SlPI37 | Solyc09g084470.2.1 | chr09:69132578..69133986 | 111 | 4.69/12584.8 | extr: 10, chlo: 1, nucl: 1, cysk: 1, golg: 1 |
| SlPI38 | Solyc09g084480.2.1 | chr09:69137050..69138791 | 111 | 6.58/12557.8 | extr: 6, cyto: 4, vacu: 2, chlo: 1, mito: 1 |
| SlPI39 | Solyc09g084490.2.1 | chr09:69140543..69142228 | 110 | 6.57/12366.6 | extr: 10, cyto: 2, nucl: 1, cysk: 1 |
| SlPI40 | Solyc09g089490.2.1 | chr09:69207347..69208204 | 117 | 6.82/13045.4 | extr: 8, vacu: 3, chlo: 1, cyto: 1, plas: 1 |
| SlPI41 | Solyc09g089500.2.1 | chr09:69211165..69212219 | 112 | 5.76/12668.9 | extr: 9, vacu: 2, chlo: 1, cyto: 1, mito: 1 |
| SlPI42 | Solyc09g089510.2.1 | chr09:69218803..69219857 | 117 | 8.64/13121.6 | extr: 10, chlo: 1, cyto: 1, mito: 1, vacu: 1 |
| SlPI43 | Solyc09g089520.2.1 | chr09:69225548..69226271 | 116 | 9.00/12977.4 | extr: 4, vacu: 4, cyto: 2, chlo: 1, nucl: 1, E.R.: 1, golg: 1 |
| SlPI44 | Solyc09g089530.2.1 | chr09:69229994..69233425 | 114 | 6.82/12715.9 | cyto: 5, extr: 5, chlo: 1, nucl: 1, mito: 1, cysk: 1 |
| SlPI45 | Solyc09g089540.2.1 | chr09:69238075..69239765 | 114 | 5.66/12700.9 | extr: 9, chlo: 1, cyto: 1, mito: 1, vacu: 1, E.R.: 1 |
| SlPI46 | Solyc09g097850.1.1 | chr09:71842796..71843200 | 134 | 9.78/14721.1 | chlo: 9, cyto: 2, extr: 2, nucl: 1 |
| SlPI47 | Solyc10g086090.1.1 | chr10:65046063..65046671 | 95 | 10.11/10682.7 | extr: 8, vacu: 2, chlo: 1, nucl: 1, cyto: 1, plas: 1 |
| SlPI48 | Solyc10g086100.1.1 | chr10:65049349..65050364 | 94 | 9.38/10521.4 | extr: 5, vacu: 5, golg: 2, chlo: 1, nucl: 1 |
| SlPI49 | Solyc11g007050.1.1 | chr11:1528822..1529124 | 100 | 7.59/10955.9 | extr: 9, vacu: 2, cyto: 1, plas: 1, golg: 1 |
| SlPI50 | Solyc11g018590.1.1 | chr11:8761757..8762969 | 102 | 6.83/11488.4 | extr: 11, chlo: 1, cyto: 1, vacu: 1 |
| SlPI51 | Solyc11g020960.1.1 | chr11:12996643..12997161 | 146 | 8.58/15636.8 | extr: 13, vacu: 1 |
| SlPI52 | Solyc11g020990.1.1 | chr11:13167991..13169189 | 360 | 7.55/39279.7 | extr: 9, chlo: 1, cyto: 1, plas: 1, vacu: 1, E.R.: 1 |
| SlPI53 | Solyc11g021020.1.1 | chr11:13195204..13195581 | 125 | 5.20/13985.9 | chlo: 5, mito: 4, extr: 4, golg: 1 |
| SlPI54 | Solyc11g021060.1.1 | chr11:13312553..13313384 | 223 | 5.32/24697.3 | extr: 11, vacu: 2, chlo: 1 |
| SlPI55 | Solyc11g022590.1.1 | chr11:14564635..14565312 | 225 | 9.37/25188.2 | extr: 6, vacu: 3, chlo: 2, cyto: 1, mito: 1, golg: 1 |
| Cis-Element | Number of Genes | Functions of Cis-Elements |
|---|---|---|
| Box 4 | 55 | part of a conserved DNA module involved in light responsiveness |
| CAAT-box | 55 | common cis-acting element in promoter and enhancer regions |
| TATA-box | 55 | core promoter element around -30 of transcription start |
| ABRE | 52 | cis-acting element involved in the abscisic acid responsiveness |
| G-box | 50 | cis-acting regulatory element involved in light responsiveness |
| ARE | 48 | cis-acting regulatory element essential for the anaerobic induction |
| G-Box | 44 | cis-acting regulatory element involved in light responsiveness |
| TCT-motif | 41 | part of a light responsive element |
| WUN-motif | 42 | wound-responsive element |
| GT1-motif | 38 | light responsive element |
| CGTCA-motif | 38 | cis-acting regulatory element involved in the MeJA-responsiveness |
| TGACG-motif | 38 | cis-acting regulatory element involved in the MeJA-responsiveness |
| GATA-motif | 29 | part of a light responsive element |
| I-box | 27 | part of a light responsive element |
| TC-rich repeats | 25 | cis-acting element involved in defense and stress responsiveness |
| chs-CMA1a | 25 | part of a light responsive element |
| TCA-element | 23 | cis-acting element involved in salicylic acid responsiveness |
| MBS | 23 | MYB binding site involved in drought-inducibility |
| LTR | 21 | cis-acting element involved in low-temperature responsiveness |
| TGA-element | 20 | auxin-responsive element |
| CAT-box | 20 | cis-acting regulatory element related to meristem expression |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Y.; Yang, W.; Yan, Q.; Chen, C.; Li, J. Genome-Wide Identification and Expression Analysis of the Protease Inhibitor Gene Families in Tomato. Genes 2020, 11, 1. https://doi.org/10.3390/genes11010001
Fan Y, Yang W, Yan Q, Chen C, Li J. Genome-Wide Identification and Expression Analysis of the Protease Inhibitor Gene Families in Tomato. Genes. 2020; 11(1):1. https://doi.org/10.3390/genes11010001
Chicago/Turabian StyleFan, Yuxuan, Wei Yang, Qingxia Yan, Chunrui Chen, and Jinhua Li. 2020. "Genome-Wide Identification and Expression Analysis of the Protease Inhibitor Gene Families in Tomato" Genes 11, no. 1: 1. https://doi.org/10.3390/genes11010001
APA StyleFan, Y., Yang, W., Yan, Q., Chen, C., & Li, J. (2020). Genome-Wide Identification and Expression Analysis of the Protease Inhibitor Gene Families in Tomato. Genes, 11(1), 1. https://doi.org/10.3390/genes11010001