Danio Rerio as Model Organism for Adenoviral Vector Evaluation

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Zebrafish Handling
2.2. Vectors
2.3. Microinjection
2.4. Incubation
2.5. Imaging
2.6. Immunohistochemistry (IHC)
2.7. Statistical Analysis
3. Results
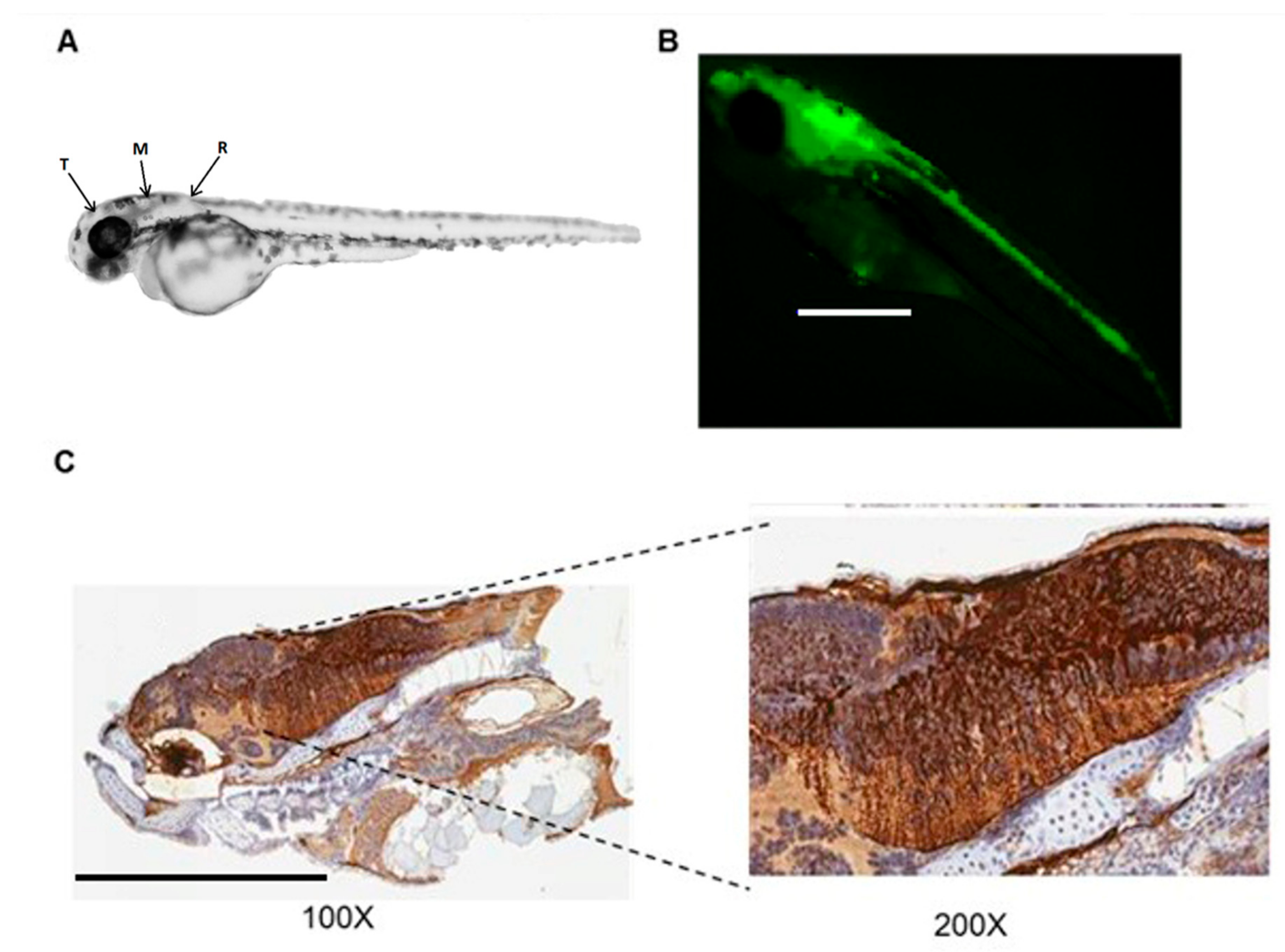
3.1. Adenoviral Vectors Transduce the Brain of Zebrafish Embryos
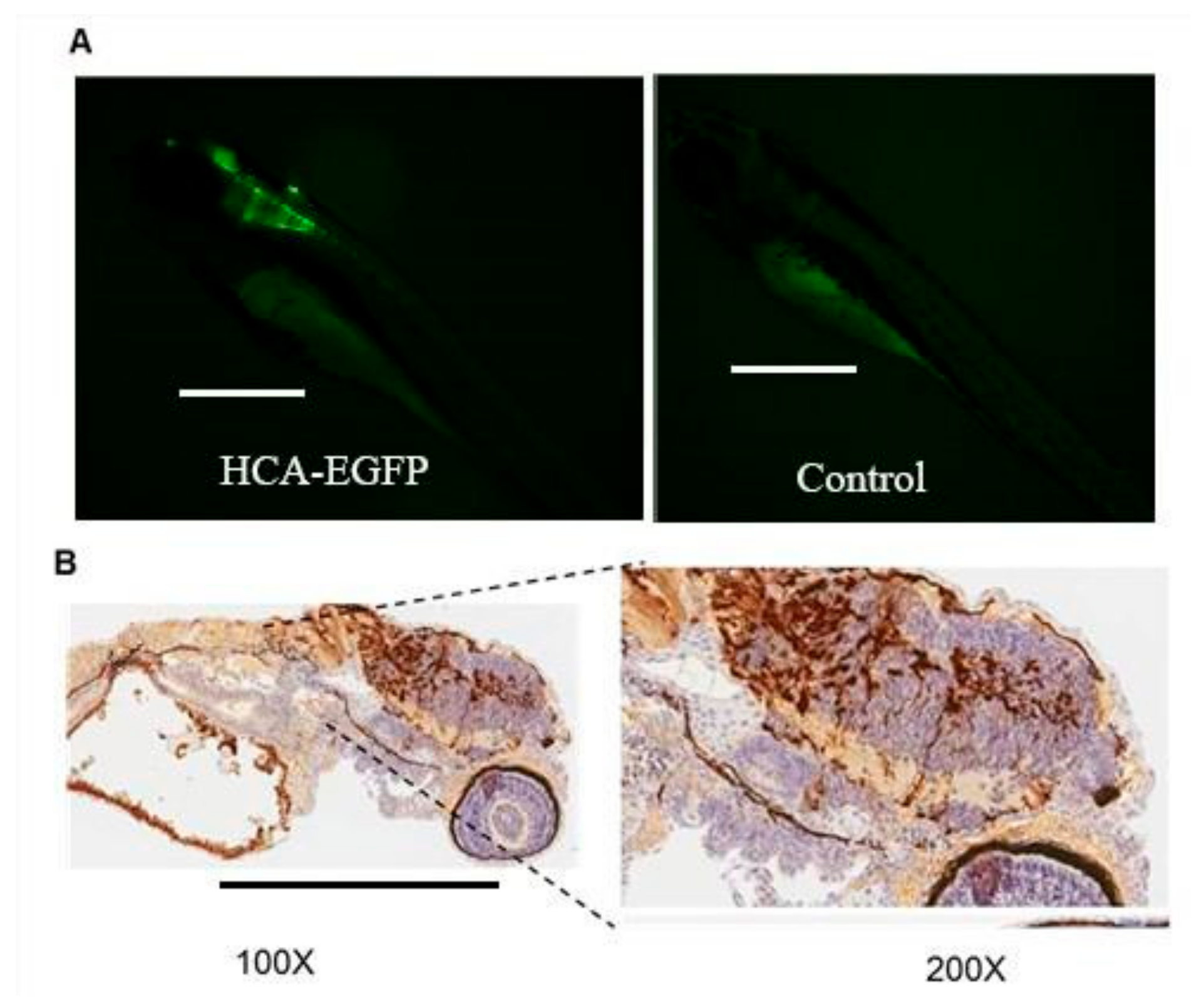
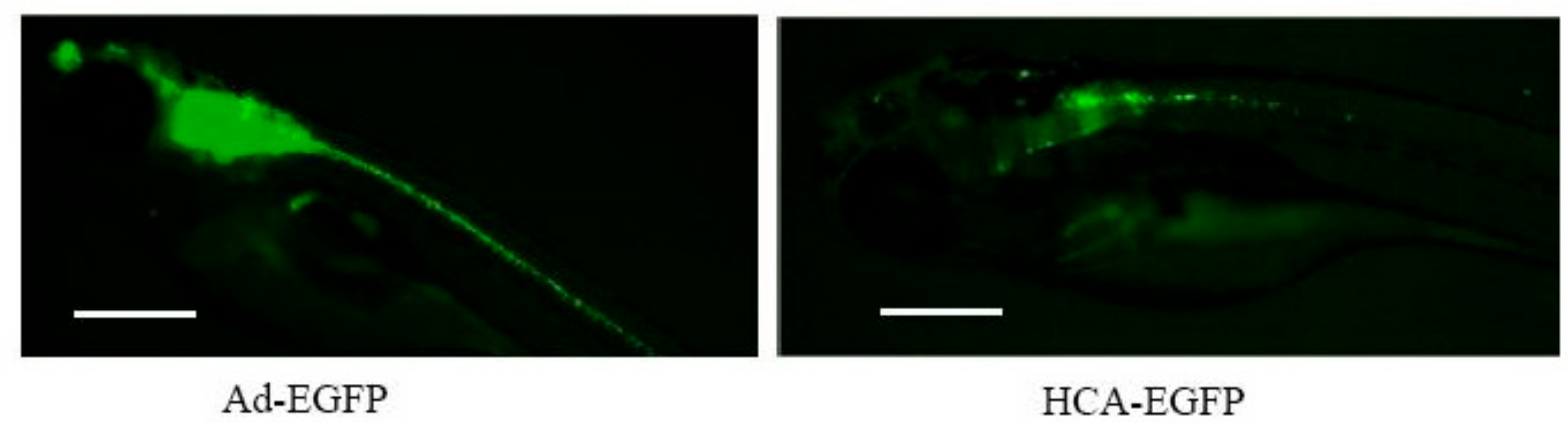
3.2. Hc-Advs Are Better Tolerated Than e1/e3-Deleted Vectors in Zebrafish
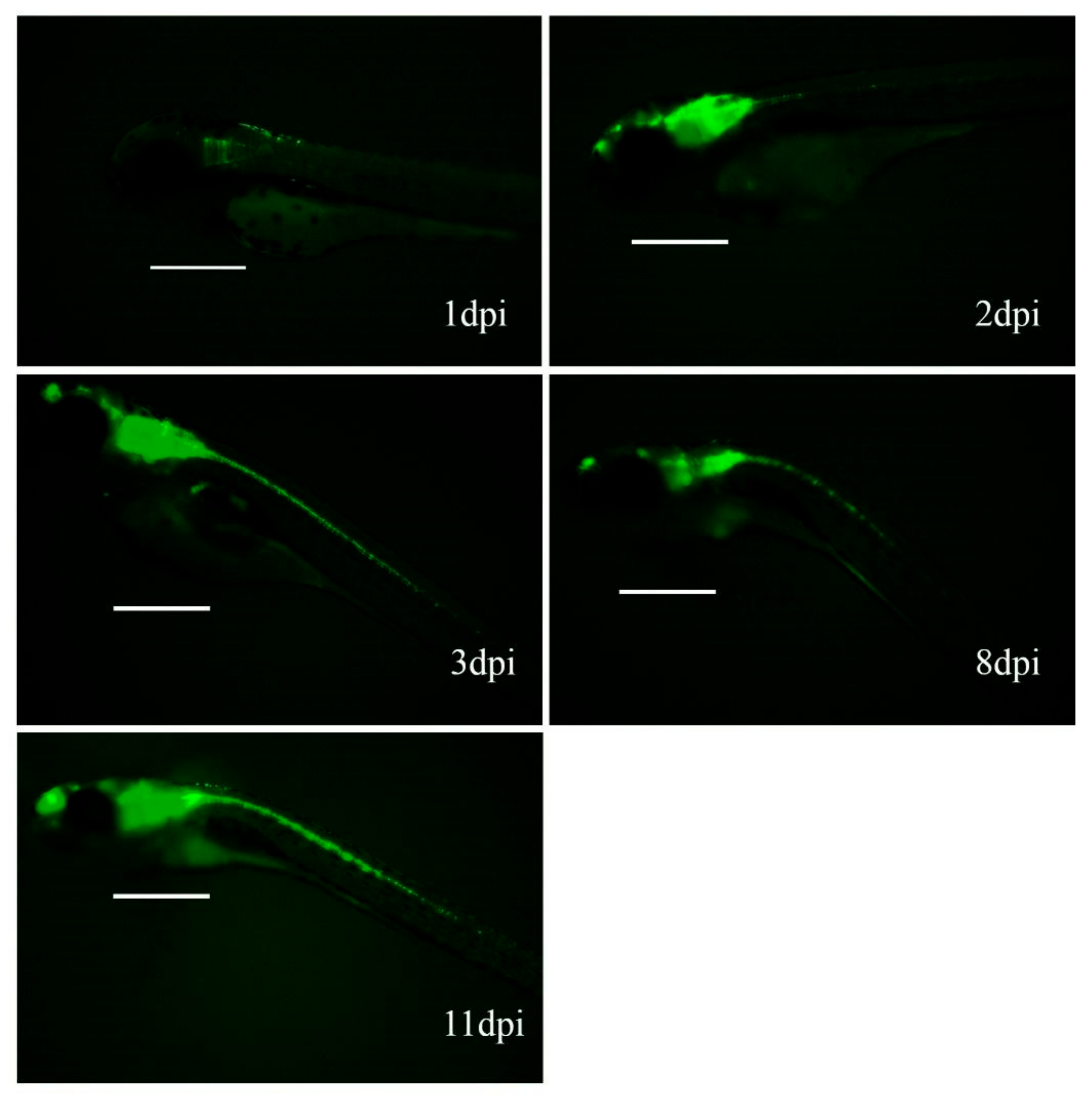
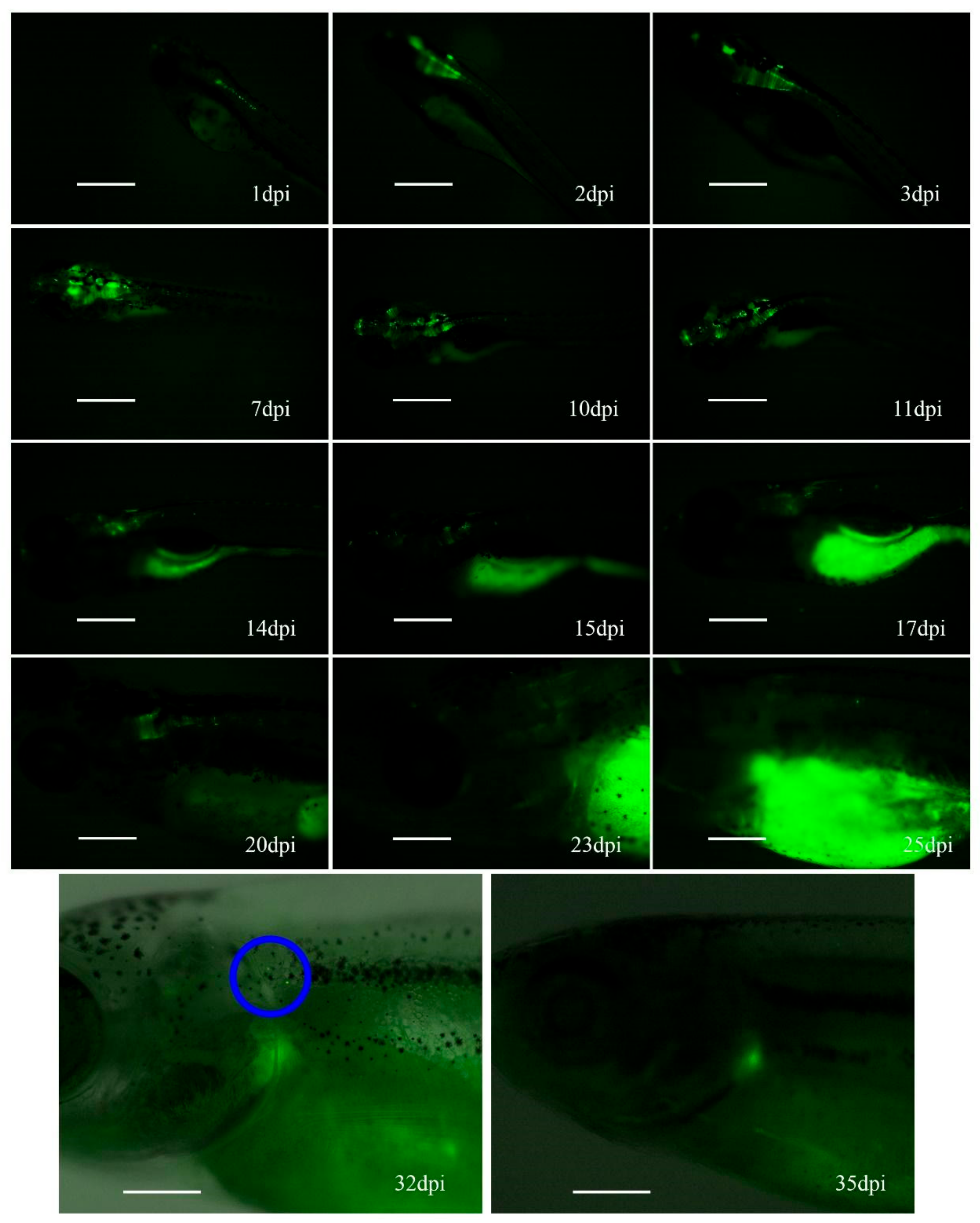
3.3. Hc-Ad Vectors Allow Sustained Expression of Transgenes in Zebrafish
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, L.; Li, F.; Dang, L.; Liang, C.; He, B.; Liu, J. In vivo delivery systems for therapeutic genome editing. Int. J. Mol. Sci. 2016, 17, 626. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K. Viral vectors in gene therapy. Diseases 2018, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Crustal, R. Adenovirus: The first effective in vivo gene delivery vector. Hum. Gene. Ther. 2014, 25, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Zirger, J.M.; Puntel, M.; Bergeron, J.; Wibowo, M.; Moridzadeh, R.; Bondale, N.; Barcia, C.; Kroeger, K.M.; Liu, C.; Castro, M.G.; et al. Immune-mediated loss of transgene expression from virally transduced brain cells is irreversible, mediated by IFNγ, perforin, and TNFα, and due to the elimination of transduced cells. Mol. Ther. 2012, 20, 808–819. [Google Scholar] [CrossRef] [PubMed]
- Cots, D.; Bosch, A.; Chillón, M. Helper dependent adenovirus vectors: Progress and future prospects. Curr. Gene Ther. 2013, 13, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Barcia, C.; Jimenez-Dalmaroni, M.; Kroeger, K.; Puntel, M.; Rapaport, A.; Larocque, D.; King, G. One-year expression from high-capacity adenoviral vectors in the brains of animals with pre-existing anti-adenoviral immunity: Clinical implications. Mol. Gene Ther. 2007, 15, 2154–2163. [Google Scholar] [CrossRef] [PubMed]
- Brunetti-Pierri, N.; Ng, T.; Iannitti, D.; Cioffi, W.; Stapleton, G.; Law, M.; Breinholt, J. Transgene expression up to 7 years in nonhuman primates following hepatic transduction with helper-dependent adenoviral vectors. Hum. Gene Ther. 2013, 24, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Nasiadka, A.; Clark, M.D. Zebrafish breeding in the laboratory environment. ILAR J. 2012, 53, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Dooley, K.; Zon, L.I. Zebrafish: A model system for the study of human diseases. Curr. Opin. Genet. Dev. 2000, 10, 252–256. [Google Scholar] [CrossRef]
- Albain, J.; Zon, L.I. Of fish and men: Using zebrafish to fight human diseases. Trends. Cell Biol. 2013, 23, 584–586. [Google Scholar]
- Howe, K.; Clark, M.; Torroja, C.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Narvaiza, I.; Aparicio, O.; Vera, M.; Razquin, N.; Bortolanza, S.; Prieto, J.; Fortes, P. Effect of adenovirus-mediated RNA interference on endogenous microRNAs in a mouse model of multidrug resistance protein 2 gene silencing. J. Virol. 2006, 80, 12236–12247. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Aparicio, M.; Mauleon, I.; Alzuguren, P.; Bunuales, M.; Gonzalez-Aseguinolaza, G.; San Martín, C.; Prieto, J.; Hernandez-Alcoceba, R. Self-inactivating helper virus for the production of high-capacity adenoviral vectors. Gene Ther. 2011, 18, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Schubert, S.; Keddig, N.; Hanel, R.; Kammann, U. Microinjection into zebrafish embryos (Danio rerio)—A useful tool in aquatic toxicity testing? Environ. Sci. Eur. 2014, 26, 32. [Google Scholar] [CrossRef]
- Yang, Y.; Ertl, H.C.J.; Wilson, J.M. MHC class I-cestricted cytotoxic T lymphocytes to viral antigens destroy hepatocytes in mice infected with E1-deleted recombinant adenoviruses. Immunity 1994, 1, 433–442. [Google Scholar] [CrossRef]
- Lam, S.H.; Chua, H.L.; Gong, Z.; Lam, T.J.; Sin, Y.M. Development and maturation of the immune system in zebrafish, Danio rerio: A gene expression profiling, in situ hybridization and immunological study. Dev. Comp. Immunol. 2004, 28, 9–28. [Google Scholar] [CrossRef]
- Zhang, B.; Shimada, Y.; Hirota, T.; Ariyoshi, M.; Kuroyanagi, J.; Nishimura, Y.; Tanaka, T. Novel immunologic tolerance of human cancer cell xenotransplants in zebrafish. Transl. Res. 2016, 170, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Parichy, D.M.; Elizondo, M.R.; Mills, M.G.; Gordon, T.N.; Engeszer, E. Normal table of post-embryonic zebrafish development: Staging by externally visible anatomy of the living fish. Dev. Dyn. 2009, 238, 2975–3015. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiments | 48 h Post-Injection (hpi) | 72 hpi | ||
|---|---|---|---|---|
| E1/E3-Deleted Vector (Ad-EGFP) | HC-AdV (HCA-EGFP) | Ad-EGFP | HCA-EGFP | |
| Positive | 45 (45.92%) | 29 (31.87%) | 55 (56.12%) | 35 (38.46%) |
| Negative | 38 (38.77%) | 52 (57.14%) | 26 (26.53%) | 44 (48.35%) |
| Dead * | 15 (15.30%) | 10 (10.99%) | 17 (17.35%) | 12 (13.19%) |
| Total | 98 | 91 | 98 | 91 |
| Vector | Mortality | ||
|---|---|---|---|
| 11 dpi | |||
| Alive | Dead | Percentage | |
| Ad-EGFP | 0 | 55 | 100% |
| HCA-EGFP | 12 | 21 | 64% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gulías, P.; Guerra-Varela, J.; Gonzalez-Aparicio, M.; Ricobaraza, A.; Vales, A.; Gonzalez-Aseguinolaza, G.; Hernandez-Alcoceba, R.; Sánchez, L. Danio Rerio as Model Organism for Adenoviral Vector Evaluation. Genes 2019, 10, 1053. https://doi.org/10.3390/genes10121053
Gulías P, Guerra-Varela J, Gonzalez-Aparicio M, Ricobaraza A, Vales A, Gonzalez-Aseguinolaza G, Hernandez-Alcoceba R, Sánchez L. Danio Rerio as Model Organism for Adenoviral Vector Evaluation. Genes. 2019; 10(12):1053. https://doi.org/10.3390/genes10121053
Chicago/Turabian StyleGulías, Paola, Jorge Guerra-Varela, Manuela Gonzalez-Aparicio, Ana Ricobaraza, Africa Vales, Gloria Gonzalez-Aseguinolaza, Rubén Hernandez-Alcoceba, and Laura Sánchez. 2019. "Danio Rerio as Model Organism for Adenoviral Vector Evaluation" Genes 10, no. 12: 1053. https://doi.org/10.3390/genes10121053
APA StyleGulías, P., Guerra-Varela, J., Gonzalez-Aparicio, M., Ricobaraza, A., Vales, A., Gonzalez-Aseguinolaza, G., Hernandez-Alcoceba, R., & Sánchez, L. (2019). Danio Rerio as Model Organism for Adenoviral Vector Evaluation. Genes, 10(12), 1053. https://doi.org/10.3390/genes10121053