Morphological and Genetic Mechanisms Underlying Awn Development in Monocotyledonous Grasses


Abstract
1. Introduction
2. Morphological and Histological Characteristics of the Awn
2.1. Morphological Features of the Grass Inflorescence
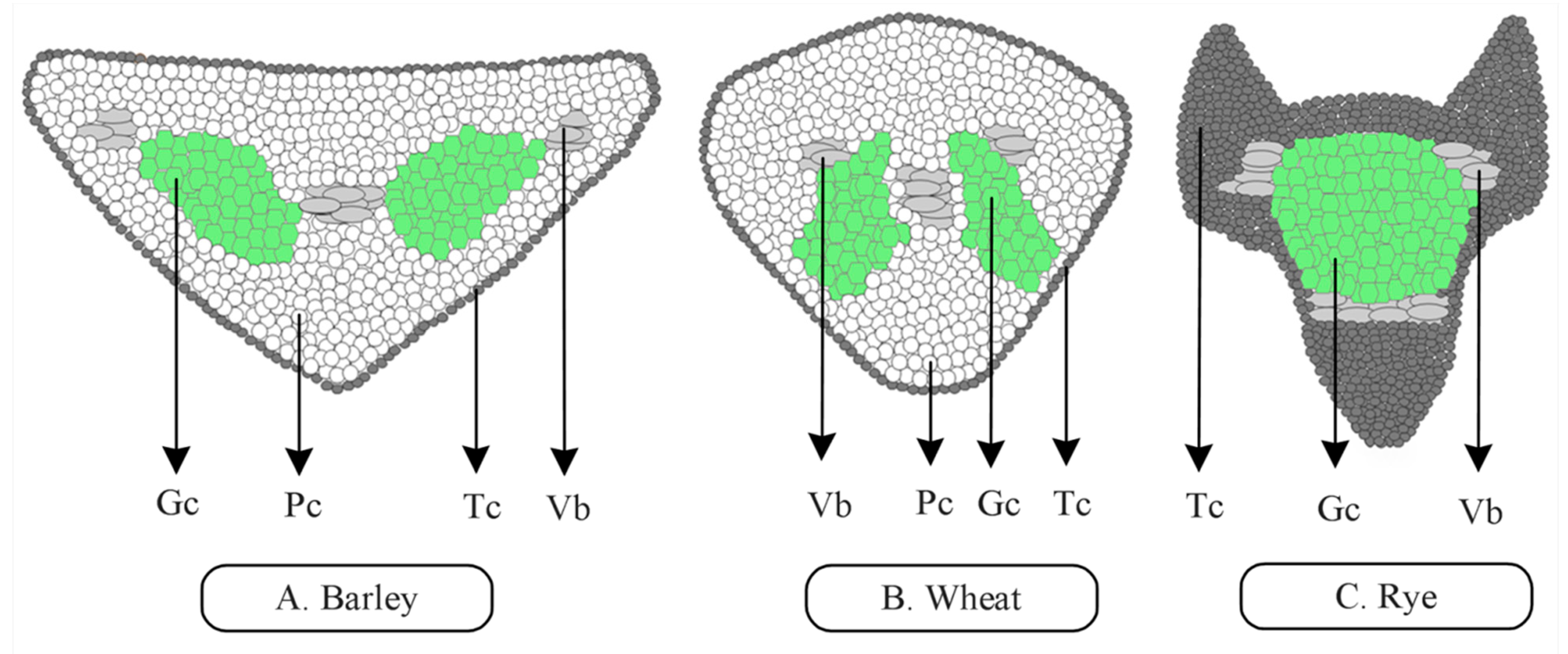
2.2. Awn Anatomy and Histology
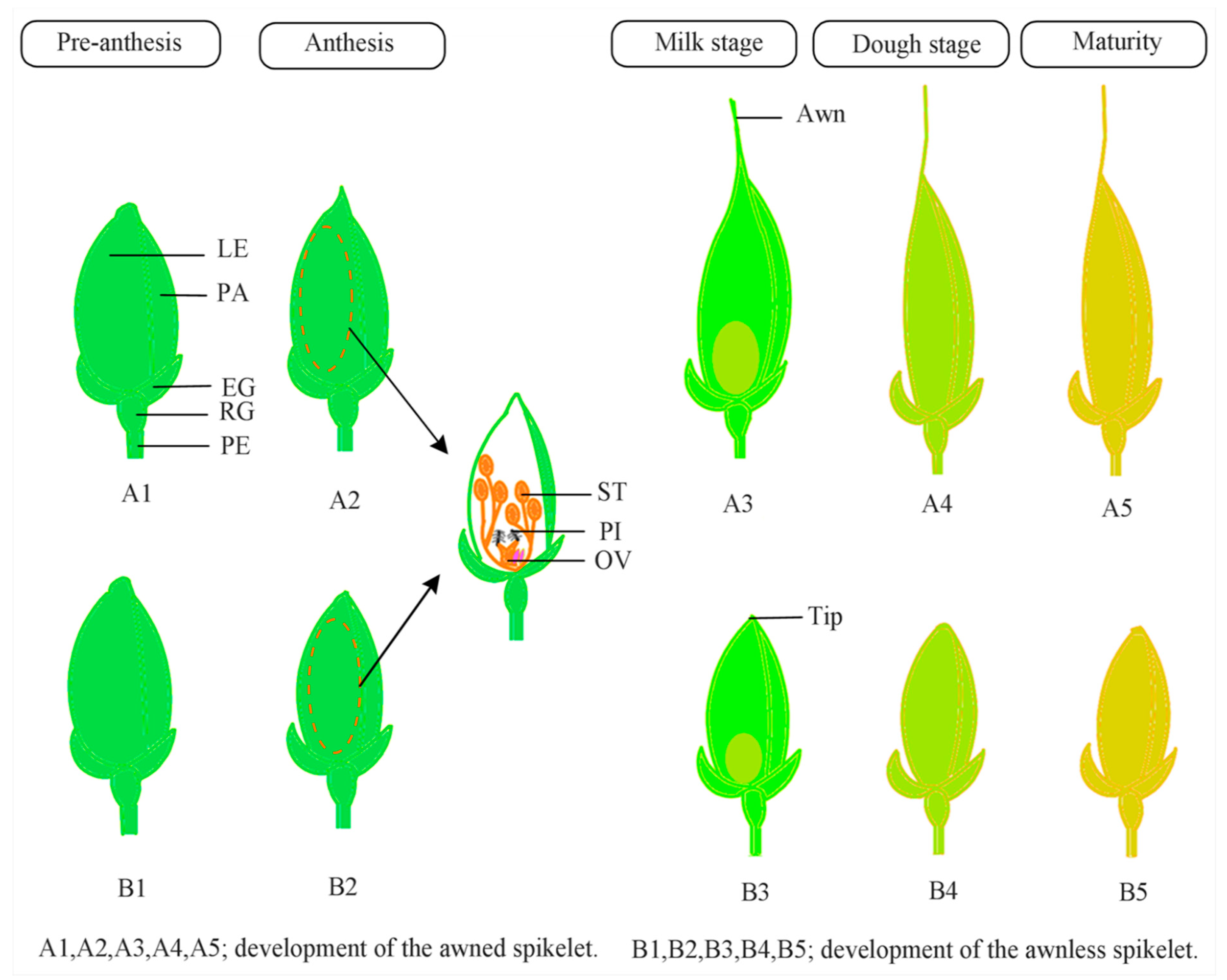
3. Awn Development Pathways
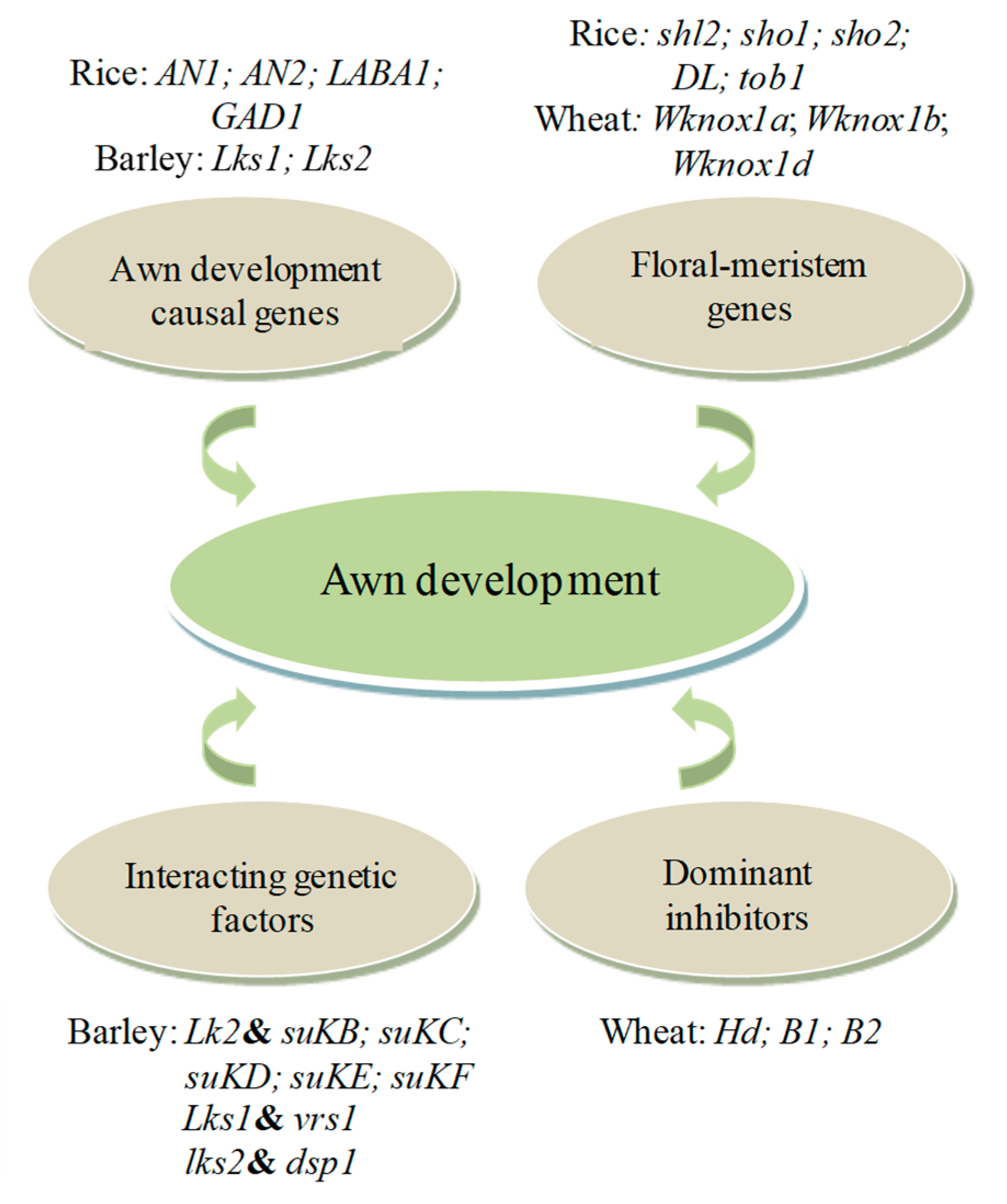
4. Genetic Basis of Awn Development in Grasses
4.1. Awn Development and Recent Genetic and Genomic Advances
4.2. Molecular Mechanisms Underlying Awn Development in Grasses
4.2.1. Mechanisms of Awn Development Causal Genes
4.2.2. Functions of Floral Meristem Genes
4.2.3. The Role of Interacting Genetic Factors
4.2.4. Regulation of Awn Formation and Length by Dominant Inhibitors
5. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yuo, T.; Yamashita, Y.; Kanamori, H.; Matsumoto, T.; Lundqvist, U.; Sato, K.; Ichii, M.; Jobling, S.A.; Taketa, S. A Short Internodes (SHI) family transcription factor gene regulates awn elongation and pistil morphology in barley. J. Exp. Bot. 2012, 63, 5223–5232. [Google Scholar] [CrossRef] [PubMed]
- Bommert, P.; Whipple, C. Grass inflorescence architecture and meristem determinacy. Semin. Cell Dev. Biol. 2017, 79, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Houston, K.; McKim, S.M.; Comadran, J.; Bonar, N.; Druka, I.; Uzrek, N.; Cirillo, E.; Guzy-Wrobelska, J.; Collins, N.C.; Halpin, C. Variation in the interaction between alleles of Hvapetala2 and microrna172 determines the density of grains on the barley inflorescence. Proc. Natl. Acad. Sci. USA 2013, 110, 16675–16680. [Google Scholar] [CrossRef] [PubMed]
- Motzo, R.; Giunta, F. Awnedness affects grain yield and kernel weight in near-isogenic lines of durum wheat. Aust. J. Agric. Res. 2002, 53, 1285–1293. [Google Scholar] [CrossRef]
- Liller, C.B.; Walla, A.; Boer, M.P.; Hedley, P.; Macaulay, M.; Effgen, S.; von Korff, M.; van Esse, G.W.; Koornneef, M. Fine mapping of a major QTL for awn length in barley using a multiparent mapping population. Theor. Appl. Genet. 2017, 130, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Komeda, N.; Asano, K.; Uehara, K.; Gamuyao, R.; Angeles-Shim, R.B.; Nagai, K.; Doi, K.; Wang, D.R.; Yasui, H. Convergent loss of awn in two cultivated rice species Oryza sativa and Oryza glaberrima is caused by mutations in different loci. G3 Genes Genomes Genet. 2015, 5, 2267–2274. [Google Scholar] [CrossRef] [PubMed]
- Elbaum, R.; Zaltzman, L.; Burgert, I.; Fratzl, P. The role of wheat awns in the seed dispersal unit. Science 2007, 316, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Maydup, M.L.; Antonietta, M.; Graciano, C.; Guiamet, J.J.; Tambussi, E.A. The contribution of the awns of bread wheat (Triticum aestivum L.) to grain filling: Responses to water deficit and the effects of awns on ear temperature and hydraulic conductance. Field Crop. Res. 2014, 167, 102–111. [Google Scholar] [CrossRef]
- Rebetzke, G.; Bonnett, D.; Reynolds, M. Awns reduce grain number to increase grain size and harvestable yield in irrigated and rainfed spring wheat. J. Exp. Bot. 2016, 67, 2573–2586. [Google Scholar] [CrossRef] [PubMed]
- McKevith, B. Nutritional aspects of cereals. Nutr. Bull. 2004, 29, 111–142. [Google Scholar] [CrossRef]
- Park, T.I.; Han, O.K.; Seo, J.H.; Choi, J.S.; Park, K.H.; Kim, J.G. New barley cultivars with improved morphological characteristics for whole crop forage in Korea. J. KoreanSoc. Grassl. Forage Sci. 2008, 28, 193–202. [Google Scholar]
- Luo, J.; Liu, H.; Zhou, T.; Gu, B.; Huang, X.; Shangguan, Y.; Zhu, J.; Li, Y.; Zhao, Y.; Wang, Y. An-1 encodes a basic helix-loop-helix protein that regulates awn development, grain size, and grain number in rice. Plant Cell 2013, 25, 3360–3376. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, S. Identification of new alleles of awnness genes, An7 and An8, in rice using Oryza glumaepatula introgression lines. Rice Genet. Newsl. 2003, 20, 19–20. [Google Scholar]
- Matsushita, S. Mapping of genes for awn in rice using Oryza meridionalis introgression lines. Rice Genet. Newsl. 2003, 20, 17–18. [Google Scholar]
- Hua, L.; Wang, D.R.; Tan, L.; Fu, Y.; Liu, F.; Xiao, L.; Zhu, Z.; Fu, Q.; Sun, X.; Gu, P.; et al. Laba1, a domestication gene associated with long, barbed awns in wild rice. Plant Cell 2015, 27, 1875–1888. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.; Zhou, T.; Luo, J.; Liu, H.; Wang, Y.; Shangguan, Y.; Zhu, J.; Li, Y.; Sang, T.; Wang, Z.; et al. An-2 encodes a cytokinin synthesis enzyme that regulates awn length and grain production in rice. Mol. Plant 2015, 8, 1635–1650. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Hua, L.; Zhu, Z.; Tan, L.; Zhao, X.; Zhang, W.; Liu, F.; Fu, Y.; Cai, H.; Sun, X. GAD1 encodes a secreted peptide that regulates grain number, grain length, and awn development in rice domestication. Plant Cell 2016, 28, 2453–2463. [Google Scholar] [CrossRef] [PubMed]
- Nishijima, R.; Ikeda, T.M.; Takumi, S. Genetic mapping reveals a dominant awn-inhibiting gene related to differentiation of the variety anathera in the wild diploid wheat Aegilops Tauschii. Genetica 2018, 146, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, M.; Iehisa, J.C.; Ohno, R.; Kimura, T.; Enoki, H.; Nishimura, S.; Nasuda, S.; Takumi, S. Three dominant awnless genes in common wheat: Fine mapping, interaction and contribution to diversity in awn shape and length. PLoS ONE 2017, 12, e0176148. [Google Scholar] [CrossRef]
- Youssef, H.M.; Eggert, K.; Koppolu, R.; Alqudah, A.M.; Poursarebani, N.; Fazeli, A.; Sakuma, S.; Tagiri, A.; Rutten, T.; Govind, G.; et al. VRS2 regulates hormone-mediated inflorescence patterning in barley. Nat. Genet. 2016, 49, 157. [Google Scholar] [CrossRef]
- Toriba, T.; Hirano, H.Y. The Drooping Leaf and Osettin2genes promote awn development in rice. Plant J. 2014, 77, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Allred, K.W. Describing the grass inflorescence. J. Range Manag. 1982, 35, 672–675. [Google Scholar] [CrossRef]
- Bonnett, O.T. Inflorescences of Maize, Wheat, Rye, Barley, and Oats: Their Initiation and Development; University of Illinois College of Agriculture: Urbana, IL, USA, 1966. [Google Scholar]
- Kinney, M.S.; Columbus, J.T.; Friar, E.A. Unisexual flower, spikelet, and inflorescence development in monoecious/dioecious Bouteloua dimorpha (Poaceae, Chloridoideae). Am. J. Bot. 2008, 95, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Schnurbusch, T. Costs and benefits of awns. J. Exp. Bot. 2016, 67, 2533. [Google Scholar] [CrossRef] [PubMed]
- Tabacchi, M.; Mantegazza, R.; Spada, A.; Ferrero, A. Morphological traits and molecular markers for classification of Echinochloa species from Italian rice fields. Weed Sci. 2006, 54, 1086–1093. [Google Scholar] [CrossRef]
- Sánchez-Ken, J.G. Lachnagrostis filiformis (Poaceae: Pooideae: Poeae: Agrostidinae) in Mexico: Known distribution and suppression of lemma awn development in terminal spikelets. Phytotaxa 2018, 350, 223–234. [Google Scholar] [CrossRef]
- Stubbendieck, J.L.; Hatch, S.L.; Landholt, L. North American Wild Landplants, a Field Guide, 2nd ed.; University of Nebraska Press: Lincoln, NE, USA, 2011. [Google Scholar]
- Kellogg, E.A. Evolutionary history of the grasses. Plant Physiol. 2001, 125, 1198–1205. [Google Scholar] [CrossRef]
- Wiebe, G.A.; Reid, D.A. Classification of Barley Varieties Grown in the United States and Canada in 1958; US Department of Agriculture: Washington, DC, USA, 1961. [Google Scholar]
- Bayles, B.B.; Clark, J.A. Classification of Wheat Varieties Grown in the United States in 1949; US Department of Agriculture: Washington, DC, USA, 1954. [Google Scholar]
- Brown, A.J. Nomenclature, variation and hybridisation in Rough Blown-grass (Poaceae: Lachnagrostis). Muelleria 2015, 33, 85–95. [Google Scholar]
- Li, X.F.; Bin, D.; Hong-Gang, W. Awn anatomy of common wheat (Triticum aestivumL.) and its relatives. Caryologia 2010, 63, 391–397. [Google Scholar] [CrossRef][Green Version]
- Peleg, Z.; Saranga, Y.; Fahima, T.; Aharoni, A.; Elbaum, R. Genetic control over silica deposition in wheat awns. Physiol.Plant. 2010, 140, 10–20. [Google Scholar] [CrossRef]
- Głazowska, S.; Murozuka, E.; Persson, D.P.; Castro, P.H.; Schjoerring, J.K. Silicon affects seed development and leaf macrohair formation in Brachypodium distachyon. Physiol. Plant. 2017, 163, 231–246. [Google Scholar] [CrossRef]
- Strömberg, C.A.; Di Stilio, V.S.; Song, Z. Functions of phytoliths in vascular plants: An evolutionary perspective. Funct. Ecol. 2016, 30, 1286–1297. [Google Scholar] [CrossRef]
- Gol, L.; Tomé, F.; von Korff, M. Floral transitions in wheat and barley: Interactions between photoperiod, abiotic stresses, and nutrient status. J. Exp. Bot. 2017, 68, 1399–1410. [Google Scholar] [CrossRef] [PubMed]
- Bonnett, O. The development of the wheat spike. J. Agric. Res. 1936, 53, 445–451. [Google Scholar]
- Ponzi, R.; Pizzolongo, G. Cytological and anatomical observations on the awn and lemma of wheat (Triticum aestivum L. cv. Ofanto). Plant Biosyst. 2005, 139, 345–348. [Google Scholar] [CrossRef]
- Bonnett, O.T. Hood and supernumerary spike development in barley. J. Agric. Res. 1938, 57, 371–377. [Google Scholar]
- Bremer-Reinders, D. The early stages of development in the rye spike. Acta Bot. Neerl. 1958, 7, 223–232. [Google Scholar] [CrossRef]
- Alqudah, A.M.; Schnurbusch, T. Awn primordium to tipping is the most decisive developmental phase for spikelet survival in barley. Funct. Plant Biol. 2014, 41, 424–436. [Google Scholar] [CrossRef]
- De Wet, J.T. Leaf anatomy and phylogeny in the tribe Danthonieae. Am. J. Bot. 1956, 43, 175–182. [Google Scholar] [CrossRef]
- Dong, R.; Dong, D.; Luo, D.; Zhou, Q.; Chai, X.; Zhang, J.; Xie, W.; Liu, W.; Dong, Y.; Wang, Y.; et al. Transcriptome analyses reveal candidate pod shattering-associated genes involved in the pod ventral sutures of common vetch (Vicia sativaL.). Front. Plant Sci. 2017, 8, 649. [Google Scholar] [CrossRef]
- Gu, X.Y.; Kianian, S.F.; Foley, M.E. Phenotypic selection for dormancy introduced a set of adaptive haplotypes from weedy into cultivated rice. Genetics 2005, 171, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Magwa, R.A.; Zhao, H.; Yao, W.; Xie, W.; Yang, L.; Xing, Y.; Bai, X. Genomewide association analysis for awn length linked to the seed shattering geneqSH1 in rice. J. Genet. 2016, 95, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Glémin, S.; Bataillon, T. A comparative view of the evolution of grasses under domestication. New Phytol. 2009, 183, 273–290. [Google Scholar] [CrossRef] [PubMed]
- Bergelson, J.; Roux, F. Towards identifying genes underlying ecologically relevant traits in Arabidopsis thaliana. Nat. Rev. Genet. 2010, 11, 867. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Morishima, H. QTL clusters reflect character associations in wild and cultivated rice. Theor. Appl. Genet. 2002, 104, 1217–1228. [Google Scholar] [CrossRef] [PubMed]
- Masoudi, B.; Mardi, M.; Hervan, E.M.; Bihamta, M.R.; Naghavi, M.R.; Nakhoda, B.; Bakhshi, B.; Ahmadi, M.; Tabatabaei, M.T.; Firouzabadi, M.H.D. Study of QTLs linked to awn length and their relationships with chloroplasts under control and saline environments in bread wheat. Genes Genomics 2019, 41, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.X.; Kim, D.M.; Ju, H.G.; Ahn, S.N. Mapping quantitative trait loci for awnness and yield component traits in isogenic lines derived from an Oryza sativa/O. rufipogon cross. J. Crop. Sci. Biotechnol. 2009, 12, 9–16. [Google Scholar] [CrossRef]
- Hart, G.; Schertz, K.; Peng, Y.; Syed, N. Genetic mapping of Sorghum bicolor (L.) Moench QTLs that control variation in tillering and other morphological characters. Theor. Appl. Genet. 2001, 103, 1232–1242. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, C.; Wu, D.; Qiao, F.; Li, W.; Duan, L.; Wang, K.; Xiao, Y.; Chen, G.; Liu, Q. High-throughput phenotyping and QTL mapping reveals the genetic architecture of maize plant growth. Plant Physiol. 2017, 173, 1554–1564. [Google Scholar] [CrossRef]
- Takamure, I. Determination of loci for Mi (Minute grain) and Lk-f (Fusayoshi long grain) on the 3rd linkage group in rice. Jpn. J. Breed. 1991, 41, 322–323. [Google Scholar]
- Sato, S.; Ishikawa, S.; Shimono, M.; Shinjyo, C. Genetic studies on an awnness gene Au-4 on chromosome 8 in rice Oryza sativa L. Jpn. J. Breed. 1996, 46, 321–327. [Google Scholar] [CrossRef]
- Xiong, L.; Liu, K.; Dai, X.; Xu, C.; Zhang, Q. Identification of genetic factors controlling domestication-related traits of rice using an F2 population of a cross between Oryza sativa and O. rufipogon. Theor. Appl. Genet. 1999, 98, 243–251. [Google Scholar] [CrossRef]
- Kubo, T.; Takano-Kai, N.; Yoshimura, A. RFLP mapping of genes for long kernel and awn on chromosome 3 in rice. Rice Genet. Newsl. 2001, 18, 26–28. [Google Scholar]
- Kishimoto, N.; Shimosaka, E.; Matsuura, S.; Saito, A. A current RFLP linkage map of rice: Alignment of the molecular map with the classical map. RGN 1992, 9, 118–124. [Google Scholar]
- Franckowiak, J.D.; Lundqvist, U. Descriptions of barley genetic stocks for 2011. Barley Genet. Newsl. 2011, 41, 54–202. [Google Scholar]
- Müller, K.J.; Romano, N.; Gerstner, O.; Garcia-Marotot, F.; Pozzi, C.; Salamini, F.; Rohde, W. The barley Hooded mutation caused by a duplication in a homeobox gene intron. Nature 1995, 374, 727. [Google Scholar] [CrossRef]
- Komatsuda, T.; Pourkheirandish, M.; He, C.; Azhaguvel, P.; Kanamori, H.; Perovic, D.; Stein, N.; Graner, A.; Wicker, T.; Tagiri, A. Six-rowed barley originated from a mutation in a homeodomain-leucine zipper I-class homeobox gene. Proc. Natl. Acad. Sci. USA 2007, 104, 1424–1429. [Google Scholar] [CrossRef]
- Roig, C.; Pozzi, C.; Santi, L.; Müller, J.; Wang, Y.; Stile, M.R.; Rossini, L.; Stanca, M.; Salamini, F. Genetics of barley hooded suppression. Genetics 2004, 167, 439–448. [Google Scholar] [CrossRef]
- Satoh, N.; Itoh, J.I.; Nagato, Y. The Shootless2 and Shootless1genes are involved in both initiation and maintenance of the shoot apical meristem through regulating the number of indeterminate cells. Genetics 2003, 164, 335–346. [Google Scholar]
- Itoh, J.I.; Kitano, H.; Matsuoka, M.; Nagato, Y. Shoot organization genes regulate shoot apical meristem organization and the pattern of leaf primordium initiation in rice. Plant Cell 2000, 12, 2161–2174. [Google Scholar] [CrossRef]
- Tanaka, W.; Toriba, T.; Ohmori, Y.; Yoshida, A.; Kawai, A.; Mayama-Tsuchida, T.; Ichikawa, H.; Mitsuda, N.; Ohme-Takagi, M.; Hirano, H.Y. The YABBY gene TONGARI-BOUSHI1 is involved in lateral organ development and maintenance of meristem organization in the rice spikelet. Plant Cell 2012, 24, 80–95. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Nagasawa, N.; Kawasaki, S.; Matsuoka, M.; Nagato, Y.; Hirano, H.Y. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell 2004, 16, 500–509. [Google Scholar] [CrossRef]
- Sameri, M.; Takeda, K.; Komatsuda, T. Quantitative trait loci controlling agronomic traits in recombinant inbred lines from a cross of oriental-and occidental-type barley cultivars. Breed. Sci. 2006, 56, 243–252. [Google Scholar] [CrossRef][Green Version]
- Yagil, E.; Stebbins, G. The morphogenetic effects of the hooded gene in barley II. Cytological and environmental factors affecting gene expression. Genetics 1969, 62, 307. [Google Scholar]
- Taketa, S.; Yuo, T.; Sakurai, Y.; Miyake, S.; Ichii, M. Molecular mapping of the short awn 2 (lks2) and dense spike 1 (dsp1) genes on barley chromosome 7H. Breed. Sci. 2011, 61, 80–85. [Google Scholar] [CrossRef]
- Sourdille, P.; Cadalen, T.; Gay, G.; Gill, B.; Bernard, M. Molecular and physical mapping of genes affecting awning in wheat. Plant Breed. 2002, 121, 320–324. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Locus Name | Locus Symbol | Associated Makers | Physical Position a | Chr. b | Phenotypic Function | Reference |
|---|---|---|---|---|---|---|---|
| Oryza sativa | AWN1 | AN1 | M6298, RM6285 | 5.88 Mb (region) | 4 | Awn formation | Luo et al. [12] |
| O. sativa | AWN2 | AN2 | FM5, FM6 | 56 kb (region) | 4 | Awn elongation | Gu et al. [16] |
| O. sativa | AWN3 | AN3 | - | - | 3 | Awn elongation | Takamure et al. [54] |
| O. sativa | AWN4 | AN4 | RT1-8a*T65 RT1-8c*T65 RT8-10*T65 | - - - | 8 | Awn elongation, inflorescence development | Sato et al. [55] |
| O. sativa | AWN5 (An5) | AN5 | RZ740 RG122 | 21.2 cM 24.5 cM | 4 | Awn elongation | Xiong et al. [56] |
| O. sativa | An-10 [An5(t)] | An10 | C1677 G1316 | 2.4 cM 2.2 cM | 3 | Awn elongation | Kubo et al. [57] |
| Oryza glumaepatula | An-11(An7) | An11 | RM3419 RM289 | 3.6 cM 3.6 cM | 5 | Awn elongation | Matsushita et al. [13] |
| O. glumaepatula | An-12(An8) | An12 | RM261 RM1359 | 12.2 cM 7.1 cM | 4 | Awn elongation | Matsushita et al. [13] |
| Oryzameridionalis | An-13(An6) | An13 | RM3496 | 11.7 cM | 8 | Awn elongation | Matsushita et al. [14] |
| O. meridionalis | An-14(An9) | An14 | RM8111 RM8051 | 3.0 cM 3.6 cM | 1 | Awn elongation | Matsushita et al. [14] |
| O. meridionalis | An-15(An10) | An15 | RM237 RM265 | 15.1 cM 3.0 cM | 1 | Awn elongation | Matsushita et al. [14] |
| O. sativa | Chromogen for Anthocyanin | C | XNpb165-1 XNpb200 | 16.4 cM 17.1 cM | 6 | Lemma, palea, and awn color | Kishimoto et al. [58] |
| O. sativa | Long and Barbed Awn 1 | LABA1 (RAE1) | M3, RM17242 | 34.6 kb (region) | 4 | Awn elongation | Hua et al. [15] |
| O. sativa | Grain Number, Grain Length, and Awn Development 1 | GAD1 (RAE2) | MX14, MX16 | 6 kb (region) | 8 | Awn elongation, grain number | Jin et al. [17] |
| O. glabberima | Regulator of Awn Elongation 3 | RAE3 | 6KG28331, 6KG30196 | 1.9 Mb (region) | 6 | Awn elongation | Furuta et al. [6] |
| Triticum aestivum | Hooded | Hd | WABM229716, WABM117400, WABM23373517.1 | 0.9 cM | 4AS | Awn suppression | Yoshioka et al. [19] |
| T. aestivum | Tipped 1 | B1 | Xgwm291 | 1.3 cM | 5AL | Awn suppression | Yoshioka et al. [19] |
| T. aestivum | Tipped 2 | B2 | WABM232658 WABM243094 | 1.3 cM 1.9 cM | 6BL | Awn suppression | Yoshioka et al. [19] |
| Aegilops tauschii | Anathera | Antr | S57615-1 Xctg211719 | 1.3 cM 3.9 cM | 5DS | Awn suppression | Nishijima et al. [18] |
| Hordeum vulgare | Awnless 1 | Lks1 | SNP 1_0619 SNP 1_1533 | 133.59 cM 141.56 cM | 2HL | Awn suppression | Franckowiak and Lundqvist [59] |
| H. vulgare | Long Awn2 | Lks2 | k06123 k04151 | 0.27 cM 1.0 cM | 7HL | Awn elongation | Yuo et al. [1] |
| H. vulgare | Hooded lemma 1 | Kap1 (Knox3) | glf3 | 25.1 cM | 4HS | Hood formation, awn suppression | Müller et al. [60] |
| H. vulgare | Six-rowed spike 1 | vrs1 | e40m36-1110S BC12348 | 0.01 cM 0.06 cM | 2H | Reduces awn length | Komatsuda et al. [61] |
| H. vulgare | K Suppressor loci suKB | suKB | E34M46 E35M39 | 16.9 cM 9.9 cM | 7H | Reduces awn length, Hood formation | Roig et al. [62] |
| H. vulgare | K Suppressor loci suKC | suKC | E35M44 E41M46 | 0.0 cM 7.8 cM | 7H | Reduces awn length, Hood formation | Roig et al. [62] |
| H. vulgare | K Suppressor loci suKD | suKD | E37M38 E41M36 | 15.9 cM 1.1 cM | 5H | Reduces awn length, Hood formation | Roig et al. [62] |
| H. vulgare | K Suppressor loci suKE | suKE | E40M43 E36M44 | 5.6 cM 0.0 cM | 7H | Hood formation | Roig et al. [62] |
| H. vulgare | K Suppressor loci suKF | suKF | E35M40 E43M32 | 5.3 cM 22.4 cM | 7H | Hood formation | Roig et al. [62] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntakirutimana, F.; Xie, W. Morphological and Genetic Mechanisms Underlying Awn Development in Monocotyledonous Grasses. Genes 2019, 10, 573. https://doi.org/10.3390/genes10080573
Ntakirutimana F, Xie W. Morphological and Genetic Mechanisms Underlying Awn Development in Monocotyledonous Grasses. Genes. 2019; 10(8):573. https://doi.org/10.3390/genes10080573
Chicago/Turabian StyleNtakirutimana, Fabrice, and Wengang Xie. 2019. "Morphological and Genetic Mechanisms Underlying Awn Development in Monocotyledonous Grasses" Genes 10, no. 8: 573. https://doi.org/10.3390/genes10080573
APA StyleNtakirutimana, F., & Xie, W. (2019). Morphological and Genetic Mechanisms Underlying Awn Development in Monocotyledonous Grasses. Genes, 10(8), 573. https://doi.org/10.3390/genes10080573