Sperm Proteome Analysis and Identification of Fertility-Associated Biomarkers in Unexplained Male Infertility


 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Participants and Semen Analysis
2.2. Inclusion and Exclusion Criteria
2.3. Sperm Protein Extraction and Quantification
2.4. Liquid Chromatography-Tandem Mass Spectrometry
- Very Low abundance: SC range 1.7–7; p ≤ 0.001 and NSAF ratio ≥2.5 for overexpressed, ≤0.4 for underexpressed proteins;
- Low abundance: SC range 8–19; p ≤ 0.01 and NSAF ratio ≥2.5 for overexpressed, ≤0.4 for underexpressed proteins;
- Medium abundance: SC range between 20 and 79; p ≤ 0.05 and NSAF ratio ≥2.0 for overexpressed, ≤0.5 for underexpressed proteins;
- High abundance: SC >80; p ≤ 0.05 and NSAF ratio ≥1.5 for overexpressed, ≤0.67 for underexpressed proteins.
2.5. Bioinformatic Analysis
2.6. Protein Selection and Validation by Western Blotting
2.7. Statistical Analysis
3. Results
3.1. Semen Parameters
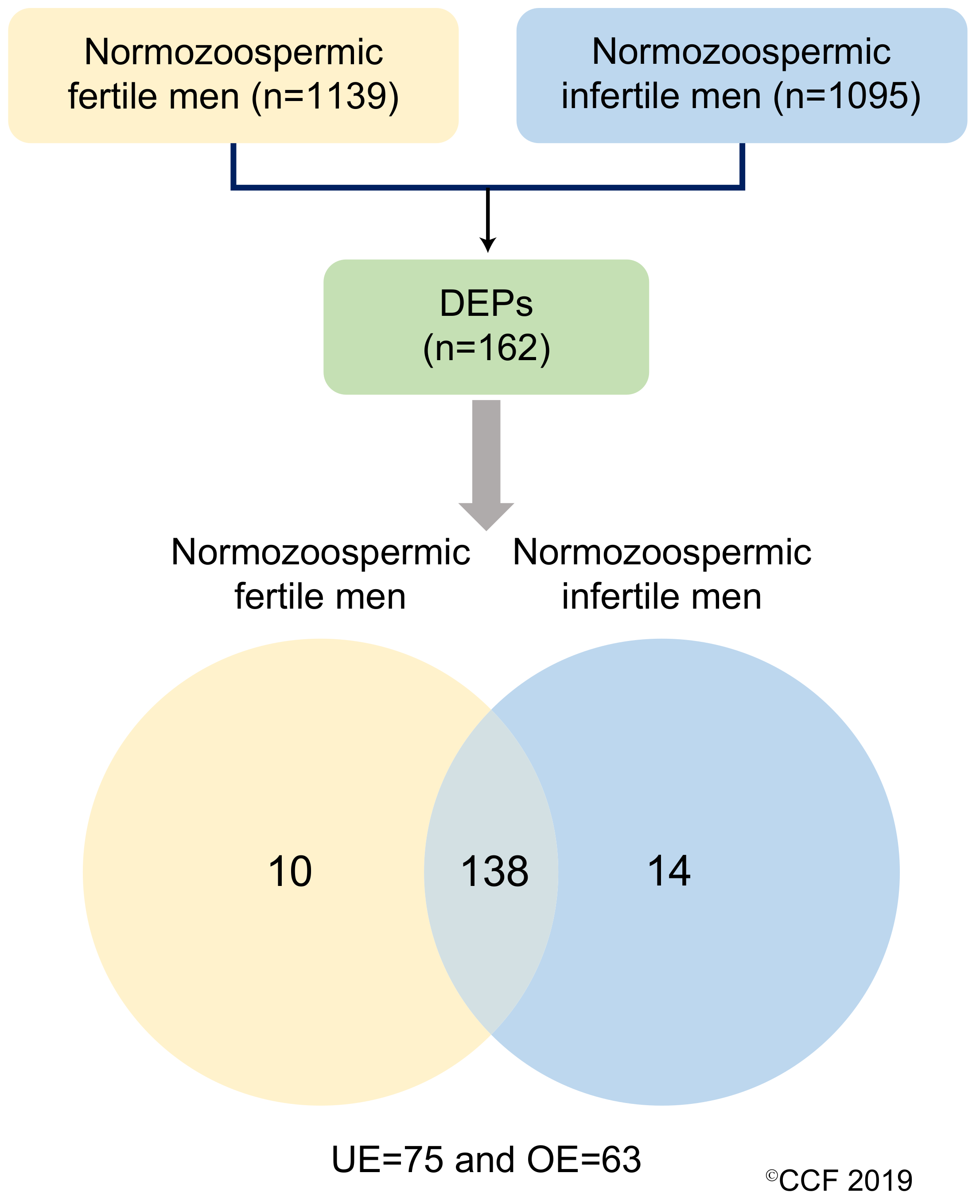
3.2. Sperm Proteome of Normozoospermic Fertile and Normozoospermic Infertile Men
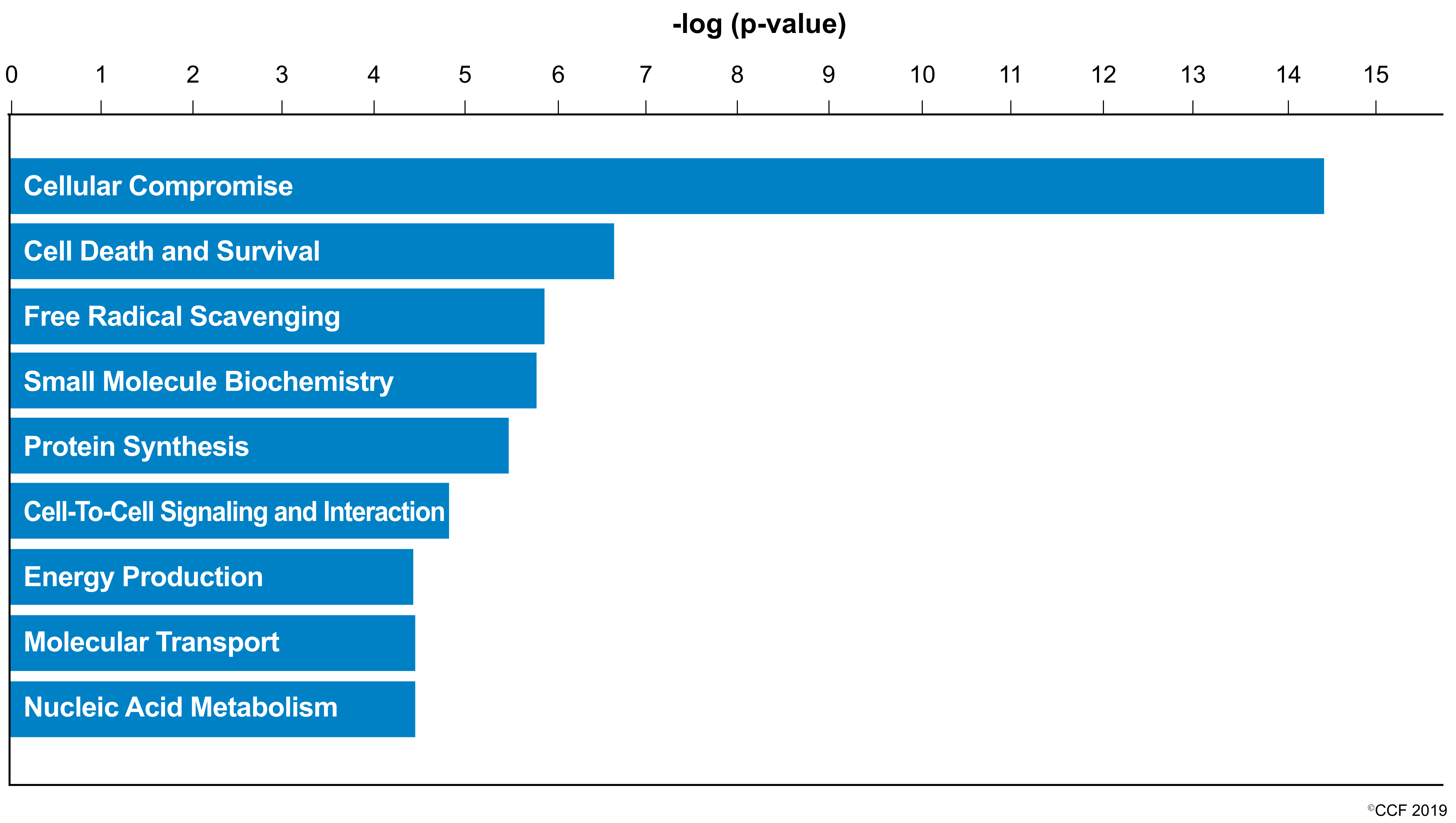
3.3. Key Canonical Pathways Enriched in Normozoospermic Infertile Men
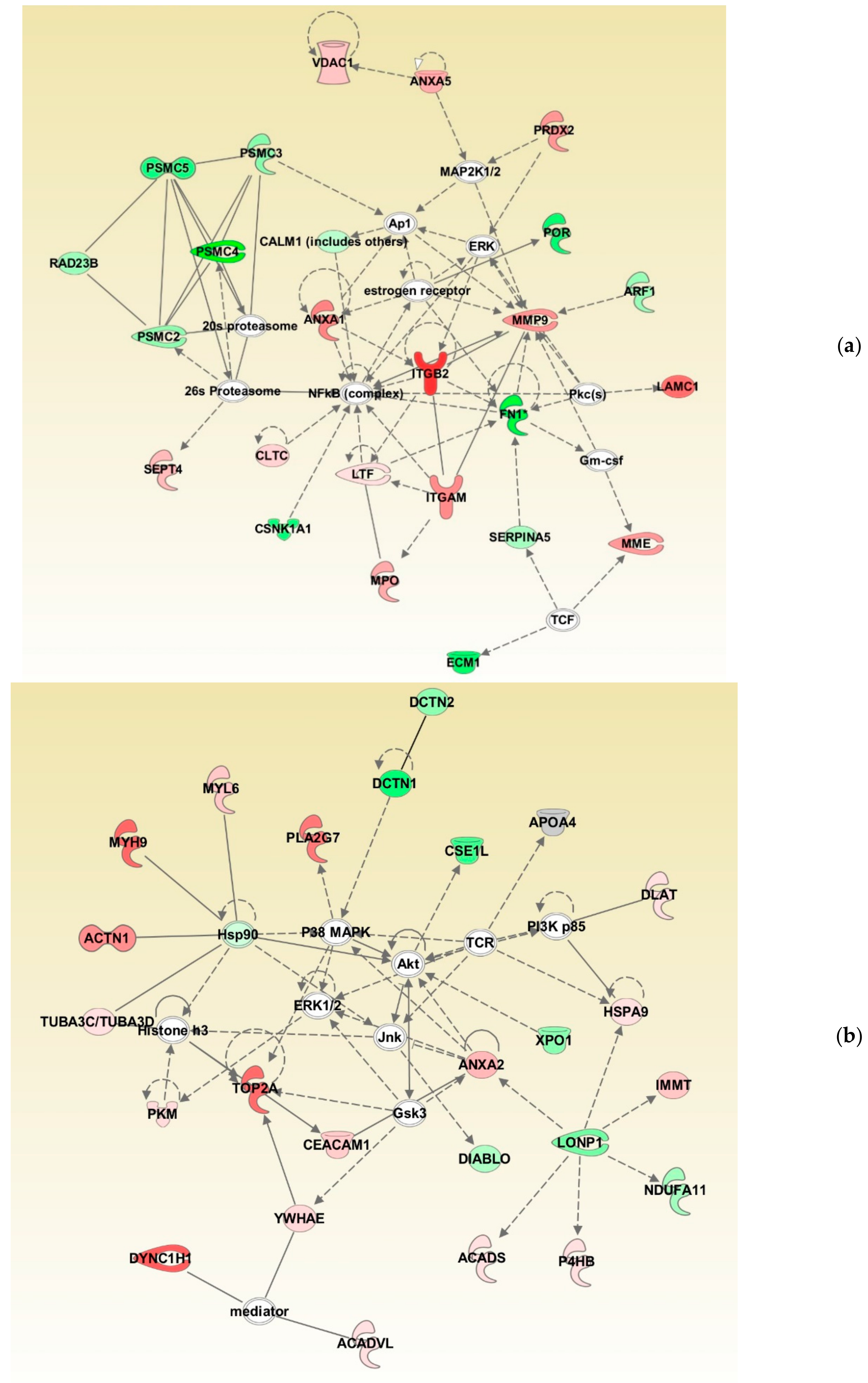
3.4. Protein Networks in Normozoospermic Infertile Men
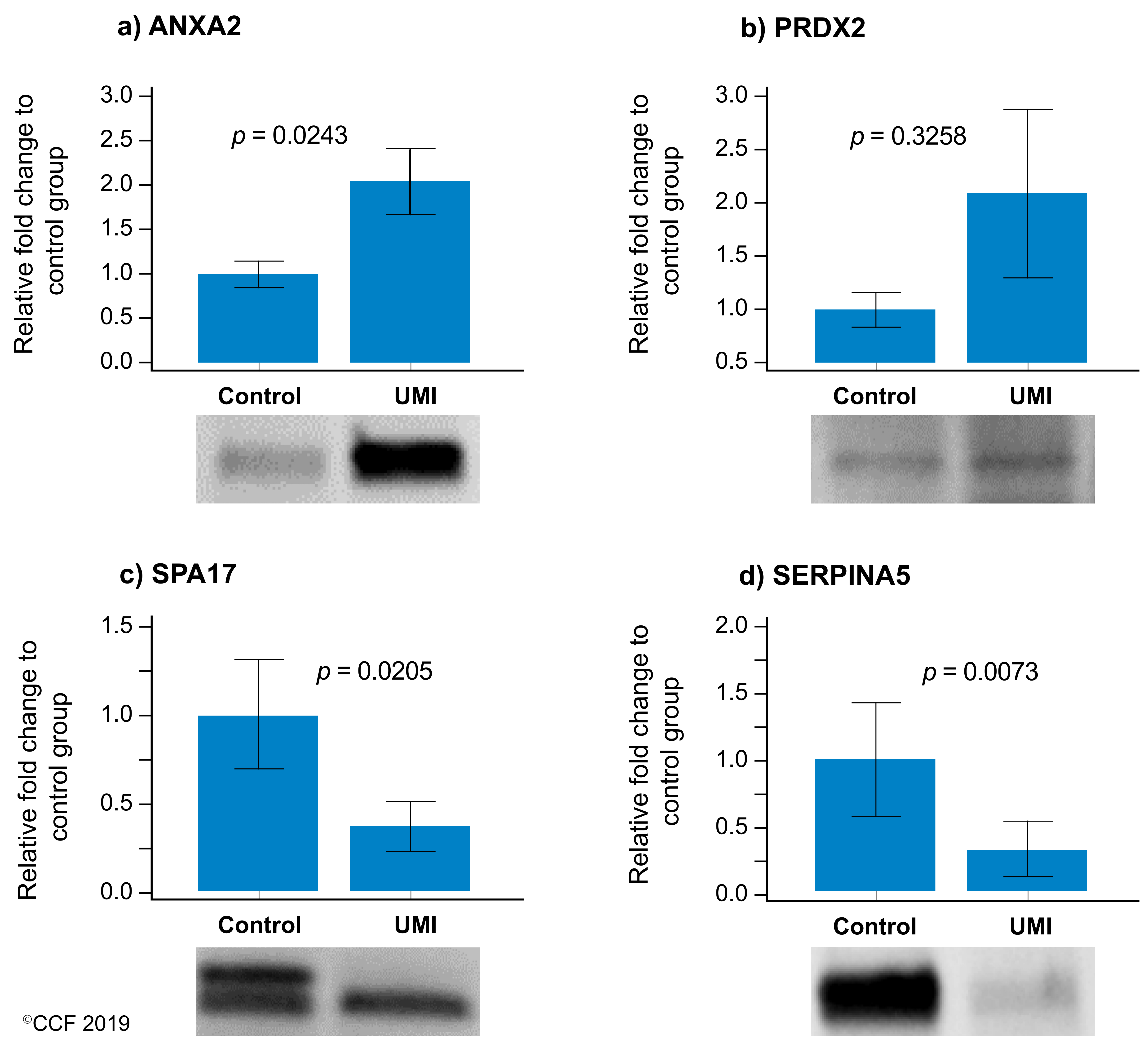
3.5. Western Blot Analysis of Validated DEPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jarow, J.P.; Sharlip, I.D.; Belker, A.M.; Lipshultz, L.I.; Sigman, M.; Thomas, A.J.; Schlegel, P.N.; Howards, S.S.; Nehra, A.; Damewood, M.D. Best practice policies for male infertility. J. Urol. 2002, 167, 2138–2144. [Google Scholar] [CrossRef]
- Nieschlag, S. Andrology: Male Reproductive Health and Dysfunction; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Hamada, A.; Esteves, S.C.; Agarwal, A. Unexplained male infertility: Potential causes and management. Hum. Androl. 2011, 1, 2–16. [Google Scholar] [CrossRef]
- Doshi, S.B.; Sharma, R.K.; Agarwal, A. Oxidative stress in unexplained male infertility. In Unexplained Infertility: Pathophysiology, Evaluation and Treatment; Schattman, G.L., Esteves, S.C., Agarwal, A., Eds.; Springer: New York, NY, USA, 2015; pp. 81–89. [Google Scholar]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; WHO: Geneva, Switzerland, 2010. [Google Scholar]
- Wallach, E.E.; Moghissi, K.S.; Wallach, E.E. Unexplained infertility. Fertil. Steril. 1983, 39, 5–21. [Google Scholar] [CrossRef]
- Panner Selvam, M.K.; Agarwal, A. Update on the proteomics of male infertility: A systematic review. Arab J. Urol. 2018, 16, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Hu, H.; Wang, Z.; Chen, X.; Yang, F.; Zhu, Z.; Fang, P.; Dai, J.; Wang, L.; Shi, H. Proteomic characteristics of spermatozoa in normozoospermic patients with infertility. J. Proteom. 2012, 75, 5426–5436. [Google Scholar] [CrossRef] [PubMed]
- Bracke, A.; Peeters, K.; Punjabi, U.; Hoogewijs, D.; Dewilde, S. A search for molecular mechanisms underlying male idiopathic infertility. Reprod. Biomed. Online 2018, 36, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Azpiazu, R.; Amaral, A.; Castillo, J.; Estanyol, J.M.; Guimerà, M.; Ballescà, J.L.; Balasch, J.; Oliva, R. High-throughput sperm differential proteomics suggests that epigenetic alterations contribute to failed assisted reproduction. Hum. Reprod. 2014, 29, 1225–1237. [Google Scholar] [CrossRef]
- Légaré, C.; Droit, A.; Fournier, F.; Bourassa, S.; Force, A.; Cloutier, F.; Tremblay, R.; Sullivan, R. Investigation of male infertility using quantitative comparative proteomics. J. Proteome Res. 2014, 13, 5403–5414. [Google Scholar] [CrossRef]
- Pixton, K.L.; Deeks, E.D.; Flesch, F.M.; Moseley, F.L.C.; Björndahl, L.; Ashton, P.R.; Barratt, C.L.R.; Brewis, I.A. Sperm proteome mapping of a patient who experienced failed fertilization at IVF reveals altered expression of at least 20 proteins compared with fertile donors: Case report. Hum. Reprod. 2004, 19, 1438–1447. [Google Scholar] [CrossRef]
- Frapsauce, C.; Pionneau, C.; Bouley, J.; Delarouziere, V.; Berthaut, I.; Ravel, C.; Antoine, J.-M.; Soubrier, F.; Mandelbaum, J. Proteomic identification of target proteins in normal but nonfertilizing sperm. Fertil. Steril. 2014, 102, 372–380. [Google Scholar] [CrossRef]
- Lopez, J. Two-dimensional electrophoresis in proteome expression analysis. J. Chromatogr. B 2007, 849, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Bartolomé, S.; Deutsch, E.W.; Binz, P.-A.; Jones, A.R.; Eisenacher, M.; Mayer, G.; Campos, A.; Canals, F.; Bech-Serra, J.-J.; Carrascal, M. Guidelines for reporting quantitative mass spectrometry based experiments in proteomics. J. Proteom. 2013, 95, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Gupta, S.; Sharma, R. Cryopreservation of Client Depositor Semen. In Andrological Evaluation of Male Infertility; Springer: Cham, Switzerland, 2016; pp. 113–133. [Google Scholar]
- Agarwal, A.; Ayaz, A.; Samanta, L.; Sharma, R.; Assidi, M.; Abuzenadah, A.M.; Sabanegh, E. Comparative proteomic network signatures in seminal plasma of infertile men as a function of reactive oxygen species. Clin. Proteom. 2015, 12, 23. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, G.; Liu, J.; Zhu, P.; Wang, J.; Wang, Y.; Wang, W.; Li, N.; Wang, X.; Zhang, C.; et al. iTRAQ-based analysis of sperm proteome from normozoospermic men achieving the rescue-ICSI pregnancy after the IVF failure. Clin. Proteom. 2018, 15, 27. [Google Scholar] [CrossRef] [PubMed]
- McReynolds, S.; Dzieciatkowska, M.; Stevens, J.; Hansen, K.C.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Toward the identification of a subset of unexplained infertility: A sperm proteomic approach. Fertil. Steril. 2014, 102, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Mayorga-Torres, B.J.M.; Camargo, M.; Cadavid, Á.P.; du Plessis, S.S.; Cardona Maya, W.D. Are oxidative stress markers associated with unexplained male infertility? Andrologia 2017, 49, e12659. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Durairajanayagam, D.; Halabi, J.; Peng, J.; Vazquez-Levin, M. Proteomics, oxidative stress and male infertility. Reprod. BioMed. Online 2014, 29, 32–58. [Google Scholar] [CrossRef]
- Sharma, R.; Agarwal, A.; Mohanty, G.; Hamada, A.J.; Gopalan, B.; Willard, B.; Yadav, S.; du Plessis, S. Proteomic analysis of human spermatozoa proteins with oxidative stress. Reprod. Biol. Endocrinol. 2013, 11, 48. [Google Scholar] [CrossRef]
- Hamada, A.J.; Montgomery, B.; Agarwal, A. Male infertility: A critical review of pharmacologic management. Expert Opin. Pharmacother. 2012, 13, 2511–2531. [Google Scholar] [CrossRef]
- Morielli, T.; O’Flaherty, C. Oxidative stress impairs function and increases redox protein modifications in human spermatozoa. Reproduction 2015, 149, 113–123. [Google Scholar] [CrossRef]
- Nakamura, N. Ubiquitination regulates the morphogenesis and function of sperm organelles. Cells 2013, 2, 732–750. [Google Scholar] [CrossRef] [PubMed]
- Kerns, K.; Morales, P.; Sutovsky, P. Regulation of sperm capacitation by the 26S proteasome: An emerging new paradigm in spermatology1. Biol. Reprod. 2016, 94, 117. [Google Scholar] [CrossRef] [PubMed]
- Redgrove, K.A.; Anderson, A.L.; Dun, M.D.; McLaughlin, E.A.; O’Bryan, M.K.; Aitken, R.J.; Nixon, B. Involvement of multimeric protein complexes in mediating the capacitation-dependent binding of human spermatozoa to homologous zonae pellucidae. Dev. Biol. 2011, 356, 460–474. [Google Scholar] [CrossRef] [PubMed]
- Teijeiro, J.M.; Roldán, M.L.; Marini, P.E. Annexin A2 and S100A10 in the mammalian oviduct. Cell Tissue Res. 2016, 363, 567–577. [Google Scholar] [CrossRef]
- He, K.L.; Deora, A.B.; Xiong, H.; Ling, Q.; Weksler, B.B.; Niesvizky, R.; Hajjar, K.A. Endothelial cell annexin A2 regulates polyubiquitination and degradation of its binding partner S100A10/p11. J. Biol. Chem. 2008, 283, 19192–19200. [Google Scholar] [CrossRef]
- Deng, S.; Jing, B.; Xing, T.; Hou, L.; Yang, Z. Overexpression of annexin A2 is associated with abnormal ubiquitination in breast cancer. Genom. Proteom. Bioinform. 2012, 10, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.R.; Miki, K.; Harper, D.B.; Eddy, E.M. A-Kinase anchoring protein 4 binding proteins in the fibrous sheath of the sperm flagellum. Biol. Reprod. 2003, 68, 2241–2248. [Google Scholar] [CrossRef]
- Vizel, R.; Hillman, P.; Ickowicz, D.; Breitbart, H. AKAP3 degradation in sperm capacitation is regulated by its tyrosine phosphorylation. Biochim. Biophys. Acta (BBA) Gen. Subj. 2015, 1850, 1912–1920. [Google Scholar] [CrossRef]
- Chiriva-Internati, M.; Gagliano, N.; Donetti, E.; Costa, F.; Grizzi, F.; Franceschini, B.; Albani, E.; Levi-Setti, P.E.; Gioia, M.; Jenkins, M.; et al. Sperm protein 17 is expressed in the sperm fibrous sheath. J. Transl. Med. 2009, 7, 61. [Google Scholar] [CrossRef]
- Intasqui, P.; Agarwal, A.; Sharma, R.; Samanta, L.; Bertolla, R.P. Towards the identification of reliable sperm biomarkers for male infertility: A sperm proteomic approach. Andrologia 2018, 50, e12919. [Google Scholar] [CrossRef]
- Wen, Y.; Richardson, R.T.; Widgren, E.E.; O’Rand, M.G. Characterization of Sp17: A ubiquitous three domain protein that binds heparin. Biochem. J. 2001, 357, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Frayne, J.; Hall, L. A re-evaluation of sperm protein 17 (Sp17) indicates a regulatory role in an A-kinase anchoring protein complex, rather than a unique role in sperm-zona pellucida binding. Reproduction 2002, 124, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Petit, F.M.; Serres, C.; Bourgeon, F.; Pineau, C.; Auer, J. Identification of sperm head proteins involved in zona pellucida binding. Hum. Reprod. 2013, 28, 852–865. [Google Scholar] [CrossRef]
- Kichine, E.; Di Falco, M.; Hales, B.F.; Robaire, B.; Chan, P. Analysis of the sperm head protein profiles in fertile men: Consistency across time in the levels of expression of heat shock proteins and peroxiredoxins. PLoS ONE 2013, 8, e77471. [Google Scholar] [CrossRef] [PubMed]
- Uhrin, P.; Dewerchin, M.; Hilpert, M.; Chrenek, P.; Schöfer, C.; Zechmeister-Machhart, M.; Krönke, G.; Vales, A.; Carmeliet, P.; Binder, B.R.; et al. Disruption of the protein C inhibitor gene results in impaired spermatogenesis and male infertility. J. Clin. Investig. 2000, 106, 1531–1539. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Geiger, M. Cell penetrating SERPINA5 (ProteinC inhibitor, PCI): More questions than answers. Semin. Cell Dev. Biol. 2017, 62, 187–193. [Google Scholar] [CrossRef]
- Moore, A.; Penfold, L.M.; Johnson, J.L.; Latchman, D.S.; Moore, H.D.M. Human sperm–egg binding is inhibited by peptides corresponding to core region of an acrosomal serine protease inhibitor. Mol. Reprod. Dev. 1993, 34, 280–291. [Google Scholar] [CrossRef]
- España, F.; Sánchez-Cuenca, J.; Fernández, P.J.; Gilabert, J.; Romeu, A.; Estellés, A.; Royo, M.; Muller, C.H. Inhibition of human sperm–zona-free hamster oocyte binding and penetration by protein C inhibitor. Andrologia 1999, 31, 217–223. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functions | p-Value | Upregulated Proteins | Downregulated Proteins |
|---|---|---|---|
| Fertilization | 0.0000147 | AKAP3, AKAP4, PLA2G7 | SPA17 |
| Binding of gonadal cell lines | 0.0000148 | ANXA5, ITGAM, ITGB2, LTF | FN1 |
| Symbol | Protein Name | Expr Fold Change | p-Value |
|---|---|---|---|
| HSP90B1 | heat shock protein 90 β family member 1 | −0.664 | 0.000359 |
| PSMC2 | 26S protease regulatory subunit 7 isoform 1 | −1.693 | 0.00723 |
| PSMC3 | 26S protease regulatory subunit 6A | −1.737 | 0.00195 |
| PSMC4 | 26S protease regulatory subunit 6B isoform 1 | −4.916 | 0.000978 |
| PSMC5 | 26S protease regulatory subunit 8 isoform 1 | −2.587 | 0.00319 |
| UCHL1 | ubiquitin carboxyl-terminal hydrolase isozyme L1 | −1.698 | 0.00524 |
| Study/Authors | Study Description | Proteomic Technique | Western Blot Validation | Limitations |
|---|---|---|---|---|
| Pixton et al., 2004 [12] | • Fertilization failure (n = 1) • Fertile donors (n = 3) | • 2DE and MS/MS • Pooled sample | • Not performed | • Less sample size • Lack of WB validation |
| Xu et al., 2012 [8] | • Fertile men (n = 10) • Infertile men (n = 10) | • 2-DE and MALDI-TOF/TOF • Pooled sample | • Proteins: PAEP, ODFP, SEMG1, PSA, GPx4Pre • Individual samples (n = 10) | • Lacks the WB validation of proteins associated with fertilization process, except for ODFP |
| Frapsauce et al., 2014 [13] | • Fertilization failure with IVF (n = 3) • Successful fertilization with IVF (n = 3) | • 2D-DIGE and MS • Pooled sample | • Not performed | • Use of DIGE • Lack of WB validation |
| McReynolds et al., 2014 [19] | • Infertile men (n = 12) | • LC-MS/MS • Pooled sample | • Protein: CLU • Individual samples (n = 12) | • WB validation of only one protein CLU |
| Azpiazu et al., 2014 [10] | • No pregnancy with IVF (n = 15) • Pregnancy with IVF (n = 16) | • LC-MS/MS • Pooled sample (n = 10) | • Protein: SRPK1 • Individual samples ○ No pregnancy (n = 6) ○ Pregnancy (n = 7) | • WB validation of only one protein SRPK1 |
| Légaré et a., 2014 [11] | • Fertile (n = 3+ n = 3) • Infertile (n = 6) • IVF failure (n = 4) | • iTRAQ and LC-MS/MS • Two pools of fertile group • Single pool of infertile and IVF failure group | • Proteins: SEMG1, PIP, GAPDHS, PGK2 • Individual samples • Fertile (n = 13) • IVF failure (n = 14) | • Lacks the WB validation of proteins associated with fertilization process |
| Liu et al., 2018 [18] | • Pregnancy with IVF (n = 20) • Pregnancy with ICSI (n = 20) | • iTRAQ and LC-MS/MS • Pooled sample | • Proteins: ZPBP1, ACRBP • Individual samples (n = 12) | • Lacks the comparison with control group (fertile men) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panner Selvam, M.K.; Agarwal, A.; Pushparaj, P.N.; Baskaran, S.; Bendou, H. Sperm Proteome Analysis and Identification of Fertility-Associated Biomarkers in Unexplained Male Infertility. Genes 2019, 10, 522. https://doi.org/10.3390/genes10070522
Panner Selvam MK, Agarwal A, Pushparaj PN, Baskaran S, Bendou H. Sperm Proteome Analysis and Identification of Fertility-Associated Biomarkers in Unexplained Male Infertility. Genes. 2019; 10(7):522. https://doi.org/10.3390/genes10070522
Chicago/Turabian StylePanner Selvam, Manesh Kumar, Ashok Agarwal, Peter Natesan Pushparaj, Saradha Baskaran, and Hocine Bendou. 2019. "Sperm Proteome Analysis and Identification of Fertility-Associated Biomarkers in Unexplained Male Infertility" Genes 10, no. 7: 522. https://doi.org/10.3390/genes10070522
APA StylePanner Selvam, M. K., Agarwal, A., Pushparaj, P. N., Baskaran, S., & Bendou, H. (2019). Sperm Proteome Analysis and Identification of Fertility-Associated Biomarkers in Unexplained Male Infertility. Genes, 10(7), 522. https://doi.org/10.3390/genes10070522