Further Insights into the Architecture of the PN Promoter That Controls the Expression of the bzd Genes in Azoarcus



Abstract
1. Introduction
2. Material and Methods
2.1. Bacterial Strains, Plasmids, and Growth Conditions
2.2. Molecular Biology Techniques
2.3. Sequence Data Analyses
2.4. Overproduction and Purification of His6-BzdR, His6-NBzdRL, and His6-FNR* Proteins
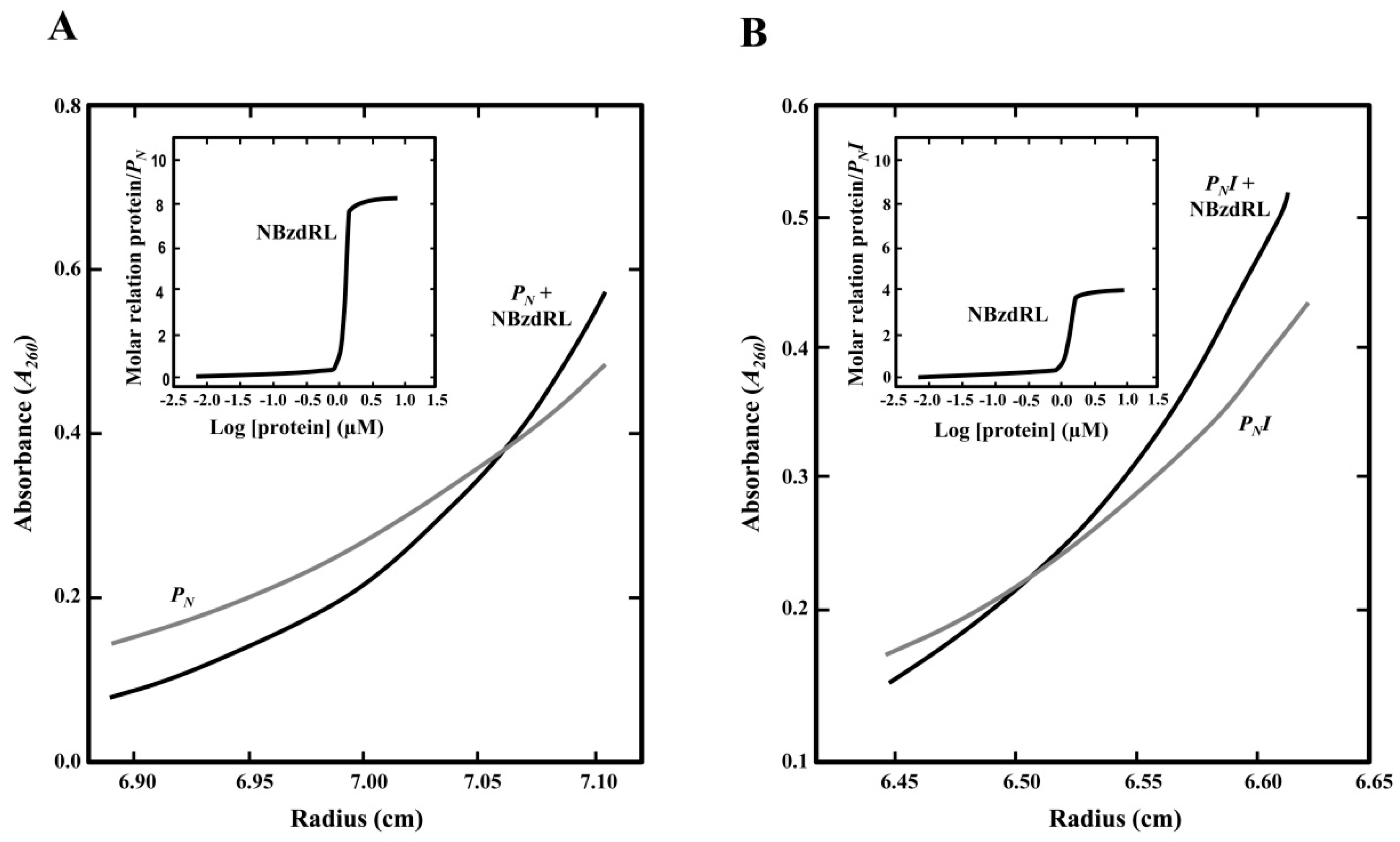
2.5. Analytical Ultracentrifugation Assays
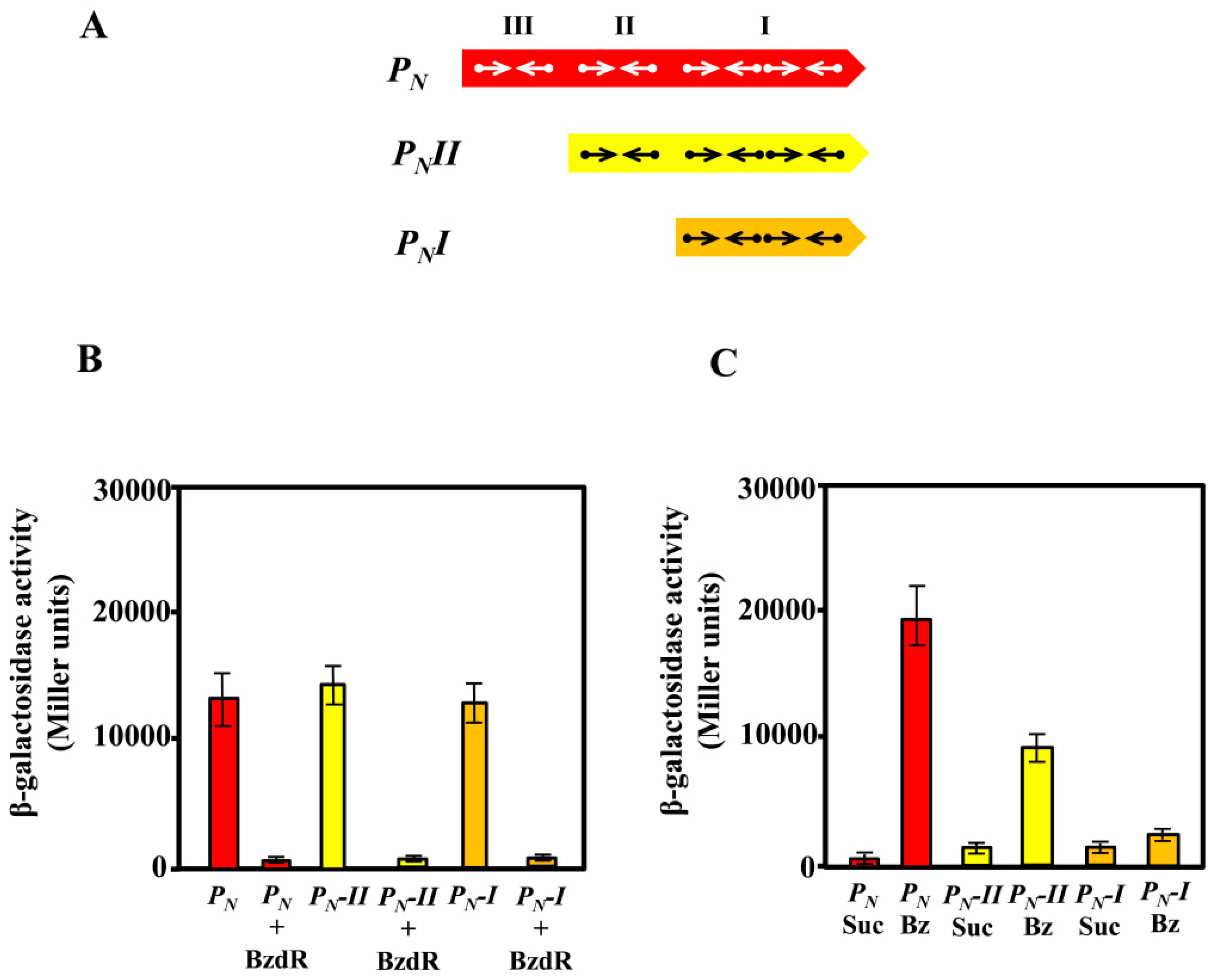
2.6. Construction of PNI::lacZ and PNII::lacZ Translational Fusions and β-Galactosidase Assays
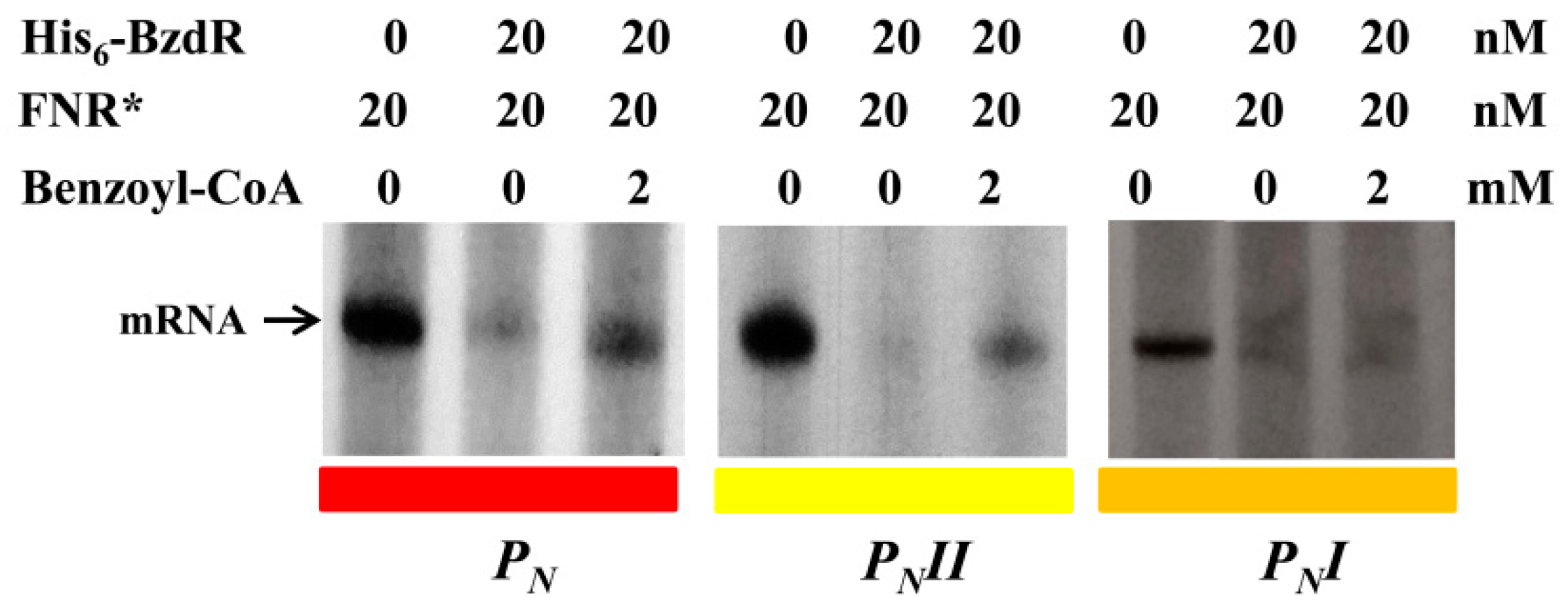
2.7. In Vitro Transcription Assays
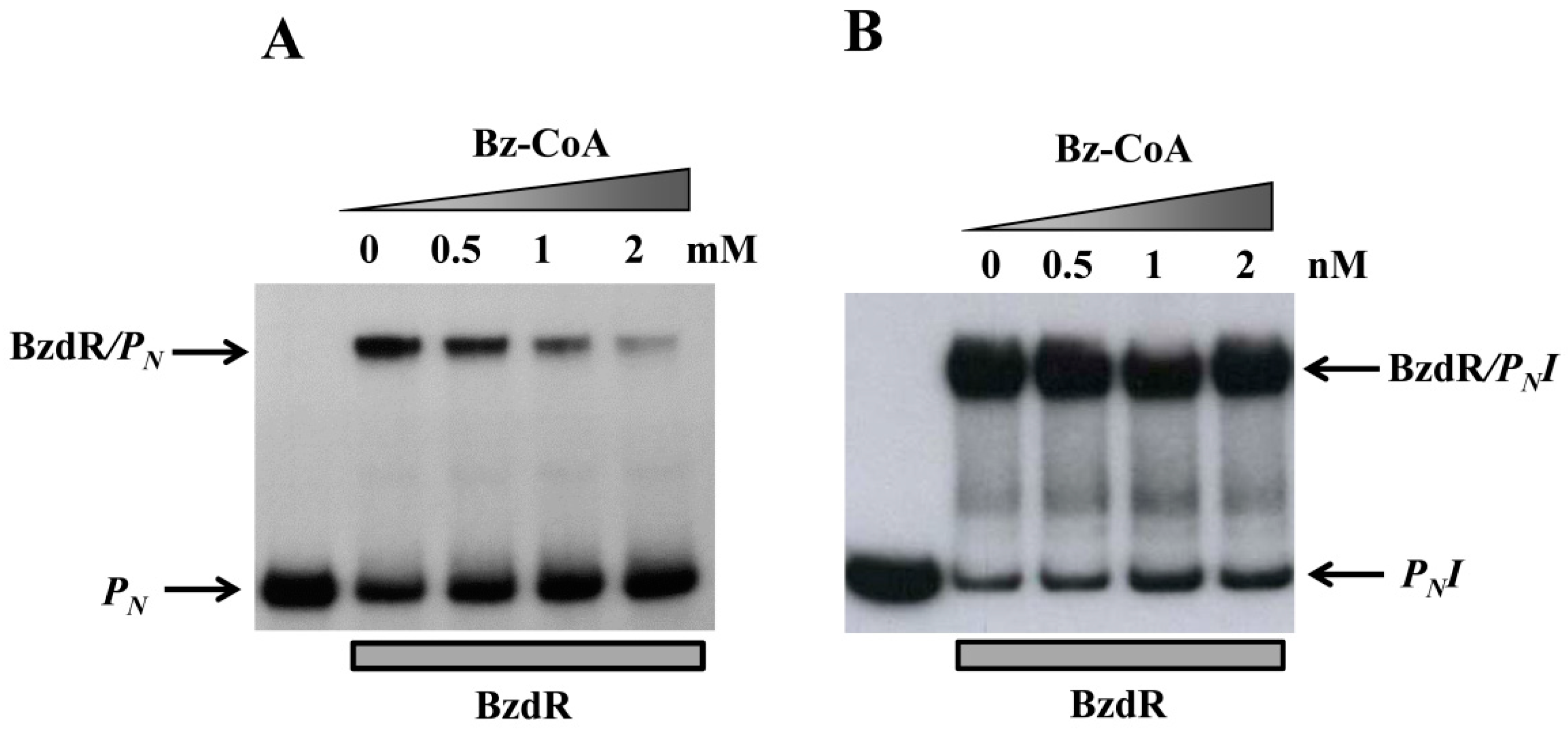
2.8. Gel Retardation Assays
2.9. Analysis of Protein–DNA Interaction by Atomic Force Microscopy
3. Results and Discussion
3.1. The Genetic Organization of the bzd Cluster and the Architecture of the PN Promoter Are Conserved in the Azoarcus/Aromatoleum Genus
3.2. In Vivo Studies on the Activity of Truncated PN Promoters
3.3. Operator Regions II and III Are Needed for the Benzoyl-CoA-Dependent De-Repression of the PN Promoter
3.4. Visualization of the BzdR/PN Complex
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pieper, D.H.; Reineke, W. Engineering bacteria for bioremediation. Curr. Opin. Biotechnol. 2000, 11, 262–270. [Google Scholar] [CrossRef]
- Fuchs, G.; Boll, M.; Heider, J. Microbial degradation of aromatic compounds—From one strategy to four. Nat. Rev. Microbiol. 2011, 9, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Brzeszcz, J.; Kaszycki, P. Aerobic bacteria degrading both n-alkanes and aromatic hydrocarbons: And undervalued strategy for metabolic diversity and flexibility. Biodegradation 2018, 29, 359–407. [Google Scholar] [CrossRef] [PubMed]
- Díaz, E.; Jiménez, J.I.; Nogales, J. Aerobic degradation of aromatic compounds. Curr. Opin. Biotechnol. 2013, 24, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.; Jauregui, R.; Vilchez-Vargas, R.; Junca, H.; Pieper, D.H. AromaDeg, a novel database for phylogenomics of aerobic bacterial degradation of aromatics. Database 2014, 2014, bau118. [Google Scholar] [CrossRef] [PubMed]
- Rabus, R.; Boll, M.; Heider, J.; Meckenstock, R.U.; Buckel, W.; Einsle, O.; Ermler, U.; Golding, B.T.; Gunsalus, R.P.; Kroneck, P.M.H.; et al. Anaerobic microbial degradation of hydrocarbons: From enzymatic reactions to the environment. J. Mol. Microbiol. Biotechnol. 2016, 26, 5–28. [Google Scholar] [CrossRef]
- Lovley, D.R. Biorremediation. Anaerobes to the rescue. Science 2001, 293, 1444–1446. [Google Scholar] [CrossRef]
- Carmona, M.; Zamarro, M.T.; Blázquez, B.; Durante-Rodríguez, G.; Juárez, J.F.; Valderrama, J.A.; López-Barragán, M.J.; García, J.L.; Díaz, E. Anaerobic catabolism of aromatic compounds: A genetic and genomic view. Microbiol. Mol. Biol. Rev. 2009, 73, 71–133. [Google Scholar] [CrossRef]
- Boll, M.; Löffler, C.; Morris, B.E.; Kung, J.W. Anaerobic degradation of homocyclic aromatic compounds via arylcarboxyl-coenzyme A esters: Organisms, strategies and key enzymes. Environ. Microbiol. 2014, 16, 612–627. [Google Scholar] [CrossRef]
- Durante-Rodríguez, G.; Gómez-Álvarez, H.; Blázquez, B.; Fernández-Llamosas, H.; Martín-Moldes, Z.; Sanz, D.; Nogales, J.; Carmona, M.; Díaz, E. Anaerobic pathways for the catabolism of aromatic compounds. In Lignin Valorization: Emerging Approaches; Beckham, T., Ed.; The Royal Society of Chemistry Publishing: Cambridge, UK, 2018; pp. 333–390. [Google Scholar]
- Egland, P.G.; Pelletier, D.A.; Dispensa, M.; Gibson, J.; Harwood, C.S. A cluster of bacterial genes for anaerobic benzene ring biodegradation. Proc. Natl. Acad. Sci. USA 1997, 94, 6484–6489. [Google Scholar] [CrossRef]
- Wischgoll, S.; Heintz, D.; Peters, F.; Erxleben, A.; Sarnighausen, E.; Reski, R.; van Dorsselaer, A.; Boll, M. Gene clusters involved in anaerobic benzoate degradation in Geobacter metallireducens. Mol. Microbiol. 2005, 58, 1238–1252. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.E.; He, Q.; Nevin, K.P.; He, Z.; Zhou, J.; Lovley, D.R. Genomic and microarray analysis of aromatics degradation in Geobacter metallireducens and comparison to a Geobacter isolate from a contaminated field site. BMC Genom. 2007, 8, 180. [Google Scholar] [CrossRef] [PubMed]
- McInerney, M.J.; Rohlin, L.; Mouttaki, H.; Kim, U.; Krupp, R.S.; Rios-Hernandez, L.; Sieber, J.; Struchtemeyer, C.G.; Bhattacharyya, A.; Campbell, J.W.; et al. The genome of Syntrophus aciditrophicus: Life at the thermodynamic limit of microbial growth. Proc. Natl. Acad. Sci. USA 2007, 104, 7600–7605. [Google Scholar] [CrossRef] [PubMed]
- Wöhlbrand, L.; Jacob, J.H.; Kube, M.; Mussmann, M.; Jarling, R.; Beck, A.; Amann, R.; Wilkes, H.; Reinhardt, R.; Rabus, R. Complete genome, catabolic sub-proteomes and key-metabolites of Desulfobacula toluolica Tol2, a marine, aromatic compound-degrading, sulfate-reducing bacterium. Environ. Microbiol. 2013, 15, 1334–1355. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, H.; Hirakawa, Y.; Greenberg, E.P.; Harwood, C.S. BadR and BadM proteins transcriptionally regulate two operons needed for anaerobic benzoate degradation by Rhodopseudomonas palustris. Appl. Environ. Microbiol. 2015, 81, 4253–4262. [Google Scholar] [CrossRef] [PubMed]
- Van Drisse, C.M.; Escalante-Semerena, J.C. Small-molecule acetylation controls the degradation of benzoate and photosynthesis in Rhodopseudomonas palustris. MBio 2018, 16, e01895-18. [Google Scholar]
- Ueki, T. Identification of a transcriptional repressor involved in benzoate metabolism in Geobacter bemidjiensis. Appl. Environ. Microbiol. 2011, 77, 7058–7062. [Google Scholar] [CrossRef] [PubMed]
- Juárez, J.F.; Zamarro, M.T.; Barragán, M.J.; Blázquez, B.; Boll, M.; Kuntze, K.; García, J.L.; Díaz, E.; Carmona, M. Identification of the Geobacter metallireducens bamVW two-component system, involved in transcriptional regulation of aromatic degradation. Appl. Environ. Microbiol. 2010, 76, 383–385. [Google Scholar] [CrossRef]
- Barragán, M.J.; Blázquez, B.; Zamarro, M.T.; Mancheño, J.M.; García, J.L.; Díaz, E.; Carmona, M. BzdR, a repressor that controls the anaerobic catabolism of benzoate in Azoarcus sp. CIB, is the first member of a new subfamily of transcriptional regulators. J. Biol. Chem. 2005, 280, 10683–10694. [Google Scholar] [CrossRef]
- Durante-Rodríguez, G.; Valderrama, J.A.; Mancheño, J.M.; Rivas, G.; Alfonso, C.; Arias-Palomo, E.; Llorca, O.; García, J.L.; Díaz, E.; Carmona, M. Biochemical characterization of the transcriptional regulator BzdR from Azoarcus sp. CIB. J. Biol. Chem. 2010, 285, 35694–35705. [Google Scholar] [CrossRef]
- Rabus, R.; Wöhlbrand, L.; Thies, D.; Meyer, M.; Reinhold-Hurek, B.; Kämpfer, P. Aromatoleum gen. nov., a novel genus accommodating the phylogenetic lineage including Azoarcus evansii and related species, and proposal of Aromatoleum aromaticum sp. nov., Aromatoleum petrolei sp. nov., Aromatoleum bremense sp. nov., Aromatoleum toluolicum sp. nov. and Aromatoleum diolicum sp. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 982–997. [Google Scholar] [PubMed]
- López-Barragán, M.J.; Carmona, M.; Zamarro, M.T.; Thiele, B.; Boll, M.; Fuchs, G.; García, J.L.; Díaz, E. The bzd gene cluster, coding for anaerobic benzoate catabolism, in Azoarcus sp. strain CIB. J. Bacteriol. 2004, 186, 5762–5774. [Google Scholar] [CrossRef] [PubMed]
- Durante-Rodríguez, G.; Zamarro, M.T.; García, J.L.; Díaz, E.; Carmona, M. Oxygen-dependent regulation of the central pathway for the anaerobic catabolism of aromatic compounds in Azoarcus sp. strain CIB. J. Bacteriol. 2006, 188, 2343–2354. [Google Scholar] [CrossRef] [PubMed]
- Valderrama, J.A.; Shingler, V.; Carmona, M.; Díaz, E. AccR is a master regulator involved in carbon catabolite repression of the anaerobic catabolism of aromatic compounds in Azoarcus sp. CIB. J. Biol. Chem. 2014, 289, 1892–1904. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Rusell, D. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- De Lorenzo, V.; Timmis, K.N. Analysis and construction of stable phenotypes in gram-negative bacteria with Tn5- and Tn10-derived minitransposons. Methods Enzymol. 1994, 235, 386–405. [Google Scholar] [PubMed]
- Casadaban, M.J. Transposition and fusion of the lac genes to selected promoters in Escherichia coli using bacteriophage lambda and Mu. J. Mol. Biol. 1976, 104, 541–555. [Google Scholar] [CrossRef]
- Wing, H.J.; Green, J.; Guest, J.R.; Busby, S.J. Role of activating region 1 of Escherichia coli FNR protein in transcription activation at class II promoter. J. Biol. Chem. 2000, 275, 29061–29065. [Google Scholar] [CrossRef]
- Durante-Rodríguez, G.; Mancheño, J.M.; Rivas, G.; Alfonso, C.; García, J.L.; Díaz, E.; Carmona, M. Identification of a missing link in the evolution of an enzyme into a transcriptional regulator. PLoS ONE 2013, 8, e57518. [Google Scholar] [CrossRef]
- Kovach, M.; Elzer, P.H.; Hill, D.S.; Robertson, G.T.; Farris, M.A.; Roop, R.M.I.; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- Marshall, C.; Labrousse, V.; Kreimer, M.; Weichart, D.; Kolb, A.; Hengge-Aronis, R. Molecular analysis of the regulation of csiD, a carbon starvation-inducible gene in Escherichia coli that is exclusively dependent on σs and requires activation by cAMP-CRP. J. Mol. Biol. 1998, 276, 339–353. [Google Scholar] [CrossRef]
- Ferrández, A.; Miñambres, B.; García, B.; Olivera, E.R.; Luengo, J.M.; García, J.L.; Díaz, E. Catabolism of phenylacetic acid in Escherichia coli. Characterization of a new aerobic hybrid pathway. J. Biol. Chem. 1998, 273, 25974–26986. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Press: New York, NY, USA, 1972; pp. 352–355. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI) Server. Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 26 June 2019).
- Wilbur, W.J.; Lipman, D.J. Rapid similarity searches of nucleic acid and protein data banks. Proc. Natl. Acad. Sci. USA 1983, 80, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Claverie, F.; Magasanik, B. Positive and negative effects of DNA bending on activation of transcription from a distant site. J. Mol. Biol. 1992, 227, 996–1008. [Google Scholar] [CrossRef]
- Carmona, M.; Magasanik, B. Activation of transcription at σ54-dependent promoters on linear templates requires intrinsic or induced bending of the DNA. J. Mol. Biol. 1996, 261, 348–356. [Google Scholar] [CrossRef]
- Collins, T.J. ImageJ for microscopy. Biotechniques 2007, 43, 25–30. [Google Scholar] [CrossRef]
- Moreno-Herrero, F.; De Pablo, P.J.; Fernández-Sánchez, R.; Colchero, J.; Gómez-Herrero, J.; Baró, A.M. Scanning force microscopy jumping and tapping modes in liquids. Appl. Phys. Lett. 2002, 81, 2620–2622. [Google Scholar] [CrossRef]
- Horcas, I.; Fernández, R.; Gómez-Rodríguez, J.M.; Colchero, J.; Gómez-Herrero, J.; Baro, A.M. WSXM: A software for scanning probe microscopy and a tool for nanotechnology. Rev. Sci. Instrum. 2007, 78, 013705. [Google Scholar] [CrossRef]
- Rabus, R.; Trautwein, K.; Wöhlbrand, L. Towards habitat-oriented systems biology of “Aromatoleum aromaticum” EbN1: Chemical sensing, catabolic network modulation and growth control in anaerobic aromatic compound degradation. Appl. Microbiol. Biotechnol. 2014, 98, 3371–3388. [Google Scholar] [CrossRef] [PubMed]
- Del Solar, G.H.; Pérez-Martín, J.; Espinosa, M. Plasmid pLS1-enconded RepA protein regulates transcription from repAB promoter by binding to a DNA sequence containing a 13-base pair symmetric element. J. Biol. Chem. 1990, 265, 12569–12575. [Google Scholar] [PubMed]
- Gomis-Ruth, F.X.; Sola, M.; Acebo, P.; Parraga, A.; Guasch, A.; Eritja, R.; González, A.; Espinosa, M.; del Solar, G.; Coll, M. The structure of plasmid-encoded transcriptional repressor CopG unligated and bound to its operator. EMBO J. 1998, 17, 7404–7415. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, M.M.; Ramos, J.L.; Gallegos, M.T.; Marqués, S. Critical nucleotides in the upstream region of the XylS-dependent TOL meta-cleavage pathway operon promoter as deduced from analysis of mutants. J. Biol. Chem. 1999, 274, 2286–2290. [Google Scholar] [CrossRef] [PubMed]
- Cowles, C.E.; Nichols, N.N.; Harwood, C.S. BenR, a XylS homologue regulates three different pathways of aromatic acid degradation in Pseudomonas putida. J. Bacteriol. 2000, 182, 6339–6346. [Google Scholar] [CrossRef] [PubMed]
- Körner, H.; Sofia, H.J.; Zumft, W.G. Phylogeny of the bacterial superfamily of Crp-Fnr transcriptional regulators: Exploiting the metabolic spectrum by controlling alterative gene programas. FEMS Microbiol. Rev. 2003, 27, 559–592. [Google Scholar] [CrossRef]
- Hansma, H. Surface biology of DNA by atomic force microscopy. Annu. Rev. Phys. Chem. 2001, 52, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Rivetti, C.; Guthold, M. Single DNA molecule analysis of transcription complexes. Methods Enzymol. 2003, 371, 34–50. [Google Scholar]
- Yang, Y.; Sass, L.; Du, C.; Hsieh, P.; Erie, D. Determination of protein-DNA binding constants and specificities from statistical analyses of single molecule: MutS-DNA interactions. Nucleic Acids Res. 2005, 33, 4322–4334. [Google Scholar] [CrossRef]
- Zhang, J.; Zeuner, Y.; Kleefeld, A.; Unden, G.; Janshoff, A. Multiple site-specific binding of Fis protein to Escherichia coli nuoA-N promoter DNA and its impact on DNA topology visualized by means of scanning force microscopy. ChemBioChem 2004, 5, 1286–1289. [Google Scholar] [CrossRef]
- Gutiérrez-del-Arroyo, P. Caracterización Mediante Microscopía de Fuerzas Atómicas de Complejos de Regulación Transcripcional. Ph.D. Thesis, Universidad Autónoma de Madrid, Madrid, Spain, 2009. [Google Scholar]
- Jia, Y.; Bi, L.; Li, F.; Chen, Y.; Zhang, C.; Zhang, X. α-shaped DNA loops induced by MutS. Biochem. Biophys. Res. Commun. 2008, 372, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Lacal, J.; Guazzaroni, M.E.; Gutiérrez del Arroyo, P.; Busch, A.; Vélez, M.; Krell, T.; Ramos, J.L. Two level of comparativeness in the binding of TodT to the tod operon promoter. J. Mol. Biol. 2008, 384, 1037–1047. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Relevant Genotype and Characteristic(s) | Reference |
|---|---|---|
| Escherichia coli strain | ||
| DH5α | endA1 hsdR17 supE44 thi-1 recA1 gyrA(Nalr) relA1 Δ(argF-lac) U169 depR Φ80dlacd(lacZ) M15 | [26] |
| S17-1λpir | Tpr Smr recA thi hsdRM+ RP4::2-Tc::Mu::Km λpir phage lysogen | [27] |
| M15 | Strain for regulated high-level expression with pQE vector | Qiagen * |
| MC4100 | F-, araD319, Δ(argF-lac)U169 rpsL150 (Smr) relA1 flbB5301 deoC1 ptsF25 rbs | [28] |
| Azoarcus strain | ||
| CIB | Wild-type strain | [23] |
| Plasmids | ||
| pQE32 | Apr, oriColE1 T5 promoter lac operator, λt0/E. coli rrnB T1 terminators, N-terminal His6 | Qiagen * |
| pQE32-His6BzdR | Apr, pQE32 derivative harboring the His6-bzdR gene | [20] |
| pQE60-His6Fnr* | Apr, pQE60 derivative harboring the His6- FNR* gene under the control of T5 promoter lac operator | [29] |
| pQE32-His6NBzdRL | Apr, pQE32 derivative harboring the His6- NBzdRL fragment | [30] |
| pREP4 | Kmr, plasmid that expresses the lac I repressor | Qiagen * |
| pECOR7 | Apr, pUC19 harboring a 7.1-kb EcoRI DNA fragment containing the bzdRNO genes | [23] |
| pBBR1MCS-5 | Gmr, oripBBR1MCS Mob+ lacZα, broad-host- range cloning and expression vector | [31] |
| pBBR5PN | Gmr, pBBR1MCS-5 derivative harboring a PN::lacZ translational fusion | [20] |
| pBBR5PNI | Gmr, pBBR1MCS-5 derivative harboring a PNI::lacZ translational fusion | This work |
| pBBR5PNII | Gmr, pBBR1MCS-5 derivative harboring a PNII::lacZ translational fusion | This work |
| pJCD01 | Apr, oriColE1, polylinker of pUC19 flanked by rpoC and rrnBT1T2 terminators | [32] |
| pJCD-PN | Apr, pJCD01 derivative harboring a 585-bp EcoRI fragment that includes the PN promoter | [24] |
| pJCD-PNI | Apr, pJCD01 derivative harboring a 139-bp EcoRI fragment that includes the PNI promoter | This work |
| pJCD-PNII | Apr, pJCD01 derivative harboring a 209-bp EcoRI fragment that includes the PNII promoter | This work |
| pSJ3-PN | Apr, pSJ3 derivative harboring a PN::lacZ translational fusion | [20] |
| pSJ3-PNI | Apr, pSJ3 derivative harboring a PNI::lacZ translational fusion | This work |
| pSJ3-PNII | Apr, pSJ3 derivative harboring a PNII::lacZ translational fusion | This work |
| pCK01 | Cmr, oripSC101, low copy number cloning vector, polylinker flanked by NotI sites | [33] |
| pCK01-BzdR | Cmr, pCK01 derivative harboring a DNA fragment containing the bzdR gene | [20] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durante-Rodríguez, G.; Gutiérrez-del-Arroyo, P.; Vélez, M.; Díaz, E.; Carmona, M. Further Insights into the Architecture of the PN Promoter That Controls the Expression of the bzd Genes in Azoarcus. Genes 2019, 10, 489. https://doi.org/10.3390/genes10070489
Durante-Rodríguez G, Gutiérrez-del-Arroyo P, Vélez M, Díaz E, Carmona M. Further Insights into the Architecture of the PN Promoter That Controls the Expression of the bzd Genes in Azoarcus. Genes. 2019; 10(7):489. https://doi.org/10.3390/genes10070489
Chicago/Turabian StyleDurante-Rodríguez, Gonzalo, Paloma Gutiérrez-del-Arroyo, Marisela Vélez, Eduardo Díaz, and Manuel Carmona. 2019. "Further Insights into the Architecture of the PN Promoter That Controls the Expression of the bzd Genes in Azoarcus" Genes 10, no. 7: 489. https://doi.org/10.3390/genes10070489
APA StyleDurante-Rodríguez, G., Gutiérrez-del-Arroyo, P., Vélez, M., Díaz, E., & Carmona, M. (2019). Further Insights into the Architecture of the PN Promoter That Controls the Expression of the bzd Genes in Azoarcus. Genes, 10(7), 489. https://doi.org/10.3390/genes10070489