Research Progress on Plant RING-Finger Proteins


Abstract
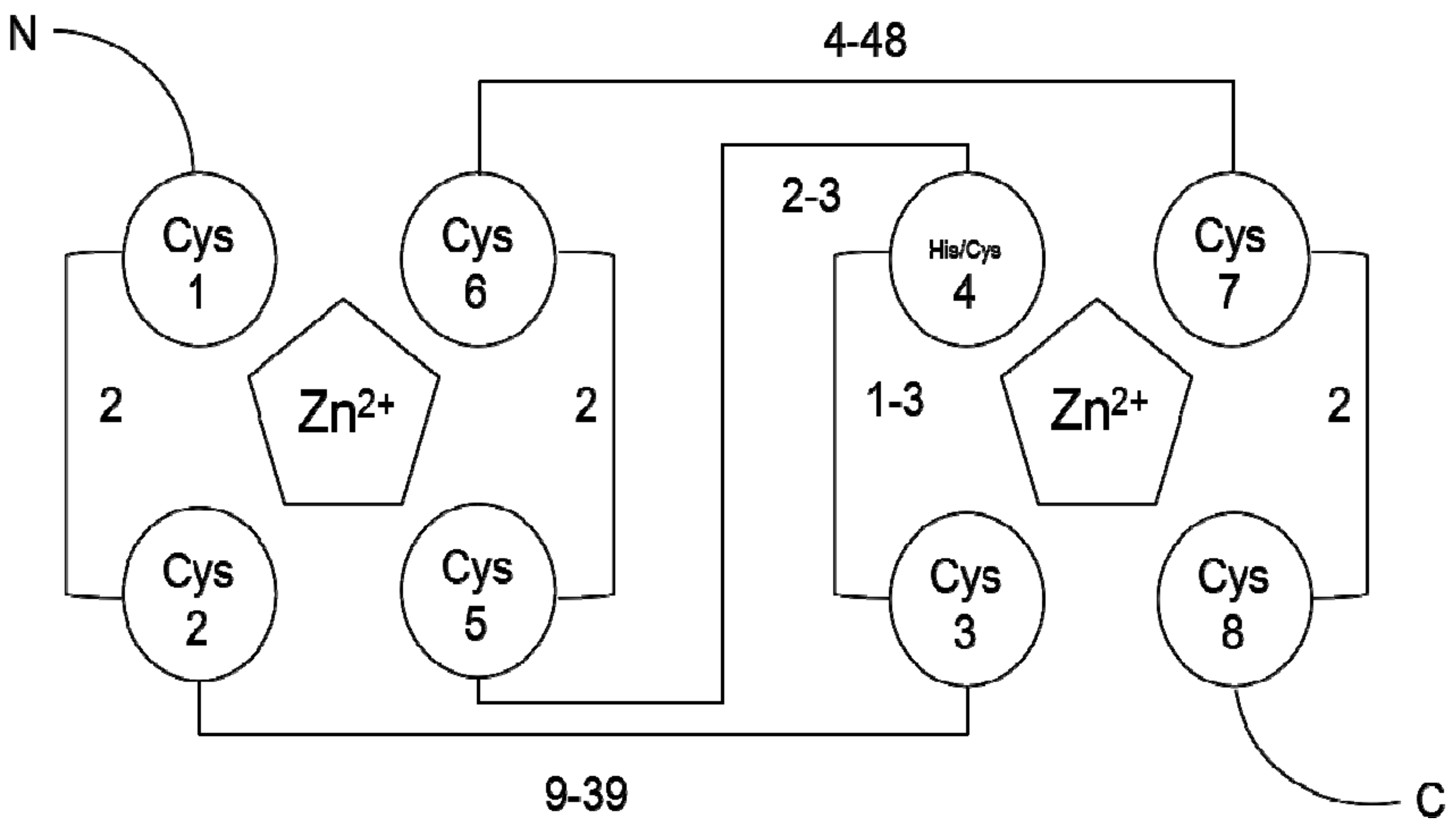
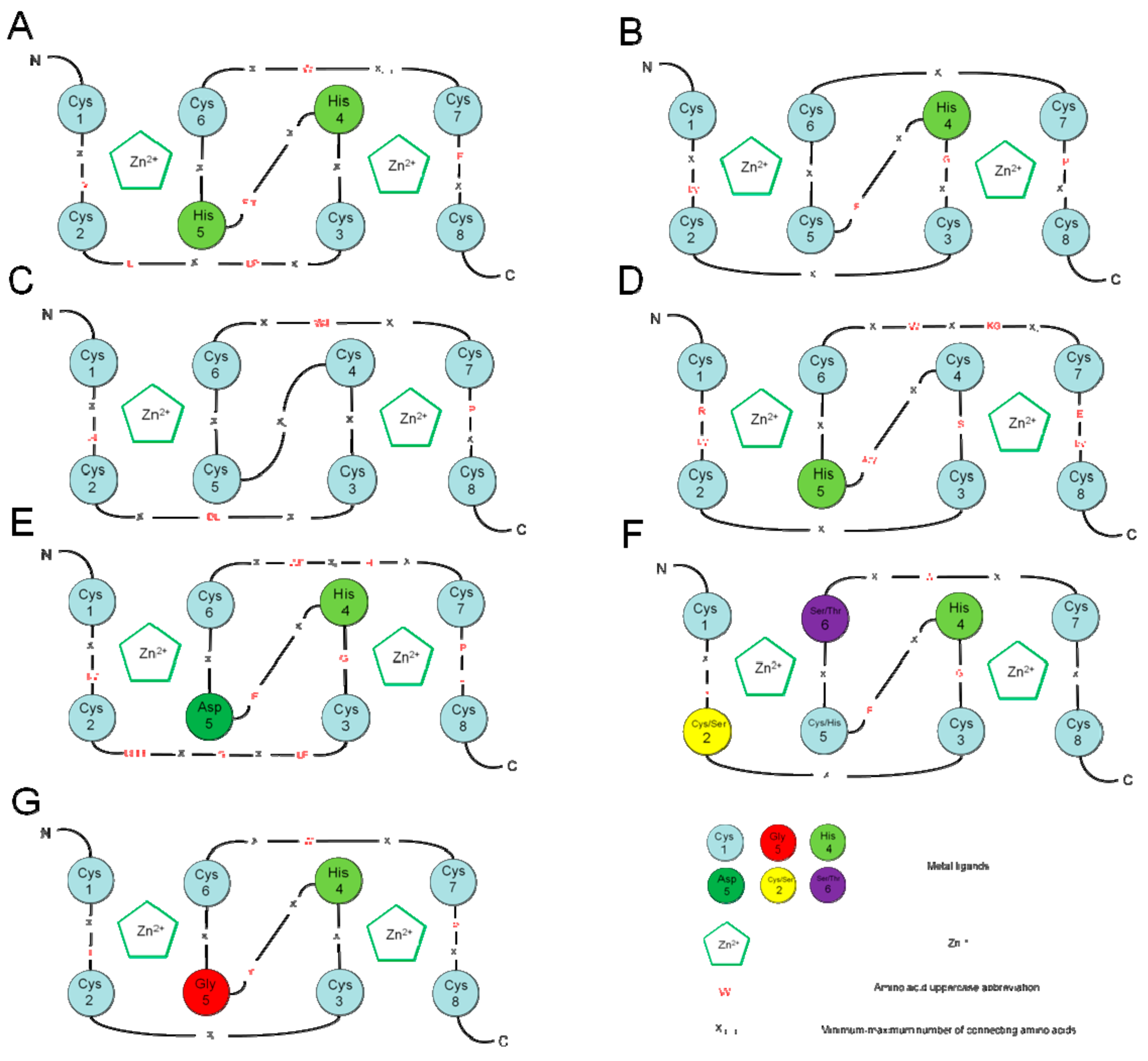
1. Introduction
2. RING-Finger Protein Subcellular Localization
3. RING-Finger Protein Functions
3.1. RING-Finger Proteins Are Involved in Plant Growth and Development
3.2. RING-Finger Proteins Are Involved in Plant Stress Resistance
3.2.1. RING-Finger Proteins Are Involved in Plant Drought Resistance
3.2.2. RING-Finger Proteins Are Involved in Salt and Aluminium Resistance
3.2.3. RING-Finger Proteins Are Involved in Temperature Stress
3.2.4. RING-Finger Proteins Are Involved in Biotic Stress
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Glickman, M.H.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef]
- Borden, K.L. RING domains: Master builders of molecular scaffolds? J. Mol. Biol. 2000, 295, 1103–1112. [Google Scholar] [CrossRef]
- Fleury, D.; Himanen, K.; Cnops, G.; Nelissen, H.; Boccardi, T.M.; Maere, S.; Beemster, G.T.; Neyt, P.; Anami, S.; Robles, P.; et al. The Arabidopsis thaliana homolog of yeast BRE1 has a function in cell cycle regulation during early leaf and root growth. Plant Cell 2007, 19, 417–432. [Google Scholar] [CrossRef]
- Zeba, N.; Isbat, M.; Kwon, N.J.; Lee, M.O.; Kim, S.R.; Hong, C.B. Heat-inducible C3HC4 type RING zinc finger protein gene from Capsicum annuum enhances growth of transgenic tobacco. Planta 2009, 229, 861–871. [Google Scholar] [CrossRef]
- Yang, Y.; Fu, D.; Zhu, C.; He, Y.; Zhang, H.; Liu, T.; Li, X.; Wu, C. The RING-Finger Ubiquitin Ligase HAF1 Mediates Heading date 1 Degradation during Photoperiodic Flowering in Rice. Plant Cell 2015, 27, 2455–2468. [Google Scholar] [CrossRef]
- Li, W.T.; He, M.; Wang, J.; Wang, Y.P. Zinc finger protein (ZFP) in plants-A review. Plant Omics 2013, 6, 474–480. [Google Scholar] [CrossRef]
- Kosarev, P.; Mayer, K.F.; Hardtke, C.S. Evaluation and classification of RING-finger domains encoded by the Arabidopsis genome. Genome Biol. 2002, 3, 0016.1–0016.12. [Google Scholar] [CrossRef] [PubMed]
- Metzger, M.B.; Pruneda, J.N.; Klevit, R.E.; Weissman, A.M. RING-type E3 ligases: Master manipulators of E2 ubiquitin-conjugating enzymes and ubiquitination. Biochim. Biophys. Acta 2014, 1843, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Hauksdottir, H.; Troy, A.; Herschleb, J.; Kraft, E.; Callis, J. Functional analysis of the RING-type ubiquitin ligase family of Arabidopsis. Plant Physiol. 2005, 137, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Freemont, P.S.; Hanson, I.M.; Trowsdale, J. A novel cysteine-rich sequence motif. Cell 1991, 64, 483–484. [Google Scholar] [CrossRef]
- Borden, K.L.; Freemont, P.S. The RING finger domain: A recent example of a sequence-structure family. Curr. Opin. Struct. Biol. 1996, 6, 395–401. [Google Scholar] [CrossRef]
- Jimenez-Lopez, D.; Munoz-Belman, F.; Gonzalez-Prieto, J.M.; Aguilar-Hernandez, V.; Guzman, P. Repertoire of plant ring e3 ubiquitin ligases revisited: New groups counting gene families and single genes. PLoS ONE 2018, 13, e0203442. [Google Scholar] [CrossRef]
- Hong, J.K.; Choi, H.W.; Hwang, I.S.; Hwang, B.K. Role of a novel pathogen-induced pepper C3-H-C4 type RING-finger protein gene, CaRFPI, in disease susceptibility and osmotic stress tolerance. Plant Mol. Biol. 2007, 63, 571–588. [Google Scholar] [CrossRef] [PubMed]
- Guzman, P. ATLs and BTLs, plant-specific and general eukaryotic structurally-related E3 ubiquitin ligases. Plant Sci. Int. J. Exp. Plant Biol. 2014, 215, 69–75. [Google Scholar] [CrossRef]
- Lim, S.D.; Yim, W.C.; Moon, J.C.; Kim, D.S.; Lee, B.M.; Jang, C.S. A gene family encoding RING finger proteins in rice: Their expansion, expression diversity, and co-expressed genes. Plant Mol. Biol. 2010, 72, 369–380. [Google Scholar] [CrossRef]
- Lim, S.D.; Hwang, J.G.; Jung, C.G.; Hwang, S.G.; Moon, J.C.; Jang, C.S. Comprehensive analysis of the rice RING E3 ligase family reveals their functional diversity in response to abiotic stress. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2013, 20, 299–314. [Google Scholar] [CrossRef]
- Liu, Q.G.; Yang, J.L.; Wang, Z.C.; Xu, X.M.; Mao, X.L.; Li, D.D.; Hu, X.Q.; Jin, D.C.; Li, C.H. Genome-wide Classification, Identification and Expression Profile of the C3HC4-type RING Finger Gene Family in Poplar (Populus trichocarpa). Plant Mol. Biol. Rep. 2015, 33, 1740–1754. [Google Scholar] [CrossRef]
- Alam, I.; Yang, Y.Q.; Wang, Y.; Zhu, M.L.; Wang, H.B.; Chalhoub, B.; Lu, Y.H. Genome-wide identification, evolution and expression analysis of RING finger protein genes in Brassica rapa. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Yang, Y.Q.; Lu, Y.H. Genome-wide survey, characterization, and expression analysis of RING finger protein genes in Brassica oleracea and their syntenic comparison to Brassica rapa and Arabidopsis thaliana. Genome 2018, 61, 685–697. [Google Scholar] [CrossRef]
- Li, Y.; Wu, B.; Yu, Y.; Yang, G.; Wu, C.; Zheng, C. Genome-wide analysis of the RING finger gene family in apple. Mol. Genet. Genom. MGG 2011, 286, 81–94. [Google Scholar] [CrossRef]
- Yang, L.; Miao, M.; Lyu, H.; Cao, X.; Li, J.; Li, Y.; Li, Z.; Chang, W. Genome wide indentification, evolution and expression analysis of RING-finger gene family in Solanum lycopersicum. Int. J. Mol. Sci. 2019, 30, 4864. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, M.Y.; Zhao, J.; Zhang, Y.C.; Xie, Q.J.; Chen, D.H. Genome-wide analysis of RING finger proteins in the smallest free-living photosynthetic eukaryote Ostreococus tauri. Mar. Genom. 2016, 26, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Ni, M. RED AND FAR-RED INSENSITIVE 2, a RING-domain zinc finger protein, mediates phytochrome-controlled seedling deetiolation responses. Plant Physiol. 2006, 140, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Wang, L.; Xu, Y.; Chen, N.; Ma, Q.; Li, F.; Chong, K. Overexpression of OsCOIN, a putative cold inducible zinc finger protein, increased tolerance to chilling, salt and drought, and enhanced proline level in rice. Planta 2007, 226, 1007–1016. [Google Scholar] [CrossRef]
- Xia, Z.; Liu, Q.; Wu, J.; Ding, J. ZmRFP1, the putative ortholog of SDIR1, encodes a RING-H2 E3 ubiquitin ligase and responds to drought stress in an ABA-dependent manner in maize. Gene 2012, 495, 146–153. [Google Scholar] [CrossRef]
- Lee, H.K.; Cho, S.K.; Son, O.; Xu, Z.; Hwang, I.; Kim, W.T. Drought stress-induced Rma1H1, a RING membrane-anchor E3 ubiquitin ligase homolog, regulates aquaporin levels via ubiquitination in transgenic Arabidopsis plants. Plant Cell 2009, 21, 622–641. [Google Scholar] [CrossRef]
- Lim, S.D.; Cho, H.Y.; Park, Y.C.; Ham, D.J.; Lee, J.K.; Jang, C.S. The rice RING finger E3 ligase, OsHCI1, drives nuclear export of multiple substrate proteins and its heterogeneous overexpression enhances acquired thermotolerance. J. Exp. Bot. 2013, 64, 2899–2914. [Google Scholar] [CrossRef]
- Gao, W.; Liu, W.; Zhao, M.; Li, W.X. NERF encodes a RING E3 ligase important for drought resistance and enhances the expression of its antisense gene NFYA5 in Arabidopsis. Nucleic Acids Res. 2015, 43, 607–617. [Google Scholar] [CrossRef]
- Kim, J.H.; Lim, S.D.; Jang, C.S. Oryza sativa heat-induced RING finger protein 1 (OsHIRP1) positively regulates plant response to heat stress. Plant Mol. Biol. 2019, 99, 545–559. [Google Scholar] [CrossRef]
- Ryu, M.Y.; Cho, S.K.; Kim, W.T. The Arabidopsis C3H2C3-type RING E3 ubiquitin ligase AtAIRP1 is a positive regulator of an abscisic acid-dependent response to drought stress. Plant Physiol. 2010, 154, 1983–1997. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Q.; Liu, Z.; Yang, H.; Wang, J.; Li, X.; Yang, Y. Arabidopsis C3HC4-RING finger E3 ubiquitin ligase AtAIRP4 positively regulates stress-responsive abscisic acid signaling. J. Integr. Plant Biol. 2016, 58, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.M.; Huang, S.; Liu, Y.Q.; Bian, M.D.; Shi, W.L.; Zuo, Z.C.; Yang, Z.M. Overexpression of A RING finger ubiquitin ligase gene AtATRF1 enhances aluminium tolerance in Arabidopsis thaliana. J. Plant Biol. 2017, 60, 66–74. [Google Scholar] [CrossRef]
- Berrocal-Lobo, M.; Stone, S.; Yang, X.; Antico, J.; Callis, J.; Ramonell, K.M.; Somerville, S. ATL9, a RING zinc finger protein with E3 ubiquitin ligase activity implicated in chitin- and NADPH oxidase-mediated defense responses. PLoS ONE 2010, 5, e14426. [Google Scholar] [CrossRef]
- Disch, S.; Anastasiou, E.; Sharma, V.K.; Laux, T.; Fletcher, J.C.; Lenhard, M. The E3 ubiquitin ligase BIG BROTHER controls arabidopsis organ size in a dosage-dependent manner. Curr. Biol. 2006, 16, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, E.; Ge, X.; Gong, Q.; Butt, H.; Zhang, C.; Yang, Z.; Li, F.; Zhang, X. Overexpressed BRH1, a RING finger gene, alters rosette leaf shape in Arabidopsis thaliana. Sci. China Life Sci. 2018, 61, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Hardtke, C.S.; Okamoto, H.; Stoop-Myer, C.; Deng, X.W. Biochemical evidence for ubiquitin ligase activity of the Arabidopsis COP1 interacting protein 8 (CIP8). Plant J. Cell Mol. Biol. 2002, 30, 385–394. [Google Scholar] [CrossRef]
- Deng, X.W.; Caspar, T.; Quail, P.H. cop1: A regulatory locus involved in light-controlled development and gene expression in Arabidopsis. Genes Dev. 1991, 5, 1172–1182. [Google Scholar] [CrossRef]
- Von Arnim, A.G.; Deng, X.W. Ring finger motif of Arabidopsis thaliana COP1 defines a new class of zinc-binding domain. J. Biol. Chem. 1993, 268, 19626–19631. [Google Scholar]
- Qin, F.; Sakuma, Y.; Tran, L.S.; Maruyama, K.; Kidokoro, S.; Fujita, Y.; Fujita, M.; Umezawa, T.; Sawano, Y.; Miyazono, K.; et al. Arabidopsis DREB2A-interacting proteins function as RING E3 ligases and negatively regulate plant drought stress-responsive gene expression. Plant Cell 2008, 20, 1693–1707. [Google Scholar] [CrossRef]
- Park, J.H.; Kang, C.H.; Nawkar, G.M.; Lee, E.S.; Paeng, S.K.; Chae, H.B.; Chi, Y.H.; Kim, W.Y.; Yun, D.J.; Lee, S.Y. EMR, a cytosolic-abundant ring finger E3 ligase, mediates ER-associated protein degradation in Arabidopsis. New Phytol. 2018, 220, 163–177. [Google Scholar] [CrossRef]
- Voiniciuc, C.; Dean, G.H.; Griffiths, J.S.; Kirchsteiger, K.; Hwang, Y.T.; Gillett, A.; Dow, G.; Western, T.L.; Estelle, M.; Haughn, G.W. Flying saucer1 is a transmembrane RING E3 ubiquitin ligase that regulates the degree of pectin methylesterification in Arabidopsis seed mucilage. Plant Cell 2013, 25, 944–959. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, H.J.; Park, C.M. HOS1 acts as a key modulator of hypocotyl photomorphogenesis. Plant Signal. Behav. 2017, 12, e1315497. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Xiong, L.; Gong, Z.; Ishitani, M.; Stevenson, B.; Zhu, J.K. The Arabidopsis HOS1 gene negatively regulates cold signal transduction and encodes a RING finger protein that displays cold-regulated nucleo--cytoplasmic partitioning. Genes Dev. 2001, 15, 912–924. [Google Scholar] [CrossRef] [PubMed]
- Marino, D.; Froidure, S.; Canonne, J.; Ben Khaled, S.; Khafif, M.; Pouzet, C.; Jauneau, A.; Roby, D.; Rivas, S. Arabidopsis ubiquitin ligase MIEL1 mediates degradation of the transcription factor MYB30 weakening plant defence. Nat. Commun. 2013, 4, 1476. [Google Scholar] [CrossRef]
- Chen, M.; Ni, M. RFI2, a RING-domain zinc finger protein, negatively regulates CONSTANS expression and photoperiodic flowering. Plant J. Cell Mol. Biol. 2006, 46, 823–833. [Google Scholar] [CrossRef]
- Cheng, M.C.; Hsieh, E.J.; Chen, J.H.; Chen, H.Y.; Lin, T.P. Arabidopsis RGLG2, functioning as a RING E3 ligase, interacts with AtERF53 and negatively regulates the plant drought stress response. Plant Physiol. 2012, 158, 363–375. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, C.; Li, Y.; Zheng, N.; Chen, H.; Zhao, Q.; Gao, T.; Guo, H.; Xie, Q. SDIR1 is a RING finger E3 ligase that positively regulates stress-responsive abscisic acid signaling in Arabidopsis. Plant Cell 2007, 19, 1912–1929. [Google Scholar] [CrossRef]
- Xie, Q.; Guo, H.S.; Dallman, G.; Fang, S.; Weissman, A.M.; Chua, N.H. SINAT5 promotes ubiquitin-related degradation of NAC1 to attenuate auxin signals. Nature 2002, 419, 167–170. [Google Scholar] [CrossRef]
- Tian, M.; Lou, L.; Liu, L.; Yu, F.; Zhao, Q.; Zhang, H.; Wu, Y.; Tang, S.; Xia, R.; Zhu, B.; et al. The RING finger E3 ligase STRF1 is involved in membrane trafficking and modulates salt-stress response in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2015, 82, 81–92. [Google Scholar] [CrossRef]
- Ko, J.H.; Yang, S.H.; Han, K.H. Upregulation of an Arabidopsis RING-H2 gene, XERICO, confers drought tolerance through increased abscisic acid biosynthesis. Plant J. Cell Mol. Biol. 2006, 47, 343–355. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, H.; Yang, Y.; Li, G.; Yang, Y.; Wang, X.; Basnayake, B.M.; Li, D.; Song, F. Functional analysis reveals pleiotropic effects of rice RING-H2 finger protein gene OsBIRF1 on regulation of growth and defense responses against abiotic and biotic stresses. Plant Mol. Biol. 2008, 68, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Jantasuriyarat, C.; Zhao, Q.; Zhang, H.; Chen, S.; Liu, J.; Liu, L.; Tang, S.; Park, C.H.; Wang, X.; et al. The SINA E3 ligase OsDIS1 negatively regulates drought response in rice. Plant Physiol. 2011, 157, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Park, G.G.; Park, J.J.; Yoon, J.; Yu, S.N.; An, G. A RING finger E3 ligase gene, Oryza sativa Delayed Seed Germination 1 (OsDSG1), controls seed germination and stress responses in rice. Plant Mol. Biol. 2010, 74, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.H.; Liu, C.C.; Wu, S.J.; Kuo, Y.Y.; Lu, C.A.; Wu, C.R.; Lian, P.J.; Hong, C.Y.; Ke, Y.T.; Huang, J.H.; et al. Expression of a gene encoding a rice RING zinc-finger protein, OsRZFP34, enhances stomata opening. Plant Mol. Biol. 2014, 86, 125–137. [Google Scholar] [CrossRef]
- Park, Y.C.; Chapagain, S.; Jang, C.S. The microtubule-associated RING finger protein 1 (OsMAR1) acts as a negative regulator for salt-stress response through the regulation of OCPI2 (O. sativa chymotrypsin protease inhibitor 2). Planta 2018, 247, 875–886. [Google Scholar] [CrossRef]
- Bae, H.; Kim, S.K.; Cho, S.K.; Kang, B.G.; Kim, W.T. Overexpression of OsRDCP1, a rice RING domain-containing E3 ubiquitin ligase, increased tolerance to drought stress in rice (Oryza sativa L.). Plant Sci. Int. J. Exp. Plant Biol. 2011, 180, 775–782. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, C.; Wei, C.; Liu, X.; Wang, M.; Yu, F.; Xie, Q.; Tu, J. The RING Finger Ubiquitin E3 Ligase OsHTAS Enhances Heat Tolerance by Promoting H2O2-Induced Stomatal Closure in Rice. Plant Physiol. 2016, 170, 429–443. [Google Scholar] [CrossRef]
- Park, Y.C.; Chapagain, S.; Jang, C.S. A Negative Regulator in Response to Salinity in Rice: Oryza sativa Salt-, ABA- and Drought-Induced RING Finger Protein 1 (OsSADR1). Plant Cell Physiol. 2018, 59, 575–589. [Google Scholar] [CrossRef]
- Park, Y.C.; Lim, S.D.; Moon, J.C.; Jang, C.S. A rice really interesting new gene H2-type E3 ligase, OsSIRH2-14, enhances salinity tolerance via ubiquitin/26S proteasome-mediated degradation of salt-related proteins. Plant Cell Environ. 2019, 42, 3061–3076. [Google Scholar] [CrossRef]
- Hwang, S.G.; Kim, J.J.; Lim, S.D.; Park, Y.C.; Moon, J.C.; Jang, C.S. Molecular dissection of Oryza sativa salt-induced RING Finger Protein 1 (OsSIRP1): Possible involvement in the sensitivity response to salinity stress. Physiol. Plant. 2016, 158, 168–179. [Google Scholar] [CrossRef]
- Lim, C.W.; Baek, W.; Lee, S.C. The Pepper RING-Type E3 Ligase CaAIRF1 Regulates ABA and Drought Signaling via CaADIP1 Protein Phosphatase Degradation. Plant Physiol. 2017, 173, 2323–2339. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.; Lim, C.W.; Lee, S.C. A pepper RING-type E3 ligase, CaASRF1, plays a positive role in drought tolerance via modulation of CaAIBZ1 stability. Plant J. Cell Mol. Biol. 2019, 98, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.W.; Baek, W.; Lee, S.C. Roles of pepper bZIP protein CaDILZ1 and its interacting partner RING-type E3 ligase CaDSR1 in modulation of drought tolerance. Plant J. Cell Mol. Biol. 2018, 96, 452–467. [Google Scholar] [CrossRef] [PubMed]
- Brugiere, N.; Zhang, W.; Xu, Q.; Scolaro, E.J.; Lu, C.; Kahsay, R.Y.; Kise, R.; Trecker, L.; Williams, R.W.; Hakimi, S.; et al. Overexpression of RING Domain E3 Ligase ZmXerico1 Confers Drought Tolerance through Regulation of ABA Homeostasis. Plant Physiol. 2017, 175, 1350–1369. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Xu, W.; Wang, J.; Wang, L.; Yao, W.; Yang, Y.; Xu, Y.; Ma, F.; Du, Y.; Wang, Y. The Chinese wild grapevine (Vitis pseudoreticulata) E3 ubiquitin ligase Erysiphe necator-induced RING finger protein 1 (EIRP1) activates plant defense responses by inducing proteolysis of the VpWRKY11 transcription factor. New Phytol. 2013, 200, 834–846. [Google Scholar] [CrossRef]
- Wang, L.; Xie, X.; Yao, W.; Wang, J.; Ma, F.; Wang, C.; Yang, Y.; Tong, W.; Zhang, J.; Xu, Y.; et al. RING-H2-type E3 gene VpRH2 from Vitis pseudoreticulata improves resistance to powdery mildew by interacting with VpGRP2A. J. Exp. Bot. 2017, 68, 1669–1687. [Google Scholar] [CrossRef]
- Liu, Y.; Li, L.; Zhang, L.; Lv, Q.; Zhao, Y.; Li, X. Isolation and identification of wheat gene TaDIS1 encoding a RING finger domain protein, which negatively regulates drought stress tolerance in transgenic Arabidopsis. Plant Sci. Int. J. Exp. Plant Biol. 2018, 275, 49–59. [Google Scholar] [CrossRef]
- Kam, J.; Gresshoff, P.; Shorter, R.; Xue, G.P. Expression analysis of RING zinc finger genes from Triticum aestivum and identification of TaRZF70 that contains four RING-H2 domains and differentially responds to water deficit between leaf and root. Plant Sci. 2007, 173, 650–659. [Google Scholar] [CrossRef]
- Yang, X.; Sun, C.; Hu, Y.; Lin, Z. Molecular cloning and characterization of a gene encoding RING zinc finger ankyrin protein from drought-tolerant Artemisia desertorum. J. Biosci. 2008, 33, 103–112. [Google Scholar] [CrossRef]
- Jung, Y.J.; Lee, I.H.; Nou, I.S.; Lee, K.D.; Rashotte, A.M.; Kang, K.K. BrRZFP1 a Brassica rapa C3HC4-type RING zinc finger protein involved in cold, salt and dehydration stress. Plant Biol. 2013, 15, 274–283. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, N.; Chen, P.; Gao, M.; Liu, J.; Wang, Y.; Zhao, T.; Li, Y.; Gai, J. Overexpression of a soybean ariadne-like ubiquitin ligase gene GmARI1 enhances aluminum tolerance in Arabidopsis. PLoS ONE 2014, 9, e111120. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cai, K.; Yin, J.; Chao, H.; Ren, Y.; Jin, L.; Cao, Y.; Duanmu, D.; Zhang, Z. A C3HC4-type RING finger protein regulates rhizobial infection and nodule organogenesis in Lotus japonicus. J. Integr. Plant Biol. 2018, 60, 878–896. [Google Scholar] [CrossRef] [PubMed]
- Karlowski, W.M.; Hirsch, A.M. The over-expression of an alfalfa RING-H2 gene induces pleiotropic effects on plant growth and development. Plant Mol. Biol. 2003, 52, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Dos Reis, S.P.; Tavares Lde, S.; Costa Cde, N.; Brigida, A.B.; de Souza, C.R. Molecular cloning and characterization of a novel RING zinc-finger protein gene up-regulated under in vitro salt stress in cassava. Mol. Biol. Rep. 2012, 39, 6513–6519. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.X.; Cheng, Z.W.; Liu, M.J.; Yang, X.F.; Qiu, D.W. C3HC4-Type RING Finger Protein NbZFP1 Is Involved in Growth and Fruit Development in Nicotiana benthamiana. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Su, X.; Liu, J.; Wang, M. The RING-H2 finger gene 1 (RHF1) encodes an E3 ubiquitin ligase and participates in drought stress response in Nicotiana tabacum. Genetica 2013, 141, 11–21. [Google Scholar] [CrossRef]
- Qi, S.L.; Lin, Q.F.; Zhu, H.S.; Gao, F.H.; Zhang, W.H.; Hua, X.J. The RING Finger E3 Ligase SpRing is a Positive Regulator of Salt Stress Signaling in Salt-Tolerant Wild Tomato Species. Plant Cell Physiol. 2016, 57, 528–539. [Google Scholar] [CrossRef]
- Dong, C.H.; Agarwal, M.; Zhang, Y.; Xie, Q.; Zhu, J.K. The negative regulator of plant cold responses, HOS1, is a RING E3 ligase that mediates the ubiquitination and degradation of ICE1. Proc. Natl. Acad. Sci. USA 2006, 103, 8281–8286. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene Expression and Signal Transduction in Water-Stress Response. Plant Physiol. 1997, 115, 327–334. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Gampala, S.S.; Rock, C.D. Abscisic acid signaling in seeds and seedlings. Plant Cell 2002, 14, S15–S45. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene Name | Plant Species | Type | Subcellular Localization | References |
|---|---|---|---|---|
| AtAIRP1 | A. thaliana | RING-HC | Cell membrane | Ryu M.Y. et al., 2010 [30] |
| AtAIRP4 | A. thaliana | RING-HC | Cytoplasm | Yang L. et al., 2016 [31] |
| AtATRF1 | A. thaliana | RING-HC | Nucleus | Qin X. et al., 2017 [32] |
| ATL9 | A. thaliana | RING-HC | Endoplasmic network | Beerocal M. et al., 2010 [33] |
| BIG BROTHER | A. thaliana | RING-H2 | Dish S. et al., 2006 [34] | |
| BRH1 | A. thaliana | RING-H2 | Wang X. et al., 2018 [35] | |
| CIP8 | A. thaliana | RING-H2 | Hardtke C.S. et al., 2002 [36] | |
| COP1 | A. thaliana | RING-HC | Nucleus | Deng X.W. et al., 1991, Von Arnim A.G. et al., 1993 [37,38] |
| DRIP1 | A. thaliana | RING-HC | Nucleus | Qin F. et al., 2008 [39] |
| DRIP2 | A. thaliana | RING-HC | Nucleus | Qin F. et al., 2008 [39] |
| EMR | A. thaliana | RING-HC | Cytoplasm | Park J.H. et al., 2018 [40] |
| FLY1 | A. thaliana | RING-H2 | Golgi apparatus | Voiniciuc C. et al., 2013 [41] |
| HOS1 | A. thaliana | RING-C2 | Nucleus | Lee H. et al.,2001, Dong C.H. et al., 2006, Kima J.H. et al., 2017 [42,43] |
| HUB1 | A. thaliana | RING-HC | Fleury D. et al., 2007 [3] | |
| MIEL1 | A. thaliana | RING-H2 | Nucleus | Marino D. et al., 2013 [44] |
| NERF | A. thaliana | RING-HC | Nucleus | Gao W. et al., 2015 [28] |
| RFI2 | A. thaliana | RING-H2 | Nucleus | Chen M.J. et al., 2006 [45] |
| RGLG2 | A. thaliana | RING-HC | Cell membrane | Cheng M.C. et al., 2012 [46] |
| Rma1 | A. thaliana | RING-HC | Endoplasmic network | Lee H.K. et al., 2009 [26] |
| SDIR1 | A. thaliana | RING-H2 | Cell membrane | Zhang Y.Y. et al., 2007 [47] |
| SINAT5 | A. thaliana | RING-HC | Nucleus | Xie Q. et al., 2002 [48] |
| STRF1 | A. thaliana | RING-H2 | Cytoplasm and cell membrane | Tian M.M. et al., 2015 [49] |
| XERICO | A. thaliana | RING-H2 | Ko J.H. et al., 2006 [50] | |
| OsBIRF1 | O. sativa | RING-H2 | Liu H.Z. et al., 2008 [51] | |
| OsCOIN | O. sativa | RING-C2 | Nuclear and cytoplasm | Liu K.M. et al., 2007 [24] |
| OsDIS1 | O. sativa | RING-H2 | Nucleus | Ning Y. et al., 2011 [52] |
| OsDSG1 | O. sativa | RING-H2 | Park G.G. et al., 2010 [53] | |
| OsHAF1 | O. sativa | RING-HC | Yang Y. et al., 2015 [5] | |
| OsHCI1 | O. sativa | RING-HC | Golgi apparatus | Lim S.D. et al., 2013 [27] |
| OsHIRP1 | O. sativa | RING-HC | Nucleus | Kim J.H. et al., 2019 [29] |
| OsHTAS | O. sativa | RING-H2 | Nuclear and cytoplasm | Liu J.P. et al., 2016 [54] |
| OsMAR1 | O. sativa | RING-H2 | Related to the microtubules | Park Y.C. et al., 2018 [55] |
| OsRDCP1 | O. sativa | RING-HC | Cell membrane | Bae H. et al., 2011 [56] |
| OsRZFP34 | O. sativa | RING-HC | Hus K.H. et al., 2014 [57] | |
| OsSADR1 | O. sativa | RING-H2 | Nucleus | Hwang S.G. et al., 2018 [58] |
| OsSIRH2-14 | O. sativa | RING-H2 | Cell membrane, cytoplasm, Golgi | Park Y.C. et al., 2019 [59] |
| OsSIRP1 | O. sativa | RING-HC | Cytoplasm | Hwang S.G. et al., 2016 [60] |
| CaAIRF1 | C. annuum L. | RING-HC | Nucleus | Lim C.W. et al., 2017 [61] |
| CaASRF1 | C. annuum L. | RING-H2 | Nuclear and cytoplasm | Joo H. et al., 2019 [62] |
| CaDSR1 | C. annuum L. | RING-H2 | Nuclear and cytoplasm | Lim C.W. et al., 2018 [63] |
| CaRFP1 | C. annuum L. | RING-HC | Hong J.K. et al., 2007 [13] | |
| CaRZFP1 | C. annuum L. | RING-HC | Zeba N. et al., 2009 [4] | |
| RmaIH1 | C. annuum L. | RING-HC | Endoplasmic network | Lee H.K. et al., 2009 [26] |
| ZmRFP1 | Z. mays L. | RING-H2 | Cell membrane | Xia Z.L. et al., 2012 [25] |
| ZmXerico1 | Z. mays L. | RING-H2 | Cytoplasm | Brugière N. et al., 2017 [64] |
| ZmXerico2 | Z. mays L. | RING-H2 | Cytoplasm | Brugière N. et al., 2017 [64] |
| EIRP1 | V. pseudoreticulata | RING-HC | Nucleus | Yu Y. et al., 2013 [65] |
| VpRH2 | V. pseudoreticulata | RING-H2 | Cytoplasm and cell membrane | Wan G.L. et al., 2017 [66] |
| TaDIS1 | T. aestivuml | RING-HC | Liu Y. et al., 2018 [67] | |
| TaRZF70 | T. aestivuml | RING-H2 | Kam J. et al., 2007 [68] | |
| AdZFP1 | A. dracunculus L. | RING-HC | YanG.X. et al., 2008 [69] | |
| BrRZFP1 | B. rapa | RING-HC | Cytoplasm and cell membrane | Jun Y.J. et al., 2013 [70] |
| GmARI1 | G. max | RING-HC | Nucleus | Zhang X.L. et al., 2014 [71] |
| LjCZF1 | L. japonicus | RING-HC | Cytoplasm and cell membrane | Cai K. et al., 2018 [72] |
| MsRH2-1 | M. sativa | RING-H2 | Karlowski W.M. et al., 2003 [73] | |
| MeRZF | M. esculenta | RING-H2 | Cell membrane | Reis S.P.D. et al., 2012 [74] |
| NbZFP1 | N. benthamiana | RING-HC | Chloroplast | Wu W.X. et al., 2014 [75] |
| NtRHF1 | N. tabacum | RING-H2 | Xia Z.L. et al., 2012 [76] | |
| SpRing | S. lycopersicum | RING-H2 | Endoplasmic network | Qi S.L. et al., 2016 [77] |
| Gene Name | AGI Loci | Protein | Function | References |
|---|---|---|---|---|
| BIG BROTHER | AT3g63530 | E3 ligase | regulator of Arabidopsis floral organ size | Dish S. et al., 2006 [34] |
| BRH1 | AT3g61460 | E3 ligase | alters rosette leaf shape | Wang X. et al., 2018 [35] |
| CIP8 | AT5g64920 | E3 ligase | regulator of photomorphogenesis | Hardtke C.S. et al., 2002 [36] |
| COP1 | AT2g32950 | E3 ligase | regulator of photomorphogenesis | Von Arnim A.G. et al., 1993 [38] |
| CaRZFP1 | regulator of root development | Zeba N. et al., 2009 [13] | ||
| EMR | AT4g26400 | E3 ligase | involved in the degradation of ER-associated protein | Park J.H. et al., 2018 [40] |
| FLY1 | AT4g28370 | E3 ligase | regulates the degree of pectin methylesterification in seed mucilage | Voiniciuc C. et al., 2013 [41] |
| HOS1 | AT2g39810 | E3 ligase | regulator of photomorphogenesis and flowering time | Lee H. et al. 2001, Kima J. Het al., 2017 [42,43] |
| HUB1 | AT2g44950 | E3 ligase | regulator of root development | Fleury D. et al., 2007 [3] |
| LjCZF1 | E3 ligase | a positive regulator of symbiotic nodulation | Cai K. et al., 2018 [72] | |
| MsRH2-1 | regulator of root and nodule development | Karlowski W.M. et al., 2003 [73] | ||
| NbZFP1 | regulator of fruit development, plant height, and leaf spacing | Wu W.X. et al., 2014 [75] | ||
| OsHAF1 | E3 ligase | regulator of photomorphogenesis | Park G.G. et al., 2010 [5] | |
| RFI2 | AT2g47700 | E3 ligase | regulator of flowering time | Chen M.J. et al., 2006 [45] |
| SINAT5 | AF480944 | E3 ligase | regulator of lateral root development | Xie Q. et al., 2002 [48] |
| Gene Name | AGI Loci | Protein | Function | References |
|---|---|---|---|---|
| AdZFP1 | regulator of plant tolerance to drought stress | Yang X. et al., 2008 [69] | ||
| AtAIRP1 | AT4G23450 | E3 ligase | regulator of plant tolerance to drought stress | Ryu M.Y. et al., 2010 [30] |
| AtAIRP4 | AT5G58787 | E3 ligase | regulator of plant tolerance to drought stress | Yang L. et al., 2016 [31] |
| AtATRF1 | E3 ligase | regulator of plant tolerance to drought stress | Qin X. et al., 2017 [32] | |
| ATL9 | AT2g35000 | E3 ligase | regulator of plant resistance to viable nutrient pathogens | Beerocaccobo M. et al., 2010 [33] |
| BrRZFP1 | regulator of plant tolerance to drought and cold stress | Jung Y.J. et al., 2013 [70] | ||
| CaAIRF1 | E3 ligase | regulator of plant tolerance to drought stress | Lim C.W. et al., 2017 [61] | |
| CaASRF1 | E3 ligase | regulator of plant tolerance to drought stress | Joo H. et al., 2019 [62] | |
| CaDSR1 | E3 ligase | regulator of plant tolerance to drought stress | Lim C.W. et al., 2018 [63] | |
| CaRFP1 | E3 ligase | regulator of plant tolerance to salt stress | Hong J.K. et al., 2007 [13] | |
| DRIP1 | E3 ligase | regulator of plant tolerance to drought stress | Qin F. et al., 2008 [39] | |
| DRIP2 | E3 ligase | regulator of plant tolerance to drought stress | Qin F. et al., 2008 [39] | |
| EIRP1 | E3 ligase | involved in pathogen defense | Yu Y. et al., 2013 [65] | |
| GmARI1 | E3 ligase | regulator of plant tolerance to Aluminium stress | Zhang X.L. et al., 2014 [73] | |
| HOS1 | AT2G39810 | E3 ligase | regulator of plant tolerance to cold stress | Dong C.H. et al., 2006 [78] |
| MeRZF | regulator of plant tolerance to salt stress | Reis S.P.D. et al., 2012 [74] | ||
| MIEL1 | AT5G18650 | E3 ligase | regulator of plant tolerance to biotic stress | Marino D. et al., 2013 [44] |
| NbZFP1 | regulator of plant tolerance to tobacco mosaic virus | Wu W.X. et al., 2014 [75] | ||
| NERF | E3 ligase | regulator of plant tolerance to drought stress | Gao W. et al., 2015 [28] | |
| NtRHF1 | E3 ligase | regulator of plant tolerance to drought stress | Xia Z.L. et al., 2012 [76] | |
| OsBIRF1 | regulator of plant tolerance to drought stress | Liu H.Z. et al., 2008 [51] | ||
| OsCOIN | regulator of plant tolerance to cold stress | Liu K.M. et al., 2007 [24] | ||
| OsDSG1 | E3 ligase | regulator of plant tolerance to drought stress | Liu K.M. et al., 2007 [53] | |
| OsHCI1 | E3 ligase | regulator of plant tolerance to high temperature stress | Lim S.D. et al., 2013 [27] | |
| OsHIRP1 | E3 ligase | regulator of plant tolerance to high temperature stress | Kim J.H. et al., 2019 [29] | |
| OsHTAS | E3 ligase | regulator of plant tolerance to high temperature stress | Liu J.P. et al., 2016 [54] | |
| OsMAR1 | E3 ligase | regulator of plant tolerance to salt stress | Park Y.C. et al., 2018 [55] | |
| OsRDCP1 | E3 ligase | regulator of plant tolerance to drought stress | Bae H. et al., 2011 [56] | |
| OsRZFP34 | regulator of plant tolerance to high temperature stress | Hus K.H. et al., 2014 [57] | ||
| OsSADR1 | E3 ligase | regulator of plant tolerance to drought stress | Hwang S.G. et al., 2018 [58] | |
| OsSIRH2-14 | E3 ligase | regulator of plant tolerance to salt stress | Park Y.C. et al., 2019 [59] | |
| OsSIRP1 | E3 ligase | regulator of plant tolerance to salt stress | Hwang S.G. et al., 2016 [60] | |
| RGLG2 | AT5G14420 | E3 ligase | regulator of plant tolerance to drought stress | Cheng M.C. et al., 2012 [46] |
| Rma1 | AT4G03510 | E3 ligase | regulator of plant tolerance to drought stress | Lee H.K. et al., 2009 [26] |
| RmaIH1 | E3 ligase | regulator of plant tolerance to drought stress | Lee H.K. et al., 2009 [26] | |
| SDIR1 | AT3G55530 | E3 ligase | regulator of plant tolerance to salt stress | Lee H.K. et al., 2009 [47] |
| SpRing | regulator of plant tolerance to salt stress | Qi S.L. et al., 2016 [77] | ||
| STRF1 | E3 ligase | regulator of plant tolerance to salt stress | Tian M.M. et al., 2015 [49] | |
| TaDIS1 | regulator of plant tolerance to drought stress | Liu Y. et al., 2018 [67] | ||
| TaRZF70 | regulator of plant tolerance to drought stress | Kam J. et al., 2007 [68] | ||
| VpRH2 | KU296022 | improves resistance to powdery mildew fungus | Wang L. et al., 2017 [66] | |
| XERICO | AT2G04240 | E3 ligase | regulator of plant tolerance to drought stress | Ko J.H. et al., 2006 [50] |
| ZmRFP1 | E3 ligase | regulator of plant tolerance to drought stress | Xia Z.L. et al., 2012 [25] | |
| ZmXerico1 | E3 ligase | regulator of plant tolerance to drought stress | Brugière N. et al., 2017 [64] | |
| ZmXerico2 | E3 ligase | regulator of plant tolerance to drought stress | Brugière N. et al., 2017 [64] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Sun, Y.; Ahmed, R.I.; Ren, A.; Xie, M. Research Progress on Plant RING-Finger Proteins. Genes 2019, 10, 973. https://doi.org/10.3390/genes10120973
Sun J, Sun Y, Ahmed RI, Ren A, Xie M. Research Progress on Plant RING-Finger Proteins. Genes. 2019; 10(12):973. https://doi.org/10.3390/genes10120973
Chicago/Turabian StyleSun, Jinhao, Yuhe Sun, Rana Imtiaz Ahmed, Angyan Ren, and Minmin Xie. 2019. "Research Progress on Plant RING-Finger Proteins" Genes 10, no. 12: 973. https://doi.org/10.3390/genes10120973
APA StyleSun, J., Sun, Y., Ahmed, R. I., Ren, A., & Xie, M. (2019). Research Progress on Plant RING-Finger Proteins. Genes, 10(12), 973. https://doi.org/10.3390/genes10120973