Daily Regulation of Phototransduction, Circadian Clock, DNA Repair, and Immune Gene Expression by Heme Oxygenase in the Retina of Drosophila

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
Animals and Procedures
3. Data Analysis
4. Results
4.1. Day-Night Differences in Gene Expression in the Retina of Drosophila
4.2. Effects of Ho Silencing on Gene Expression in the Retina of Drosophila
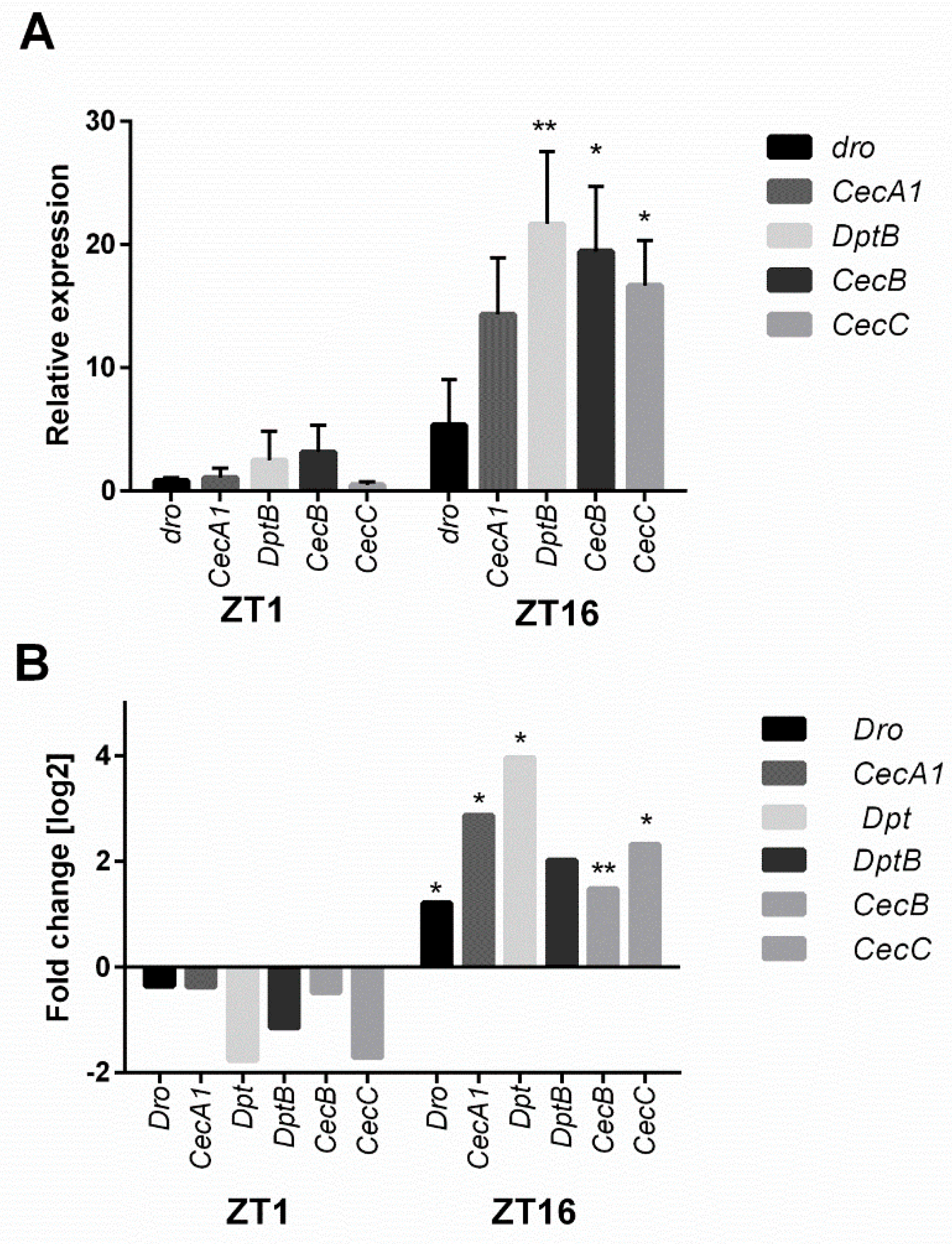
4.3. Confirmation of Microarray Data with the Real-Time PCR Method
5. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Byk, T.; Bar-Yaacov, M.; Doza, Y.N.; Minke, B.; Selinger, Z. Regulatory arrestin cycle secures the fidelity and maintenance of the fly photoreceptor cell. Proc. Natl. Acad. Sci. USA 1993, 90, 1907–1911. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Barber, T.R.; Xia, H.; Ready, D.F.; Hardie, R.C. Depletion of PtdIns(4,5)P2 underlies retinal degeneration in Drosophila trp mutants. J. Cell Sci. 2013, 126, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Shieh, B.H. Molecular genetics of retinal degeneration a Drosophila perspective. Fly 2011, 5, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Organisciak, D.T.; Vaughan, D.K. Retinal light damage: Mechanisms and protection. Prog. Retin. Eye Res. 2010, 29, 113–134. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.M.; Christianson, J.S.; Sapp, R.J.; Stark, W.S. Visual receptor cycle in normal and period mutant Drosophila: Microspectrophotometry, electrophysiology, and ultrastructural morphometry. Vis. Neurosci. 1992, 9, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Pyza, E.; Meinertzhagen, I.A. Circadian rhythms in screening pigment and invaginating organelles in photoreceptor terminals of the housefly’s first optic neuropile. J. Neurobiol. 1997, 32, 517–529. [Google Scholar] [CrossRef]
- Górska-Andrzejak, J.; Makuch, R.; Stefan, J.; Görlich, A.; Semik, D.; Pyza, E. Circadian expression of the presynaptic active zone protein bruchpilot in the lamina of Drosophila melanogaster. Dev. Neurobiol. 2013, 73, 14–26. [Google Scholar] [CrossRef]
- Lee, C.; Bae, K.; Edery, I. PER and TIM inhibit the DNA binding activity of a Drosophila CLOCK-CYC/dBMAL1 heterodimer without disrupting formation of the heterodimer: A basis for circadian transcription. Mol. Cell. Biol. 1999, 19, 5316–5325. [Google Scholar] [CrossRef]
- Ozturk, N.; Selby, C.P.; Annayev, Y.; Zhong, D.; Sancar, A. Reaction mechanism of Drosophila cryptochrome. Proc. Natl. Acad. Sci. USA 2011, 108, 516–521. [Google Scholar] [CrossRef]
- Ceriani, M.F.; Darlington, T.K.; Staknis, D.; Más, P.; Petti, A.A.; Weitz, C.J.; Kay, S.A. Light-dependent sequestration of TIMELESS by CRYPTOCHROME. Science 1999, 285, 553–556. [Google Scholar] [CrossRef]
- Naidoo, N.; Song, W.; Hunter-Ensor, M.; Sehgal, A. A role for the proteasome in the light response of the timeless clock protein. Science 1999, 285, 1737–1741. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Hardin, P.E. Drosophila photoreceptors contain an autonomous circadian oscillator that can function without period mRNA cycling. J. Neurosci. 1998, 18, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Damulewicz, M.; Loboda, A.; Bukowska-Strakova, K.; Jozkowicz, A.; Dulak, J.; Pyza, E. Clock and clock-controlled genes are differently expressed in the retina, lamina and in selected cells of the visual system of Drosophila melanogaster. Front. Cell. Neurosci. 2015, 9, 353. [Google Scholar] [CrossRef] [PubMed]
- Siwicki, K.K.; Eastman, C.; Petersen, G.; Rosbash, M.; Hall, J.C. Antibodies to the period gene product of Drosophila reveal diverse tissue distribution and rhythmic changes in the visual system. Neuron 1988, 1, 141–150. [Google Scholar] [CrossRef]
- Yoshii, T.; Todo, T.; Wülbeck, C.; Stanewsky, R.; Helfrich-Förster, C. Cryptochrome is present in the compound eyes and a subset of Drosophila’s clock neurons. J. Comp. Neurol. 2008, 508, 952–966. [Google Scholar] [CrossRef]
- Zerr, D.M.; Hall, J.C.; Rosbash, M.; Siwicki, K.K. Circadian fluctuations of period protein immunoreactivity in the CNS and the visual system of Drosophila. J. Neurosci. 1990, 10, 2749–2762. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Jazwa, A.; Grochot-Przeczek, A.; Rutkowski, A.J.; Cisowski, J.; Agarwal, A.; Jozkowicz, A.; Dulak, J. Heme oxygenase-1 and the vascular bed: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2008, 10, 1767–1812. [Google Scholar] [CrossRef]
- Cui, L.; Yoshioka, Y.; Suyari, O.; Kohno, Y.; Zhang, X.; Adachi, Y.; Ikehara, S.; Yoshida, T.; Yamaguchi, M.; Taketani, S. Relevant expression of Drosophila heme oxygenase is necessary for the normal development of insect tissues. Biochem. Biophys. Res. Commun. 2008, 377, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Damulewicz, M.; Loboda, A.; Jozkowicz, A.; Dulak, J.; Pyza, E. Haeme oxygenase protects against UV light DNA damages in the retina in clock-dependent manner. Sci. Rep. 2017, 7, 5197. [Google Scholar] [CrossRef]
- Ida, H.; Suyari, O.; Shimamura, M.; Tien Tai, T.; Yamaguchi, M.; Taketani, S. Genetic link between heme oxygenase and the signaling pathway of DNA damage in Drosophila melanogaster. Tohoku J. Exp. Med. 2013, 231, 117–125. [Google Scholar] [CrossRef]
- Damulewicz, M.; Loboda, A.; Jozkowicz, A.; Dulak, J.; Pyza, E. Interactions between the circadian clock and heme oxygenase in the retina of Drosophila melanogaster. Mol. Neurobiol. 2017, 54, 4953–4962. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.Q.; Liu, L.P.; Binari, R.; Hardy, R.; Shim, H.S.; Cavallaro, A.; Booker, M.; Pfeiffer, B.D.; Markstein, M.; Wang, H.; et al. A Drosophila resource of transgenic RNAi lines for neurogenetics. Genetics 2009, 182, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Poudel, S.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. PANTHER version 10: Expanded protein families and functions, and analysis tools. Nucleic Acids Res. 2016, 44, D336–D342. [Google Scholar] [CrossRef] [PubMed]
- Claridge-Chang, A.; Wijnen, H.; Naef, F.; Boothroyd, C.; Rajewsky, N.; Young, M.W. Circadian regulation of gene expression systems in the Drosophila head. Neuron 2001, 32, 657–671. [Google Scholar] [CrossRef]
- McDonald, M.J.; Rosbash, M. Microarray analysis and organization of circadian gene expression in Drosophila. Cell 2001, 107, 567–578. [Google Scholar] [CrossRef]
- Ueda, H.R.; Matsumoto, A.; Kawamura, M.; Iino, M.; Tanimura, T.; Hashimoto, S. Genome-wide transcriptional orchestration of circadian rhythms in Drosophila. J. Biol. Chem. 2002, 277, 14048–14052. [Google Scholar] [CrossRef]
- Ceriani, M.F.; Hogenesch, J.B.; Yanovsky, M.; Panda, S.; Straume, M.; Kay, S.A. Genome-wide expression analysis in Drosophila reveals genes controlling circadian behavior. J. Neurosci. 2002, 22, 9305–9319. [Google Scholar] [CrossRef]
- Lin, Y.; Han, M.; Shimada, B.; Wang, L.; Gibler, T.M.; Amarakone, A.; Awad, T.A.; Stormo, G.D.; Van Gelder, R.N.; Taghert, P.H. Influence of the period-dependent circadian clock on diurnal, circadian, and aperiodic gene expression in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2002, 99, 9562–9567. [Google Scholar] [CrossRef]
- Wijnen, H.; Naef, F.; Boothroyd, C.; Claridge-Chang, A.; Young, M.W. Control of daily transcript oscillations in Drosophila by light and the circadian clock. PLoS Genet. 2006, 2, 326–343. [Google Scholar] [CrossRef]
- Kula-Eversole, E.; Nagoshi, E.; Shang, Y.; Rodriguez, J.; Allada, R.; Rosbash, M. Surprising gene expression patterns within and between PDF-containing circadian neurons in Drosophila. Proc. Natl. Acad. Sci. USA 2010, 107, 13497–13502. [Google Scholar] [CrossRef] [PubMed]
- Hartman, S.J.; Menon, I.; Haug-Collet, K.; Colley, N.J. Expression of rhodopsin and arrestin during the light-dark cycle in Drosophila. Mol. Vis. 2001, 7, 95–100. [Google Scholar] [PubMed]
- Hardie, R.C.; Raghu, P. Visual transduction in Drosophila. Nature 2001, 413, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Mazzotta, G.; Rossi, A.; Leonardi, E.; Mason, M.; Bertolucci, C.; Caccin, L.; Spolaore, B.; Martin, A.J.M.; Schlichting, M.; Grebler, R.; et al. Fly cryptochrome and the visual system. Proc. Natl. Acad. Sci. USA 2013, 110, 6163–6168. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Emerson, M.; Su, H.S.; Sehgal, A. Response of the timeless protein to light correlates with behavioral entrainment and suggests a nonvisual pathway for circadian photoreception. Neuron 1998, 21, 215–223. [Google Scholar] [CrossRef]
- Loboda, A.; Jozkowicz, A.; Dulak, J. HO-1/CO system in tumor growth, angiogenesis and metabolism—Targeting HO-1 as an anti-tumor therapy. Vascul. Pharmacol. 2015, 74, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Weis, S.; Yang, G.; Weng, Y.H.; Helston, R.; Rish, K.; Smith, A.; Bordner, J.; Polte, T.; Gaunitz, F.; et al. Heme oxygenase-1 protein localizes to the nucleus and activates transcription factors important in oxidative stress. J. Biol. Chem. 2007, 282, 20621–20633. [Google Scholar] [CrossRef]
- Shieh, B.H.; Kristaponyte, I.; Hong, Y. Distinct roles of arrestin 1 protein in photoreceptors during Drosophila development. J. Biol. Chem. 2014, 289, 18526–18534. [Google Scholar] [CrossRef]
- Ekengren, S.; Hultmark, D. A family of Turandot-related genes in the humoral stress response of Drosophila. Biochem. Biophys. Res. Commun. 2001, 284, 998–1003. [Google Scholar] [CrossRef]
- Mecklenburg, K.L.; Takemori, N.; Komori, N.; Chu, B.; Hardie, R.C.; Matsumoto, H.; O’Tousa, J.E. Retinophilin is a light-regulated phosphoprotein required to suppress photoreceptor dark noise in Drosophila. J. Neurosci. 2010, 30, 1238–1249. [Google Scholar] [CrossRef]
- Ravi, D.; Wiles, A.M.; Bhavani, S.; Ruan, J.; Leder, P.; Bishop, A.J.R. A network of conserved damage survival pathways revealed by a genomic RNAi screen. PLoS Genet. 2009, 5. [Google Scholar] [CrossRef]
- Lee, J.E.; Edery, I. Circadian regulation in the ability of Drosophila to combat pathogenic infections. Curr. Biol. 2008, 18, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Shirasu-Hiza, M.M.; Dionne, M.S.; Pham, L.N.; Ayres, J.S.; Schneider, D.S. Interactions between circadian rhythm and immunity in Drosophila melanogaster. Curr. Biol. 2007, 17, 353–355. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.C.; Liao, C.W.; Pan, R.L.; Juang, J.L. Infection-induced intestinal oxidative stress triggers organ-to-organ immunological communication in Drosophila. Cell Host Microbe 2012, 11, 410–417. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cecB_For | CTTCGTCTTTGTGGCACTCATC |
| cecB_Rev | CTGACCAATGCGTTCGATTTT |
| dptB_For | TCCTGATCCCCGAGAGATTG |
| dptB_Rev | CAGATCGAATCCTTGCTTTGG |
| cecA1_For | GACCTCACTGCAATATCAATATCTTT |
| cecA1_Rev | GGTGATGGCCAGAATGAGAG |
| dro_For | ATTTGTCCACCACTCCAAGC |
| dro_Rev | GATGACTTCTCCGCGGTATG |
| cecC_For | CATCAGTCGCTCAGTTTCCA |
| cecC_Rev | TTCCCAGTCCTTGAATGGTT |
| trp_For | GCCCACCGAAATAACTACGA |
| trp_Rev | TATGCGTTGATCCTCGACTG |
| trpl_For | CCAAGAAACGAAAGGAGCAG |
| trpl_Rev | GAAACATCCATTCCGCTGTT |
| per_For | AAGAGCACCTTCTGCGTGAT |
| per_Rev | AGAATCTCGTCGGGAACCTT |
| norpA_For | CCACAACATGTCCTCGTTTG |
| norpA_Rev | TGGTTGAGTTGCATGGGTAA |
| rpl32_For | AGAAGCGCAAGGAGATTGTC |
| rpl32_Rev | ATGGTGCTGCTATCCCAATC |
| Biological Process | ZT1 | ZT16 | ||
|---|---|---|---|---|
| Upregulation | Downregulation | Upregulation | Down-Regulation | |
| Sensory Perception | Gr94a Ir92a CG12645 Lush Gr5a Obp69a | Obp83b Obp84a Obp28a Or49b Obp69a Gk | ||
| Transport | Porin2 ppk31 CG7458 Ndae1 CG30354 CG30345 Ykt6 CG5780 | CG1208 CG42260 spir Mppe | CG6125 CG15096 CG8051 Mdr50 inaF-C ppk22 | |
| Ubiquitination | Roc1b | |||
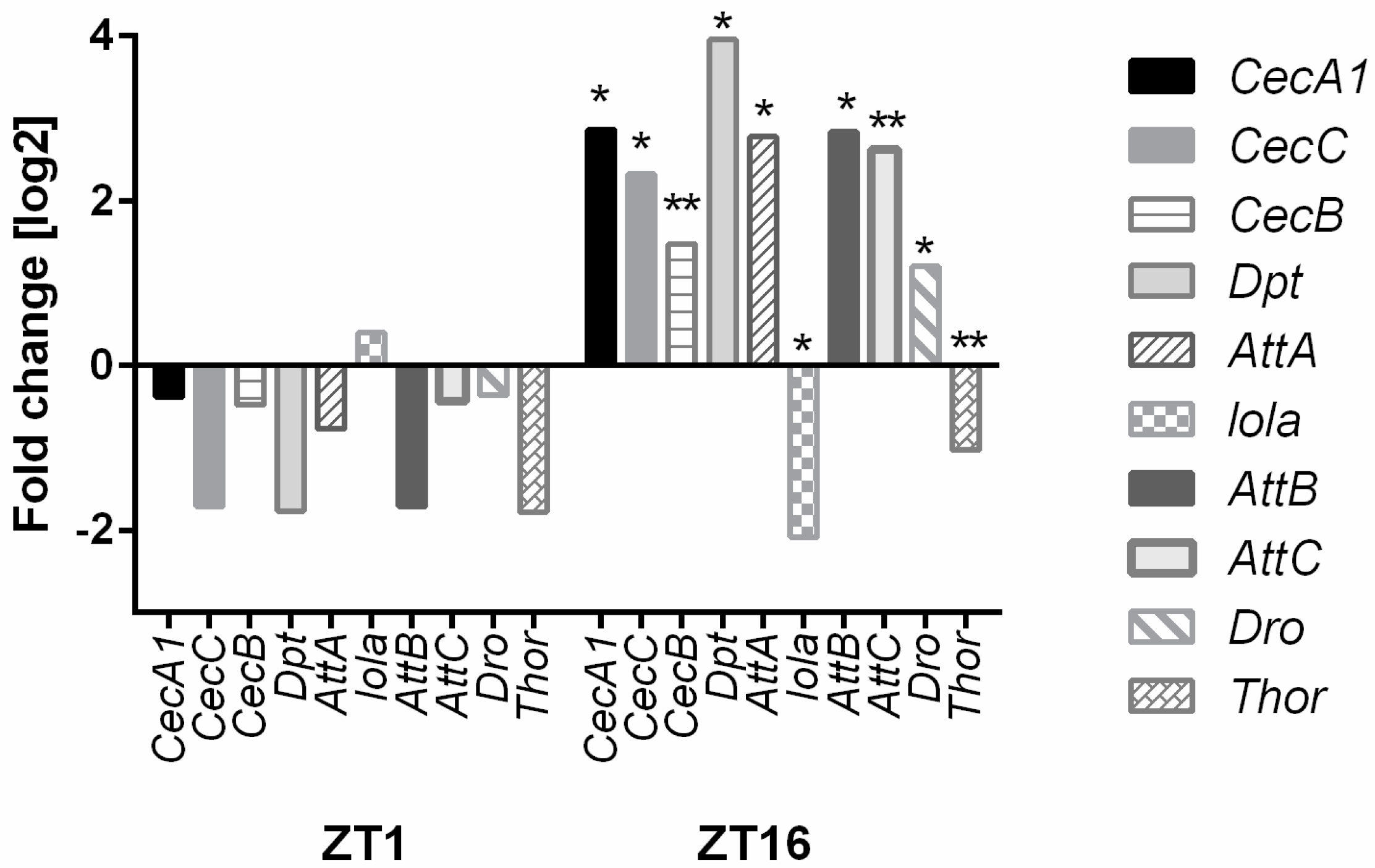
| Defense | Drs Rel CG10764 | Gb | AttD Dpt CecC Thor AttB AttA AttC CecA1 | |
| Translation | CG6094 CG10881 | |||
| Transcription | Hb Ubx CG9727 Vsx2 Sug H15 | CG18599 Samuel | lin-28 sug Crg-1 | |
| Metabolic Processes | CG17544 Oys | CG6432 CG11453 | CG11391 Pepck | |
| Oxido-reduction | CG10639 CG15864 CG9150 | Fbp2 CG6852 Cyp305a1 CG5653 | Prx2540-2 Cyp4e3 CG9629 CG10131 Sardh Cyp6a14 CG10512 | CG9747 Fmo-2 CG7724 |
| Wound Healing | Cht5 | Cht5 | ||
| Ca2+ Binding | CG30378 TpnC47D | CG4495 | ||
| Cytoskeleton | Ppcs | Sls Mwh Unc-115a | CG5023 | |
| Lipase | CG1986 CG10116 | CG10116 | ||
| DNA Damage | Corp | Pdk1 mus308 | ||
| Proteolysis | CG30287 | CG17571 CG5255 CG4914 | CG5849 CG4914 | |
| CuticleMetabolism | CG42494 Cpr67B | Cpr92F CG15754 Cpr5C | ||
| Endopeptidase Inhibitor | CG42464 | |||
| Response to Stress | CG11498 | |||
| Cell Cycle | CycB3 ana2 | ana2 Spc25 dgt3 Incenp | ||
| Signal Transduction | 5-HT2B Graf dlg1 hbs RhoGAP100F | Tsp26A | TyrRII LKR CCAP | Oamb Proc-R |
| Metal Ion Binding | CG42249 | CG18446 MtnD Amy-p | ||
| Phosphatase | Aph-4 | CG11425 | Aph-4 | |
| Hydrolase | CG7997 Tobi | Jhe | ||
| ATPase | CG9492 | CG11069 | ||
| Kinase | CkIIalpha-i3 CG15547 | |||
| Transferase | CG3038 | |||
| Nucleic Acid Binding | CG3165 | CG11563 CG7556 Adat | cid CG45050 qkr58E-1 | |
| Lipid Binding | CheB42b | |||
| Phagocytosis | Prx3 NimA | Prx3 | ||
| Zinc Binding | CG31053 RAF2 | CAH1 App | ||
| Phototransduction | Rh5 | |||
| Geotaxis | CG34353 | |||
| Clock | Cry | |||
| Response to ER Stress | CG42678 | |||
| Septate Junction | Tsp2A | |||
| Unknown | CG13742 CG43894 CG43391 scpr-C abd-A CG3611 CG43816 CG17564 CG15186 CG15140 CG32413 CG34279 CG13337 CG12688 CG44437 CG13449 CG4998 CG12321 CG44158 CG42869 CG11905 | CG10051 CG32548 CG45060 G6P CG44163 CG31415 CG13058 Osi24 CG13332 CG12034 CG43117 CG15728 pyd3 Sfp77F pre-mod(mdg4)-B CG17777 CG42272 scpr-B CG45603 CG11380 CG17378 | Os-C α5 CG17572 CG12239 CG11674 OS9 Tsp42Er CG4962 CG30271 a10 CG10332 Lsp1beta CG45061 pre-mod(mdg4)-T CG11585 MESR3 CG18170 CG13042 CG14075 Lsp1alpha CG43236 | CG13840 CG44434 CG11889 CG45307 Sfp77F CG17777 scpr-B CG17104 CG13931 CG8568 |
| GO Category ZT1 vs. ZT1 GMR>hoRNAi | Gene Count | p-Value | Genes |
|---|---|---|---|
| Locomotion (GO:0040011) | 27 | 1.46 E-01 | NetA, RhoGAP100F, CG34353, Ppcs, Trpm, fra, oys, ey, Nmda1, hb, Roc1b, CG3857, Tsp26A, how, Cry, cry, abd-A, w, Galphaq, trp, Ced-12, dlg1, sls, stum, Unc-115a, CG9317, klar, E(Pc) |
| Sensory Perception of Light (GO:0050953) | 5 | 3.37 E-01 | Rh5, cry, Cry, trp, CG9317 |
| Response to External Stimuli (GO:0009605) | 34 | 3.86 E-01 | NetA, RhoGAP100F, NimC1, Atg6, CG34353, Trpm, fra, Su(var)2-10, CG2051, Rh5, Nmda1, Drsl5, rdgB, CG3857, mthl8, trp, Cry, cry, CG13890, Drs, sug, w, Galphaq, Gr5a, dlg1, Diedel, gb, Sik2, Fuca, stum, Unc-115a, CG9317, E9Pc, Rel, srpk79D |
| Geotaxis (GO:0042332) Response to Gravity (GO:0009629) | 5 | 5.79 E-01 | CG34353, CG3857, Cry, cry, w, dlg1 |
| Genes Upregulated at ZT1 in GMR>hoRNAi Strain | Function |
| Arr1 | Phototransduction |
| CG17108 | Unknown |
| MntA | Metal homeostasis |
| eIF-4a | Response to DNA damage |
| Mt:Cyt-b | Electron transport |
| RpL13 | Translation |
| RpS25 | Translation |
| RplP2 | Translation |
| CG3457 | Unknown |
| IM2 | Defense response |
| RpL4 | Translation |
| CG4374 | Metal binding |
| To | Circadian clock |
| RpS27A | Translation |
| CG9821 | Unknown |
| Trf4-1 | DNA polymerase |
| Awd | Kinase |
| Spt20 | Histone acetylation |
| IM4 | Defense response |
| Ip259 | Phagocytosis |
| Genes Downregulated at ZT1in GMR>hoRNAi Strain | Function |
| Obp99b | Sensory perception |
| Nplp3 | Signaling pathway |
| Arr2 | Phototransduction |
| ninaE | Phototransduction |
| Pdh | Phototransduction |
| Mt:ATPase6 | ATPase |
| Ggamma30A | Phototransduction |
| mt:Col | Unknown |
| CG2233 | Unknown |
| Trp | Phototransduction |
| Obp44a | Sensory perception |
| RpL27A | Translation |
| Mt:ColII | Unknown |
| Nplp2 | Defense response |
| GstE12 | Detoxification |
| Rtp | Unknown |
| RpS28b | Translation |
| TotA | Response to UV, oxidative stress, bacterium |
| CG4842 | Oxido-reduction |
| Sea | Prevents chromosomal breaks |
| GO Category | Gene Count | p-Value | Genes |
|---|---|---|---|
| Antibacterial Humoral Response (GO:0019731) | 10 | 8.00 E-08 | CecA1, CecC, AttA, Thor, Dpt, CecB, AttB, AttC, CecA1, Dro |
| Antimicrobial Humoral Response (GO:0019730) Humoral Immune Response (GO: 0006959) | 11 | 1.67 E-05 4.25 E-04 | CecA1, CecC, AttA, Thor, Dpt, CecB, AttB, AttC, CecA1, Dro, lola |
| Defense Response to Gram-Positive Bacterium (GO:0050830) | 9 | 2.29 E-04 | AttD, CecA1, CecC, AttA, CecB, AttB, AttC, CecA1, Dro |
| Oxidation-Reduction Process (GO:0055114) | 26 | 2.18 E-02 | CG33099, CG9747, CG9629, Sardh, Cyp4e3, Lkr, CG18003, Cyp309a1, Prx3, CG10131, MtnD, Acox57D-d, Cyp6a14, CG10512, Cyp9h1, CG11236, CG18170, CG3397, CG14688, Prx2540-2, CG7724, Cyp4p2, CG14630, Sodh-2, Fmo-2, GILT3 |
| Cellular Macromolecule Metabolic Process (GO:0044260) | 17 | 5.03 E-02 | CG13085, Su(var)2-10, CG43143, Lkr, mus308, fng, qrk58E-1, MED23, S6k, Sse, Fak, RpS21, btl, p38c, Pdk1, Incenp, CG18853 |
| Single-Organism Metabolic Process (GO:0044710) | 46 | 1.40 E-01 | CG33099, CG9747, CG10184, CG9629, Sardh, Cyp4e3, Vkor, Lkr, CG18003, CG5379, Cyp309a2, Prx3, CG10131, Thor, CG10116, MtnD, Crz, Acox57D-d, Pepck, Cyp6a14, fng, CG10512, CG7059, CG6465, AcCoAS, dob, Cyp9h1, CG11236, CG18170, S6k, CG11425, CG3397, Jhe, CG6415, CG14688, Gnmt, Prx2540-2, CG7724, Sk1, ade3, Cyp4p2, CG14630, Sodh-2, Fmo-2, Lsd-1, GILT3 |
| Defense Response to Other Organism (GO:0098542) | 14 | 2.28 E-01 | AttD, Drsl6, CecA1, Su(var)2-10, CecC, AttA, Thor, Dpt, CecB, GNBP-like3, AttB, AttC, CecA1, Dro |
| Immune Response (GO:0006955) | 12 | 2.83 E-01 | CecA1, CecC, AttA,Thor, Dpt, CecB, AttB, AttC, PGRP-SC2, lola, Dro |
| Response to Bacterium (GO:0009617) | 13 | 3.05 E-01 | AttD, CecA1, Su(var)2-10, CecC, AttA, Thor, Dpt, CecB, AttB, AttC, CecA1, p38c, Dro |
| Response to External Biotic Stimulus (GO:0043207) | 16 | 3.07 E-01 | AttD, Drsl6, CecA1, Su(var)2-10, CecC, AttA, Thor, Dpt, CecB, GNBP-like3, AttB, AttC, CecA1, lola, p38c, Dro |
| Genes Upregulated at ZT16 in GMR>hoRNAi Strain | Function |
| Rh6 | Phototransduction |
| TotA | Response to UV, oxidative stress, bacterium |
| Trp | Phototransduction |
| RpS13 | Translation |
| Xport | Phototransduction |
| eIF-4a | DNA damage |
| RpL37A | Translation |
| CG4962 | Unknown |
| Rtp | Unknown |
| Ggamma30A | Phototransduction |
| Cam | DNA damage |
| RpS7 | Translation |
| RpS26 | Translation |
| CG4000 | Unknown |
| Hsc70-4 | Chaperon |
| RpL8 | Translation |
| RpS28b | Translation |
| CG1561 | Unknown |
| CG2016 | Unknown |
| RpLP1 | Translation |
| Genes Downregulated at ZT16 in GMR>hoRNAi Strain | Function |
| Obp99b | Sensory perception |
| Arr1 | Phototransduction |
| Arr2 | Phototransduction |
| CG17108 | Unknown |
| CG6503 | Unknown |
| Tsf1 | Defense response |
| Lsp2 | Defense response |
| mt:ATPase6 | ATPase |
| Nplp2 | Signaling pathway |
| Nplp3 | Signaling pathway |
| mt:Cyt-b | Oxido-reduction |
| CG2233 | Unknown |
| Adhr | Oxido-reduction |
| mt:Col | Unknown |
| Pepck | Glucose metabolism |
| RpLP2 | Translation |
| CG34166 | Unknown |
| mt:ND4 | Oxido-reduction |
| Noe | Unknown |
| CG30197 | Peptidase inhibitor |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damulewicz, M.; Świątek, M.; Łoboda, A.; Dulak, J.; Bilska, B.; Przewłocki, R.; Pyza, E. Daily Regulation of Phototransduction, Circadian Clock, DNA Repair, and Immune Gene Expression by Heme Oxygenase in the Retina of Drosophila. Genes 2019, 10, 6. https://doi.org/10.3390/genes10010006
Damulewicz M, Świątek M, Łoboda A, Dulak J, Bilska B, Przewłocki R, Pyza E. Daily Regulation of Phototransduction, Circadian Clock, DNA Repair, and Immune Gene Expression by Heme Oxygenase in the Retina of Drosophila. Genes. 2019; 10(1):6. https://doi.org/10.3390/genes10010006
Chicago/Turabian StyleDamulewicz, Milena, Michał Świątek, Agnieszka Łoboda, Józef Dulak, Bernadetta Bilska, Ryszard Przewłocki, and Elżbieta Pyza. 2019. "Daily Regulation of Phototransduction, Circadian Clock, DNA Repair, and Immune Gene Expression by Heme Oxygenase in the Retina of Drosophila" Genes 10, no. 1: 6. https://doi.org/10.3390/genes10010006
APA StyleDamulewicz, M., Świątek, M., Łoboda, A., Dulak, J., Bilska, B., Przewłocki, R., & Pyza, E. (2019). Daily Regulation of Phototransduction, Circadian Clock, DNA Repair, and Immune Gene Expression by Heme Oxygenase in the Retina of Drosophila. Genes, 10(1), 6. https://doi.org/10.3390/genes10010006