1. Introduction
The rapid recognition of living and dead cells in order to clear cell debris is important for the maintenance of the central nervous system (CNS) microenvironment and for immunological tolerance and resolution of inflammation. Microglia, the resident immune cells of the CNS, contribute in a major way to the regulation of inflammation [
1]. They function as a crucial defence against various types of insult to tissues and cells within the brain [
2,
3], with the ability to exist in either a resting state (ramified) or an active state (amoeboid) [
4]. In the case of inflammation, infection or trauma, microglia rapidly convert into an activated state and become phagocytic [
3]. Also, whilst in an active state, microglia can secrete both pro- and anti-inflammatory cytokines, the latter including interleukin 10 (IL-10) and transforming growth factor β (TGF-β) [
4,
5]. The anti-inflammatory cytokines are produced in order to antagonise the pro-inflammatory and immune responses in the CNS [
5,
6,
7]. The anti-inflammatory functions of IL-10 include blocking the activity of Th17 cells and inhibiting the production of pro-inflammatory cytokines such as TNF-α, IL-1 and IFN-γ [
8]. During the resolution phase of inflammation, TGF-β is upregulated, which enables the draining of leukocytes, tissue repair [
9,
10,
11], as well as myelination [
12]. Upon upregulation, through binding to TGF-β receptors and subsequent phosphorylation of Smad receptors, TGF-β eventually contributes to the suppression of inflammation [
9,
13,
14].
We and others have previously shown that the vitamin K-dependent protein Gas6 is an important modulator of cellular processes also in the CNS, such as cell survival, inflammation, proliferation, migration and removal of apoptotic cells and debris [
15,
16,
17]. Gas6 is a ligand for the TAM (Tyro3, Axl, Mer) receptor tyrosine kinases (RTKs) [
15,
16,
17]. Of major significance is also that Gas6–TAM signalling appears capable of regulating the immune system, including the activity of phagocytes such as macrophages and microglia. TAM triple-receptor-deficient mice suffer from a lymphoproliferative disorder that is accompanied by broad-spectrum autoimmunity [
17,
18]. TAM signalling has also been shown to play a role in oligodendrocyte survival [
19] as well as to enhance myelination in a cuprizone model of demyelination [
20]. Direct administration of Gas6 into the CNS was protective in experimental autoimmune encephalomyelitis, whilst deletion of Gas6 enhanced inflammation and delayed recovery [
21]. In addition, we have reported that Gas6–TAM signalling can protect from demyelination in an ex vivo demyelination model [
15]. These findings indicate Gas6 has the ability to negatively regulate neuroinflammation and the immune response as well as induce brain regeneration, all of which make it a potential therapeutic target for relevant disorders such as multiple sclerosis (MS). In the present study, we show that Gas6 exerts anti-inflammatory and pro-repair roles through upregulation, respectively, of IL-10 and TGF-β in the mouse optic nerve and in mixed glial cell cultures.
2. Materials and Methods
2.1. Animals
Wildtype and Tyro3−/−, Axl−/− and Mer−/− C57/BL6 mice, aged postnatal day 0 to 2 (P0–2), were used for mixed and primary glial cultures, whereas adult, 2-month old, mice were used for ex vivo experiments with optic nerve cultures. The three TAM knockout strains were obtained from The Jackson Laboratories, Bar Harbor, ME, USA. All experiments involving animals were performed in accordance with the Home Office Animals (Scientific Procedures) Act 1986 under a UK Home Office project licence (licence number PC2238199; 22/8/2017), as well as following approval from the University of Portsmouth ethics review committee (AWERB). Animals were killed humanely by cervical dislocation, and tissues were removed rapidly and placed in ice-cold saline prior to experiments.
2.2. Optic Nerve Culture
Mouse optic nerve cultures were set up for incubation over several days as previously described [
22]. Briefly, optic nerves with eyeball attached were removed from 2-month-old mice and placed in ice-cold artificial cerebrospinal fluid containing 124 mM NaCl, 2.5 mM KCl, 26 mM NaHCO
3, 2 mM MgSO
4, 1.25 mM KH
2PO
4, 10 mM glucose, 4 mM sucrose, 2.5 mM CaCl
2, pH 7.4. The tissue was placed onto semi-porous membrane inserts (0.4 mm; Millipore, Watford, UK) in a six-well culture plate and covered with 2 mL culture medium consisting of 50% Opti-MEM, 25% Hanks Balanced Salt Solution, 25% horse serum (Gibco Invitrogen, Paisley, UK), supplemented with 25 mM D-glucose (Sigma, Gillingham, UK) and antibiotics (penicillin G sodium 10,000 U/mL, streptomycin sulphate 1000 mg/mL; Gibco Invitrogen) at a 1:500 dilution. For experiments, recombinant Gas6 protein [
15] was added directly to the culture medium to a final concentration of 400 ng/mL; culture medium alone was used as control treatment. Treatments were replenished on the second day of incubation. After 3 days, optic nerves were detached from the eye, and the tissue was processed for subsequent applications. All experiments were repeated using cultures prepared from different animals at different times.
2.3. Cell Culture
2.3.1. Primary Mixed Glial Cell Culture
Mixed glial cell cultures were established from mice at postnatal ages P0–2 according to the method described by Mecha et al. [
23]. Briefly, after dissection of the brain and removal of the brain stem and cerebellum, the forebrain was placed into cold Dulbecco’s modified eagle medium (DMEM), and the meninges were removed under a dissecting microscope (VWR, Lutterworth, UK). The forebrain was transferred into a 50 mL falcon tube containing 1 mL cold DMEM, where it was triturated and dissociated using a serum-coated glass Pasteur pipette. The tube was centrifuged at 168×
g for 10 min. The pellet was re-suspended in warm DMEM 10:10:1 containing 10% foetal calf serum (FCS), 10% horse serum and 1% penicillin/streptomycin. Re-suspended cell pellets were added to tissue culture flasks or 24-well plates pre-coated with poly-D-lysine (Sigma). Cells were incubated at 37 °C in a humidified atmosphere of 5% CO
2 in air for minimum 10 days. Microscopic analysis of the cultures verified a consistent proportion of astrocytes/microglia across the various cultures. Following the incubation setup period, the cells were processed either for setting up pure glial cell cultures or for direct use in experiments, treating with recombinant Gas6, lipopolysaccharide (LPS, 10 ng/mL; Sigma) or medium only. LPS is a Gram-negative bacterial cell wall component that can stimulate the production of a number of inflammatory cytokines in various cells. Following 24 h of incubation, the cells were processed for subsequent applications.
2.3.2. Primary Pure Glial Cell Culture
Microglia
Following isolation and culture of mixed glial populations over 10 days as described above, separation of distinct glial cell types was performed as described by Mecha et al. [
23]. Briefly, flasks containing settled mixed glial cultures were shaken orbitally at 230 rpm for 3 h at 37 °C. The suspension of cells following shaking, containing detached microglia, was collected and immediately added to warm DMEM 10:10:1 in tissue culture flasks. The cell suspension was centrifuged for 10 min at 168×
g, the cell pellet was resuspended in warm DMEM 10:10:1, and cells were added to a 24-well plate and incubated at 37 °C in a humidified atmosphere of 5% CO
2 in air. Immunofluorescence analysis of cultures for distinct cell type markers, Iba1 (microglia), GFAP (astrocytes), Sox10 (oligodendrocyte precursor cells, OPCs), verified their identity and purity at ≥95%. The cells then underwent various treatments for experiments, as described in the Section Results.
Astrocytes
According to the method described by Mecha et al. [
23], following the initial detachment of microglia from the mixed glial cultures, the remaining cells were placed on an incubating orbital shaker and shaken at 260 rpm overnight at 37 °C. The following day, the cell suspension, mostly containing oligodendrocyte-lineage cells, was removed, and the remaining attached astrocytes were detached from the culture plastic surface by tryptic digestion (TrypLE
™ Express; Gibco Invitrogen). The cell pellet following centrifugation was resuspended in warm DMEM 5:5:1 containing 5% FCS, 5% horse serum and 1% penicillin/streptomycin. Cells were added to a 24-well plate and incubated at 37 °C in a humidified atmosphere of 5% CO
2 in air. As stated above, immunostaining analysis verified the cultures’ identity and purity at ≥95%. The cells then underwent various treatments for experiments, as described in the Section Results.
2.4. Reverse-Transcription Quantitative PCR
Total RNA was isolated from cultured optic nerves as well as cultured mixed glial cells using a total RNA extraction kit (RNeasy Mini Kit; Qiagen, Manchester, UK). RNA was reverse-transcribed (Applied BioSystems, Loughborough, UK) to produce cDNA for subsequent real-time quantitative polymerase chain reaction (qPCR) analysis. qPCR analysis was performed using specific primers and fluorescent hydrolysis probes for each gene (Integrated DNA Technologies; Leuven, Belgium) within a reaction master mix (FastStart Essential DNA Probes Master; Roche, Burgess Hill, UK). The expression levels of all genes in the samples were normalised against the expression of the mouse
Gapdh gene as reference gene. All samples were analysed based on the ΔC
T and ΔΔC
T method, where ΔC
T is C
T[Target gene]-C
T[Housekeeping gene], and 2
−ΔCT represents the relative gene expression. ΔΔC
T is ΔC
T[Gas6]-ΔC
T[Mock], and 2
−ΔΔCT shows fold up- or downregulation of the gene of interest, whereby values >1 and <1 are deemed as corresponding to up- and downregulation, respectively [
24].
2.5. SDS-PAGE and Western Blot
Total protein was extracted from cultured optic nerves for SDS-PAGE analysis of proteins. Tissues were homogenised in lysis buffer composed of 50 mM Tris-HCl, 150 mM NaCl, 1% Triton X-100, 0.5% NP-40, 1 mM EDTA, 10 mM Na
4P
2O
7, pH 8.0. Equal amounts of total protein were loaded onto a 10% polyacrylamide gel, and proteins were separated by SDS-PAGE as previously described [
25]. The separated proteins in the gel were transferred to a polyvinylidene fluoride membrane (Immobilon-P; Millipore, Watford, UK). Membranes were first blocked in 3% non-fat dry milk in 25 mM Tris, 150 mM NaCl, 0.05% Tween-20, pH 8.0, for 1 h at room temperature (RT), after which they were incubated with primary antibodies at 4 °C overnight. Primary antibodies were against β-actin (dilution, 1:10000, A2066; Sigma) and myelin basic protein (MBP, dilution, 1:500, MAB386; Millipore, Darmstadt, Germany). Washing of the membrane (25 mM Tris-HCl, 150 mM NaCl, 0.05% Tween-20, pH 8.0) was followed by 1 h incubation with a horseradish peroxidase-conjugated secondary antibody recognising the appropriate primary antibody (1:5000; Promega, Southampton, UK; Dako, Glostrup, Denmark) at RT. A chemiluminescence detection reagent (Luminata Forte Western HRP Substrate; Millipore) was used to develop the signal, and bands were visualized by a CCD-based gel imager (Bio-Rad ChemiDoc
™ MP, Hemel Hempstead, UK). Band intensities were quantified by densitometry using ImageJ software, with band intensities of the proteins of interest being normalised against those of actin protein bands in every sample lane.
2.6. IL-10 ELISA
The cell culture supernatant was collected from mixed glial cultures following experimental treatments as described in the Section Results, and samples were stored at −80 °C. ELISA for mouse IL-10 was performed in duplicate wells per each sample, following the manufacturer’s instructions (mouse IL-10 ELISA kit, Bio-Techne, Abingdon, UK).
2.7. Statistical Analysis
All statistical analyses were performed using the software Prism 6 (GraphPad Inc, La Jolla, CA, USA). Animal tissue cultures were randomly assigned to the treatment groups. All results are expressed as mean ± SEM, with each experiment performed a minimum 3 times, as specified in the figure legend, using multiple replicates per treatment. Differences between multiple treatments in mixed glial cultures were compared using paired parametric student t test and, in optic nerve cultures, were compared using unpaired parametric t test. A p value of less than 0.05 was considered statistically significant.
4. Discussion
Previously, we and others have shown that all three TAM receptors are expressed in CNS tissues with distinct expression profiles and that there is a postnatal increase in Tyro3 and Axl expression concomitant with the timeline of myelination [
15,
26]. However, the involvement of the individual TAM receptors in the biology of specific CNS glial cells remains to be comprehensively characterised. Here, we have determined the expression of Gas6 and the TAM receptors in mouse primary pure glial cell cultures incorporating astrocytes, microglia and OPCs. Also in this study, we have probed further the molecular mechanisms behind the action of Gas6 in the mouse optic nerve, which we previously showed to respond to Gas6 with an increase in OPC numbers, MBP expression and myelination [
15]. We observed here that the pro-myelinating effect of Gas6 in the optic nerve occurs through the induction of the anti-inflammatory cytokine IL-10. This effect was reproduced using mixed glial cell cultures, and experiments with TAM single-knockout cells revealed the necessity of both Tyro3 and Axl receptors mediating in concert the Gas6 effect. These results therefore suggest that the recognised role of Gas6–TAM signalling in regulating inflammation is linked to its role in CNS myelination.
The expression of TAM receptors in various regions of the CNS has previously been reported, with Tyro3 being the most prominent TAM overall [
26]. Here, we observed that microglia strongly expressed Mer and Axl receptors at mRNA level, whereas astrocytes expressed Axl and appeared to be the only glial cells showing Tyro3 expression. In contrast, OPCs had negligible expression of all these genes. We did also attempt Western blot and flow cytometry as approaches for protein detection in the glial cultures; however, these primary cells were not amenable to efficient protein extraction or did not supply sufficient yields for flow experiments (data not shown). The mRNA expression analysis also revealed microglia to be the main glial cell source of Gas6, showing strong expression. These findings are largely in line with other observations, though with some differences. For example, our observation of Axl expression in microglia contrasts with that of another study which showed Axl expression to be weak [
27]. This, however, may be due to differences in the level of expression captured at the time of analysis, tissue source, as well as age. We have shown previously that TAM expression increases during postnatal development through to adulthood [
15]. A separate study also showed that Tyro3 expression begins to increase shortly after birth, significantly augmenting until reaching its peak at P24 [
26]. Therefore, this can explain the overall negligible expression of all TAMs and Gas6 in our neonatal OPC cultures. Nevertheless, the combined findings of these studies show that TAMs appear to be important in the CNS after birth and, moreover, appear to play distinct roles in different glial cells.
IL-10 is a well-known immunoregulatory cytokine that acts on both haematopoietic and non-haematopoietic cells to control inflammatory responses and immune reactions, also within the brain [
28,
29]. Amongst its effects is the suppression of secretion of proinflammatory cytokines by monocytes, macrophages and, likely, microglia [
28,
30,
31]. We report here that Gas6 significantly upregulated IL-10 gene expression in cultured optic nerve, which was replicated in cultured mixed glial cells. However, the effect of Gas6 on IL-10 at the protein level as measured by ELISA was not significant; this is most likely due to the low overall protein levels detected in the culture medium and to factors including mRNA translation rate, protein secretion per cell, sufficient cell numbers in the sample, degree of dilution in the medium. Moreover, the intercellular communication model we propose involves IL-10 signalling between closely associating cells in the tissue, and hence, small changes in IL-10 protein concentration in the culture medium may not properly reflect the more impactful in situ role of IL-10. This new finding supports the anti-inflammatory, pro-myelinating and pro-repair role that Gas6–TAM signalling is thought to mediate in the CNS and is in keeping with our previous report of Gas6 upregulation of IL-13 and concomitant downregulation of MMP-9, EphA1 and GFAP genes in the optic nerve [
15]. It is also in keeping with the finding that Gas6 increased IL-10 expression in macrophages and was itself stimulated by IL-10 [
32].
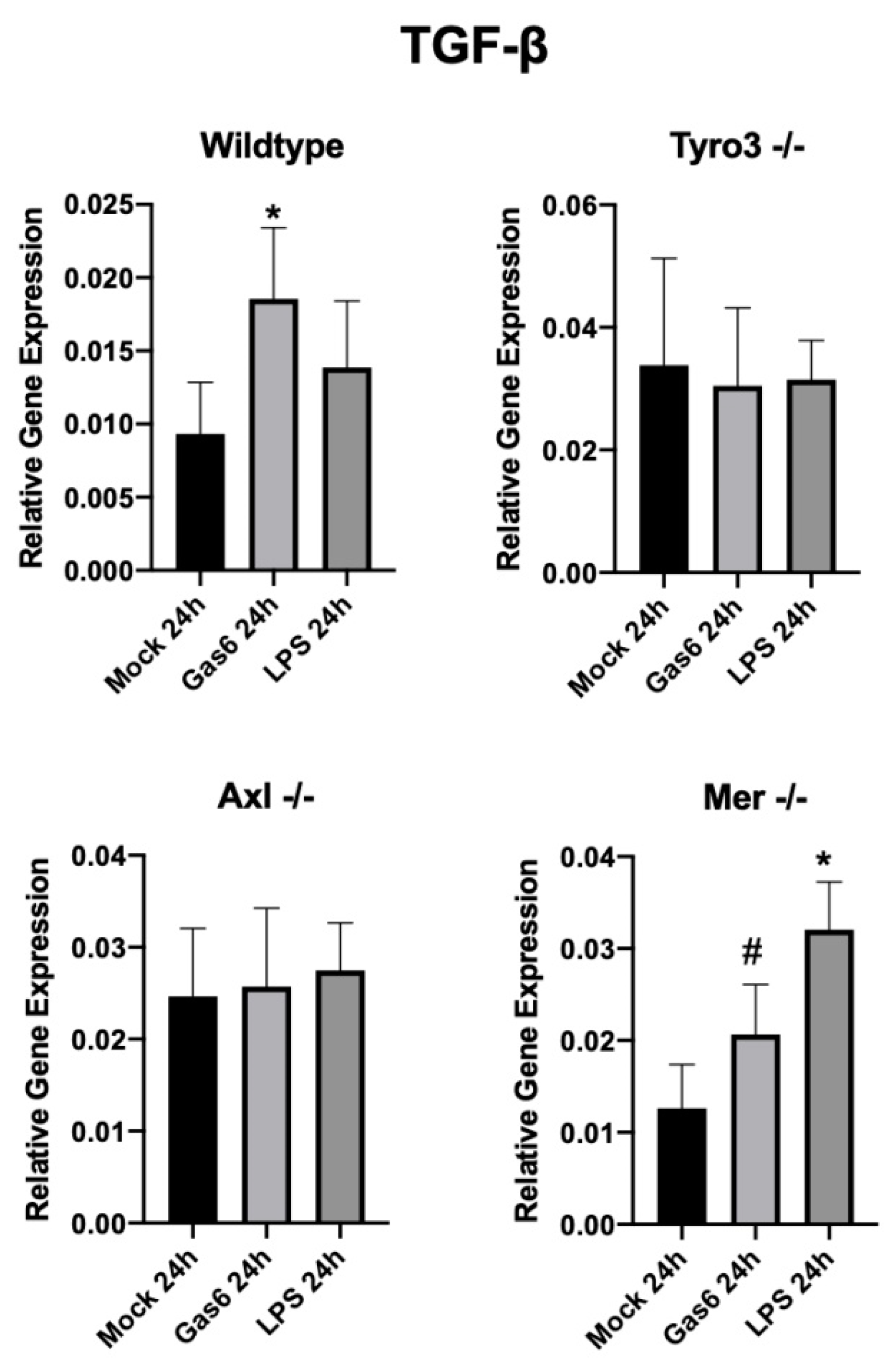
In addition, we observed that Gas6 stimulated an increase in TGF-β expression in both optic nerve and mixed glial cell cultures. Interestingly, a link between IL-10 and TGF-β signalling in regulating CNS inflammation has been reported, whereby IL-10 released by microglia during inflammation acts on astrocytes to stimulate the latter’s release of TGF-β, which then can act on microglia to suppress inflammation [
29]. Furthermore, we used tissue/cells from TAM single-receptor knockout mice in order to identify the specific TAM receptor(s) mediating Gas6 response. Gas6-mediated upregulation of IL-10 and TGF-β expression was abolished in both Axl- and Tyro3-deficient models, whereas it still occurred in the absence of Mer. Therefore, it appears that the joint presence of both Axl and Tyro3 receptors is necessary for Gas6 to be able to exert its anti-inflammatory and pro-repair functions through IL-10 and TGF-β induction. Although the molecular mechanism for this arrangement is not yet known, we speculate that it may involve heterodimerisation amongst these two TAMs and/or a parallel signalling via the separate TAMs that may be most efficacious when combined [
33,
34,
35]. In support of this, in Rat2 cells, Tyro3 and Axl were shown to be closely associated with the ability to cross-phosphorylate and enhance one another’s activation [
36].
Considering the fact that both optic nerve and mixed glial culture models contain glial cells, it is most likely that microglia are the main source of IL-10, including in response to Gas6. We also added Gas6 directly to pure microglial cultures; however, no significant increase in IL-10 expression was observed (data not shown), which may suggest an indirect cellular pathway of Gas6 ligand stimulation. In experiments where conditioned medium from astrocytes treated with Gas6 was added to microglia, a slight but insignificant increase in IL-10 expression was observed (data not shown). These supplementary observations therefore indicate that a full effect of Gas6 on IL-10 induction requires a direct physical interaction between microglia and astrocytes, as afforded in the optic nerve and mixed glial cell culture models. Moreover, a crucial role for astrocytes in this mechanism is also indicated by the fact that the presence of Tyro3 is necessary for the Gas6 effect, this TAM receptor being appreciably expressed only in astrocytes.
We previously reported that Gas6 significantly upregulated MBP expression in the optic nerve and protected against experimental demyelination in brain slice cultures [
15]. Moreover, Gas6 has been shown to be capable of boosting remyelination in a cuprizone-induced demyelination model [
20]. In addition to its primary role as an anti-inflammatory cytokine, IL-10 may also be involved in CNS regeneration and myelination, as it has been shown to boost myelination and to reduce scar formation in a rodent nerve repair model [
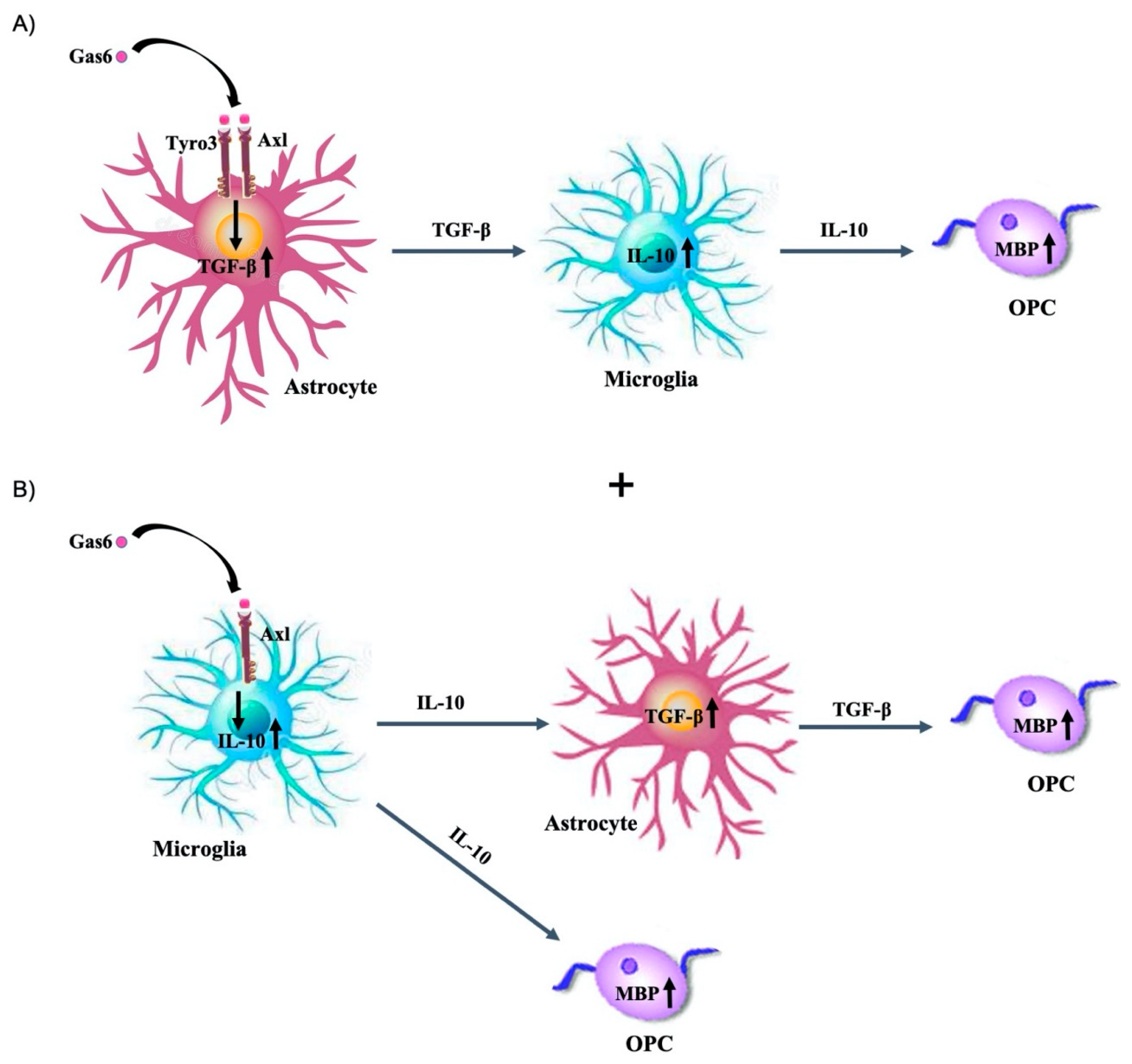
37]. Our data indicate an indirect effect of Gas6 on MBP via IL-10 induction, although the intercellular signalling mechanism behind this effect requires full characterisation. One model is that Gas6 stimulates first Tyro3 and Axl receptors on astrocytes, causing the release of TGF-β from these cells, which subsequently acts on microglia to cause IL-10 upregulation (
Figure 6A). Such a link is consistent with previous observations indicating that IL-10 upregulation in macrophages occurred in parallel with TGF-β increase and was blocked by a neutralising anti-TGF-β antibody. Also, concomitant to the astrocyte response, Gas6 could induce IL-10 upregulation in microglia, presumably via Axl, prior to TGF-β upregulation, and as a result, upregulated IL-10 could induce myelination, whilst TGF-β released from astrocytes would push OPCs towards a pathway that favours myelination (
Figure 6B).
In conclusion, this study has shown the discrete expression of TAM receptors and their ligand Gas6 in various glial cell types and that Gas6 upregulates IL-10 and TGF-β expression in both optic nerve and mixed glial cell cultures. The activation of both Axl and Tyro3 in astrocytes appears to be necessary for the effect of Gas6, and the positive effect of Gas6 on myelination is mediated through IL-10 induction and release. Therefore, these findings support a prominent role for Gas6 as an anti-inflammatory molecule in conjunction with its pro-myelinating and pro-repair functions in the CNS.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





