Neutrophil Extracellular Traps in Periodontitis

 ,
,  and
and 
Abstract
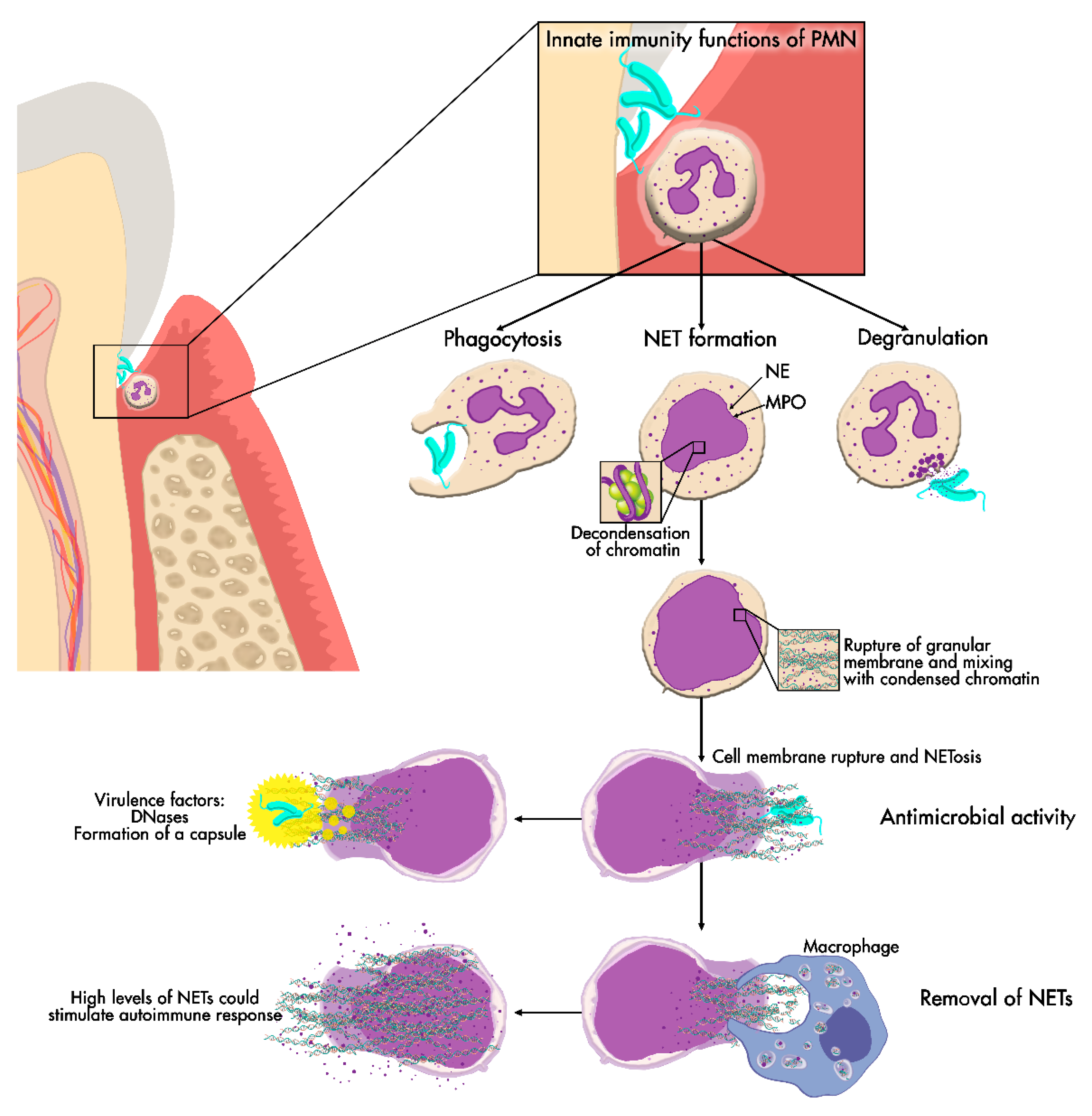
1. Periodontitis
2. Periodontal Neutrophils
3. Neutrophil Extracellular Traps (NETs)
3.1. NET Formation
3.2. Microbicidal Effects of NETs
3.3. Microorganisms’ Strategies to Escape the Action of NETs
3.4. Removal of NETs
4. NETs and Periodontitis
4.1. NETs in Periodontitis Studies
4.2. Microbial Agents Alter NET Formation
4.3. Defective Neutrophils and Impaired NET Formation in Periodontitis
5. Role of NETs in Systemic Diseases
The Relationship between Rheumatoid Arthritis, Periodontitis and NETs
6. Conclusions and Future Research Lines
Funding
Conflicts of Interest
References
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Darveau, R.P. Periodontitis: A polymicrobial disruption of host homeostasis. Nat. Rev. Microbiol. 2010, 8, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Nibali, L.; Henderson, B.; Sadiq, S.T.; Donos, N. Genetic dysbiosis: The role of microbial insults in chronic inflammatory diseases. J. Oral. Microbiol. 2014, 6, 22962. [Google Scholar] [CrossRef]
- Nauseef, W.M. Proteases, neutrophils, and periodontitis: The net effect. J. Clin. Investig. 2014, 124, 4237–4239. [Google Scholar] [CrossRef] [PubMed]
- Bascones-Martinez, A.; Munoz-Corcuera, M.; Noronha, S.; Mota, P.; Bascones-Ilundain, C.; Campo-Trapero, J. Host defence mechanisms against bacterial aggression in periodontal disease: Basic mechanisms. Med. Oral Patol. Oral Cir. Bucal. 2009, 14, e680–e685. [Google Scholar] [CrossRef][Green Version]
- Kassebaum, N.J.; Bernabe, E.; Dahiya, M.; Bhandari, B.; Murray, C.J.; Marcenes, W. Global burden of severe periodontitis in 1990–2010: A systematic review and meta-regression. J. Dent. Res. 2014, 93, 1045–1053. [Google Scholar] [CrossRef]
- Frencken, J.E.; Sharma, P.; Stenhouse, L.; Green, D.; Laverty, D.; Dietrich, T. Global epidemiology of dental caries and severe periodontitis—A comprehensive review. J. Clin. Periodontol. 2017, 44, S94–S105. [Google Scholar] [CrossRef]
- Marcenes, W.; Kassebaum, N.J.; Bernabe, E.; Flaxman, A.; Naghavi, M.; Lopez, A.; Murray, C.J. Global burden of oral conditions in 1990–2010: A systematic analysis. J. Dent. Res. 2013, 92, 592–597. [Google Scholar] [CrossRef]
- Eke, P.I.; Dye, B.A.; Wei, L.; Slade, G.D.; Thornton-Evans, G.O.; Borgnakke, W.S.; Taylor, G.W.; Page, R.C.; Beck, J.D.; Genco, R.J. Update on prevalence of periodontitis in adults in the united states: Nhanes 2009 to 2012. J. Periodontol. 2015, 86, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Mesa, F.; Liebana, J.; Galindo-Moreno, P.; J. O’Valle, F. Oral pathogens, immunity, and periodontal diseases. Curr. Immunol. Rev. 2011, 7, 83–91. [Google Scholar] [CrossRef]
- Jaillon, S.; Galdiero, M.R.; Del Prete, D.; Cassatella, M.A.; Garlanda, C.; Mantovani, A. Neutrophils in innate and adaptive immunity. Semin. Immunopathol. 2013, 35, 377–394. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.A.; Krauss, J. Neutrophils in periodontal inflammation. Front. Oral Biol. 2012, 15, 56–83. [Google Scholar] [PubMed]
- Cortes-Vieyra, R.; Rosales, C.; Uribe-Querol, E. Neutrophil functions in periodontal homeostasis. J. Immunol. Res. 2016, 2016, 1396106. [Google Scholar] [CrossRef]
- De Smit, M.; Westra, J.; Vissink, A.; Doornbos-van der Meer, B.; Brouwer, E.; van Winkelhoff, A.J. Periodontitis in established rheumatoid arthritis patients: A cross-sectional clinical, microbiological and serological study. Arthritis Res. Ther. 2012, 14, R222. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.B.; Wright, H.J.; Roberts, A.; Ling-Mountford, N.; Cooper, P.R.; Chapple, I.L. Neutrophil hyper-responsiveness in periodontitis. J. Dent. Res. 2007, 86, 718–722. [Google Scholar] [CrossRef]
- Eash, K.J.; Greenbaum, A.M.; Gopalan, P.K.; Link, D.C. Cxcr2 and cxcr4 antagonistically regulate neutrophil trafficking from murine bone marrow. J. Clin. Investig. 2010, 120, 2423–2431. [Google Scholar] [CrossRef] [PubMed]
- Thorbert-Mros, S.; Larsson, L.; Berglundh, T. Cellular composition of long-standing gingivitis and periodontitis lesions. J. Periodontal. Res. 2015, 50, 535–543. [Google Scholar] [CrossRef]
- Moonen, C.G.J.; Hirschfeld, J.; Cheng, L.; Chapple, I.L.C.; Loos, B.G.; Nicu, E.A. Oral neutrophils characterized: Chemotactic, phagocytic, and neutrophil extracellular trap (net) formation properties. Front. Immunol. 2019, 10, 635. [Google Scholar] [CrossRef]
- Hirschfeld, J. Neutrophil subsets in periodontal health and disease: A mini review. Front. Immunol. 2019, 10, 3001. [Google Scholar] [CrossRef]
- Zeidler, C.; Germeshausen, M.; Klein, C.; Welte, K. Clinical implications of ela2-, hax1-, and g-csf-receptor (csf3r) mutations in severe congenital neutropenia. Br. J. Haematol. 2009, 144, 459–467. [Google Scholar] [CrossRef]
- Mc Guire, P.J.; Cunningham-Rundles, C.; Ochs, H.; Diaz, G.A. Oligoclonality, impaired class switch and b-cell memory responses in whim syndrome. Clin. Immunol. 2010, 135, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, E.; Hajishengallis, G. Neutrophil homeostasis and periodontal health in children and adults. J. Dent. Res. 2014, 93, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.M.; Brenchley, L.; Moutsopoulos, N.M. Primary immunodeficiencies reveal the essential role of tissue neutrophils in periodontitis. Immunol. Rev. 2019, 287, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G. New developments in neutrophil biology and periodontitis. Periodontology 2000 2020, 82, 78–92. [Google Scholar] [CrossRef]
- Almarza Novoa, E.; Kasbekar, S.; Thrasher, A.J.; Kohn, D.B.; Sevilla, J.; Nguyen, T.; Schwartz, J.D.; Bueren, J.A. Leukocyte adhesion deficiency-i: A comprehensive review of all published cases. J. Allergy Clin. Immunol. Pract. 2018, 6, 1418–1420. [Google Scholar] [CrossRef] [PubMed]
- Hanna, S.; Etzioni, A. Leukocyte adhesion deficiencies. Ann. N. Y. Acad. Sci. 2012, 1250, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.Y.; Chavakis, E.; Czabanka, M.A.; Langer, H.F.; Fraemohs, L.; Economopoulou, M.; Kundu, R.K.; Orlandi, A.; Zheng, Y.Y.; Prieto, D.A.; et al. Del-1, an endogenous leukocyte-endothelial adhesion inhibitor, limits inflammatory cell recruitment. Science 2008, 322, 1101–1104. [Google Scholar] [CrossRef]
- Kourtzelis, I.; Li, X.; Mitroulis, I.; Grosser, D.; Kajikawa, T.; Wang, B.; Grzybek, M.; von Renesse, J.; Czogalla, A.; Troullinaki, M.; et al. Del-1 promotes macrophage efferocytosis and clearance of inflammation. Nat. Immunol. 2019, 20, 40–49. [Google Scholar] [CrossRef]
- Roberts, H.M.; Ling, M.R.; Insall, R.; Kalna, G.; Spengler, J.; Grant, M.M.; Chapple, I.L. Impaired neutrophil directional chemotactic accuracy in chronic periodontitis patients. J. Clin. Periodontol. 2015, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Moutsopoulos, N.M.; Konkel, J.; Sarmadi, M.; Eskan, M.A.; Wild, T.; Dutzan, N.; Abusleme, L.; Zenobia, C.; Hosur, K.B.; Abe, T.; et al. Defective neutrophil recruitment in leukocyte adhesion deficiency type i disease causes local il-17-driven inflammatory bone loss. Sci. Transl. Med. 2014, 6, 229–240. [Google Scholar] [CrossRef]
- Moutsopoulos, N.M.; Zerbe, C.S.; Wild, T.; Dutzan, N.; Brenchley, L.; DiPasquale, G.; Uzel, G.; Axelrod, K.C.; Lisco, A.; Notarangelo, L.D.; et al. Interleukin-12 and interleukin-23 blockade in leukocyte adhesion deficiency type 1. N. Engl. J. Med. 2017, 376, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, D.; Kumar, S.; Desai, J.; Anders, H.J. Neutrophil extracellular traps in tissue pathology. Histol. Histopathol. 2017, 32, 203–213. [Google Scholar] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Rizo, V.; Martinez-Guzman, M.A.; Iniguez-Gutierrez, L.; Garcia-Orozco, A.; Alvarado-Navarro, A.; Fafutis-Morris, M. Neutrophil extracellular traps and its implications in inflammation: An overview. Front. Immunol. 2017, 8, 81. [Google Scholar] [CrossRef] [PubMed]
- Desai, J.; Kumar, S.V.; Mulay, S.R.; Konrad, L.; Romoli, S.; Schauer, C.; Herrmann, M.; Bilyy, R.; Muller, S.; Popper, B.; et al. Pma and crystal-induced neutrophil extracellular trap formation involves ripk1-ripk3-mlkl signaling. Eur. J. Immunol. 2016, 46, 223–229. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Boeltz, S.; Amini, P.; Anders, H.J.; Andrade, F.; Bilyy, R.; Chatfield, S.; Cichon, I.; Clancy, D.M.; Desai, J.; Dumych, T.; et al. To net or not to net:Current opinions and state of the science regarding the formation of neutrophil extracellular traps. Cell Death Differ. 2019, 26, 395–408. [Google Scholar] [CrossRef]
- Sierra, J.M.; Fuste, E.; Rabanal, F.; Vinuesa, T.; Vinas, M. An overview of antimicrobial peptides and the latest advances in their development. Expert Opin. Biol. Ther. 2017, 17, 663–676. [Google Scholar] [CrossRef]
- Kang, H.K.; Kim, C.; Seo, C.H.; Park, Y. The therapeutic applications of antimicrobial peptides (amps): A patent review. J. Microbiol. 2017, 55, 1–12. [Google Scholar] [CrossRef]
- Brinkmann, V. Neutrophil extracellular traps in the second decade. J. Innate Immun. 2018, 10, 414–421. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef]
- Gupta, A.K.; Giaglis, S.; Hasler, P.; Hahn, S. Efficient neutrophil extracellular trap induction requires mobilization of both intracellular and extracellular calcium pools and is modulated by cyclosporine a. PLoS ONE 2014, 9, e97088. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.R.; Palmer, L.J.; Chapple, I.L. Neutrophil extracellular traps as a new paradigm in innate immunity: Friend or foe? Periodontology 2000 2013, 63, 165–197. [Google Scholar] [CrossRef]
- Pilsczek, F.H.; Salina, D.; Poon, K.K.; Fahey, C.; Yipp, B.G.; Sibley, C.D.; Robbins, S.M.; Green, F.H.; Surette, M.G.; Sugai, M.; et al. A novel mechanism of rapid nuclear neutrophil extracellular trap formation in response to staphylococcus aureus. J. Immunol. 2010, 185, 7413–7425. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, M.; Lindberg, M.R.; Kennett, M.J.; Xiong, N.; Wang, Y. Pad4 is essential for antibacterial innate immunity mediated by neutrophil extracellular traps. J. Exp. Med. 2010, 207, 1853–1862. [Google Scholar] [CrossRef]
- Yousefi, S.; Mihalache, C.; Kozlowski, E.; Schmid, I.; Simon, H.U. Viable neutrophils release mitochondrial DNA to form neutrophil extracellular traps. Cell Death Differ. 2009, 16, 1438–1444. [Google Scholar] [CrossRef]
- Brinkmann, V.; Zychlinsky, A. Beneficial suicide: Why neutrophils die to make nets. Nat. Rev. Microbiol. 2007, 5, 577–582. [Google Scholar] [CrossRef]
- Jorch, S.K.; Kubes, P. An emerging role for neutrophil extracellular traps in noninfectious disease. Nat. Med. 2017, 23, 279–287. [Google Scholar] [CrossRef]
- Arazna, M.; Pruchniak, M.P.; Demkow, U. Neutrophil extracellular traps in bacterial infections: Strategies for escaping from killing. Respir. Physiol. Neurobiol. 2013, 187, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Parker, H.; Albrett, A.M.; Kettle, A.J.; Winterbourn, C.C. Myeloperoxidase associated with neutrophil extracellular traps is active and mediates bacterial killing in the presence of hydrogen peroxide. J. Leukoc. Biol. 2012, 91, 369–376. [Google Scholar] [CrossRef]
- Berends, E.T.; Horswill, A.R.; Haste, N.M.; Monestier, M.; Nizet, V.; von Kockritz-Blickwede, M. Nuclease expression by staphylococcus aureus facilitates escape from neutrophil extracellular traps. J. Innate Immun. 2010, 2, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Beiter, K.; Wartha, F.; Albiger, B.; Normark, S.; Zychlinsky, A.; Henriques-Normark, B. An endonuclease allows streptococcus pneumoniae to escape from neutrophil extracellular traps. Curr. Biol. 2006, 16, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Palmer, L.J.; Chapple, I.L.; Wright, H.J.; Roberts, A.; Cooper, P.R. Extracellular deoxyribonuclease production by periodontal bacteria. J. Periodontal. Res. 2012, 47, 439–445. [Google Scholar] [CrossRef]
- Bryzek, D.; Ciaston, I.; Dobosz, E.; Gasiorek, A.; Makarska, A.; Sarna, M.; Eick, S.; Puklo, M.; Lech, M.; Potempa, B.; et al. Triggering netosis via protease-activated receptor (par)-2 signaling as a mechanism of hijacking neutrophils function for pathogen benefits. PLoS Pathog. 2019, 15, e1007773. [Google Scholar] [CrossRef]
- Jayaprakash, K.; Demirel, I.; Khalaf, H.; Bengtsson, T. The role of phagocytosis, oxidative burst and neutrophil extracellular traps in the interaction between neutrophils and the periodontal pathogen porphyromonas gingivalis. Mol. Oral Microbiol. 2015, 30, 361–375. [Google Scholar] [CrossRef]
- Doke, M.; Fukamachi, H.; Morisaki, H.; Arimoto, T.; Kataoka, H.; Kuwata, H. Nucleases from prevotella intermedia can degrade neutrophil extracellular traps. Mol. Oral Microbiol. 2017, 32, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Hakkim, A.; Furnrohr, B.G.; Amann, K.; Laube, B.; Abed, U.A.; Brinkmann, V.; Herrmann, M.; Voll, R.E.; Zychlinsky, A. Impairment of neutrophil extracellular trap degradation is associated with lupus nephritis. Proc. Natl. Acad. Sci. USA 2010, 107, 9813–9818. [Google Scholar] [CrossRef] [PubMed]
- Farrera, C.; Fadeel, B. Macrophage clearance of neutrophil extracellular traps is a silent process. J. Immunol. 2013, 191, 2647–2656. [Google Scholar] [CrossRef] [PubMed]
- White, P.; Sakellari, D.; Roberts, H.; Risafi, I.; Ling, M.; Cooper, P.; Milward, M.; Chapple, I. Peripheral blood neutrophil extracellular trap production and degradation in chronic periodontitis. J. Clin. Periodontol. 2016, 43, 1041–1049. [Google Scholar] [CrossRef]
- Moonen, C.G.; Buurma, K.G.; Faruque, M.R.; Balta, M.G.; Liefferink, E.; Bizzarro, S.; Nicu, E.A.; Loos, B.G. Periodontal therapy increases neutrophil extracellular trap degradation. Innate Immun. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yang, X.M.; Jia, S.Y. Characteristics of neutrophil extracellular traps in patients with periodontitis and gingivitis. Braz. Oral Res. 2020, 34, e015. [Google Scholar] [CrossRef] [PubMed]
- Magan-Fernandez, A.; O’Valle, F.; Abadia-Molina, F.; Munoz, R.; Puga-Guil, P.; Mesa, F. Characterization and comparison of neutrophil extracellular traps in gingival samples of periodontitis and gingivitis: A pilot study. J. Periodontal Res. 2019, 54, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.H.; Chapple, I.L.C.; Shapira, L.; Golan-Goldhirsh, A.; Gopas, J.; Polak, D. Nupharidine enhances aggregatibacter actinomycetemcomitans clearance by priming neutrophils and augmenting their effector functions. J. Clin. Periodontol. 2019, 46, 62–71. [Google Scholar] [CrossRef]
- Kaneko, C.; Kobayashi, T.; Ito, S.; Sugita, N.; Murasawa, A.; Nakazono, K.; Yoshie, H. Circulating levels of carbamylated protein and neutrophil extracellular traps are associated with periodontitis severity in patients with rheumatoid arthritis: A pilot case-control study. PLoS ONE 2018, 13, e0192365. [Google Scholar] [CrossRef]
- Fine, N.; Hassanpour, S.; Borenstein, A.; Sima, C.; Oveisi, M.; Scholey, J.; Cherney, D.; Glogauer, M. Distinct oral neutrophil subsets define health and periodontal disease states. J. Dent. Res. 2016, 95, 931–938. [Google Scholar] [CrossRef]
- Hirschfeld, J.; Dommisch, H.; Skora, P.; Horvath, G.; Latz, E.; Hoerauf, A.; Waller, T.; Kawai, T.; Jepsen, S.; Deschner, J.; et al. Neutrophil extracellular trap formation in supragingival biofilms. Int. J. Med. Microbiol. 2015, 305, 453–463. [Google Scholar] [CrossRef]
- Vitkov, L.; Klappacher, M.; Hannig, M.; Krautgartner, W.D. Neutrophil fate in gingival crevicular fluid. Ultrastruct. Pathol. 2010, 34, 25–30. [Google Scholar] [CrossRef]
- Vitkov, L.; Klappacher, M.; Hannig, M.; Krautgartner, W.D. Extracellular neutrophil traps in periodontitis. J. Periodontal. Res. 2009, 44, 664–672. [Google Scholar] [CrossRef]
- Alyami, H.M.; Finoti, L.S.; Teixeira, H.S.; Aljefri, A.; Kinane, D.F.; Benakanakere, M.R. Role of nod1/nod2 receptors in fusobacterium nucleatum mediated netosis. Microb. Pathog. 2019, 131, 53–64. [Google Scholar] [CrossRef]
- Hirschfeld, J.; White, P.C.; Milward, M.R.; Cooper, P.R.; Chapple, I.L.C. Modulation of neutrophil extracellular trap and reactive oxygen species release by periodontal bacteria. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [PubMed]
- Roberts, H.; White, P.; Dias, I.; McKaig, S.; Veeramachaneni, R.; Thakker, N.; Grant, M.; Chapple, I. Characterization of neutrophil function in papillon-lefevre syndrome. J. Leukoc. Biol. 2016, 100, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Palmer, L.J.; Damgaard, C.; Holmstrup, P.; Nielsen, C.H. Influence of complement on neutrophil extracellular trap release induced by bacteria. J. Periodontal Res. 2016, 51, 70–76. [Google Scholar] [CrossRef]
- Hirschfeld, J.; Roberts, H.M.; Chapple, I.L.; Parcina, M.; Jepsen, S.; Johansson, A.; Claesson, R. Effects of aggregatibacter actinomycetemcomitans leukotoxin on neutrophil migration and extracellular trap formation. J. Oral Microbiol. 2016, 8, 33070. [Google Scholar] [CrossRef]
- Wright, H.J.; Matthews, J.B.; Chapple, I.L.; Ling-Mountford, N.; Cooper, P.R. Periodontitis associates with a type 1 ifn signature in peripheral blood neutrophils. J. Immunol. 2008, 181, 5775–5784. [Google Scholar] [CrossRef]
- Clark, S.R.; Ma, A.C.; Tavener, S.A.; McDonald, B.; Goodarzi, Z.; Kelly, M.M.; Patel, K.D.; Chakrabarti, S.; McAvoy, E.; Sinclair, G.D.; et al. Platelet tlr4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat. Med. 2007, 13, 463–469. [Google Scholar] [CrossRef]
- White, P.; Cooper, P.; Milward, M.; Chapple, I. Differential activation of neutrophil extracellular traps by specific periodontal bacteria. Free Radic. Biol. Med. 2014, 75, S53. [Google Scholar] [CrossRef] [PubMed]
- Vitkov, L.; Hannig, M.; Minnich, B.; Herrmann, M. Periodontal sources of citrullinated antigens and tlr agonists related to ra. Autoimmunity 2018, 51, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.B.; Wright, H.J.; Roberts, A.; Cooper, P.R.; Chapple, I.L. Hyperactivity and reactivity of peripheral blood neutrophils in chronic periodontitis. Clin. Exp. Immunol. 2007, 147, 255–264. [Google Scholar] [CrossRef]
- Nicu, E.A.; Rijkschroeff, P.; Wartewig, E.; Nazmi, K.; Loos, B.G. Characterization of oral polymorphonuclear neutrophils in periodontitis patients: A case-control study. BMC Oral Health 2018, 18, 149. [Google Scholar] [CrossRef]
- Borenstein, A.; Fine, N.; Hassanpour, S.; Sun, C.; Oveisi, M.; Tenenbaum, H.C.; Glogauer, M. Morphological characterization of para- and proinflammatory neutrophil phenotypes using transmission electron microscopy. J. Periodontal Res. 2018, 53, 972–982. [Google Scholar] [CrossRef]
- Cekici, A.; Kantarci, A.; Hasturk, H.; Van Dyke, T.E. Inflammatory and immune pathways in the pathogenesis of periodontal disease. Periodontology 2000 2014, 64, 57–80. [Google Scholar] [CrossRef] [PubMed]
- Bullon, P.; Castejon-Vega, B.; Roman-Malo, L.; Jimenez-Guerrero, M.P.; Cotan, D.; Forbes-Hernandez, T.Y.; Varela-Lopez, A.; Perez-Pulido, A.J.; Giampieri, F.; Quiles, J.L.; et al. Autophagic dysfunction in patients with papillon-lefevre syndrome is restored by recombinant cathepsin c treatment. J. Allergy Clin. Immunol. 2018, 142, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- Jepsen, S.; Caton, J.G.; Albandar, J.M.; Bissada, N.F.; Bouchard, P.; Cortellini, P.; Demirel, K.; de Sanctis, M.; Ercoli, C.; Fan, J.; et al. Periodontal manifestations of systemic diseases and developmental and acquired conditions: Consensus report of workgroup 3 of the 2017 world workshop on the classification of periodontal and peri-implant diseases and conditions. J. Periodontol. 2018, 89, S237–S248. [Google Scholar] [CrossRef]
- Hahn, J.; Schauer, C.; Czegley, C.; Kling, L.; Petru, L.; Schmid, B.; Weidner, D.; Reinwald, C.; Biermann, M.H.C.; Blunder, S.; et al. Aggregated neutrophil extracellular traps resolve inflammation by proteolysis of cytokines and chemokines and protection from antiproteases. FASEB J. 2019, 33, 1401–1414. [Google Scholar] [CrossRef]
- Byrd, A.S.; O’Brien, X.M.; Johnson, C.M.; Lavigne, L.M.; Reichner, J.S. An extracellular matrix-based mechanism of rapid neutrophil extracellular trap formation in response to candida albicans. J. Immunol. 2013, 190, 4136–4148. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, S.; Urosevic, M.; Daryadel, A.; Oberholzer, P.A.; Baumann, C.; Fey, M.F.; Dummer, R.; Simon, H.U.; Yousefi, S. Induction of genes mediating interferon-dependent extracellular trap formation during neutrophil differentiation. J. Biol. Chem. 2004, 279, 44123–44132. [Google Scholar] [CrossRef]
- Kaplan, M.J.; Radic, M. Neutrophil extracellular traps: Double-edged swords of innate immunity. J. Immunol. 2012, 189, 2689–2695. [Google Scholar] [CrossRef]
- McDonald, B.; Davis, R.P.; Kim, S.J.; Tse, M.; Esmon, C.T.; Kolaczkowska, E.; Jenne, C.N. Platelets and neutrophil extracellular traps collaborate to promote intravascular coagulation during sepsis in mice. Blood 2017, 129, 1357–1367. [Google Scholar] [CrossRef]
- Cools-Lartigue, J.; Spicer, J.; McDonald, B.; Gowing, S.; Chow, S.; Giannias, B.; Bourdeau, F.; Kubes, P.; Ferri, L. Neutrophil extracellular traps sequester circulating tumor cells and promote metastasis. J. Clin. Investig. 2013, 123, 3446–3458. [Google Scholar] [CrossRef]
- Garley, M.; Dziemianczyk-Pakiela, D.; Grubczak, K.; Surazynski, A.; Dabrowska, D.; Ratajczak-Wrona, W.; Sawicka-Powierza, J.; Borys, J.; Moniuszko, M.; Palka, J.A.; et al. Differences and similarities in the phenomenon of nets formation in oral inflammation and in oral squamous cell carcinoma. J. Cancer 2018, 9, 1958–1965. [Google Scholar] [CrossRef] [PubMed]
- Khandpur, R.; Carmona-Rivera, C.; Vivekanandan-Giri, A.; Gizinski, A.; Yalavarthi, S.; Knight, J.S.; Friday, S.; Li, S.; Patel, R.M.; Subramanian, V.; et al. Nets are a source of citrullinated autoantigens and stimulate inflammatory responses in rheumatoid arthritis. Sci. Transl. Med. 2013, 5, 178ra140. [Google Scholar] [CrossRef]
- Panda, R.; Krieger, T.; Hopf, L.; Renne, T.; Haag, F.; Rober, N.; Conrad, K.; Csernok, E.; Fuchs, T.A. Neutrophil extracellular traps contain selected antigens of anti-neutrophil cytoplasmic antibodies. Front. Immunol. 2017, 8, 439. [Google Scholar] [CrossRef]
- Gupta, A.; Hasler, P.; Gebhardt, S.; Holzgreve, W.; Hahn, S. Occurrence of neutrophil extracellular DNA traps (nets) in pre-eclampsia: A link with elevated levels of cell-free DNA? Ann. NY Acad. Sci. 2006, 1075, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Drechsler, M.; Doring, Y.; Megens, R.T.; Soehnlein, O. Neutrophilic granulocytes—Promiscuous accelerators of atherosclerosis. Thromb. Haemost. 2011, 106, 839–848. [Google Scholar] [PubMed]
- Olsen, I.; Singhrao, S.K.; Potempa, J. Citrullination as a plausible link to periodontitis, rheumatoid arthritis, atherosclerosis and alzheimer’s disease. J. Oral Microbiol. 2018, 10, 1487742. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, M.; Eriksson, K.; Lee, L.; Hermansson, M.; Johansson, A.; Nicholas, A.P.; Gerasimcik, N.; Lundberg, K.; Klareskog, L.; Catrina, A.I.; et al. Increased citrullination and expression of peptidylarginine deiminases independently of p. Gingivalis and a. Actinomycetemcomitans in gingival tissue of patients with periodontitis. J. Trans. Med. 2018, 16, 214. [Google Scholar] [CrossRef]
- Corsiero, E.; Pratesi, F.; Prediletto, E.; Bombardieri, M.; Migliorini, P. Netosis as source of autoantigens in rheumatoid arthritis. Front. Immunol. 2016, 7, 485. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, Z.; Shu, D.; Xiong, Y.; He, M.; Xu, S.; Si, S.; Guo, B. Association of periodontitis with rheumatoid arthritis and the effect of non-surgical periodontal treatment on disease activity in patients with rheumatoid arthritis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 5802–5810. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, I.A.; Lee, J.Y.; Kim, K.H.; Kim, S.; Koo, K.T.; Kim, T.I.; Seol, Y.J.; Ku, Y.; Rhyu, I.C.; et al. Periodontal pathogens and the association between periodontitis and rheumatoid arthritis in korean adults. J. Periodontal Implant. Sci. 2018, 48, 347–359. [Google Scholar] [CrossRef]
- Lee, J.Y.; Choi, I.A.; Kim, J.H.; Kim, K.H.; Lee, E.Y.; Lee, E.B.; Lee, Y.M.; Song, Y.W. Association between anti-porphyromonas gingivalis or anti-alpha-enolase antibody and severity of periodontitis or rheumatoid arthritis (ra) disease activity in ra. BMC Musculoskelet. Disord. 2015, 16, 190. [Google Scholar] [CrossRef] [PubMed]
- Bizzarro, S.; Van der Velden, U.; Loos, B.G. Local disinfection with sodium hypochlorite as adjunct to basic periodontal therapy: A randomized controlled trial. J. Clin. Periodontol. 2016, 43, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Bizzarro, S.; van der Velden, U.; Teeuw, W.J.; Gerdes, V.E.A.; Loos, B.G. Effect of periodontal therapy with systemic antimicrobials on parameters of metabolic syndrome: A randomized clinical trial. J. Clin. Periodontol. 2017, 44, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Bialowas, K.; Radwan-Oczko, M.; Dus-Ilnicka, I.; Korman, L.; Swierkot, J. Periodontal disease and influence of periodontal treatment on disease activity in patients with rheumatoid arthritis and spondyloarthritis. Rheumatol. Int. 2020, 40, 455–463. [Google Scholar] [CrossRef]
- He, Y.; Yang, F.Y.; Sun, E.W. Neutrophil extracellular traps in autoimmune diseases. Chin. Med. J. Engl. 2018, 131, 1513–1519. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Studies on the Expression of NETs in Periodontitis Patients | |||||
| Author | Year | Participants | Types of samples | NET marker | Results |
| Zhang et al. [62] | 2020 | 27 periodontitis, 17 gingivitis and 20 controls | Peripheral blood neutrophils | IL-8 and TNF-alpha as NETs inducers | Periodontitis showed lower expression of IL-8 compared to controls |
| Moonen et al. A [61] | 2019 | 1st part:38 periodontitis and 38 controls 2nd part: 91 periodontitis before and after treatment | Peripheral blood neutrophils | SYTOX Green | No differences in NET degradation between healthy subjects and periodontitis. Periodontal therapy increased NET degradation |
| Magán-Fernández et al. [63] | 2019 | 6 Chronic periodontitis, 5 gingivitis and 2 controls | Gingival tissue biopsies | CitH3 and MPO | Higher H3 in gingivitis and MPO higher in periodontitis |
| Levy et al. [64] | 2019 | 3 Localized aggressive periodontitis and 3 controls and HL60 neutrophils | Peripheral blood neutrophils and HL60 neutrophils incubated with nupharidine | SYTOX Green | NET formation was higher in the neutrophils exposed to Nupharidine |
| Kaneko et al. [65] | 2018 | 40 Rheumatoid arthritis and periodontitis, 30 periodontitis and 43 controls | Serum samples | NET-associated MPO-DNA complexes by ELISA | NETs increased in the RA + periodontitis group. NETs were associated with moderate to severe periodontitis. Periodontal treatment reduced NETs |
| White et al. [60] | 2016 | Chronic periodontitis and controls (40 pairs) | Peripheral blood neutrophils stimulated with PMA or HOCl | SYTOX Green | NET formation decreased and NET removal was restored following periodontal treatment |
| Fine et al. [66] | 2016 | 17 Chronic periodontitis and 11 controls | Blood and oral neutrophils | CitH3, MPO, CD18 | Proinflammatory oral neutrophils from periodontitis showed high levels of NET formation compared to controls |
| Hirschfeld et al. [67] | 2015 | 14 Experimental gingivitis and 6 controls | Supragingival plaque, peripheral blood neutrophils | CitH3, Histone H1, CD-177, MPO, NE, Cathepsin-G. | NETs were found within the oral biofilm. Bacterial isolates tested induced NET formation. |
| Vitkov et al. [68] | 2010 | 26 Periodontitis | GCF (18); Purulent crevicular exudate (8) | Scanning electron microscopy (SEM); CitH3 and DNA | All neutrophils in the samples were citrullinated. 78% of them showed dispersed NETs |
| Vitkov et al. [69] | 2009 | 22 Chronic Periodontitis | Purulent crevicular exudate (22); Gingival biopsies (12) | Exudates: NE and DNA; Biopsies: Transmission electron microscopy (TEM) and SEM (with and without DNase). | NETs were found on all the exudate samples. DNase caused the disappearance of NETs |
| In Vitro Studies on NET Formation Induced by Periodontal Bacteria | |||||
| Author | Year | Participants | Types of samples | NET marker | Results |
| Bryzek et al. [55] | 2019 | Human donors | Peripheral blood neutrophils stimulated with different P. gingivalis strains, antigens and gingipains | NE, Hoechst 33342, ADNbc PicoGreen® and DNase I | Gingipains from P. gingivalis induce NETs formation and prevent P. gingivalis entrapment and killing |
| Alyami et al. [70] | 2019 | In vitro PMN layers | Human primary neutrophils infected with Aggregatibacter actinomycetemcomitans, P. gingivalis and F. nucleatum | SYTOX Orange, NE, CitH3, DAPI | F.nucleatum induced rapid and robust NET formation trough NOD1 and NOD 2 receptors |
| Hirschfeld et al. [71] | 2017 | 10 Healthy donors | Peripheral blood neutrophils. Stimulation with 19 periodontal bacteria | FITC NET-DNA, NE, and MPO | Certain species stimulated higher NET formation. |
| Doke et al. [57] | 2017 | Healthy donors | PMA-stimulated peripheral blood neutrophils. Nucleases from several periodontal bacteria. | SYTOX Orange, NE and DAPI | Prevotella intermedia demonstrated the highest NET degradation of all the Gram—periodontal bacteria |
| Roberts et al. [72] | 2016 | 5 Papillon–Lefévre syndrome (PLS) patients and 5 controls | Peripheral blood neutrophils stimulated with periodontal bacteria | SYTOX Green, NE, NET-bound MPO, NET-bound CG | Neutrophils from PLS patients have a reduced capacity for NET formation and a compromised antimicrobial activity |
| Palmer et al. [73] | 2016 | Healthy donors | Peripheral blood neutrophils incubated with oral bacteria in different complement blocking conditions | NET-DNA fluorometry | Complement and IgG enhance NET formation by several periodontal bacteria |
| Hirschfeld et al. [74] | 2016 | Healthy donors | Peripheral blood neutrophils with A.a., A.a. leucotoxin | Micrococcal nuclease | The leucotoxic strain of A.a. and high concentrations of A.a. leucotoxin induced NET formation |
| Jayaprakash et al. [56] | 2015 | Healthy donors | In vitro PMA-generated NETs; | FITC-labeled P. gingivalis, F-actin, DNA | P. gingivalis strains K1A and E8 induced NET formation |
| Palmer et al. [54] | 2012 | Healthy donors | In vitro PMA-generated NETs | DNase activity of periodontal bacterial species. SYTOX Green | DNase producing species caused the degradation of NETs |
| Other Studies Regarding NET Formation in Oral Neutrophils | |||||
| Moonen et al. [18] | 2019 | 9 Healthy donors | PMA-stimulated venous blood neutrophils and oral neutrophils | SYTOX Green | Oral neutrophils showed greater NET formation than circulating neutrophils in both stimulated and non-stimulated groups |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magán-Fernández, A.; Rasheed Al-Bakri, S.M.; O’Valle, F.; Benavides-Reyes, C.; Abadía-Molina, F.; Mesa, F. Neutrophil Extracellular Traps in Periodontitis. Cells 2020, 9, 1494. https://doi.org/10.3390/cells9061494
Magán-Fernández A, Rasheed Al-Bakri SM, O’Valle F, Benavides-Reyes C, Abadía-Molina F, Mesa F. Neutrophil Extracellular Traps in Periodontitis. Cells. 2020; 9(6):1494. https://doi.org/10.3390/cells9061494
Chicago/Turabian StyleMagán-Fernández, Antonio, Sarmad Muayad Rasheed Al-Bakri, Francisco O’Valle, Cristina Benavides-Reyes, Francisco Abadía-Molina, and Francisco Mesa. 2020. "Neutrophil Extracellular Traps in Periodontitis" Cells 9, no. 6: 1494. https://doi.org/10.3390/cells9061494
APA StyleMagán-Fernández, A., Rasheed Al-Bakri, S. M., O’Valle, F., Benavides-Reyes, C., Abadía-Molina, F., & Mesa, F. (2020). Neutrophil Extracellular Traps in Periodontitis. Cells, 9(6), 1494. https://doi.org/10.3390/cells9061494