C. quinoa and S. hispanica L. Seeds Provide Immunonutritional Agonists to Selectively Polarize Macrophages

 and
and 
Abstract
1. Introduction
2. Material and Methods
2.1. Cell Culture
2.2. Isolation of Protease Inhibitors (PPIs) and Cell Culture Treatments
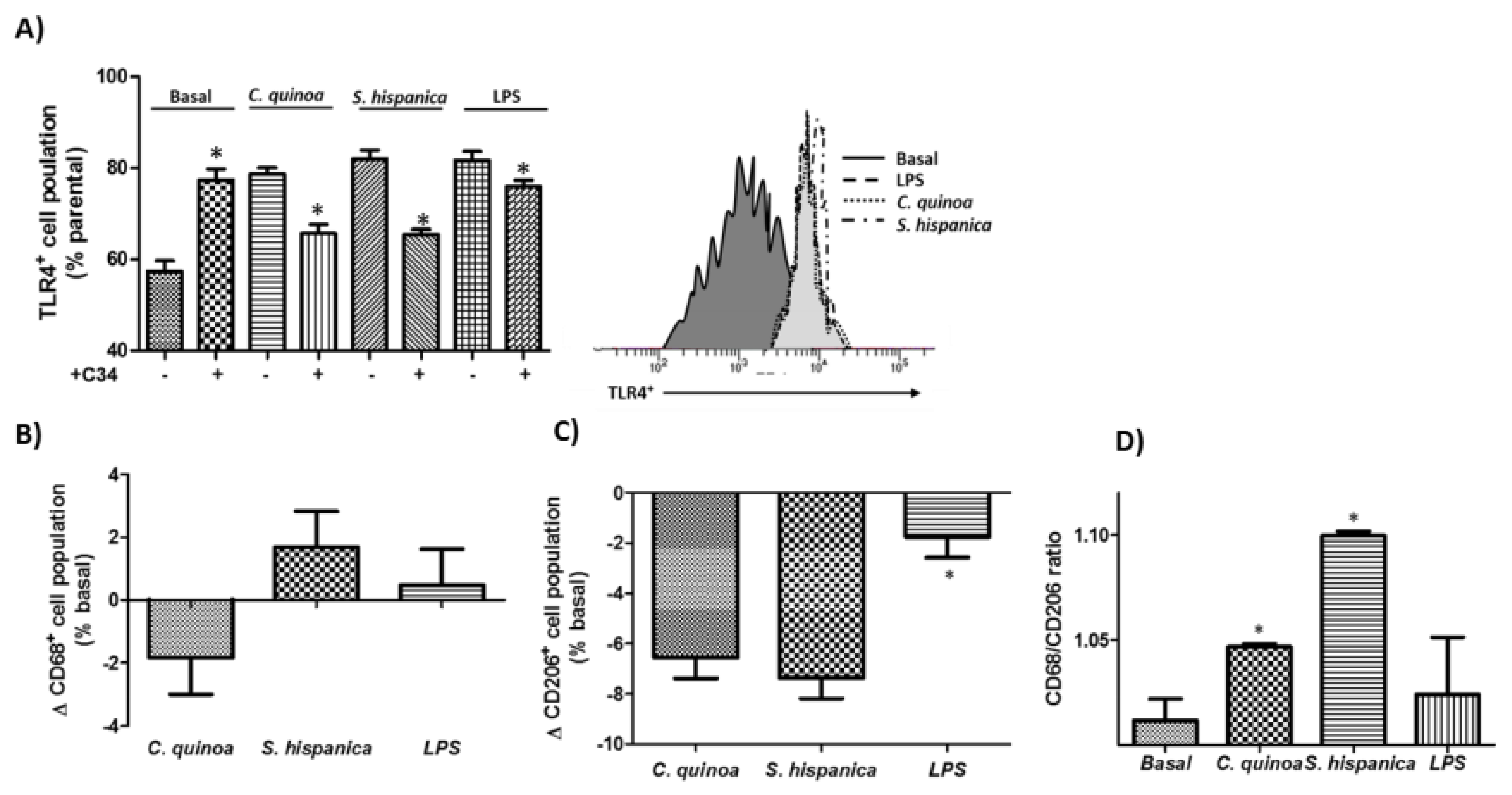
2.3. FACS Analysis for TLR4 Expression and Macrophage Phenotypic Differentiation
2.4. Cell ‘Mito’ Stress Test Assay
2.5. Measurement of Lysosomal (Neutral Red Assay) and Mitochondrial Enzyme (Test MTT) Activities
2.6. Phospholipid Assay
2.7. Relative Gene Expression Analyses
2.8. Cell Cultures Total Protein Extraction
2.9. In-Gel Digestion
2.10. Reverse Phase-Liquid Chromatography RP-LC-MS/MS Analysis
2.11. Statistical Analysis
3. Results and Discussion
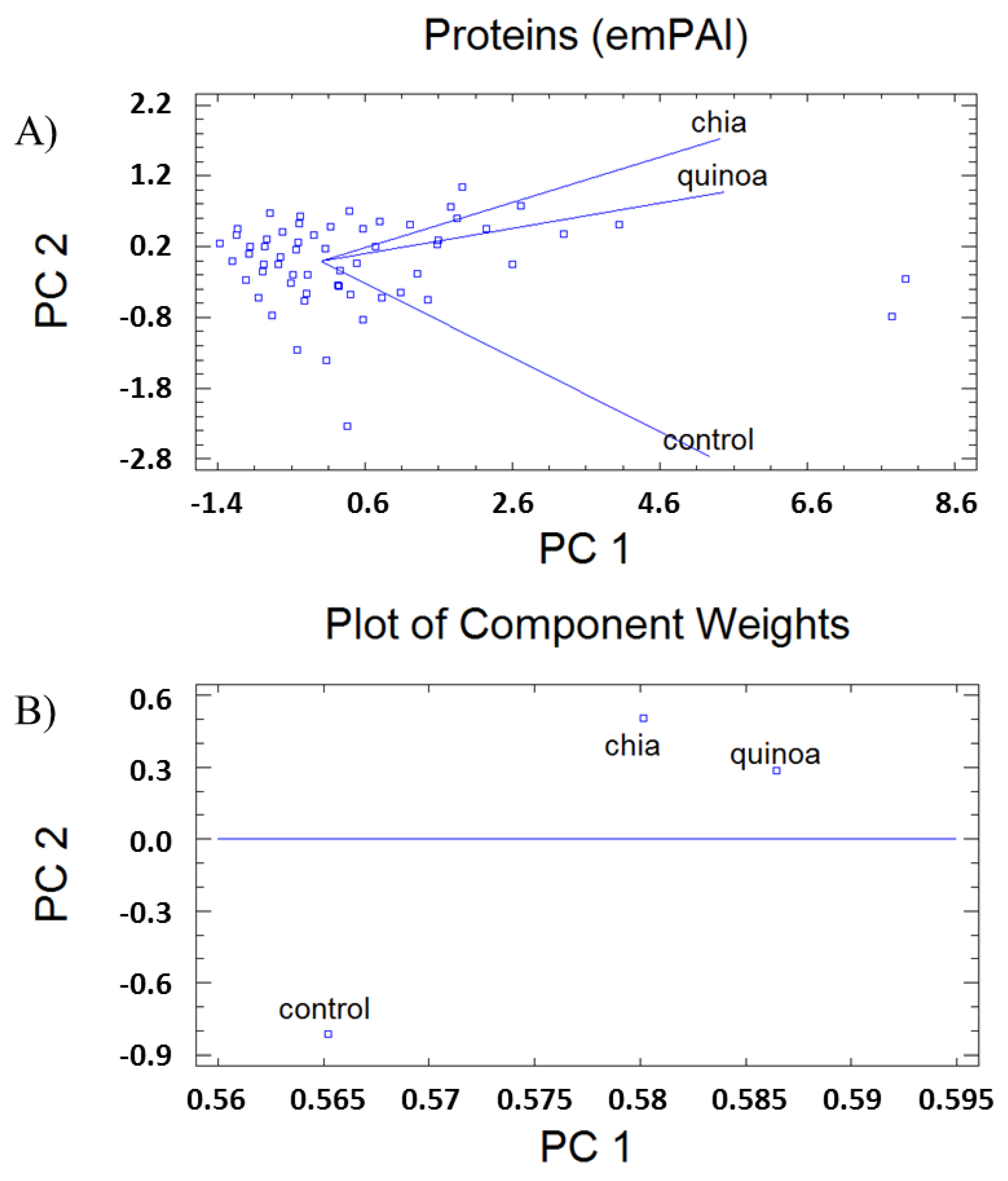
3.1. Protein Patterns
3.2. Macrophage Polarization
3.3. Immunonutritonal Changes
3.4. Phenotypic Changes
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Motta, B.M.; Grander, C.; Gögele, M.; Foco, L.; Vukovic, V.; Melotti, R.; Fuchsberger, C.; Grandi, A.D.; Cantaloni, C.; Picard, A.; et al. Microbiota, type 2 diabetes and non-alcoholic fatty liver disease: Protocol of an observational study. J. Transl. Med. 2019, 17, 408. [Google Scholar] [CrossRef] [PubMed]
- Kazankov, K.; Jørgensen, S.M.D.; Thomsen, K.L.; Møller, H.J.; Vilstrup, H.; George, J.; Schuppan, D.; Grønbæk, H. The role of macrophages in nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Junker, Y.; Zeissig, S.; Kim, S.J.; Barisani, D.; Wieser, H.; Leffler, D.A.; Zevallos, V.; Libermann, T.A.; Dillon, S.; Freitag, T.L.; et al. Wheat amylase trypsin inhibitors drive intestinal inflammation via activation of toll-like receptor 4. J. Exp. Med. 2012, 209, 2395–2408. [Google Scholar] [CrossRef] [PubMed]
- Laparra, J.M.; Fotschki, B.; Haros, C. Immunonutritional consequences of different serine-type protease inhibitors in a C57BL/6 Hepatocarcinoma model. Oncotarget 2019, 10, 760–772. [Google Scholar] [CrossRef]
- Srikanth, S.; Chen, Z. Plant Protease Inhibitors in Therapeutics-Focus on Cancer Therapy. Front. Pharmacol. 2016, 7, 470. [Google Scholar] [CrossRef]
- Laparra, J.M.; Haros, C. Plants seed protease inhibitors differentially affect innate immunity at tumor microenvironment to control hepatocarcinoma. Food Funct. 2019, 10, 4210–4219. [Google Scholar] [CrossRef]
- Zevallos, V.F.; Raker, V.; Tenzer, S.; Jimenez-Calvente, C.; Ashfaq-Khan, M.; Rüssel, N.; Pickert, G.; Schild, H.; Steinbrink, K.; Schuppan, D. Nutritional Wheat Amylase-Trypsin Inhibitors Promote Intestinal Inflammation via Activation of Myeloid Cells. Gastroenterology 2017, 152, 1100–1113. [Google Scholar] [CrossRef]
- Vega-Gálvez, A.; Miranda, M.; Vergara, J.; Uribe, E.; Puente, L.; Martínez, E.A. Nutrition facts and functional potential of quinoa (Chenopodium quinoa willd.), an ancient Andean grain: A review. J. Sci. Food Agric. 2010, 90, 2541–2547. [Google Scholar]
- EFSA. Safety of chia seeds (Salvia hispanica L.) as a novel food for extended uses pursuant to Regulation (EU) 2015/2283. EFSA J. 2019, 17, e05657. [Google Scholar]
- Brinegar, C.; Goundan, S. Isolation and characterization of chenopodin, the 11S seed storage protein of quinoa (Chenopodium quinoa). J. Agric. Food Chem. 1993, 41, 182–185. [Google Scholar] [CrossRef]
- Brinegar, C.; Sine, B.; Nwokocha, L. High-Cysteine 2S Seed Storage Proteins from Quinoa (Chenopodium quinoa). J. Agric. Food Chem. 1996, 44, 1621–1623. [Google Scholar] [CrossRef]
- Sandoval-Oliveros, M.R.; Paredes-López, O. Isolation and characterization of proteins from chia seeds (Salvia hispanica L.). J. Agric. Food Chem. 2013, 61, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Ju, C.; Tacke, F. Hepatic macrophages in homeostasis and liver diseases: From pathogenesis to novel therapeutic strategies. Cell. Mol. Immunol. 2016, 13, 316–327. [Google Scholar] [CrossRef]
- Laparra, J.M.; Alfonso-García, A.; Alegría, A.; Barberá, R.; Cilla, A. 7keto-stigmasterol and 7keto-cholesterol induce differential proteome changes to intestinal epitelial (Caco-2) cells. Food Chem. Toxicol. 2015, 84, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.L.; Escobar, J.; Izquierdo-Álvarez, A.; Gil, A.; Pérez, S.; Pereda, J.; Zapico, I.; Vento, M.; Sabater, L.; Marina, A.; et al. Disulfide stress: A novel type of oxidative stress in acute inflammation. Free Radic. Biol. Med. 2014, 70, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Alonso, R.; Pisa, D.; Marina, A.I.; Morato, E.; Rábano, A.; Rodal, I.; Carrasco, L. Evidence for Fungal Infection in Cerebrospinal Fluid and Brain Tissue from Patients with Amyotrophic Lateral Sclerosis. Int. J. Biol. Sci. 2015, 11, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Youle, R.J.; Huang, H.C. Occurrence of Low Molecular Weight and High Cysteine Containing Albumin Storage Proteins in Oilseeds of Diverse Species. Am. J. Bot. 1981, 68, 44–48. [Google Scholar] [CrossRef]
- Van den Bossche, J.; Baardman, J.; Otto, N.A.; van der Velden, S.; Neele, A.E.; van den Berg, S.M.; Luque-Martin, R.; Chen, H.J.; Boshuizen, M.C.; Ahmed, M.; et al. Mitochondrial Dysfunction Prevents Repolarization of Inflammatory Macrophages. Cell Rep. 2016, 17, 684–696. [Google Scholar] [CrossRef]
- Murakami, Y.; Hoshi, M.; Imamura, Y.; Arioka, Y.; Yamamoto, Y.; Saito, K. Remarkable role of indoleamine 2,3-dioxygenase and tryptophan metabolites in infectious diseases: Potential role in macrophage-mediated inflammatory diseases. Mediat. Inflamm. 2013, 2013, 391984. [Google Scholar] [CrossRef]
- van der Mark, V.A.; Ghiboub, M.; Marsman, C.; Zhao, J.; van Dijk, R.; Hiralall, J.K.; Ho-Mok, K.S.; Castricum, Z.; de Jonge, W.J.; Oude Elferink, R.P.; et al. Phospholipid flippases attenuate LPS-induced TLR4 signaling by mediating endocytic retrieval of Toll-like receptor 4. Cell Mol. Life Sci. 2017, 74, 715–730. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Yoshimura, A.; Kaneko, T.; Ukai, T.; Ozaki, Y.; Nakamura, H.; Li, X.; Matsumura, H.; Hara, Y.; Ogata, Y. A single nucleotide polymorphism in 3’-untranslated region contributes to the regulation of Toll-like receptor 4 translation. J. Biol. Chem. 2012, 287, 25163–25172. [Google Scholar] [CrossRef] [PubMed]
- 23 Rios, F.J.; Koga, M.M.; Pecenin, M.; Ferracini, M.; Gidlund, M.; Jancar, S. Oxidized LDL induces alternative macrophage phenotype through activation of CD36 and PAFR. Mediat. Inflamm. 2013, 2013, 198193. [Google Scholar]
- Feingold, K.R.; Shinegawa, J.K.; Cross, A.S.; Moser, A.; Grunfeld, C. Angiopoietin like protein 4 expression is decreased in activated macrophages. Biochem. Biophys. Res. Commun. 2012, 421, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Deiuliis, J.A.; Kampfrath, T.; Ying, Z.; Maiseyeu, A.; Rajagopalan, S. Lipoic acid attenuates innate immune infiltration and activation in the visceral adipose tissue of obese insulin resistant mice. Lipids 2011, 46, 1021–1032. [Google Scholar] [CrossRef]
- Galván-Peña, S.; O’Neill, L.A. Metabolic reprograming in macrophage polarization. Front. Immunol. 2014, 5, 420. [Google Scholar]
- Zhang, H. HIF-1 suppresses lipid catabolism to promote cancer progression. Mol. Cell Oncol. 2015, 2, e980184. [Google Scholar] [CrossRef]
- Wang, L.Y.; Diao, Z.L.; Zheng, J.F.; Wu, Y.R.; Zhang, Q.D.; Liu, W.H. Apelin attenuates TGF-β1-induced epithelial to mesenchymal transition via activation of PKC-ε in human renal tubular epithelial cells. Peptides 2017, 96, 44–52. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, Q. Activation of PPARγ by baicalin attenuates pulmonary hypertension in an infant rat model by suppressing HMGB1/RAGE signaling. FEBS Open Bio 2017, 7, 477–484. [Google Scholar] [CrossRef]
- Xie, L.; Liu, C.; Wang, L.; Gunawardena, H.P.; Yu, Y.; Du, R.; Taxman, D.J.; Dai, P.; Yan, Z.; Yu, J.; et al. Protein phosphatase 2A catalytic subunit α plays a MyD88-dependent, central role in the gene-specific regulation of endotoxin tolerance. Cell Rep. 2013, 3, 678–688. [Google Scholar] [CrossRef][Green Version]
- Mrakovic, A.; Kay, J.G.; Furuya, W.; Brumell, J.H.; Botelho, R.J. Rab7 and Arl8 GTPases are necessary for lysosome tubulation in macrophages. Traffic 2012, 13, 1667–1679. [Google Scholar] [CrossRef] [PubMed]
- Mallard, F.; Tang, B.L.; Galli, T.; Tenza, D.; Saint-Pol, A.; Yue, X.; Antony, C.; Hong, W.; Goud, B.; Johannes, L. Early/recycling endosomes-to-TGN transport involves two SNARE complexes and a Rab6 isoform. J. Cell Biol. 2002, 156, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lou, J.; Ouyang, C.; Chen, W.; Liu, Y.; Liu, X.; Cao, X.; Wang, J.; Lu, L. Ras-related protein Rab10 facilitates TLR4 signaling by promoting replenishment of TLR4 onto the plasma membrane. Proc. Natl. Acad. Sci. USA 2010, 107, 13806–13811. [Google Scholar] [CrossRef]
- Linehan, S.A.; Martínez-Pomares, L.; Stahl, P.D.; Gordon, S. Mannose receptor and its putative ligands in normal murine lymphoid and nonlymphoid organs: In situ expression of mannose receptor by selected macrophages, endothelial cells, perivascular microglia, and mesangial cells, but not dendritic cells. J. Exp. Med. 1999, 189, 1961–1972. [Google Scholar] [CrossRef] [PubMed]
- Gangloff, M. Different dimerisation mode for TLR4 upon endosomal acidification? Trends Biochem. Sci. 2012, 37, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Kumazoe, M.; Nakamura, Y.; Yamashita, M.; Suzuki, T.; Takamatsu, K.; Huang, Y.; Bae, J.; Yamashita, S.; Murata, M.; Yamada, S.; et al. Green Tea Polyphenol Epigallocatechin-3-gallate Suppresses Toll-like Receptor 4 Expression via Up-regulation of E3 Ubiquitin-protein Ligase RNF216. J. Biol. Chem. 2017, 292, 4077–4088. [Google Scholar] [CrossRef]
- Kumazoe, M.; Yamashita, M.; Nakamura, Y.; Takamatsu, K.; Bae, J.; Yamashita, S.; Yamada, S.; Onda, H.; Nojiri, T.; Kangawa, K.; et al. Green Tea Polyphenol EGCG Upregulates Tollip Expression by Suppressing Elf-1 Expression. J. Immunol. 2017, 199, 3261–3269. [Google Scholar] [CrossRef]
- Silverstein, R.L.; Febbraio, M. CD36, a scavenger receptor involved in immunity, metabolism, angiogenesis, and behavior. Sci. Signal. 2009, 2, re3. [Google Scholar] [CrossRef]
- Kobayashi, H. Endogenous anti-inflammatory substances, inter-alpha-inhibitor and bikunin. Biol. Chem. 2006, 387, 1545–1549. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nº (#) | Protein a (Accession) | Mr | pI | Mean Fold Difference b | pc | |
|---|---|---|---|---|---|---|
| C. quinoa | S. hispanica L. | |||||
| 1 | Heat shock protein HSP 90-beta (P08238) | 83.2 | 5.03 | 1.23 ± 0.28 | 0.89 ± 0.28 | 0.103 |
| 2 | Pyruvate kinase (P14618) | 57.9 | 7.84 | 0.13 ± 0.03 * | 0.11 ± 0.03 * | 0.446 |
| 3 | Malate dehydrogenase (P40926) | 35.5 | 8.68 | 1.92 ± 0.44 * | 1.66 ± 0.53 | 0.444 |
| 4 | Polyubiquitin-C (P0CG48) | 77.0 | 7.66 | 1.34 ± 0.31 | 1.16 ± 0.37 | 0.445 |
| 5 | Transitional endoplasmic reticulum ATPase (P55072) | 89.3 | 5.26 | 1.40 ± 0.32 | 1.22 ± 0.39 | 0.443 |
| 6 | T-complex protein 1 subunit zeta (P40227) | 58.0 | 6.68 | 0.24 ± 0.05 * | 0.21 ± 0.07 * | 0.446 |
| 7 | Protein disulfide-isomerase (P07237) | 57.1 | 4.87 | 1.30 ± 0.30 | 0.97 ± 0.31 | 0.124 |
| 8 | Peptidyl-prolyl cis-trans isomerase A (P62937) | 18.0 | 7.81 | 1.71 ± 0.38 | 1.25 ± 0.40 | 0.124 |
| 9 | 6-phosphogluconate dehydrogenase (P52209-2) | 51.8 | 7.44 | 1.61 ± 0.38 | 1.46 ± 0.46 | 0.445 |
| 10 | Ras-related protein Rab-7a (P51149) | 23.5 | 6.70 | 0.96 ± 0.22 | 0.62 ± 0.20 | 0.037 |
| 11 | Aldehyde dehydrogenase (P05091-2) | 51.0 | 6.20 | 0.28 ± 0.06 * | 0.97 ± 0.31 | 0.001 |
| 12 | D-3-phosphoglycerate dehydrogenase (O43175) | 56.6 | 6.71 | 1.68 ± 0.38 | 1.45 ± 0.47 | 0.445 |
| 13 | L-lactate dehydrogenase B chain (P07195) | 36.6 | 6.05 | 1.96 ± 0.45 * | 0.97 ± 0.31 | 0.004 |
| 14 | Sodium/potassium-transporting ATPase (P05023-3) | 109.5 | 5.31 | 1.68 ± 0.38 | 0.36 ± 0.11 * | 0.001 |
| 15 | Fructose-bisphosphate aldolase A (P04075) | 39.4 | 8.09 | 0.72 ± 0.16 | 1.45 ± 0.45 | 0.010 |
| 16 | Annexin A5 (P08758) | 35.9 | 5.05 | 2.01 ± 0.46 * | 2.04 ± 0.65 | 0.932 |
| 17 | Voltage-dependent anion-selective channel (P45880-2) | 30.4 | 7.20 | 1.34 ± 0.31 | 1.16 ± 0.38 | 0.445 |
| 18 | L-lactate dehydrogenase A chain (P00338) | 36.7 | 8.27 | 1.34 ± 0.31 | 0.87 ± 0.28 | 0.037 |
| 19 | Macrophage-capping protein (P40121-2) | 36.8 | 7.20 | 1.67 ± 0.38 | 1.45 ± 0.46 | 0.446 |
| 20 | Dolichyl-diphosphooligosaccharide-glycosyltransferase (P39656-3) | 49.0 | 6.86 | 0.33 ± 0.88 * | 0.29 ± 0.09 * | 0.446 |
| 21 | Nucleoside diphosphate kinase A (P15531) | 17.1 | 6.19 | 1.68 ± 0.27 | 1.45 ± 0.46 | 0.445 |
| 22 | T-complex protein 1 subunit delta (P50991) | 57.9 | 7.83 | 1.20 ± 0.27 | 0.21 ± 0.07 * | 0.001 |
| 23 | Ubiquitin-like modifier-activating enzyme 1 (P22314-2) | 113.7 | 5.60 | 0.67 ± 0.15 | 0.58 ± 0.17 | 0.446 |
| 24 | Serine hydroxymethyl-transferase (P34897-3) | 53.4 | 8.15 | 1.70 ± 0.38 | 1.42 ± 0.32 | 0.445 |
| 25 | Malate dehydrogenase (P40925-2) | 27.0 | 9.11 | 1.67 ± 0.38 | 1.54 ± 0.41 | 0.446 |
| 26 | Serine/threonine kinase receptor-associated protein (Q9Y3F4) | 38.4 | 5.12 | 0.42 ± 0.10 * | 0.36 ± 0.11 * | 0.446 |
| 27 | Glucose-6-phosphate isomerase (P06744) | 63.1 | 8.32 | 1.22 ± 0.25 | 0.97 ± 0.13 | 0.446 |
| 28 | T-complex protein 1 subunit α (P17987) | 60.3 | 6.11 | 1.12 ± 0.19 | 0.48 ± 0.15 * | 0.001 |
| 29 | Chloride intracellular channel protein 1 (O00299) | 26.9 | 5.17 | 1.86 ± 0.34 | 1.39 ± 0.43 | 0.446 |
| 30 | Nascent polypeptide-associated complex subunit α (Q13765) | 23.4 | 4.56 | 1.12 ± 0.25 | 0.49 ± 0.15 * | 0.002 |
| 31 | Rho GDP-dissociation inhibitor 1 (P52565) | 23.2 | 5.11 | 2.51 ± 0.57 * | 1.54 ± 0.62 | 0.013 |
| 32 | Glucose-6-phosphate 1-dehydrogenase (P11413) | 59.2 | 6.84 | 1.00 ± 0.23 | 0.87 ± 0.27 | 0.446 |
| 33 | Glyceraldehyde-3-phosphate dehydrogenase (P04406) | 36.0 | 8.46 | 0.56 ± 0.13 * | 0.48 ± 0.15 * | 0.446 |
| 34 | Dihydrolipoyl dehydrogenase (P09622-2) | 43.6 | 7.03 | 1.67 ± 0.38 | 0.72 ± 0.23 | 0.002 |
| 35 | Ras-related protein Rab-10 (P61026) | 22.5 | 8.38 | 1.68 ± 0.38 | 0.97 ± 0.26 | 0.013 |
| 36 | V-type proton ATPase catalytic subunit A (P38606-2) | 64.7 | 5.66 | 1.12 ± 0.25 | 0.51 ± 0.15 * | 0.002 |
| 37 | Poly(rC)-binding protein 1 (Q15365) | 37.5 | 7.09 | 2.24 ± 0.51 * | 1.45 ± 0.46 | 0.037 |
| 38 | Voltage-dependent anion-selective channel protein 1 (P21796) | 30.8 | 8.54 | 1.26 ± 0.29 | 0.36 ± 0.11 * | 0.001 |
| 39 | T-complex protein 1 subunit ε (P48643-2) | 49.5 | 6.19 | 1.55 ± 0.09 | 0.42 ± 0.12 * | 0.001 |
| 40 | Peptidyl-prolyl cis-trans isomerase FKBP4 (Q02790) | 51.8 | 5.43 | 0.42 ± 0.10 * | 0.38 ± 0.13 * | 0.446 |
| 41 | Ras-related protein Rab-1A (P62820) | 22.7 | 6.21 | 0.56 ± 0.13 * | 0.97 ± 0.31 | 0.025 |
| 42 | T-complex protein 1 subunit eta(Q99832-3) | 54.8 | 7.68 | 1.68 ± 0.38 | 0.49 ± 0.14 * | 0.001 |
| 43 | Ras-related protein Rab-2A (P61019-2) | 20.8 | 6.11 | 1.67 ± 0.38 | 0.72 ± 0.23 | 0.002 |
| 44 | Isocitrate dehydrogenase [NADP] cytoplasmic (O75874) | 46.6 | 7.01 | 1.51 ± 0.33 | 1.42 ± 0.08 | 0.446 |
| 45 | Isocitrate dehydrogenase [NADP], mitochondrial (P48735-2) | 45.2 | 7.75 | 2.51 ± 0.57 * | 0.72 ± 0.32 | 0.001 |
| 46 | Fermitin family homolog 3 (Q86UX7-2) | 75.4 | 6.77 | 0.56 ± 0.13 * | 0.48 ± 0.13 * | 0.446 |
| 47 | Dolichyl-diphosphooligosaccharide--protein glycosyltransferase subunit 2 (P04844-2) | 67.7 | 6.06 | 0.84 ± 0.19 | 0.79 ± 0.23 | 0.446 |
| 48 | Receptor of activated protein C kinase 1 (P63244) | 35.1 | 7.69 | 3.35 ± 0.77 * | 1.56 ± 0.46 | 0.001 |
| 49 | Ceruloplasmin (P00450) | 122.1 | 5.72 | 2.51 ± 0.57 * | 2.18 ± 0.70 | 0.446 |
| 50 | Hypoxia up-regulated protein 1 (Q9Y4L1) | 111.3 | 5.22 | 0.84 ± 0.19 | 0.76 ± 0.32 | 0.446 |
| 51 | SRP55-3 of Serine/arginine-rich splicing factor 6 (Q13247-3) | 38.4 | 11.00 | 1.67 ± 0.38 | 0.87 ± 0.41 | 0.002 |
| 52 | High mobility group protein B1 (P09429) | 24.9 | 5.74 | 0.84 ± 0.19 | 0.69 ± 0.17 | 0.446 |
| 53 | Cell division control protein 42 homolog (P60953) | 21.2 | 6.55 | 0.91 ± 0.12 | 1.67 ± 0.26 | 0.025 |
| 54 | Ran GTPase-activating protein 1 (P46060) | 63.5 | 4.68 | 1.69 ± 0.28 | 0.72 ± 0.17 | 0.002 |
| 55 | NADPH-cytochrome P450 reductase (P16435) | 76.6 | 5.58 | 1.75 ± 0.42 | 0.81 ± 0.22 | 0.002 |
| 56 | Glutamate dehydrogenase 1 (P00367-3) | 46.5 | 7.08 | 0.84 ± 0.19 | 0.76 ± 0.21 | 0.446 |
| 57 | Ras GTPase-activating-like protein IQGAP1 (P46940) | 189.1 | 6.48 | 0.84 ± 0.19 | 0.79 ± 0.18 | 0.446 |
| 58 | Proliferation-associated protein 2G4 (Q9UQ80-2) | 38.0 | 7.53 | 1.26 ± 0.29 | 0.54 ± 0.17 | 0.001 |
| 59 | Protein disulfide-isomerase A3 (P30101) | 56.7 | 6.35 | 1.33 ± 0.19 | 0.47 ± 0.09 | 0.001 |
| 60 | GTP-binding nuclear protein Ran (P62826) | 24.4 | 7.49 | 2.24 ± 0.51 * | 1.94 ± 0.62 | 0.445 |
| 61 | Calnexin (P27824) | 67.5 | 4.60 | 0.89 ± 0.19 | 0.76 ± 0.32 | 0.446 |
| 62 | Serine/threonine-protein phosphatase 2A 55 kDa regulatory subunit B (P63151) | 51.7 | 6.20 | 0.85 ± 0.16 | 1.45 ± 0.34 | 0.025 |
| 63 | Apoptosis-inducing factor 1 (O95831-3) | 66.3 | 8.94 | 0.56 ± 0.13 * | 0.48 ± 0.15 * | 0.446 |
| 64 | Phosphoglycerate kinase 1 (P00558) | 44.6 | 8.10 | 1.92 ± 0.44 * | 1.64 ± 0.36 | 0.148 |
| 65 | Phosphoglycerate mutase 1 (P18669) | 28.8 | 7.18 | 1.81 ± 0.38 | 1.25 ± 0.40 | 0.124 |
| 66 | 40S ribosomal protein S3 (P23396) | 26.7 | 9.66 | 1.66 ± 0.28 | 1.13 ± 0.36 | 0.050 |
| 67 | Rab GDP dissociation inhibitor β (P50395) | 50.6 | 6.47 | 0.42 ± 0.10 * | 0.36 ± 0.11 * | 0.446 |
| 68 | T-complex protein 1 subunit γ (P49368) | 60.5 | 6.49 | 0.21 ± 0.05 * | 1.09 ± 0.35 | 0.001 |
| 69 | T-complex protein 1 subunit τ (P50990-3) | 51.6 | 5.24 | 1.12 ± 0.25 | 0.48 ± 0.16 * | 0.001 |
| 70 | Serine/threonine-protein phosphatase PP1-α (P62136) | 37.5 | 6.33 | 1.00 ± 0.23 | 0.29 ± 0.09 * | 0.002 |
| 71 | T-complex protein 1 subunit β (P78371-2) | 52.7 | 6.44 | 2.10 ± 0.48 * | 1.56 ± 0.36 | 0.066 |
| 72 | Serine/threonine-protein phosphatase PP1-γ (P36873) | 37.0 | 6.54 | 1.26 ± 0.29 | 0.37 ± 0.10 * | 0.001 |
| 73 | Peroxiredoxin-1 (Q06830) | 22.1 | 8.13 | 1.20 ± 0.27 | 0.83 ± 0.26 | 0.066 |
| 74 | Sarcoplasmic/endoplasmic reticulum calcium ATPase 2 (P16615-2) | 109.6 | 5.36 | 1.12 ± 0.25 | 0.65 ± 0.18 * | 0.002 |
| 75 | Plasminogen activator inhibitor 1 RNA-binding protein (Q8NC51-4) | 42.4 | 8.44 | 1.67 ± 0.38 | 1.74 ± 0.42 | 0.446 |
| 76 | Proliferating cell nuclear antigen (P12004) | 28.8 | 4.69 | 0.84 ± 0.19 | 0.72 ± 0.23 | 0.446 |
| 77 | Proteasome subunit beta type-5 (P28074) | 28.5 | 6.92 | 0.42 ± 0.10 * | 0.18 ± 0.05 * | 0.002 |
| 78 | Splicing factor, proline- and glutamine-rich (P23246) | 76.1 | 9.44 | 1.12 ± 0.25 | 1.78 ± 0.52 | 0.190 |
| 79 | Protein disulfide-isomerase A4 (P13667) | 72.9 | 5.07 | 1.68 ± 0.38 | 0.97 ± 0.26 | 0.013 |
| 80 | Nucleosome assembly protein 1-like 1 (P55209-2) | 42.7 | 4.55 | 0.56 ± 0.13 * | 0.84 ± 0.16 * | 0.446 |
| 81 | Ubiquitin-conjugating enzyme E2 N (P61088) | 17.1 | 6.57 | 0.84 ± 0.19 | 0.72 ± 0.23 | 0.446 |
| 82 | Calcyclin-binding protein (Q9HB71) | 26.2 | 8.25 | 0.96 ± 0.15 | 0.87 ± 0.32 | 0.446 |
| 83 | SRP40-4 of Serine/arginine-rich splicing factor 5 (Q13243-3) | 30.8 | 11.66 | 0.81 ± 0.14 | 0.76 ± 0.13 | 0.446 |
| 84 | Prostaglandin E synthase 3 (Q15185-3) | 14.9 | 4.77 | 0.80 ± 0.05 | 0.81 ± 0.09 | 0.446 |
| 85 | V-type proton ATPase subunit C1 (P21283) | 43.9 | 7.46 | 0.88 ± 0.19 | 0.72 ± 0.06 | 0.446 |
| 86 | Heat shock protein HSP 90-α (P07900) | 84.6 | 5.02 | 1.32 ± 0.30 | 0.99 ± 0.32 | 0.135 |
| 87 | RuvB-like 1 (Q9Y265) | 42.1 | 6.38 | 0.56 ± 0.13 * | 0.48 ± 0.15 * | 0.446 |
| 88 | Phospholipase A-2-activating protein (Q9Y263) | 50.2 | 6.42 | 1.67 ± 0.38 | 1.75 ± 0.27 | 0.446 |
| 89 | Ras-related protein Rab-14 (P61106) | 87.1 | 6.37 | 5.03 ± 1.15 * | 2.91 ± 0.39 * | 0.013 |
| 90 | Prolyl endopeptidase (P48147) | 23.9 | 6.21 | 1.75 ± 0.31 | 1.45 ± 0.44 | 0.446 |
| 91 | V-type proton ATPase subunit B (P21281) | 80.6 | 5.86 | 1.67 ± 0.42 | 1.42 ± 0.12 | 0.446 |
| 92 | Ras-related protein Rab-6A (P20340) | 56.5 | 5.81 | 1.57 ± 0.38 | 1.45 ± 0.08 | 0.446 |
| 93 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit 1 (P04843) | 23.6 | 5.54 | 1.37 ± 0.36 | 2.87 ± 0.64 * | 0.025 |
| 94 | Mitochondrial import receptor subunit TOM40 homolog (O96008-2) | 68.5 | 6.38 | 3.35 ± 0.76 * | 1.28 ± 0.24 | 0.002 |
| 95 | Ras-related protein Rab-11B (Q15907) | 34.4 | 7.24 | 1.71 ± 0.38 | 1.54 ± 0.46 | 0.446 |
| 96 | Serine/arginine-rich splicing factor 1 (Q07955) | 24.5 | 5.94 | 1.69 ± 0.23 | 1.72 ± 0.31 | 0.446 |
| 97 | Inter-α-trypsin inhibitor heavy chain H2 (P19823) | 27.7 | 10.35 | 1.66 ± 0.35 | 2.91 ± 0.93 * | 0.025 |
| 98 | Ras GTPase-activating protein-binding protein 1 (Q13283) | 106.3 | 6.85 | 1.59 ± 0.37 | 1.33 ± 0.46 | 0.446 |
| 99 | Clathrin heavy chain 1 (Q00610-2) | 187.8 | 5.69 | 1.68 ± 0.38 | 0.29 ± 0.09 * | 0.001 |
| 100 | Proteasome subunit beta type-4 (P28070) | 29.2 | 5.97 | 0.84 ± 0.19 | 0.72 ± 0.23 | 0.446 |
| 101 | Guanine nucleotide-binding protein G ‘α’ (P04899) | 40.5 | 5.54 | 1.76 ± 0.26 * | 1.45 ± 0.31 | 0.013 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srdić, M.; Ovčina, I.; Fotschki, B.; Haros, C.M.; Laparra Llopis, J.M. C. quinoa and S. hispanica L. Seeds Provide Immunonutritional Agonists to Selectively Polarize Macrophages. Cells 2020, 9, 593. https://doi.org/10.3390/cells9030593
Srdić M, Ovčina I, Fotschki B, Haros CM, Laparra Llopis JM. C. quinoa and S. hispanica L. Seeds Provide Immunonutritional Agonists to Selectively Polarize Macrophages. Cells. 2020; 9(3):593. https://doi.org/10.3390/cells9030593
Chicago/Turabian StyleSrdić, Maša, Ivana Ovčina, Bartosz Fotschki, Claudia Monika Haros, and Jose Moises Laparra Llopis. 2020. "C. quinoa and S. hispanica L. Seeds Provide Immunonutritional Agonists to Selectively Polarize Macrophages" Cells 9, no. 3: 593. https://doi.org/10.3390/cells9030593
APA StyleSrdić, M., Ovčina, I., Fotschki, B., Haros, C. M., & Laparra Llopis, J. M. (2020). C. quinoa and S. hispanica L. Seeds Provide Immunonutritional Agonists to Selectively Polarize Macrophages. Cells, 9(3), 593. https://doi.org/10.3390/cells9030593