Leucine Improved Growth Performance, Muscle Growth, and Muscle Protein Deposition Through AKT/TOR and AKT/FOXO3a Signaling Pathways in Hybrid Catfish Pelteobagrus vachelli × Leiocassis longirostris


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Feeding Management
2.3. Sample Collection
2.4. Histological Analysis of Muscle
2.5. Real-Time Quantitative PCR
2.6. Western Blotting
2.7. Calculations and Statistical Analysis
3. Results
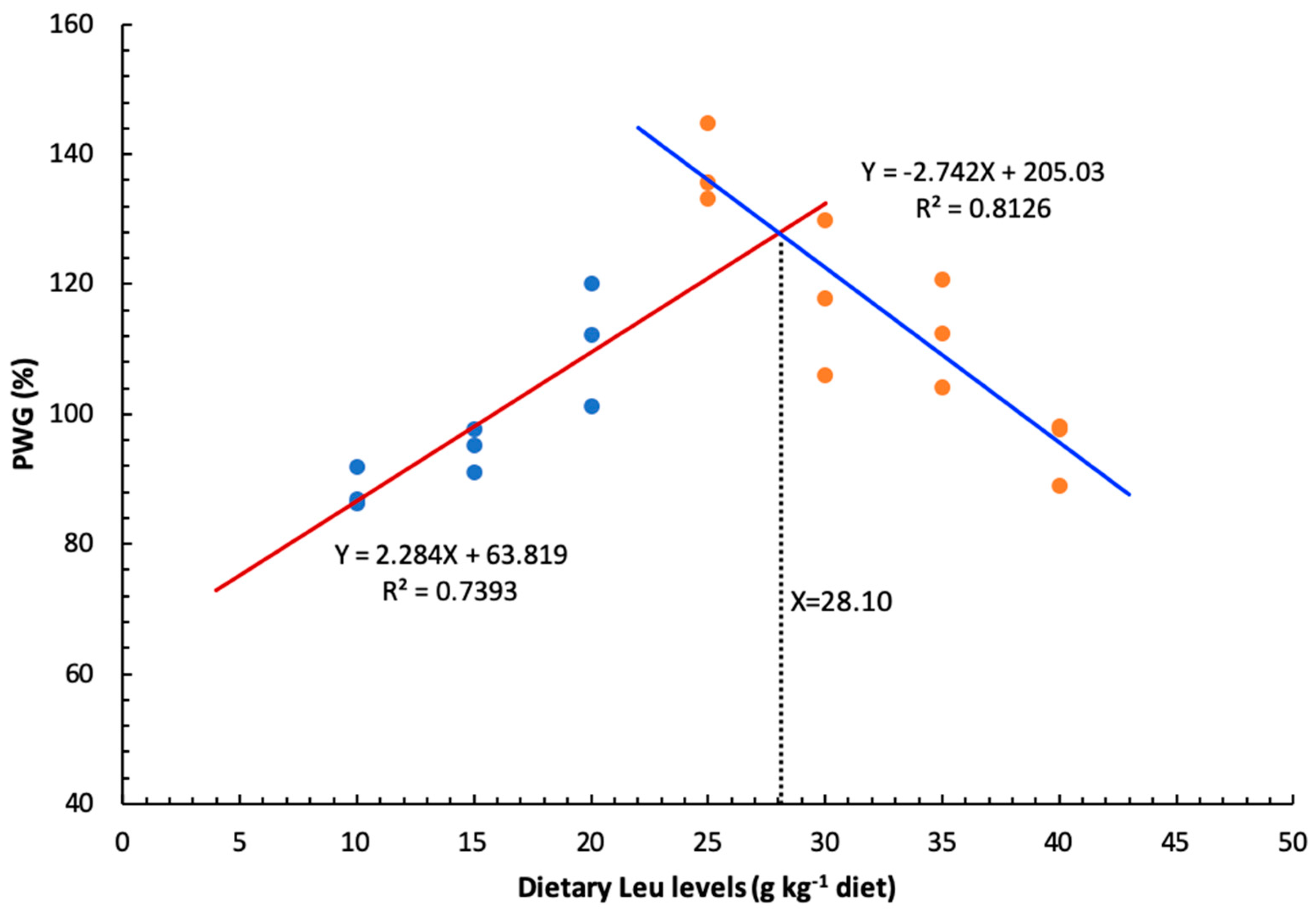
3.1. Effect of Dietary Leu on Growth Performance
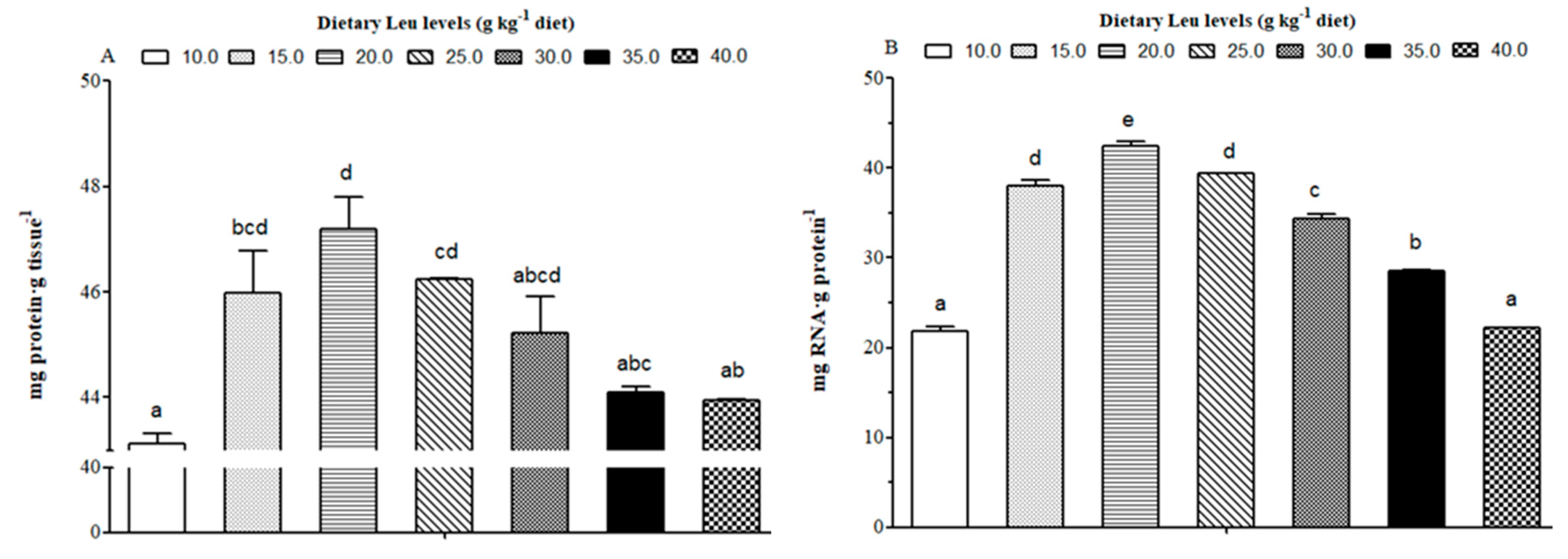
3.2. Effects of Dietary Leu on Muscle Fiber Diameter and Density, Protein Content, and RNA/Protein Ratio
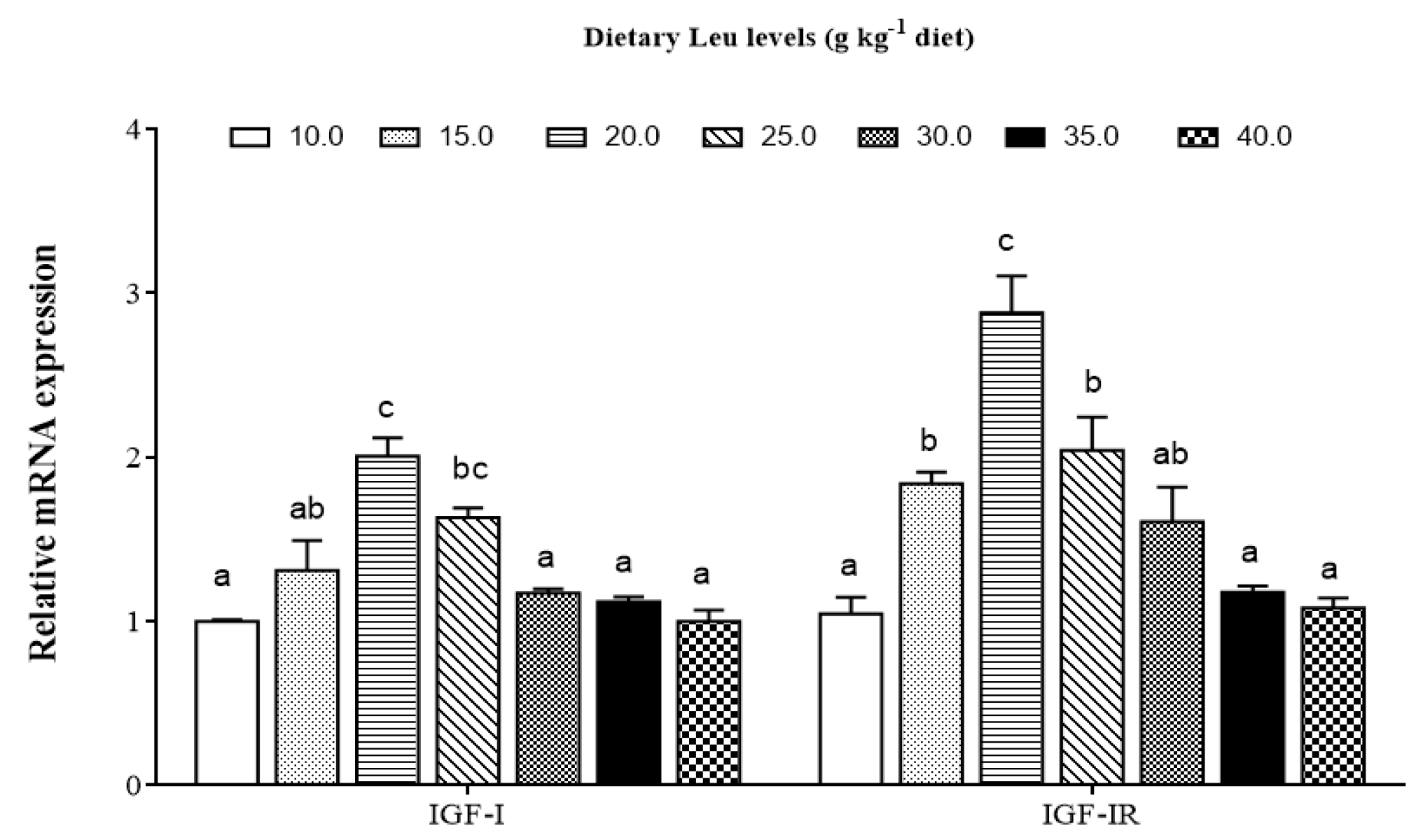
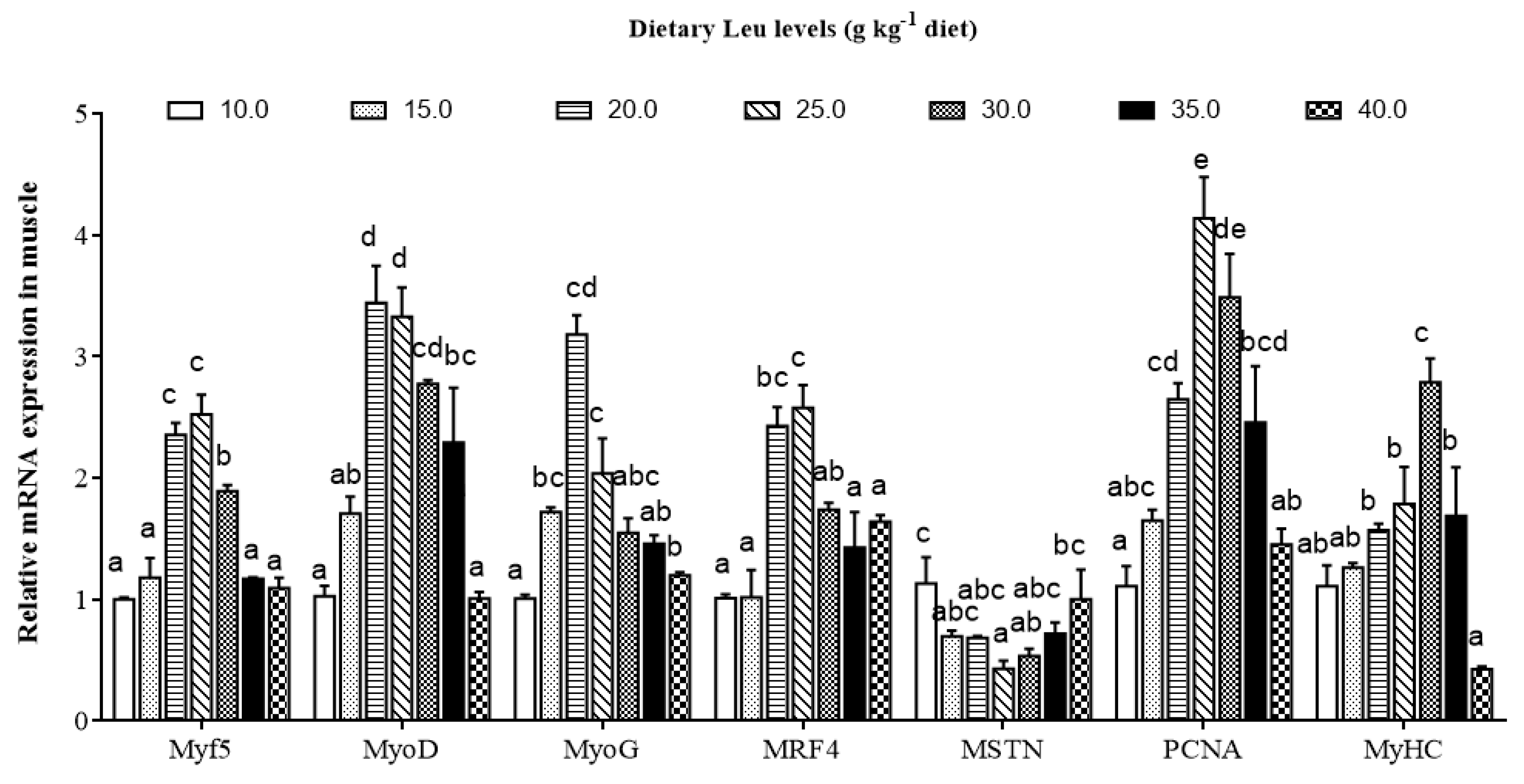
3.3. Effects of Dietary Leu on IGF-I and Muscle Growth Related Gene mRNA Expressions
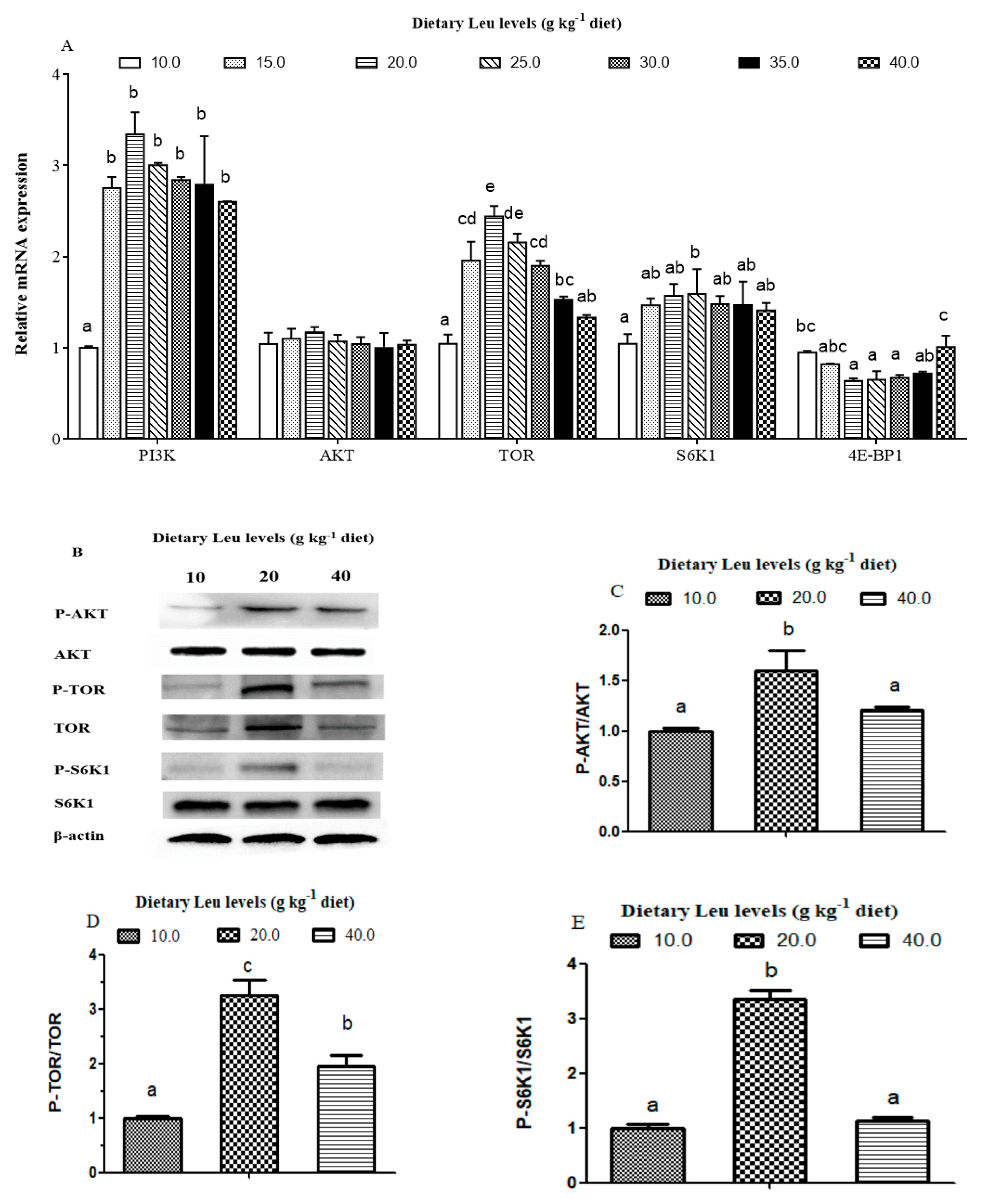
3.4. Effects of Dietary Leu on PI3K/AKT/TOR Signaling Pathway
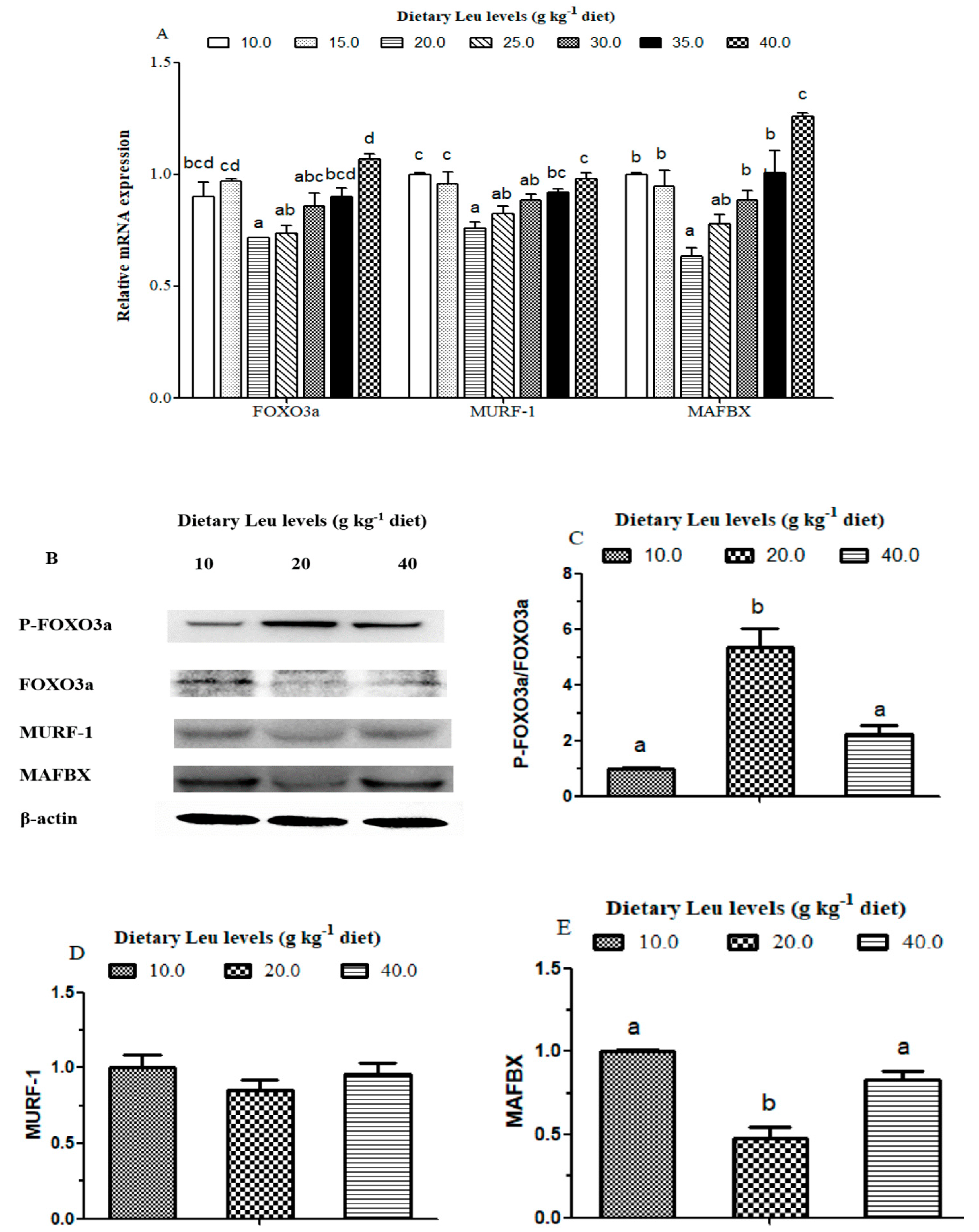
3.5. Effects of Dietary Leu on Protein Degradation Related Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vélez, E.J.; Lutfi, E.; Azizi, S.; Perelló, M.; Salmerón, C.; Riera-Codina, M.; Ibarz, A.; Fernández-Borràs, J.; Blasco, J.; Capilla, E.; et al. Understanding fish muscle growth regulation to optimize aquaculture production. Aquaculture 2017, 467, 28–40. [Google Scholar] [CrossRef]
- Periago, M.J.; Ayala, M.D.; López-Albors, O.; Abdel, I.; Martínez, C.; García-Alcázar, A.; Ros, G.; Gil, F. Muscle cellularity and flesh quality of wild and farmed sea bass, Dicentrarchus labrax L. Aquaculture 2005, 249, 175–188. [Google Scholar] [CrossRef]
- Johnston, I.A.; Bower, N.I.; Macqueen, D.J. Growth and the regulation of myotomal muscle mass in teleost fish. J. Exp. Biol. 2011, 214, 1617. [Google Scholar] [CrossRef] [PubMed]
- Rowlerson, A.; Veggetti, A. 5-Cellular mechanisms of post-embryonic muscle growth in aquaculture species. Fish Physiology; Johnston, I.A., Ed.; Academic Press: San Diego, CA, USA, 2001; Volume 18, pp. 103–140. [Google Scholar]
- Johnston, I.A.; Lee, H.; Macqueen, D.J.; Paranthaman, K.; Kawashima, C.; Anwar, A.; Kinghorn, J.R.; Dalmay, T. Embryonic temperature affects muscle fibre recruitment in adult zebrafish: Genome-wide changes in gene and microRNA expression associated with the transition from hyperplastic to hypertrophic growth phenotypes. J. Exp. Biol. 2009, 212, 1781–1793. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.W.; Duan, C.H. Insulin-like growth factor signaling in fish. Int. Rev. Cytol. 2005, 243, 215–285. [Google Scholar] [PubMed]
- Musarò, A.; McCullagh, K.J.A.; Naya, F.J.; Olson, E.N.; Rosenthal, N. IGF-1 induces skeletal myocyte hypertrophy through calcineurin in association with GATA-2 and NF-ATc1. Nature 1999, 400, 581–585. [Google Scholar] [CrossRef]
- Rommel, C.; Bodine, S.C.; Clarke, B.A.; Rossman, R.; Nunez, L.; Stitt, T.N.; Yancopoulos, G.D.; Glass, D.J. Mediation of IGF-1-induced skeletal myotube hypertrophy by PI(3)K/Akt/mTOR and PI(3)K/Akt/GSK3 pathways. Nat. Cell. Biol. 2001, 3, 1009–1013. [Google Scholar] [CrossRef]
- Rommel, C.; Clarke, B.A.; Zimmermann, S.; Nuñez, L.; Rossman, R.; Reid, K.; Moelling, K.; Yancopoulos, G.D.; Glass, D.J. Differentiation stage-specific inhibition of the Raf-MEK-ERK pathway by Akt. Science 1999, 286, 1738–1741. [Google Scholar] [CrossRef]
- Buckingham, M.; Rigby, P.W.J. Gene regulatory networks and transcriptional mechanisms that control myogenesis. Dev. Cell 2014, 28, 225–238. [Google Scholar] [CrossRef]
- Berkes, C.A.; Tapscott, S.J. MyoD and the transcriptional control of myogenesis. Semin. Cell Dev. Biol. 2005, 16, 585–595. [Google Scholar] [CrossRef]
- Watabe, S. 2. Myogenic regulatory factors. Fish Physiol. 2001, 18, 19–41. [Google Scholar]
- Lee, S.; McPherron, A.C. Regulation of myostatin activity and muscle growth. Proc. Natl. Acad. Sci. USA 2001, 98, 9306–9311. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, L.; Lu, Z.; Gao, J.E.; Chu, Y.; Li, L.; Wang, M.; Zhang, G.; Zhang, M.; Ye, J. The effects of dietary leucine on the growth performances, body composition, metabolic abilities and innate immune responses in black carp Mylopharyngodon piceus. Fish Shellfish Immun. 2017, 67, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ai, Q.; Mai, K.; Xu, W.; Cheng, Z.; He, Z. Dietary leucine requirement for juvenile large yellow croaker Pseudosciaena crocea (Richardson, 1846). J. Ocean Univ. China 2010, 9, 371–375. [Google Scholar] [CrossRef]
- Choo, P.; Smith, T.K.; Cho, C.Y.; Ferguson, H.W. Dietary excesses of leucine influence growth and body composition of rainbow trout. J. Nutr. 1991, 121, 1932–1939. [Google Scholar] [CrossRef]
- Tan, X.; Lin, H.; Huang, Z.; Zhou, C.; Wang, A.; Qi, C.; Zhao, S. Effects of dietary leucine on growth performance, feed utilization, non-specific immune responses and gut morphology of juvenile golden pompano Trachinotus ovatus. Aquaculture 2016, 465, 100–107. [Google Scholar] [CrossRef]
- Deng, Y.; Jiang, W.; Liu, Y.; Jiang, J.; Kuang, S.; Tang, L.; Wu, P.; Zhang, Y.; Feng, L.; Zhou, X. Differential growth performance, intestinal antioxidant status and relative expression of Nrf2 and its target genes in young grass carp (Ctenopharyngodon idella) fed with graded levels of leucine. Aquaculture 2014, 434, 66–73. [Google Scholar] [CrossRef]
- Wilson, R.P.; Poe, W.E.; Robinson, E.H. Leucine, isoleucine, valine and histidine requirements of fingerling channel catfish. J. Nutr. 1980, 110, 627–633. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, X.; Wu, X.; Gao, Y.; Li, X.; Dong, Y.; Yao, W. Effects of dietary leucine levels on growth, feed utilization, neuro-endocrine growth axis and TOR-related signaling genes expression of juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Aquaculture 2019, 504, 172–181. [Google Scholar] [CrossRef]
- Ren, M.; Habte-Tsion, H.; Liu, B.; Miao, L.; Ge, X.; Xie, J.; Liang, H.; Zhou, Q.; Pan, L. Dietary leucine level affects growth performance, whole body composition, plasma parameters and relative expression of TOR and TNF-ɑ in juvenile blunt snout bream, Megalobrama amblycephala. Aquaculture 2015, 448, 162–168. [Google Scholar] [CrossRef]
- Zehra, S.; Khan, M.A. Dietary leucine requirement of fingerling Catla catla (Hamilton) based on growth, feed conversion ratio, RNA/DNA ratio, leucine gain, blood indices and carcass composition. Aquacult. Int. 2015, 23, 577–595. [Google Scholar] [CrossRef]
- Farhat; Khan, M.A. Response of fingerling stinging catfish, Heteropneustes fossilis (Bloch) to varying levels of dietary L-leucine in relation to growth, feed conversion, protein utilization, leucine retention and blood parameters. Aquacult. Nutr. 2014, 20, 291–302. [Google Scholar] [CrossRef]
- Jie-Min, D.; Mu-Xue, Y.; Zhen-Yu, S.; Chu-Yi, G.; Si-Qi, Z.; Xiao-Shan, Q. Leucine promotes proliferation and differentiation of primary preterm rat satellite cells in part through mTORC1 signaling pathway. Nutrients 2015, 7, 3387–3400. [Google Scholar]
- Zhang, S.; Chen, X.; Huang, Z.; Chen, D.W.; Yu, B.; Chen, H.; Luo, J.; He, J.; Zheng, P.; Yu, J. Leucine promotes differentiation of porcine myoblasts through the protein kinase B (Akt)/Forkhead box O1 signalling pathway. Brit. J. Nutr. 2018, 119, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Averous, J.; Gabillard, J.C.; Seiliez, I.; Dardevet, D. Leucine limitation regulates myf5 and myoD expression and inhibits myoblast differentiation. Exp. Cell Res. 2012, 318, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Young, V.R. Regulation of protein synthesis and skeletal muscle growth. J. Anim. Sci. 1974, 38, 1054–1070. [Google Scholar] [CrossRef]
- Adams, G.R.; Haddad, F. The relationships among IGF-1, DNA content, and protein accumulation during skeletal muscle hypertrophy. J. Appl. Physiol. 1996, 81, 2509–2516. [Google Scholar] [CrossRef]
- Salto, R.; Vílchez, J.D.; Cabrera, E.; Guinovart, J.J.; Girón, M.D. Activation of ERK by sodium tungstate induces protein synthesis and prevents protein degradation in rat L6 myotubes. FEBS. Lett. 2014, 588, 2246–2254. [Google Scholar] [CrossRef]
- Suryawan, A.; Davis, T.A. Regulation of protein degradation pathways by amino acids and insulin in skeletal muscle of neonatal pigs. J. Anim. Sci. Biotechnol. 2014, 5, 8. [Google Scholar] [CrossRef]
- Keller, J.; Couturier, A.; Haferkamp, M.; Most, E.; Eder, K. Supplementation of carnitine leads to an activation of the IGF-1/PI3K/Akt signalling pathway and down regulates the E3 ligase MuRF1 in skeletal muscle of rats. Nutr. Metab. 2013, 10, 28. [Google Scholar] [CrossRef]
- Codina, M.; García De La Serrana, D.; Sánchez-Gurmaches, J.; Montserrat, N.; Chistyakova, O.; Navarro, I.; Gutiérrez, J. Metabolic and mitogenic effects of IGF-II in rainbow trout (Oncorhynchus mykiss) myocytes in culture and the role of IGF-II in the PI3K/Akt and MAPK signalling pathways. Gen. Comp. Endocr. 2008, 157, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, E.N.; Björnsson, B.T.; Valdés, J.A.; Einarsdottir, I.E.; Lorca, B.; Alvarez, M.; Molina, A. IGF-I/PI3K/Akt and IGF-I/MAPK/ERK pathways in vivo in skeletal muscle are regulated by nutrition and contribute to somatic growth in the fine flounder. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1532–R1542. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Wei, D.; Yin, P.; Zheng, L.; Guo, T.; Liu, Y.; Tian, L.; Niu, J. Dietary replacement of fish-meal impaired protein synthesis and immune response of juvenile Pacific white shrimp, Litopenaeus vannamei at low salinity. Comp. Biochem. Physiol. B Biochem. Molec. Biol. 2019, 228, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lu, S.; Wu, M.; Yao, W.; Jin, Z.; Wu, X. Effects of dietary protein levels on growth, feed utilization and expression of growth related genes of juvenile giant grouper (Epinephelus lanceolatus). Aquaculture 2019, 504, 369–374. [Google Scholar] [CrossRef]
- Suryawan, A.; Davis, T.A. Regulation of protein synthesis by amino acids in muscle of neonates. Front. Biosci. 2011, 16, 1445–1460. [Google Scholar] [CrossRef]
- Proud, C.G. Amino acids and mTOR signalling in anabolic function. Biochem. Soc. Trans. 2007, 35, 1187–1190. [Google Scholar] [CrossRef]
- Navé, B.T.; Ouwens, D.M.; Withers, D.J.; Alessi, D.R.; Shepherd, P.R. Mammalian target of rapamycin is a direct target for protein kinase B: Identification of a convergence point for opposing effects of insulin and amino-acid deficiency on protein translation. Biochem. J. 1999, 344, 427–431. [Google Scholar] [CrossRef]
- Cleveland, B.M.; Kenney, P.B.; Manor, M.L.; Weber, G.M. Effects of feeding level and sexual maturation on carcass and fillet characteristics and indices of protein degradation in rainbow trout (Oncorhynchus mykiss). Aquaculture 2012, 338–341, 228–236. [Google Scholar] [CrossRef]
- Paul, P.K.; Kumar, A. TRAF6 coordinates the activation of autophagy and ubiquitin-proteasome systems in atrophying skeletal muscle. Autophagy 2011, 7, 555–556. [Google Scholar] [CrossRef]
- Ge, X.; Zhang, Y.; Jiang, H. Signaling pathways mediating the effects of insulin-like growth factor-I in bovine muscle satellite cells. Mol. Cell. Endocrinol. 2013, 372, 23–29. [Google Scholar] [CrossRef]
- White, J.P.; Gao, S.; Puppa, M.J.; Sato, S.; Welle, S.L.; Carson, J.A. Testosterone regulation of Akt/mTORC1/FoxO3a signaling in skeletal muscle. Mol. Cell. Endocrinol. 2013, 365, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef]
- Kops, G.J.P.L.; Ruiter, N.D.D.; De Vries-Smits, A.M.M.; Powell, D.R.; Bos, J.L.; Burgering, B.M.T. Direct control of the forkhead transcription factor AFX by protein kinase B. Nature 1999, 398, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Glass, D.J. Molecular mechanisms modulating muscle mass. Trends Mol. Med. 2003, 9, 344–350. [Google Scholar] [CrossRef]
- Palma, L.D.; Marinelli, M.; Pavan, M.; Orazi, A. Ubiquitin ligases MuRF1 and MAFbx in human skeletal muscle atrophy. Joint Bone Spine 2008, 75, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Cruz, B.; Oliveira, A.; Gomes-Marcondes, M.C.C. L-leucine dietary supplementation modulates muscle protein degradation and increases pro-inflammatory cytokines in tumour-bearing rats. Cytokine 2017, 96, 253–260. [Google Scholar] [CrossRef]
- Cleveland, B.M.; Weber, G.M. Effects of insulin-like growth factor-I, insulin, and leucine on protein turnover and ubiquitin ligase expression in rainbow trout primary myocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R341–R350. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, X.; Xu, S.; Xie, J.; Xiang, K.; Feng, L.; Liu, Y.; Jiang, W.; Wu, P.; Zhao, J.; et al. Dietary tryptophan affects growth performance, digestive and absorptive enzyme activities, intestinal antioxidant capacity, and appetite and GH–IGF axis-related gene expression of hybrid catfish (Pelteobagrus vachelli♀ × Leiocassis longirostris♂). Fish Physiol. Biochem. 2019, 45, 1627–1647. [Google Scholar] [CrossRef]
- Jiao, S.; Tan, X.; Sui, Y.; You, F. Muscle fibre type composition in the lateral muscle of olive flounder Paralichthys olivaceus. Acta Histochem. 2019, 121, 1–6. [Google Scholar] [CrossRef]
- Jiang, W.; Liu, Y.; Jiang, J.; Wu, P.; Feng, L.; Zhou, X. Copper exposure induces toxicity to the antioxidant system via the destruction of Nrf2/ARE signaling and caspase-3-regulated DNA damage in fish muscle: Amelioration by myo-inositol. Aquat. Toxicol. 2015, 159, 245–255. [Google Scholar] [CrossRef]
- Skiba-Cassy, S.; Lansard, M.; Panserat, S.; Médale, F. Rainbow trout genetically selected for greater muscle fat content display increased activation of liver TOR signaling and lipogenic gene expression. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R1421–R1429. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Han, J.; Xue, M.; Yu, H.; Huang, H.; Wu, X.; Zheng, Y.; Qin, Y.; Liang, X. Growth and feed intake regulation responses to anorexia, adaptation and fasting in Japanese seabss, Lateolabrax japonicas when fishmeal is totally replaced by plant protein. Aquaculture 2019, 498, 528–538. [Google Scholar] [CrossRef]
- Jiang, W.; Wen, H.; Liu, Y.; Jiang, J.; Wu, P.; Zhao, J.; Kuang, S.; Tang, L.; Tang, W.; Zhang, Y.; et al. Enhanced muscle nutrient content and flesh quality, resulting from tryptophan, is associated with anti-oxidative damage referred to the Nrf2 and TOR signalling factors in young grass carp (Ctenopharyngodon idella): Avoid tryptophan deficiency or excess. Food Chem. 2016, 199, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Khan, M.A. Dietary branched-chain amino acid valine, isoleucine and leucine requirements of fingerling Indian major carp, Cirrhinus mrigala (Hamilton). Brit. J. Nutr. 2006, 96, 450–460. [Google Scholar]
- Abidi, S.F.; Khan, M.A. Dietary leucine requirement of fingerling Indian major carp, Labeo rohita (Hamilton). Aquac. Res. 2007, 38, 478–486. [Google Scholar] [CrossRef]
- Kim, S.S.; Lee, K.J. Comparison of leucine requirement in olive flounder (Paralichthys olivaceus) by free or synthetic dipeptide forms of leucine. Anim. Feed Sci. Tech. 2013, 183, 195–201. [Google Scholar] [CrossRef]
- Yamamoto, T.; Shima, T.; Furuita, H. Antagonistic effects of branched-chain amino acids induced by excess protein-bound leucine in diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 2004, 232, 539–550. [Google Scholar] [CrossRef]
- Hughes, S.G.; Rumsey, G.L.; Nesheim, M.C. Dietary requirements for essential branched-chain amino acids by lake trout. Trans. Am. Fish. Soc. 1983, 112, 812–817. [Google Scholar] [CrossRef]
- Greek Walker, M.; Pull, G. A survey of red and white muscle in marine fish. J. Fish Biol. 2006, 7, 295–300. [Google Scholar] [CrossRef]
- Xu, Y.; Tan, Q.; Kong, F.; Yu, H.; Zhu, Y.; Yao, J.; Abouel Azm, F.R. Fish growth in response to different feeding regimes and the related molecular mechanism on the changes in skeletal muscle growth in grass carp (Ctenopharyngodon idellus). Aquaculture 2019, 512, 734295. [Google Scholar] [CrossRef]
- Martín-Pérez, M.; Fernandez-Borras, J.; Ibarz, A.; Felip, O.; Fontanillas, R.; Gutierrez, J.; Blasco, J. Naturally occurring stable isotopes reflect changes in protein turnover and growth in gilthead sea bream (Sparus aurata) juveniles under different dietary protein levels. J. Agric. Food Chem. 2013, 61, 8924–8933. [Google Scholar] [CrossRef] [PubMed]
- Tirapegui, J.; Ribeiro, S.M.L.; Pires, I.S.D.O.; Rogero, M.M. Effects of two different levels of dietary protein on body composition and protein nutritional status of growing rats. Nutrients 2012, 4, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- Areta, J.L.; Hawley, J.A.; Ye, J.; Chan, M.H.S.; Coffey, V.G. Increasing leucine concentration stimulates mechanistic target of rapamycin signaling and cell growth in C2C12 skeletal muscle cells. Nutr. Res. 2014, 34, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Rius-Francino, M.; Acerete, L.; Jiménez-Amilburu, V.; Capilla, E.; Navarro, I.; Gutiérrez, J. Differential effects on proliferation of GH and IGFs in sea bream (Sparus aurata) cultured myocytes. Gen. Comp. Endocr. 2011, 172, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Azizi, S.; Nematollahi, M.A.; Mojazi Amiri, B.; Vélez, E.J.; Lutfi, E.; Navarro, I.; Capilla, E.; Gutiérrez, J. Lysine and leucine deficiencies affect myocytes development and IGF signaling in gilthead sea bream (Sparus aurata). PLoS ONE 2016, 11, e0147618. [Google Scholar] [CrossRef]
- Church, D.D.; Schwarz, N.A.; Spillane, M.B.; McKinley-Barnard, S.K.; Andre, T.L.; Ramirez, A.J.; Willoughby, D.S. L-leucine increases skeletal muscle IGF-1 but does not differentially increase Akt/mTORC1 signaling and serum IGF-1 compared to ursolic acid in response to resistance exercise in resistance-trained men. J. Am. Coll. Nutr. 2016, 35, 627–638. [Google Scholar] [CrossRef]
- Johnston, I.A. Muscle development and growth: Potential implications for flesh quality in fish. Aquaculture 1999, 177, 99–115. [Google Scholar] [CrossRef]
- Coolican, S.A.; Samuel, D.S.; Ewton, D.Z.; Mcwade, F.J.; Florini, J.R. The mitogenic and myogenic actions of insulin-like growth factors utilize distinct signaling pathways. J. Biol. Chem. 1997, 272, 6653–6662. [Google Scholar] [CrossRef]
- Montserrat, N.; Capilla, E.; Navarro, I.; Gutiérrez, J. Metabolic Effects of Insulin and IGFs on gilthead sea bream (Sparus aurata) muscle cells. Front. Endocrinol. 2012, 3, 55. [Google Scholar] [CrossRef]
- Rescan, P.Y. Muscle growth patterns and regulation during fish ontogeny. Gen. Comp. Endocr. 2005, 142, 111–116. [Google Scholar] [CrossRef]
- Johnson, S.E.; Allen, R.E. Proliferating cell nuclear antigen (PCNA) is expressed in activated rat skeletal muscle satellite cells. J. Cell Physiol. 1993, 154, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Rescan, P.Y. Regulation and functions of myogenic regulatory factors in lower vertebrates. Compar. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 130, 1–12. [Google Scholar] [CrossRef]
- Cornelison, D.D.W.; Olwin, B.B.; Rudnicki, M.A.; Wold, B.J. MyoD−/− satellite cells in single-fiber culture are differentiation defective and MRF4 deficient. Dev. Biol. 2000, 224, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Biga, P.R.; Goetz, F.W. Zebrafish and giant danio as models for muscle growth: Determinate vs. indeterminate growth as determined by morphometric analysis. Am. J. Physiol. Regul. Integr. Compar. Physiol. 2006, 291, R1327–R1337. [Google Scholar] [CrossRef]
- Chen, X.; Huang, Z.; Chen, D.; Yang, T.; Liu, G. MicroRNA-27a is induced by leucine and contributes to leucine-induced proliferation promotion in C2C12 cells. Int. J. Mol. Sci. 2013, 14, 14076–14084. [Google Scholar] [CrossRef] [PubMed]
- Rebhan, Y.; Funkenstein, B. Inhibition of fish myostatin activity by recombinant fish follistatin and myostatin prodomain: Potential implications for enhancing muscle growth in farmed fish. Aquaculture 2008, 284, 231–238. [Google Scholar] [CrossRef]
- Mobley, C.B.; Fox, C.D.; Ferguson, B.S.; Amin, R.H.; Dalbo, V.J.; Baier, S.; Rathmacher, J.A.; Wilson, J.M.; Roberts, M.D. L-leucine, beta-hydroxy-beta-methylbutyric acid (HMB) and creatine monohydrate prevent myostatin-induced Akirin-1/Mighty mRNA down-regulation and myotube atrophy. J. Int. Soc. Sports Nutr. 2014, 11, 38. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, X.; Yu, B.; He, J.; Chen, D.W. MicroRNA-27a promotes myoblast proliferation by targeting myostatin. Biochem. Bioph. Res. Commun. 2012, 423, 265–269. [Google Scholar] [CrossRef]
- Michelato, M.; Zaminhan, M.; Boscolo, W.R.; Nogaroto, V.; Vicari, M.; Artoni, R.F.; Furuya, V.R.B.; Furuya, W.M. Dietary histidine requirement of Nile tilapia juveniles based on growth performance, expression of muscle-growth-related genes and haematological responses. Aquaculture 2017, 467, 63–70. [Google Scholar] [CrossRef]
- Alami-Durante, H.; Bazin, D.; Cluzeaud, M.; Fontagné-Dicharry, S.; Kaushik, S.; Geurden, I. Effect of dietary methionine level on muscle growth mechanisms in juvenile rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 483, 273–285. [Google Scholar] [CrossRef]
- Childress, C.J.; Fuller, S.A.; Rawles, S.D.; Beck, B.H.; Gaylord, T.G.; Barrows, F.T.; McEntire, M.E. Lysine supplementation of commercial fishmeal-free diet in hybrid striped bass Morone chrysops x M. saxatilis affects expression of growth-related genes. Aquacult. Nutr. 2016, 22, 738–744. [Google Scholar] [CrossRef]
- Houlihan, D.F.; Hall, S.J.; Gray, C. Effects of ration on protein turnover in cod. Aquaculture 1989, 79, 103–110. [Google Scholar] [CrossRef]
- Cantley, L.C. The phosphoinositide 3-kinase pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef] [PubMed]
- Engelman, J.A.; Luo, J.; Cantley, L.C. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat. Rev. Genet. 2006, 7, 606–619. [Google Scholar] [CrossRef]
- Xu, Y.; Li, N.; Xiang, R.; Sun, P. Emerging roles of the p38 MAPK and PI3K/AKT/mTOR pathways in oncogene-induced senescence. Trends Biochem. Sci. 2014, 39, 268–276. [Google Scholar] [CrossRef]
- Montserrat, N.; Sánchez-Gurmaches, J.; García De La Serrana, D.; Navarro, M.I.; Gutiérrez, J. IGF-I binding and receptor signal transduction in primary cell culture of muscle cells of gilthead sea bream: Changes throughout in vitro development. Cell Tissue Res. 2007, 330, 503–513. [Google Scholar] [CrossRef]
- Seiliez, I.; Gabillard, J.; Skiba-Cassy, S.; Garcia-Serrana, D.; Gutiérrez, J.; Kaushik, S.; Panserat, S.; Tesseraud, S. An in vivo and in vitro assessment of TOR signaling cascade in rainbow trout (Oncorhynchus mykiss). Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R329–R335. [Google Scholar] [CrossRef]
- Lansard, M.; Panserat, S.; Plagnes-Juan, E.; Seiliez, I.; Skiba-Cassy, S. Integration of insulin and amino acid signals that regulate hepatic metabolism-related gene expression in rainbow trout: Role of TOR. Amino Acids 2010, 39, 801–810. [Google Scholar] [CrossRef]
- Kimball, S.R. Regulation of global and specific mRNA translation by amino acids. J. Nutr. 2002, 132, 883–886. [Google Scholar] [CrossRef]
- Hara, K.; Yonezawa, K.; Weng, Q.P.; Kozlowski, M.T.; Belham, C.; Avruch, J. Amino acid sufficiency and mTOR regulate p70 S6 kinase and eIF-4E BP1 through a common effector mechanism. J. Biol. Chem. 1998, 273, 14484–14494. [Google Scholar] [CrossRef]
- Kimball, S.R.; Shantz, L.M.; Horetsky, R.L.; Jefferson, L.S. Leucine regulates translation of specific mRNAs in L6 myoblasts through mTOR-mediated changes in availability of eIF4E and phosphorylation of ribosomal protein S6. J. Biol. Chem. 1999, 274, 11647–11652. [Google Scholar] [CrossRef] [PubMed]
- Sekuli, A.; Hudson, C.C.; Homme, J.L.; Yin, P.; Otterness, D.M.; Karnitz, L.M.; Abraham, R.T. A direct linkage between the phosphoinositide 3-kinase-AKT signaling pathway and the mammalian target of rapamycin in mitogen-stimulated and transformed cells. Cancer Res. 2000, 60, 3504–3513. [Google Scholar]
- Lang, C.H.; Frost, R.A. Endotoxin disrupts the leucine-signaling pathway involving phosphorylation of mTOR, 4E-BP1, and S6K1 in skeletal muscle. J. Cell. Physiol. 2005, 203, 144–155. [Google Scholar] [CrossRef]
- Zhao, J.; Brault, J.; Schild, A.; Cao, P.; Sandri, M.; Schiaffino, S.; Lecker, S.; Goldberg, A. FoxO3 coordinately activates protein degradation by the autophagic/lysosomal and proteasomal pathways in atrophying muscle cells. Cell Metab. 2008, 6, 472–483. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C.; Latres, E.; Baumhueter, S.; Lai, V.K.M.; Nunez, L.; Clarke, B.A.; Poueymirou, W.T.; Panaro, F.J.; Na, E.; Dharmarajan, K.; et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 2001, 294, 1704–1708. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Bai, K.; He, J.; Su, W.; Dong, L.; Zhang, L.; Wang, T. Leucine improves growth performance of intrauterine growth retardation piglets by modifying gene and protein expression related to protein synthesis. Nutrition 2016, 32, 114–121. [Google Scholar] [CrossRef]
- Borgenvik, M.; Apró, W.; Blomstrand, E. Intake of branched-chain amino acids influences the levels of MAFbx mRNA and MuRF-1 total protein in resting and exercising human muscle. Am. J. Physiol. Endoc. Metab. 2011, 302, E510–E521. [Google Scholar] [CrossRef]
- Buse, M.G.; Reid, S.S. Leucine. A possible regulator of protein turnover in muscle. J. Clin. Invest. 1975, 56, 1250–1261. [Google Scholar] [CrossRef]
- Sacheck, J.M.; Hyatt, J.K.; Raffaello, A.; Jagoe, R.T.; Roy, R.R.; Edgerton, V.R.; Lecker, S.H.; Goldberg, A.L. Rapid disuse and denervation atrophy involve transcriptional changes similar to those of muscle wasting during systemic diseases. FASEB. J. 2006, 21, 140–155. [Google Scholar] [CrossRef]
- Sacheck, J.M.; Ohtsuka, A.; McLary, S.C.; Goldberg, A.L. IGF-I stimulates muscle growth by suppressing protein breakdown and expression of atrophy-related ubiquitin ligases, atrogin-1 and MuRF1. Am. J. Physiol. Endoc. Metab. 2004, 287, E591–E601. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Nutrients contenta | ||
|---|---|---|---|
| Fish meal | 170.0 | Crude protein | 384.7 |
| Casein | 10.0 | Crude fat | 71.0 |
| Gelatin | 10.0 | Available phosphorus | 14.8 |
| Amino acid premixb | 170.0 | ω-3 | 15.2 |
| Leucine-glycine premixc | 120.0 | ω-6 | 13.9 |
| α-starch | 210.0 | ||
| Corn starch | 168.3 | ||
| Fish oil | 30.0 | ||
| Soybean oil | 26.0 | ||
| Vitamin premixd | 10.0 | ||
| Mineral element premixe | 20.0 | ||
| Ca(H2PO4)2 | 40.0 | ||
| Choline chloride (50%) | 10.0 | ||
| Ethoxyquin (30%) | 0.5 | ||
| Cellulose | 5.0 | ||
| XanthophyII | 0.2 |
| Amino Acid | Dietary Leu Level (g kg−1 Diet) | 384.7 g kg−1 Muscle Protein | ||||||
|---|---|---|---|---|---|---|---|---|
| 10 | 15 | 20 | 25 | 30 | 35 | 40 | ||
| Essential amino acid | ||||||||
| Threonine | 1.93 | 1.89 | 1.95 | 1.94 | 1.90 | 1.87 | 1.93 | 1.94 |
| Valine | 1.34 | 1.38 | 1.41 | 1.36 | 1.39 | 1.42 | 1.43 | 1.40 |
| Methionine | 1.26 | 1.25 | 1.19 | 1.23 | 1.23 | 1.25 | 1.22 | 1.21 |
| Isoleucine | 1.17 | 1.14 | 1.12 | 1.16 | 1.20 | 1.18 | 1.17 | 1.18 |
| Leucine | 1.11 | 1.48 | 2.02 | 2.52 | 3.04 | 3.57 | 4.02 | 2.53 |
| Phenylalanine | 1.81 | 1.79 | 1.80 | 1.82 | 1.75 | 1.83 | 1.76 | 1.77 |
| Histidine | 0.84 | 0.76 | 0.80 | 0.82 | 0.79 | 0.81 | 0.85 | 0.83 |
| Lysine | 2.61 | 2.59 | 2.62 | 2.57 | 2.65 | 2.58 | 2.55 | 2.62 |
| Arginine | 2.57 | 2.55 | 2.47 | 2.60 | 2.58 | 2.61 | 2.49 | 2.53 |
| Nonessential amino acid | ||||||||
| Aspartic acid | 4.41 | 4.50 | 4.62 | 4.55 | 4.48 | 4.68 | 4.60 | 4.56 |
| Serine | 1.91 | 1.93 | 1.87 | 1.95 | 2.01 | 1.94 | 2.05 | 1.97 |
| Glutamic acid | 6.82 | 6.77 | 6.68 | 6.71 | 6.75 | 6.69 | 6.66 | 6.62 |
| Glycine | 2.22 | 2.28 | 2.26 | 2.30 | 2.32 | 2.29 | 2.25 | 2.21 |
| Alanine | 2.73 | 2.85 | 2.80 | 2.78 | 2.71 | 2.90 | 2.87 | 2.82 |
| Cystine | 0.21 | 0.20 | 0.19 | 0.25 | 0.18 | 0.21 | 0.17 | 0.23 |
| Tyrosine | 1.51 | 1.51 | 1.47 | 1.45 | 1.55 | 1.56 | 1.49 | 1.50 |
| Proline | 1.44 | 1.48 | 1.51 | 1.57 | 1.48 | 1.50 | 1.53 | 1.46 |
| Name | Sequence (5′–3′) | OAT (°C) | Accession Number |
|---|---|---|---|
| Myf5-QF | CTCCAGTCCTTCATCATCCACC | 64.9 | MK253547 |
| Myf5-QR | CACTCGCACTCTGACCTTCGT | ||
| MyoD-QF | CCTAATCAGAGGCTTCCCA | 55.5 | HM363525 |
| MyoD-QR | TCACCGCTGTATTGTTCCA | ||
| MyoG-QF | TACTTTTTCCCCGAACAGC | 57.6 | HQ246723 |
| MyoG-QR | TCCAGTCCTACATTGCCAGA | ||
| MRF4-QF | CAGACTGTCAGAGGACGGGG | 52.8 | MK281342 |
| MRF4-QR | CAGCCTTCTCTTTGGTGGGA | ||
| MSTN-QF | ACGCCACTACCGAGACCG | 64.6 | DQ767967 |
| MSTN-QR | CTCAATACCCCAGTTTGTTTCC | ||
| PCNA-QF | GTTGATGGACTTGGATGTGGA | 60.1 | MK281343 |
| PCNA-QR | CGTTGCTGGTTTGGGAGA | ||
| MyHC-QF | GCAATGAAGGAGAACTATG | 60.0 | MK440319 |
| MyHC-QR | TCACACTTTCCTCAGCGT | ||
| IGF-I-QF | ATCTGGGTAATGTGACTGCCGA | 56.8 | KX434878 |
| IGF-I-QR | TTCATCATCTCCGCCCTTGC | ||
| IGF-IR-QF | ACACCGATGAGGGAAACTGG | 56.6 | MG773202 |
| IGF-IR-QR | GTGGATGAAGGACGGGAACA | ||
| PI3K-QF | GTGAATGGGAAAGACGCT | 62.6 | MG773208 |
| PI3K-QR | GCACACAGGACTCCAGATGA | ||
| AKT-QF | TCTACCCTTTACACCTGCTGAC | 61.7 | KX131157.1 |
| AKT-QR | GATGGCTGGGATTGCTTTC | ||
| TOR-QF | GACAAACGGAGGAAGGAGG | 58.2 | MG773199 |
| TOR-QR | TCATCAGGAAAGAAGAGGGACT | ||
| 4E-BP1-QF | ACGCCACCCAGTTGCCTA | 62.6 | MG773207 |
| 4E-BP1-QR | GGATGCTTTTGCTGCCGAC | ||
| S6K1-QF | GCAAACTGAATCTCCCACCC | 61.7 | MG773195 |
| S6K1-QR | AGGCTTGAAAGGCGGCTC | ||
| MURF-1-QF | CCGTTTTGAGGTGGTGCT | 53.6 | MK756118 |
| MURF-1- QR | TGTTCTCCAGTTGTTGCTTGTA | ||
| MAFBX-QF | AACCTCTGTCACTACCACTTCACT | 54.8 | MK812970 |
| MAFBX- QR | GGTCGCTGTACTGCTCTTTATG | ||
| FOXO3a-QF | GACTTCCGCTCTCGCACTAA | 60.5 | MK562423 |
| FOXO3a-QR | ATCATCAGCAACCTCATCCACT | ||
| β-actin-QF | CCTAAAGCCAACAGGGAAAA | 59 | EU161066 |
| β-actin-QR | ATGGGGCAGAGCATAACC | ||
| 18S-QF | CCTGAGAAACGGCTACCACATCC | 57.1 | KP938527 |
| 18S-QR | AGCAACTTTAATATACGCTATTGGAG |
| Leu | 10.0 | 15.0 | 20.0 | 25.0 | 30.0 | 35.0 | 40.0 |
|---|---|---|---|---|---|---|---|
| IBW | 23.21 ± 0.13 | 22.98 ± 0.14 | 22.90 ± 0.20 | 23.19 ± 0.27 | 23.32 ± 0.24 | 23.30 ± 0.25 | 23.34 ± 0.19 |
| Survival | 96.67 ± 1.93 | 96.67 ± 0.00 | 95.56 ± 2.94 | 98.89 ± 1.11 | 95.56 ± 1.11 | 97.78 ± 2.22 | 94.44 ± 2.93 |
| FBW | 43.73 ± 0.70a | 44.73 ± 0.13a | 48.22 ± 1.32a | 54.55 ± 0.87b | 49.48 ± 1.58ab | 47.82 ± 2.22a | 45.53 ± 1.17a |
| PWG | 88.35 ± 1.81a | 94.70 ± 1.95ab | 111.20 ± 5.51ab | 137.89 ± 3.56c | 117.91 ± 11.96bc | 112.42 ± 8.26ab | 94.94 ± 2.99ab |
| SGR | 1.13 ± 0.02a | 1.19 ± 0.02a | 1.33 ± 0.05ab | 1.55 ± 0.03b | 1.34 ± 0.07ab | 1.27 ± 0.08a | 1.19 ± 0.03a |
| FI | 37.57 ± 1.19bc | 38.60 ± 0.54bc | 42.04 ± 1.84c | 43.30 ± 3.74c | 38.00 ± 0.57bc | 34.25 ± 1.02ab | 31.66 ± 1.74a |
| FE | 53.05 ± 1.42a | 56.87 ± 2.79a | 59.08 ± 1.15a | 74.87 ± 2.67b | 71.85 ± 4.15b | 69.01 ± 4.27b | 67.34 ± 1.48b |
| PER | 2.06 ± 0.03a | 2.09 ± 0.07a | 2.41 ± 0.03ab | 3.11 ± 0.37b | 2.53 ± 0.21ab | 2.52 ± 0.08ab | 2.58 ± 0.01ab |
| Regressions | |||||||
| YPWG = -0.1535X2 + 8.1194X + 16.5380 | X = 26.45 | R2 = 0.7833 | P = 0.047 | ||||
| YSGR = -0.0012X2 + 0.0647X + 0.5707 | X = 26.96 | R2 = 0.7366 | P = 0.075 | ||||
| YFI = -0.03199X2 + 1.382X + 26.57 | X = 21.60 | R2 = 0.8765 | P = 0.015 | ||||
| YFE = -0.043X2 + 2.7213X + 27.73 | X = 31.65 | R2 = 0.7959 | P = 0.042 | ||||
| YPER = -0.0019X2 + 0.1148X + 1.0029 | X = 30.21 | R2 = 0.5735 | P = 0.182 | ||||
| Leu | 10.0 | 15.0 | 20.0 | 25.0 | 30.0 | 35.0 | 40.0 |
|---|---|---|---|---|---|---|---|
| DI | 28.85 ± 0.42a | 29.36 ± 0.92a | 34.35 ± 2.92ab | 38.48 ± 1.85b | 33.31 ± 1.64ab | 29.74 ± 0.31a | 28.63 ± 0.77a |
| DE | 64.00 ± 1.79a | 79.4 ± 3.97b | 88.4 ± 4.53bc | 103.5 ± 7.58c | 90.6 ± 2.5bc | 95.4 ± 1.81bc | 87 ± 6.82bc |
| Independent Parameters | Dependent Parameters | Correlation Coefficients | P |
|---|---|---|---|
| PWG | FE | 0.796 | 0.032 |
| PER | 0.872 | 0.011 | |
| IGF-I mRNA | Muscle fiber diameter | 0.724 | 0.066 |
| Myf5 mRNA | 0.837 | 0.019 | |
| MyoD mRNA | 0.820 | 0.024 | |
| MyoG mRNA | 0.977 | 0.000 | |
| MRF4 mRNA | 0.768 | 0.044 | |
| PI3K mRNA | 0.637 | 0.124 | |
| AKT mRNA | 0.855 | 0.014 | |
| TOR mRNA | 0.897 | 0.006 | |
| 4E-BP mRNA | -0.703 | 0.078 | |
| S6K1 mRNA | 0.639 | 0.122 | |
| FOXO3a mRNA | -0.809 | 0.028 | |
| MURF-1 mRNA | -0.923 | 0.003 | |
| MAFBX mRNA | -0.883 | 0.008 | |
| Protein content | IGF-I mRNA | 0.912 | 0.004 |
| Myf5 mRNA | 0.801 | 0.031 | |
| MyoD mRNA | 0.803 | 0.030 | |
| MyoG mRNA | 0.902 | 0.005 | |
| MRF4 mRNA | 0.670 | 0.099 | |
| MSTN mRNA | -0.736 | 0.059 | |
| PI3K mRNA | 0.782 | 0.038 | |
| AKT mRNA | 0.822 | 0.023 | |
| TOR mRNA | 0.987 | 0.000 | |
| 4E-BP mRNA | -0.730 | 0.062 | |
| S6K1 mRNA | 0.791 | 0.034 | |
| FOXO3a mRNA | -0.649 | 0.115 | |
| MURF-1 mRNA | -0.838 | 0.019 | |
| MAFBX mRNA | -0.808 | 0.028 | |
| TOR mRNA | Myf5 mRNA | 0.840 | 0.018 |
| MyoD mRNA | 0.876 | 0.010 | |
| MyoG mRNA | 0.895 | 0.006 | |
| MRF4 mRNA | 0.713 | 0.072 | |
| MSTN mRNA | -0.814 | 0.026 | |
| PCNA mRNA | 0.672 | 0.098 | |
| 4E-BP mRNA | Myf5 mRNA | -0.806 | 0.029 |
| MyoD mRNA | -0.963 | 0.000 | |
| MyoG mRNA | 0.697 | 0.082 | |
| MRF4 mRNA | -0.647 | 0.116 | |
| MSTN mRNA | 0.887 | 0.008 | |
| PCNA mRNA | -0.851 | 0.015 | |
| MyHC mRNA | 0.801 | 0.030 | |
| S6K1 mRNA | Myf5 mRNA | 0.677 | 0.095 |
| MyoD mRNA | 0.760 | 0.047 | |
| MyoG mRNA | 0.663 | 0.105 | |
| MRF4 mRNA | 0.698 | 0.081 | |
| MSTN mRNA | -0.839 | 0.018 | |
| PCNA mRNA | 0.706 | 0.076 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Li, J.-Y.; Jiang, Q.; Zhou, X.-Q.; Feng, L.; Liu, Y.; Jiang, W.-D.; Wu, P.; Zhou, J.; Zhao, J.; et al. Leucine Improved Growth Performance, Muscle Growth, and Muscle Protein Deposition Through AKT/TOR and AKT/FOXO3a Signaling Pathways in Hybrid Catfish Pelteobagrus vachelli × Leiocassis longirostris. Cells 2020, 9, 327. https://doi.org/10.3390/cells9020327
Zhao Y, Li J-Y, Jiang Q, Zhou X-Q, Feng L, Liu Y, Jiang W-D, Wu P, Zhou J, Zhao J, et al. Leucine Improved Growth Performance, Muscle Growth, and Muscle Protein Deposition Through AKT/TOR and AKT/FOXO3a Signaling Pathways in Hybrid Catfish Pelteobagrus vachelli × Leiocassis longirostris. Cells. 2020; 9(2):327. https://doi.org/10.3390/cells9020327
Chicago/Turabian StyleZhao, Ye, Jin-Yang Li, Qin Jiang, Xiao-Qiu Zhou, Lin Feng, Yang Liu, Wei-Dan Jiang, Pei Wu, Jian Zhou, Juan Zhao, and et al. 2020. "Leucine Improved Growth Performance, Muscle Growth, and Muscle Protein Deposition Through AKT/TOR and AKT/FOXO3a Signaling Pathways in Hybrid Catfish Pelteobagrus vachelli × Leiocassis longirostris" Cells 9, no. 2: 327. https://doi.org/10.3390/cells9020327
APA StyleZhao, Y., Li, J.-Y., Jiang, Q., Zhou, X.-Q., Feng, L., Liu, Y., Jiang, W.-D., Wu, P., Zhou, J., Zhao, J., & Jiang, J. (2020). Leucine Improved Growth Performance, Muscle Growth, and Muscle Protein Deposition Through AKT/TOR and AKT/FOXO3a Signaling Pathways in Hybrid Catfish Pelteobagrus vachelli × Leiocassis longirostris. Cells, 9(2), 327. https://doi.org/10.3390/cells9020327