Understanding Pseudomonas aeruginosa–Host Interactions: The Ongoing Quest for an Efficacious Vaccine


Abstract
1. Introduction
2. P. aeruginosa Virulence Factors and Adaptation to the Lung Environment
3. Host Immune Response against P. aeruginosa
3.1. Recognition
3.2. The Innate Immune Response
3.3. The Adaptive Response
3.4. The Importance of Novel Animal Models
4. Vaccination against P. aeruginosa: An Overview of the Last 50 Years
4.1. Lipopolysaccharide
4.2. Alginate
4.3. Flagellar Antigens
4.4. Type 4 Pili
4.5. Outer Membrane Proteins
4.5.1. OprF Major Porin and OprI Lipoprotein
4.5.2. OprL, OprH, OprG and Others
4.6. Type III Secretion System
4.7. Extracellular Products
4.7.1. Exotoxin A
4.7.2. Other Extracellular Products
4.8. Other Bacterial Components
4.9. Whole-Cell Killed and Live-Attenuated P. aeruginosa Vaccines
4.10. DNA Vaccines
5. Adjuvants
5.1. Definition, Function, Classification, and Licensing
5.2. Adjuvants in Vaccines against P. aeruginosa
6. Lessons from P. aeruginosa Vaccines and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Azam, M.W.; Khan, A.U. Updates on the pathogenicity status of Pseudomonas aeruginosa. Drug Discov. Today 2019, 24, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Bianconi, I.; Jeukens, J.; Freschi, L.; Alcalá-Franco, B.; Facchini, M.; Boyle, B.; Molinaro, A.; Kukavica-Ibrulj, I.; Tümmler, B.; Levesque, R.C.; et al. Comparative genomics and biological characterization of sequential Pseudomonas aeruginosa isolates from persistent airways infection. BMC Genom. 2015, 16, 1105. [Google Scholar] [CrossRef] [PubMed]
- Tümmler, B. Emerging therapies against infections with Pseudomonas aeruginosa. F1000Res 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Hamouche, J.E.; Wang, G.; Smith, M.; Yin, C.; Dhand, A.; Dimitrova, N.; Fallon, J.T. Integrated Genome-Wide Analysis of an Isogenic Pair of Pseudomonas aeruginosa Clinical Isolates with Differential Antimicrobial Resistance to Ceftolozane/Tazobactam, Ceftazidime/Avibactam, and Piperacillin/Tazobactam. Int. J. Mol. Sci. 2020, 21, 1026. [Google Scholar] [CrossRef] [PubMed]
- Center for Disease Control and Prevention (USA). Antibiotic Resistance Threats in the United States. 2019. Available online: https://www.cdc.gov/drugresistance/biggest-threats.html (accessed on 10 August 2020).
- Bianconi, I.; Alcalá-Franco, B.; Scarselli, M.; Dalsass, M.; Buccato, S.; Colaprico, A.; Marchi, S.; Masignani, V.; Bragonzi, A. Genome-Based Approach Delivers Vaccine Candidates against Pseudomonas aeruginosa. Front. Immunol. 2018, 9, 3021. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and Treatment of Multidrug-Resistant and Extensively Drug-Resistant Pseudomonas aeruginosa Infections. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef]
- Adlbrecht, C.; Wurm, R.; Depuydt, P.; Spapen, H.; Lorente, J.A.; Staudinger, T.; Creteur, J.; Zauner, C.; Meier-Hellmann, A.; Eller, P.; et al. Efficacy, immunogenicity, and safety of IC43 recombinant Pseudomonas aeruginosa vaccine in mechanically ventilated intensive care patients-a randomized clinical trial. Crit. Care 2020, 24, 74. [Google Scholar] [CrossRef]
- Grimwood, K.; Kyd, J.M.; Owen, S.J.; Massa, H.M.; Cripps, A.W. Vaccination against respiratory Pseudomonas aeruginosa infection. Hum. Vaccines Immunother. 2015, 11, 14–20. [Google Scholar] [CrossRef]
- Marshall, B.; AF, A.F.; Loeffler, D.; Elbert, A.; O’Neil, T.; Rush, T.; Rizvi, S. Patient Registry Annual Data Report; Cystic Fibrosis Foundation: Bethesda, MD, USA, 2018; Volume 2018. [Google Scholar]
- King, J.D.; Kocíncová, D.; Westman, E.L.; Lam, J.S. Lipopolysaccharide biosynthesis in Pseudomonas aeruginosa. Innate Immun. 2009, 15, 261–312. [Google Scholar] [CrossRef]
- Alshalchi, S.A.; Anderson, G.G. Expression of the lipopolysaccharide biosynthesis gene lpxD affects biofilm formation of Pseudomonas aeruginosa. Arch. Microbiol. 2015, 197, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Huszczynski, S.M.; Lam, J.S.; Khursigara, C.M. The role of Pseudomonas aeruginosa lipopolysaccharide in bacterial pathogenesis and physiology. Pathogens 2019, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, S.; Bouffartigues, E.; Bodilis, J.; Maillot, O.; Lesouhaitier, O.; Feuilloley, M.G.J.; Orange, N.; Dufour, A.; Cornelis, P. Structure, function and regulation of Pseudomonas aeruginosa porins. FEMS Microbiol. Rev. 2017, 41, 698–722. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, I.; Parales, R.E.; Krell, T.; Hill, J.E. Pseudomonas chemotaxis. FEMS Microbiol. Rev. 2015, 39, 17–46. [Google Scholar] [CrossRef]
- Haiko, J.; Westerlund-Wikström, B. The role of the bacterial flagellum in adhesion and virulence. Biology 2013, 2, 1242–1267. [Google Scholar] [CrossRef]
- Burrows, L.L. Pseudomonas aeruginosa twitching motility: Type IV pili in action. Annu. Rev. Microbiol. 2012, 66, 493–520. [Google Scholar] [CrossRef]
- Lee, K.; Yoon, S.S. Pseudomonas aeruginosa biofilm, a programmed bacterial life for fitness. J. Microbiol. Biotechnol. 2017, 27, 1053–1064. [Google Scholar] [CrossRef]
- Yan, S.; Wu, G. Can Biofilm Be Reversed through Quorum Sensing in Pseudomonas aeruginosa? Front. Microbiol. 2019, 10, 1582. [Google Scholar] [CrossRef]
- Mann, E.E.; Wozniak, D.J. Pseudomonas biofilm matrix composition and niche biology. FEMS Microbiol. Rev. 2012, 36, 893–916. [Google Scholar] [CrossRef]
- Bleves, S.; Viarre, V.; Salacha, R.; Michel, G.P.; Filloux, A.; Voulhoux, R. Protein secretion systems in Pseudomonas aeruginosa: A wealth of pathogenic weapons. Int. J. Med. Microbiol. 2010, 300, 534–543. [Google Scholar] [CrossRef]
- Pena, R.T.; Blasco, L.; Ambroa, A.; González-Pedrajo, B.; Fernández-García, L.; López, M.; Bleriot, I.; Bou, G.; García-Contreras, R.; Wood, T.K.; et al. Relationship Between Quorum Sensing and Secretion Systems. Front. Microbiol. 2019, 10, 1100. [Google Scholar] [CrossRef] [PubMed]
- Anantharajah, A.; Mingeot-Leclercq, M.P.; Van Bambeke, F. Targeting the Type Three Secretion System in Pseudomonas aeruginosa. Trends Pharmacol. Sci. 2016, 37, 734–749. [Google Scholar] [CrossRef] [PubMed]
- Michalska, M.; Wolf, P. Pseudomonas Exotoxin A: Optimized by evolution for effective killing. Front. Microbiol. 2015, 6, 963. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.; McDermott, C.; Anoopkumar-Dukie, S.; McFarland, A.J.; Forbes, A.; Perkins, A.V.; Davey, A.K.; Chess-Williams, R.; Kiefel, M.J.; Arora, D.; et al. Cellular Effects of Pyocyanin, a Secreted Virulence Factor of Pseudomonas aeruginosa. Toxins 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Strateva, T.; Mitov, I. Contribution of an arsenal of virulence factors to pathogenesis of Pseudomonas aeruginosa infections. Ann. Microbiol. 2011, 61, 717–732. [Google Scholar] [CrossRef]
- Zulianello, L.; Canard, C.; Köhler, T.; Caille, D.; Lacroix, J.S.; Meda, P. Rhamnolipids are virulence factors that promote early infiltration of primary human airway epithelia by Pseudomonas aeruginosa. Infect. Immun. 2006, 74, 3134–3147. [Google Scholar] [CrossRef]
- Halldorsson, S.; Gudjonsson, T.; Gottfredsson, M.; Singh, P.K.; Gudmundsson, G.H.; Baldursson, O. Azithromycin maintains airway epithelial integrity during Pseudomonas aeruginosa infection. Am. J. Respir. Cell Mol. Biol. 2010, 42, 62–68. [Google Scholar] [CrossRef]
- Alfiniyah, C.; Bees, M.A.; Wood, A.J. Quorum machinery: Effect of the Las system in Rhl regulation of P. aeruginosa. In AIP Conference Proceedings; AIP Publishing LLC.: College Park, MD, USA, 2019; Volume 2192, p. 060001. [Google Scholar] [CrossRef]
- Malhotra, S.; Hayes, D.J.; Wozniak, D.J.J. Cystic fibrosis and Pseudomonas aeruginosa: The host-microbe interface. Clin. Microbiol. Rev. 2019, 32, e00138-18. [Google Scholar] [CrossRef]
- Cornelis, P.; Dingemans, J. Pseudomonas aeruginosa adapts its iron uptake strategies in function of the type of infections. Front. Cell. Infect. Microbiol. 2013, 3, 75. [Google Scholar] [CrossRef]
- Dauner, M.; Skerra, A. Scavenging Bacterial Siderophores with Engineered Lipocalin Proteins as an Alternative Antimicrobial Strategy. ChemBioChem 2020, 21, 601–606. [Google Scholar] [CrossRef]
- Lee, J.; Zhang, L. The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell 2015, 6, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Gellatly, S.L.; Hancock, R.E. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Haq, I.J.; Gray, M.A.; Garnett, J.P.; Ward, C.; Brodlie, M. Airway surface liquid homeostasis in cystic fibrosis: Pathophysiology and therapeutic targets. Thorax 2016, 71, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Winstanley, C.; O’Brien, S.; Brockhurst, M.A. Pseudomonas aeruginosa Evolutionary Adaptation and Diversification in Cystic Fibrosis Chronic Lung Infections. Trends Microbiol. 2016, 24, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Cullen, L.; McClean, S. Bacterial Adaptation during Chronic Respiratory Infections. Pathogens 2015, 4, 66–89. [Google Scholar] [CrossRef]
- Folkesson, A.; Jelsbak, L.; Yang, L.; Johansen, H.K.; Ciofu, O.; Høiby, N.; Molin, S. Adaptation of Pseudomonas aeruginosa to the cystic fibrosis airway: An evolutionary perspective. Nat. Rev. Microbiol. 2012, 10, 841–851. [Google Scholar] [CrossRef]
- Marvig, R.L.; Damkiær, S.; Khademi, S.M.; Markussen, T.M.; Molin, S.; Jelsbak, L. Within-host evolution of Pseudomonas aeruginosa reveals adaptation toward iron acquisition from hemoglobin. mBio 2014, 5, e00966-14. [Google Scholar] [CrossRef]
- Cullen, L.; Weiser, R.; Olszak, T.; Maldonado, R.F.; Moreira, A.S.; Slachmuylders, L.; Brackman, G.; Paunova-Krasteva, T.S.; Zarnowiec, P.; Czerwonka, G.; et al. Phenotypic characterization of an international Pseudomonas aeruginosa reference panel: Strains of cystic fibrosis (CF) origin show less in vivo virulence than non-CF strains. Microbiology 2015, 161, 1961–1977. [Google Scholar] [CrossRef]
- Bianconi, I.; D’Arcangelo, S.; Esposito, A.; Benedet, M.; Piffer, E.; Dinnella, G.; Gualdi, P.; Schinella, M.; Baldo, E.; Donati, C.; et al. Persistence and m icroevolution of Pseudomonas aeruginosa in the cystic fibrosis lung: A single-patient longitudinal genomic study. Front. Microbiol. 2018, 9, 3242. [Google Scholar] [CrossRef]
- Park, J.E.; Yung, R.; Stefanowicz, D.; Shumansky, K.; Akhabir, L.; Durie, P.R.; Corey, M.; Zielenski, J.; Dorfman, R.; Daley, D.; et al. Cystic fibrosis modifier genes related to Pseudomonas aeruginosa infection. Genes Immun. 2011, 12, 370–377. [Google Scholar] [CrossRef]
- Emond, M.J.; Louie, T.; Emerson, J.; Chong, J.X.; Mathias, R.A.; Knowles, M.R.; Rieder, M.J.; Tabor, H.K.; Nickerson, D.A.; Barnes, K.C.; et al. Exome Sequencing of Phenotypic Extremes Identifies CAV2 and TMC6 as Interacting Modifiers of Chronic Pseudomonas aeruginosa Infection in Cystic Fibrosis. PLoS Genet. 2015, 11, e1005273. [Google Scholar] [CrossRef]
- Di Paola, M.; Park, A.J.; Ahmadi, S.; Roach, E.J.; Wu, Y.S.; Struder-Kypke, M.; Lam, J.S.; Bear, C.E.; Khursigara, C.M. SLC6A14 Is a Genetic Modifier of Cystic Fibrosis That Regulates Pseudomonas aeruginosa Attachment to Human Bronchial Epithelial Cells. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Castaldo, A.; Cernera, G.; Iacotucci, P.; Cimbalo, C.; Gelzo, M.; Comegna, M.; Di Lullo, A.M.; Tosco, A.; Carnovale, V.; Raia, V.; et al. TAS2R38 is a novel modifier gene in patients with cystic fibrosis. Sci. Rep. 2020, 10, 5806. [Google Scholar] [CrossRef] [PubMed]
- McIsaac, S.M.; Stadnyk, A.W.; Lin, T.J. Toll-like receptors in the host defense against Pseudomonas aeruginosa respiratory infection and cystic fibrosis. J. Leukoc. Biol. 2012, 92, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Lovewell, R.R.; Patankar, Y.R.; Berwin, B. Mechanisms of phagocytosis and host clearance of Pseudomonas aeruginosa. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L591–L603. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.K.; Kazmierczak, B.I. Inflammation: A Double-Edged Sword in the Response to Pseudomonas aeruginosa Infection. J. Innate Immun. 2017, 9, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Ramphal, R.; Balloy, V.; Huerre, M.; Si-Tahar, M.; Chignard, M. TLRs 2 and 4 are not involved in hypersusceptibility to acute Pseudomonas aeruginosa lung infections. J. Immunol. 2005, 175, 3927–3934. [Google Scholar] [CrossRef]
- Skerrett, S.J.; Wilson, C.B.; Liggitt, H.D.; Hajjar, A.M. Redundant Toll-like receptor signaling in the pulmonary host response to Pseudomonas aeruginosa. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L312–L322. [Google Scholar] [CrossRef]
- Pène, F.; Grimaldi, D.; Zuber, B.; Sauneuf, B.; Rousseau, C.; El Hachem, C.; Martin, C.; Belaïdouni, N.; Balloy, V.; Mira, J.P.; et al. Toll-like receptor 2 deficiency increases resistance to Pseudomonas aeruginosa pneumonia in the setting of sepsis-induced immune dysfunction. J. Infect. Dis. 2012, 206, 932–942. [Google Scholar] [CrossRef]
- Benmohamed, F.; Medina, M.; Wu, Y.Z.; Maschalidi, S.; Jouvion, G.; Guillemot, L.; Chignard, M.; Manoury, B.; Touqui, L. Toll-like receptor 9 deficiency protects mice against Pseudomonas aeruginosa lung infection. PLoS ONE 2014, 9, e90466. [Google Scholar] [CrossRef]
- Morris, A.E.; Liggitt, H.D.; Hawn, T.R.; Skerrett, S.J. Role of Toll-like receptor 5 in the innate immune response to acute P. aeruginosa pneumonia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L1112–L1119. [Google Scholar] [CrossRef] [PubMed]
- Vijay-Kumar, M.; Carvalho, F.A.; Aitken, J.D.; Fifadara, N.H.; Gewirtz, A.T. TLR5 or NLRC4 is necessary and sufficient for promotion of humoral immunity by flagellin. Eur. J. Immunol. 2010, 40, 3528–3534. [Google Scholar] [CrossRef] [PubMed]
- Cendra, M.D.M.; Christodoulides, M.; Hossain, P. Signaling Mediated by Toll-l ike receptor 5 sensing of Pseudomonas aeruginosa flagellin influences IL-1 β and IL-18 production by primary fibroblasts derived from the human cornea. Front. Cell. Infect. Microbiol. 2017, 7, 130. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jeon, J.; Bai, F.; Jin, S.; Wu, W.; Ha, U.H. The Pseudomonas aeruginosa HSP70-like protein DnaK induces IL-1β expression via TLR4-dependent activation of the NF-κB and JNK signaling pathways. Comp. Immunol. Microbiol. Infect. Dis. 2019, 67, 101373. [Google Scholar] [CrossRef] [PubMed]
- Tolle, L.; Yu, F.S.; Kovach, M.A.; Ballinger, M.N.; Newstead, M.W.; Zeng, X.; Nunez, G.; Standiford, T.J. Redundant and cooperative interactions between TLR5 and NLRC4 in protective lung mucosal immunity against Pseudomonas aeruginosa. J. Innate Immun. 2015, 7, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Dosunmu, E.F.; Emeh, R.O.; Dixit, S.; Bakeer, M.K.; Coats, M.T.; Owen, D.R.; Pillai, S.R.; Singh, S.R.; Dennis, V.A. The anti-microbial peptide TP359 attenuates inflammation in human lung cells infected with Pseudomonas aeruginosa via TLR5 and MAPK pathways. PLoS ONE 2017, 12, e0176640. [Google Scholar] [CrossRef]
- Pang, Z.; Junkins, R.D.; Raudonis, R.; MacNeil, A.J.; McCormick, C.; Cheng, Z.; Lin, T.J. Regulator of calcineurin 1 differentially regulates TLR-dependent MyD88 and TRIF signaling pathways. PLoS ONE 2018, 13, e0197491. [Google Scholar] [CrossRef]
- Nakamura, S.; Iwanaga, N.; Seki, M.; Fukudome, K.; Oshima, K.; Miyazaki, T.; Izumikawa, K.; Yanagihara, K.; Miyazaki, Y.; Mukae, H.; et al. Toll-Like Receptor 4 Agonistic Antibody Promotes Host Defense against Chronic Pseudomonas aeruginosa Lung Infection in Mice. Infect. Immun. 2016, 84, 1986–1993. [Google Scholar] [CrossRef]
- Lavoie, E.G.; Wangdi, T.; Kazmierczak, B.I. Innate immune responses to Pseudomonas aeruginosa infection. Microbes Infect. 2011, 13, 1133–1145. [Google Scholar] [CrossRef]
- Mijares, L.A.; Wangdi, T.; Sokol, C.; Homer, R.; Medzhitov, R.; Kazmierczak, B.I. Airway epithelial MyD88 restores control of Pseudomonas aeruginosa murine infection via an IL-1-dependent pathway. J. Immunol. 2011, 186, 7080–7088. [Google Scholar] [CrossRef]
- Hussain, S.; Johnson, C.G.; Sciurba, J.; Meng, X.; Stober, V.P.; Liu, C.; Cyphert-Daly, J.M.; Bulek, K.; Qian, W.; Solis, A.; et al. TLR5 participates in the TLR4 receptor complex and promotes MyD88-dependent signaling in environmental lung injury. Elife 2020, 9, e50458. [Google Scholar] [CrossRef] [PubMed]
- Alhazmi, A. NOD-like receptor(s) and host immune responses with Pseudomonas aeruginosa infection. Inflamm. Res. 2018, 67, 479–493. [Google Scholar] [CrossRef] [PubMed]
- McHugh, B.J.; Wang, R.; Li, H.N.; Beaumont, P.E.; Kells, R.; Stevens, H.; Young, L.; Rossi, A.G.; Gray, R.D.; Dorin, J.R.; et al. Cathelicidin is a “fire alarm”, generating protective NLRP3-dependent airway epithelial cell inflammatory responses during infection with Pseudomonas aeruginosa. PLoS Pathog. 2019, 15, e1007694. [Google Scholar] [CrossRef] [PubMed]
- Ralhan, A.; Laval, J.; Lelis, F.; Ballbach, M.; Grund, C.; Hector, A.; Hartl, D. Current Concepts and Controversies in Innate Immunity of Cystic Fibrosis Lung Disease. J. Innate Immun. 2016, 8, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Huus, K.E.; Joseph, J.; Zhang, L.; Wong, A.; Aaron, S.D.; Mah, T.F.; Sad, S. Clinical Isolates of Pseudomonas aeruginosa from Chronically Infected Cystic Fibrosis Patients Fail to Activate the Inflammasome during Both Stable Infection and Pulmonary Exacerbation. J. Immunol. 2016, 196, 3097–3108. [Google Scholar] [CrossRef]
- Bitto, N.J.; Baker, P.J.; Dowling, J.K.; Wray-McCann, G.; De Paoli, A.; Tran, L.S.; Leung, P.L.; Stacey, K.J.; Mansell, A.; Masters, S.L.; et al. Membrane vesicles from Pseudomonas aeruginosa activate the noncanonical inflammasome through caspase-5 in human monocytes. Immunol. Cell Biol. 2018, 96, 1120–1130. [Google Scholar] [CrossRef]
- Amiel, E.; Lovewell, R.R.; O’Toole, G.A.; Hogan, D.A.; Berwin, B. Pseudomonas aeruginosa evasion of phagocytosis is mediated by loss of swimming motility and is independent of flagellum expression. Infect. Immun. 2010, 78, 2937–2945. [Google Scholar] [CrossRef]
- Rada, B. Interactions between Neutrophils and Pseudomonas aeruginosa in Cystic Fibrosis. Pathogens 2017, 6. [Google Scholar] [CrossRef]
- Geddes-McAlister, J.; Kugadas, A.; Gadjeva, M. Tasked with a Challenging Objective: Why Do Neutrophils Fail to Battle Pseudomonas aeruginosa biofilms. Pathogens 2019, 8, 283. [Google Scholar] [CrossRef]
- Giacalone, V.D.; Margaroli, C.; Mall, M.A.; Tirouvanziam, R. Neutrophil Adaptations upon Recruitment to the Lung: New Concepts and Implications for Homeostasis and Disease. Int. J. Mol. Sci. 2020, 21, 851. [Google Scholar] [CrossRef]
- Laucirica, D.R.; Garratt, L.W.; Kicic, A. Progress in Model Systems of Cystic Fibrosis Mucosal Inflammation to Understand Aberrant Neutrophil Activity. Front. Immunol. 2020, 11, 595. [Google Scholar] [CrossRef] [PubMed]
- Kurahashi, K.; Sawa, T.; Ota, M.; Kajikawa, O.; Hong, K.; Martin, T.R.; Wiener-Kronish, J.P. Depletion of phagocytes in the reticuloendothelial system causes increased inflammation and mortality in rabbits with Pseudomonas aeruginosa pneumonia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L198–L209. [Google Scholar] [CrossRef] [PubMed]
- Kragh, K.N.; Alhede, M.; Jensen, P.; Moser, C.; Scheike, T.; Jacobsen, C.S.; Seier Poulsen, S.; Eickhardt-Sørensen, S.R.; Trøstrup, H.; Christoffersen, L.; et al. Polymorphonuclear leukocytes restrict growth of Pseudomonas aeruginosa in the lungs of cystic fibrosis patients. Infect. Immun. 2014, 82, 4477–4486. [Google Scholar] [CrossRef] [PubMed]
- Carevic, M.; Öz, H.; Fuchs, K.; Laval, J.; Schroth, C.; Frey, N.; Hector, A.; Bilich, T.; Haug, M.; Schmidt, A.; et al. CXCR1 Regulates Pulmonary Anti-Pseudomonas Host Defense. J. Innate Immun. 2016, 8, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.C.; Strieter, R.M.; Mehrad, B.; Newstead, M.W.; Zeng, X.; Standiford, T.J. CXC chemokine receptor CXCR2 is essential for protective innate host response in murine Pseudomonas aeruginosa pneumonia. Infect. Immun. 2000, 68, 4289–4296. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Carlson, E.; Diaconu, E.; Pearlman, E. CXCL1/KC and CXCL5/LIX are selectively produced by corneal fibroblasts and mediate neutrophil infiltration to the corneal stroma in LPS keratitis. J. Leukoc. Biol. 2007, 81, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Hou, Y.; Sun, F.; Yang, Z.; Li, C. Dysregulated Chemokine Signaling in Cystic Fibrosis Lung Disease: A Potential Therapeutic Target. Curr. Drug Targets 2016, 17, 1535–1544. [Google Scholar] [CrossRef]
- Skopelja-Gardner, S.; Theprungsirikul, J.; Lewis, K.A.; Hammond, J.H.; Carlson, K.M.; Hazlett, H.F.; Nymon, A.; Nguyen, D.; Berwin, B.L.; Hogan, D.A.; et al. Regulation of Pseudomonas aeruginosa-Mediated Neutrophil Extracellular Traps. Front. Immunol. 2019, 10, 1670. [Google Scholar] [CrossRef]
- Muntaka, S.; Almuhanna, Y.; Jackson, D.; Singh, S.; Afryie-Asante, A.; Cámara, M.; Martínez-Pomares, L. Gamma Interferon and Interleukin-17A Differentially Influence the Response of Human Macrophages and Neutrophils to Pseudomonas aeruginosa Infection. Infect. Immun. 2019, 87, e00814-18. [Google Scholar] [CrossRef]
- Omar, T.; Ziltener, P.; Chamberlain, E.; Cheng, Z.; Johnston, B. Mice Lacking γδ T Cells Exhibit Impaired Clearance of Pseudomonas aeruginosa Lung Infection and Excessive Production of Inflammatory Cytokines. Infect. Immun. 2020, 88, e00171-20. [Google Scholar] [CrossRef]
- Garai, P.; Berry, L.; Moussouni, M.; Bleves, S.; Blanc-Potard, A.B. Killing from the inside: Intracellular role of T3SS in the fate of Pseudomonas aeruginosa within macrophages revealed by mgtC and oprF mutants. PLoS Pathog. 2019, 15, e1007812. [Google Scholar] [CrossRef] [PubMed]
- Mishra, M.; Ressler, A.; Schlesinger, L.S.; Wozniak, D.J. Identification of OprF as a complement component C3 binding acceptor molecule on the surface of Pseudomonas aeruginosa. Infect. Immun. 2015, 83, 3006–3014. [Google Scholar] [CrossRef] [PubMed]
- Mauch, R.M.; Jensen, P.; Moser, C.; Levy, C.E.; Høiby, N. Mechanisms of humoral immune response against Pseudomonas aeruginosa biofilm infection in cystic fibrosis. J. Cyst. Fibros. 2018, 17, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Johansen, H.K.; Hougen, H.P.; Rygaard, J.; Høiby, N. Interferon-gamma (IFN-gamma) treatment decreases the inflammatory response in chronic Pseudomonas aeruginosa pneumonia in rats. Clin. Exp. Immunol. 1996, 103, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Barr, H.; Liu, Y.C.; Robins, A.; Heeb, S.; Williams, P.; Fogarty, A.; Cámara, M.; Martínez-Pomares, L. Granulocyte-macrophage colony stimulatory factor enhances the pro-inflammatory response of interferon-γ-treated macrophages to Pseudomonas aeruginosa infection. PLoS ONE 2015, 10, e0117447. [Google Scholar] [CrossRef] [PubMed]
- Domingue, J.C.; Drewes, J.L.; Merlo, C.A.; Housseau, F.; Sears, C.L. Host responses to mucosal biofilms in the lung and gut. Mucosal. Immunol. 2020, 13, 413–422. [Google Scholar] [CrossRef]
- Seibold, M.A. Interleukin-13 Stimulation Reveals the Cellular and Functional Plasticity of the Airway Epithelium. Ann. Am. Thorac. Soc. 2018, 15, S98–S102. [Google Scholar] [CrossRef]
- Baker, S.M.; McLachlan, J.B.; Morici, L.A. Immunological considerations in the development of Pseudomonas aeruginosa vaccines. Hum. Vaccines Immunother. 2020, 16, 412–418. [Google Scholar] [CrossRef]
- Wu, W.; Huang, J.; Duan, B.; Traficante, D.C.; Hong, H.; Risech, M.; Lory, S.; Priebe, G.P. Th17-stimulating protein vaccines confer protection against Pseudomonas aeruginosa pneumonia. Am. J. Respir. Crit. Care Med. 2012, 186, 420–427. [Google Scholar] [CrossRef]
- Hamaoka, S.; Naito, Y.; Katoh, H.; Shimizu, M.; Kinoshita, M.; Akiyama, K.; Kainuma, A.; Moriyama, K.; Ishii, K.J.; Sawa, T. Efficacy comparison of adjuvants in PcrV vaccine against Pseudomonas aeruginosa pneumonia. Microbiol. Immunol. 2017, 61, 64–74. [Google Scholar] [CrossRef]
- Schaefers, M.M.; Duan, B.; Mizrahi, B.; Lu, R.; Reznor, G.; Kohane, D.S.; Priebe, G.P. PLGA-encapsulation of the Pseudomonas aeruginosa PopB vaccine antigen improves Th17 responses and confers protection against experimental acute pneumonia. Vaccine 2018, 36, 6926–6932. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.M.; Pociask, D.; Clements, J.D.; McLachlan, J.B.; Morici, L.A. Intradermal vaccination with a Pseudomonas aeruginosa vaccine adjuvanted with a mutant bacterial ADP-ribosylating enterotoxin protects against acute pneumonia. Vaccine 2019, 37, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; McAleer, J.P.; Lin, Y.; Paterson, D.L.; Zheng, M.; Alcorn, J.F.; Weaver, C.T.; Kolls, J.K. Th17 cells mediate clade-specific, serotype-independent mucosal immunity. Immunity 2011, 35, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Bedoya, S.K.; Lam, B.; Lau, K.; Larkin, J. Th17 cells in immunity and autoimmunity. Clin. Dev. Immunol. 2013, 2013, 986789. [Google Scholar] [CrossRef] [PubMed]
- Bayes, H.K.; Ritchie, N.D.; Evans, T.J. Interleukin-17 Is Required for Control of Chronic Lung Infection Caused by Pseudomonas aeruginosa. Infect. Immun. 2016, 84, 3507–3516. [Google Scholar] [CrossRef]
- Iwanaga, N.; Kolls, J.K. Updates on T helper type 17 immunity in respiratory disease. Immunology 2019, 156, 3–8. [Google Scholar] [CrossRef]
- Neill, D.R.; Saint, G.L.; Bricio-Moreno, L.; Fothergill, J.L.; Southern, K.W.; Winstanley, C.; Christmas, S.E.; Slupsky, J.R.; McNamara, P.S.; Kadioglu, A.; et al. The B lymphocyte differentiation factor (BAFF) is expressed in the airways of children with CF and in lungs of mice infected with Pseudomonas aeruginosa. PLoS ONE 2014, 9, e95892. [Google Scholar] [CrossRef]
- Pan, T.; Tan, R.; Li, M.; Liu, Z.; Wang, X.; Tian, L.; Liu, J.; Qu, H. IL17-Producing γδ T Cells May Enhance Humoral Immunity during Pulmonary Pseudomonas aeruginosa infection in mice. Front. Cell. Infect. Microbiol. 2016, 6, 170. [Google Scholar] [CrossRef]
- Mauch, R.M.; Nørregaard, L.L.; Ciofu, O.; Levy, C.E.; Høiby, N. IgG avidity to Pseudomonas aeruginosa over the course of chronic lung biofilm infection in cystic fibrosis. J. Cyst. Fibros. 2018, 17, 356–359. [Google Scholar] [CrossRef]
- Li, Y.; Jin, L.; Chen, T. The Effects of Secretory IgA in the Mucosal Immune System. Biomed. Res. Int. 2020, 2020, 2032057. [Google Scholar] [CrossRef]
- Aanæs, K. Bacterial sinusitis can be a focus for initial lung colonisation and chronic lung infection in patients with cystic fibrosis. J. Cyst. Fibros. 2013, 12 (Suppl. 2), S1–S20. [Google Scholar] [CrossRef]
- Mauch, R.M.; Rossi, C.L.; Nolasco da Silva, M.T.; Bianchi Aiello, T.; Ribeiro, J.D.; Ribeiro, A.F.; Høiby, N.; Levy, C.E. Secretory IgA-mediated immune response in saliva and early detection of Pseudomonas aeruginosa in the lower airways of pediatric cystic fibrosis patients. Med. Microbiol. Immunol. 2019, 208, 205–213. [Google Scholar] [CrossRef] [PubMed]
- McCarron, A.; Donnelley, M.; Parsons, D. Airway disease phenotypes in animal models of cystic fibrosis. Respir. Res. 2018, 19, 54. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Yan, Z.; Engelhardt, J.F. Viral Vectors, Animal Models, and Cellular Targets for Gene Therapy of Cystic Fibrosis Lung Disease. Hum. Gene Ther. 2020, 31, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Semaniakou, A.; Croll, R.P.; Chappe, V. Animal Models in the Pathophysiology of Cystic Fibrosis. Front. Pharmacol. 2018, 9, 1475. [Google Scholar] [CrossRef] [PubMed]
- Cigana, C.; Lorè, N.I.; Riva, C.; De Fino, I.; Spagnuolo, L.; Sipione, B.; Rossi, G.; Nonis, A.; Cabrini, G.; Bragonzi, A. Tracking the immunopathological response to Pseudomonas aeruginosa during respiratory infections. Sci. Rep. 2016, 6, 21465. [Google Scholar] [CrossRef] [PubMed]
- Kukavica-Ibrulj, I.; Facchini, M.; Cigana, C.; Levesque, R.C.; Bragonzi, A. Assessing Pseudomonas aeruginosa virulence and the host response using murine models of acute and chronic lung infection. Methods Mol. Biol. 2014, 1149, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Worgall, S. 40 years on: Have we finally got a vaccine for Pseudomonas aeruginosa? Future Microbiol. 2012, 7, 1333–1335. [Google Scholar] [CrossRef] [PubMed]
- Miler, J.M.; Spilsbury, J.F.; Jones, R.J.; Roe, E.A.; Lowbury, E.J. A new polyvalent Pseudomonas vaccine. J. Med. Microbiol. 1977, 10, 19–27. [Google Scholar] [CrossRef]
- Cryz, S.J.; Furer, E.; Sadoff, J.C.; Germanier, R. Pseudomonas aeruginosa immunotype 5 polysaccharide-toxin A conjugate vaccine. Infect. Immun. 1986, 52, 161–165. [Google Scholar] [CrossRef]
- Zuercher, A.W.; Horn, M.P.; Que, J.U.; Ruedeberg, A.; Schoeni, M.H.; Schaad, U.B.; Marcus, P.; Lang, A.B. Antibody responses induced by long-term vaccination with an octovalent conjugate Pseudomonas aeruginosa vaccine in children with cystic fibrosis. FEMS Immunol. Med. Microbiol. 2006, 47, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Zuercher, A.W.; Imboden, M.A.; Jampen, S.; Bosse, D.; Ulrich, M.; Chtioui, H.; Lauterburg, B.H.; Lang, A.B. Cellular immunity in healthy volunteers treated with an octavalent conjugate Pseudomonas aeruginosa vaccine. Clin. Exp. Immunol. 2006, 143, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Bennett-Guerrero, E.; McIntosh, T.J.; Barclay, G.R.; Snyder, D.S.; Gibbs, R.J.; Mythen, M.G.; Poxton, I.R. Preparation and preclinical evaluation of a novel liposomal complete-core lipopolysaccharide vaccine. Infect. Immun. 2000, 68, 6202–6208. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.E.; Bryan, L.E. Studies on the ability of alginate to act as a protective immunogen against infection with Pseudomonas aeruginosa in animals. J. Infect. Dis. 1985, 151, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Pennington, J.E.; Pier, G.B.; Sadoff, J.C.; Small, G.J. Active and passive immunization strategies for Pseudomonas aeruginosa pneumonia. Rev. Infect. Dis. 1986, 8 (Suppl. 4), S426–S433. [Google Scholar] [CrossRef]
- Pier, G.B.; Small, G.J.; Warren, H.B. Protection against mucoid Pseudomonas aeruginosa in rodent models of endobronchial infections. Science 1990, 249, 537–540. [Google Scholar] [CrossRef]
- Cryz, S.J.; Fürer, E.; Que, J.U. Synthesis and characterization of a Pseudomonas aeruginosa alginate-toxin A conjugate vaccine. Infect. Immun. 1991, 59, 45–50. [Google Scholar] [CrossRef]
- Theilacker, C.; Coleman, F.T.; Mueschenborn, S.; Llosa, N.; Grout, M.; Pier, G.B. Construction and characterization of a Pseudomonas aeruginosa mucoid exopolysaccharide-alginate conjugate vaccine. Infect. Immun. 2003, 71, 3875–3884. [Google Scholar] [CrossRef]
- Holder, I.A.; Wheeler, R.; Montie, T.C. Flagellar preparations from Pseudomonas aeruginosa: Animal protection studies. Infect. Immun. 1982, 35, 276–280. [Google Scholar] [CrossRef]
- Holder, I.A.; Naglich, J.G. Experimental studies of the pathogenesis of infections due to Pseudomonas aeruginosa: Immunization using divalent flagella preparations. J. Trauma 1986, 26, 118–122. [Google Scholar] [CrossRef]
- Campodónico, V.L.; Llosa, N.J.; Grout, M.; Döring, G.; Maira-Litrán, T.; Pier, G.B. Evaluation of flagella and flagellin of Pseudomonas aeruginosa as vaccines. Infect. Immun. 2010, 78, 746–755. [Google Scholar] [CrossRef] [PubMed]
- Shajiei, A.; Malal, A.R.; Shahabi, G.; Farhoudi, R.; Faezi, S.; Tebianian, M.; Sohrabi, N.; Mahdavi, M. Pseudomonas aeruginosa recombinant flagellin induced poly-isotypic humoral immune responses in the Balb/C mice. Jundishapur J. Microbiol. 2013, 6, e6760. [Google Scholar] [CrossRef]
- Faezi, S.; Safarloo, M.; Amirmozafari, N.; Nikokar, I.; Siadat, S.D.; Holder, I.A.; Mahdavi, M. Protective efficacy of Pseudomonas aeruginosa type-A flagellin in the murine burn wound model of infection. APMIS 2014, 122, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Behrouz, B.; Mahdavi, M.; Amirmozafari, N.; Fatemi, M.J.; Irajian, G.; Bahroudi, M.; Hashemi, F.B. Immunogenicity of Pseudomonas aeruginosa recombinant b-type fagellin as a vaccine candidate: Protective efficacy in a murine burn wound sepsis model. Burns 2016, 16, 30026–30028. [Google Scholar] [CrossRef] [PubMed]
- Laghaei, P.; Hashemi, F.B.; Irajian, G.; Korpi, F.; Amirmozafari, N.; Behrouz, B. Immunogenicity and protective efficacy of Pseudomonas aeruginosa type a and b flagellin vaccines in a burned mouse model. Mol. Immunol. 2016, 74, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Campodónico, V.L.; Llosa, N.J.; Bentancor, L.V.; Maira-Litran, T.; Pier, G.B. Efficacy of a conjugate vaccine containing polymannuronic acid and flagellin against experimental Pseudomonas aeruginosa lung infection in mice. Infect. Immun. 2011, 79, 3455–3464. [Google Scholar] [CrossRef]
- Faezi, S.; Bahrmand, A.R.; Mahdavi, M.; Siadat, S.D.; Sardari, S.; Nikokar, I.; Khanaki, K.; Mirzajani, E.; Goudarzi, G. Preparation of Pseudomonas aeruginosa alginate-flagellin immunoconjugate. Biologicals 2017, 47, 11–17. [Google Scholar] [CrossRef]
- Farajnia, S.; Peerayeh, S.N.; Tanomand, A.; Majidi, J.; Goudarzi, G.; Naghili, B.; Rahbarnia, L. Protective efficacy of recombinant exotoxin A-flagellin fusion protein against Pseudomonas aeruginosa infection. Can. J. Microbiol. 2015, 61, 60–64. [Google Scholar] [CrossRef]
- Weimer, E.T.; Lu, H.; Kock, N.D.; Wozniak, D.J.; Mizel, S.B. A fusion protein vaccine containing OprF epitope 8, OprI, and type A and B flagellins promotes enhanced clearance of non-mucoid Pseudomonas aeruginosa. Infect. Immun. 2009, 77, 2356–2366. [Google Scholar] [CrossRef]
- Weimer, E.T.; Ervin, S.E.; Wozniak, D.J.; Mizel, S.B. Immunization of young African green monkeys with OprF epitope 8-OprI-type A- and B-flagellin fusion proteins promotes the production of protective antibodies against nonmucoid Pseudomonas aeruginosa. Vaccine 2009, 27, 6762–6769. [Google Scholar] [CrossRef]
- Hassan, R.; El-Naggar, W.; Abd El-Aziz, A.M.; Shaaban, M.; Kenawy, H.I.; Ali, Y.M. Immunization with outer membrane proteins (OprF and OprI) and flagellin B protects mice from pulmonary infection with mucoid and nonmucoid Pseudomonas aeruginosa. J. Microbiol. Immunol. Infect. 2018, 51, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Umelo-Njaka, E.; Nomellini, J.F.; Bingle, W.H.; Glasier, L.G.; Irvin, R.T.; Smit, J. Expression and testing of Pseudomonas aeruginosa vaccine candidate proteins prepared with the Caulobacter crescentus S-layer protein expression system. Vaccine 2001, 19, 1406–1415. [Google Scholar] [CrossRef]
- Ohama, M.; Hiramatsu, K.; Miyajima, Y.; Kishi, K.; Nasu, M.; Kadota, J. Intratracheal immunization with pili protein protects against mortality associated with Pseudomonas aeruginosa pneumonia in mice. FEMS Immunol. Med. Microbiol. 2006, 47, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Banadkoki, A.Z.; Keshavarzmehr, M.; Afshar, Z.; Aleyasin, N.; Fatemi, M.J.; Behrouz, B.; Hashemi, F.B. Protective effect of pilin protein with alum+naloxone adjuvant against acute pulmonary Pseudomonas aeruginosa infection. Biologicals 2016, 44, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Hertle, R.; Mrsny, R.; Fitzgerald, D.J. Dual-function vaccine for Pseudomonas aeruginosa: Characterization of chimeric exotoxin A-pilin protein. Infect. Immun. 2001, 69, 6962–6969. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.C.; Tham, D.M.; Feng, W.; Huang, F.; Embaie, S.; Liu, K.; Dean, D.; Hertle, R.; Fitzgerald, D.J.; Mrsny, R.J. Intranasal immunization strategy to impede pilin-mediated binding of Pseudomonas aeruginosa to airway epithelial cells. Infect. Immun. 2005, 73, 7705–7717. [Google Scholar] [CrossRef]
- Korpi, F.; Hashemi, F.B.; Irajian, G.; Fatemi, M.J.; Laghaei, P.; Behrouz, B. Flagellin and pilin immunization against multi-drug resistant Pseudomonas aeruginosa protects mice in the burn wound sepsis model. Immunol. Lett. 2016, 176, 8–17. [Google Scholar] [CrossRef]
- Hashemi, F.B.; Behrouz, B.; Irajian, G.; Laghaei, P.; Korpi, F.; Fatemi, M.J. A trivalent vaccine consisting of “flagellin A+B and pilin” protects against Pseudomonas aeruginosa infection in a murine burn model. Microb. Pathog. 2020, 138, 103697. [Google Scholar] [CrossRef]
- Gholami, M.; Chirani, A.S.; Razavi, S.; Falak, R.; Irajian, G. Immunogenicity of a fusion protein containing PilQ and disulphide turn region of PilA from Pseudomonas aeruginosa in mice. Lett. Appl. Microbiol. 2017, 65, 439–445. [Google Scholar] [CrossRef]
- Gholami, M.; Chirani, A.S.; Falak, R.; Moshiri, M.; Razavi, S.; Irajian, G. Induction of Specific Humoral Immune Response in Mice against a Pseudomonas aeruginosa chimeric PilQ/PilA protein. Rep. Biochem. Mol. Biol. 2018, 7, 38–44. [Google Scholar]
- Horzempa, J.; Held, T.K.; Cross, A.S.; Furst, D.; Qutyan, M.; Neely, A.N.; Castric, P. Immunization with a Pseudomonas aeruginosa 1244 pilin provides O-antigen-specific protection. Clin. Vaccine Immunol. 2008, 15, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Cachia, P.J.; Glasier, L.M.; Hodgins, R.R.; Wong, W.Y.; Irvin, R.T.; Hodges, R.S. The use of synthetic peptides in the design of a consensus sequence vaccine for Pseudomonas aeruginosa. J. Pept. Res. 1998, 52, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Kao, D.J.; Churchill, M.E.; Irvin, R.T.; Hodges, R.S. Animal protection and structural studies of a consensus sequence vaccine targeting the receptor binding domain of the type IV pilus of Pseudomonas aeruginosa. J. Mol. Biol. 2007, 374, 426–442. [Google Scholar] [CrossRef] [PubMed]
- Ahmadbeigi, Y.; Chirani, A.S.; Soleimani, N.; Mahdavi, M.; Goudarzi, M. Immunopotentiation of the engineered low-molecular-weight pilin targeting Pseudomonas aeruginosa: A combination of immunoinformatics investigation and active immunization. Mol. Immunol. 2020, 124, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Finke, M.; Duchêne, M.; Eckhardt, A.; Domdey, H.; von Specht, B.U. Protection against experimental Pseudomonas aeruginosa infection by recombinant P. aeruginosa lipoprotein I expressed in Escherichia coli. Infect. Immun. 1990, 58, 2241–2244. [Google Scholar] [CrossRef] [PubMed]
- Finke, M.; Muth, G.; Reichhelm, T.; Thoma, M.; Duchêne, M.; Hungerer, K.D.; Domdey, H.; von Specht, B.U. Protection of immunosuppressed mice against infection with Pseudomonas aeruginosa by recombinant P. aeruginosa lipoprotein I and lipoprotein I-specific monoclonal antibodies. Infect. Immun. 1991, 59, 1251–1254. [Google Scholar] [CrossRef]
- Matthews-Greer, J.M.; Robertson, D.E.; Gilleland, L.B.; Gilleland, H.E. Pseudomonas aeruginosa outer membrane protein F produced in Escherichia coli retains vaccine efficacy. Curr. Microbiol. 1990, 20, 171–175. [Google Scholar] [CrossRef]
- von Specht, B.U.; Gabelsberger, J.; Knapp, B.; Hundt, E.; Schmidt-Pilger, H.; Bauernsachs, S.; Lenz, U.; Domdey, H. Immunogenic efficacy of differently produced recombinant vaccines candidates against Pseudomonas aeruginosa infections. J. Biotechnol. 2000, 83, 3–12. [Google Scholar] [CrossRef]
- Jing, H.; Zhang, X.; Zou, J.; Yuan, Y.; Chen, Z.; Liu, D.; Wu, W.; Yang, F.; Lu, D.; Zou, Q.; et al. Oligomerization of IC43 resulted in improved immunogenicity and protective efficacy against Pseudomonas aeruginosa lung infection. Int. J. Biol. Macromol. 2020, 159, 174–182. [Google Scholar] [CrossRef]
- Cui, Z.; Han, D.; Sun, X.; Zhang, M.; Feng, X.; Sun, C.; Gu, J.; Tong, C.; Lei, L.; Han, W. Mannose-modified chitosan microspheres enhance OprF-OprI-mediated protection of mice against Pseudomonas aeruginosa infection via induction of mucosal immunity. Appl. Microbiol. Biotechnol. 2015, 99, 667–680. [Google Scholar] [CrossRef]
- Bahey-El-Din, M.; Mohamed, S.A.; Sheweita, S.A.; Haroun, M.; Zaghloul, T.I. Recombinant N-terminal outer membrane porin (OprF) of Pseudomonas aeruginosa is a promising vaccine candidate against both P. aeruginosa and some strains of Acinetobacter baumannii. Int. J. Med. Microbiol. 2020, 310, 151415. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.Y.; Shang, H.F.; Chen, T.L.; Lin, C.P.; Hui, C.F.; Hwang, J. Recombinant protein composed of Pseudomonas exotoxin A, outer membrane proteins I and F as vaccine against P. aeruginosa infection. Appl. Microbiol. Biotechnol. 1999, 52, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Gilleland, H.E.; Gilleland, L.B.; Fowler, M.R. Vaccine efficacies of elastase, exotoxin A, and outer-membrane protein F in preventing chronic pulmonary infection by Pseudomonas aeruginosa in a rat model. J. Med. Microbiol. 1993, 38, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Fakoor, M.H.; Mousavi Gargari, S.L.; Owlia, P.; Sabokbar, A. Protective Efficacy of the OprF/OprI/PcrV Recombinant Chimeric Protein Against Pseudomonas aeruginosa. Infect. Drug Resist. 2020, 13, 1651–1661. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Parlane, N.A.; Wedlock, D.N.; Rehm, B.H. Bioengineering a bacterial pathogen to assemble its own particulate vaccine capable of inducing cellular immunity. Sci. Rep. 2017, 7, 41607. [Google Scholar] [CrossRef]
- Ingle, S.A.; Mahajan, K.; Sunil Kumar, B.V.; Singh, S.; Agrawal, R.; Verma, R. Over-Expression and Immunogenicity of Outer Membrane Protein L (OprL) of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2015, 87, 217–223. [Google Scholar] [CrossRef]
- Gao, C.; Yang, F.; Wang, Y.; Liao, Y.; Zhang, J.; Zeng, H.; Zou, Q.; Gu, J. Vaccination with a recombinant OprL fragment induces a Th17 response and confers serotype-independent protection against Pseudomonas aeruginosa infection in mice. Clin. Immunol. 2017, 183, 354–363. [Google Scholar] [CrossRef]
- Liu, C.; Pan, X.; Xia, B.; Chen, F.; Jin, Y.; Bai, F.; Priebe, G.; Cheng, Z.; Jin, S.; Wu, W. Construction of a Protective Vaccine against Lipopolysaccharide-Heterologous Pseudomonas aeruginosa strains based on expression profiling of outer membrane proteins during infection. Front. Immunol. 2018, 9, 1737. [Google Scholar] [CrossRef]
- Sawa, T.; Yahr, T.L.; Ohara, M.; Kurahashi, K.; Gropper, M.A.; Wiener-Kronish, J.P.; Frank, D.W. Active and passive immunization with the Pseudomonas V antigen protects against type III intoxication and lung injury. Nat. Med. 1999, 5, 392–398. [Google Scholar] [CrossRef]
- Naito, Y.; Hamaoka, S.; Kinoshita, M.; Kainuma, A.; Shimizu, M.; Katoh, H.; Moriyama, K.; Ishii, K.J.; Sawa, T. The protective effects of nasal PcrV-CpG oligonucleotide vaccination against Pseudomonas aeruginosa pneumonia. Microbiol. Immunol. 2018, 62, 774–785. [Google Scholar] [CrossRef]
- Fakoor, M.H.; Owlia, P.; Mousavi Gargari, S.L.; Sabokbar, A. In-Silico Analysis and Protective Efficacy of the PcrV Recombinant Vaccine against Pseudomonas aeruginosa in the Burned and PA-Infected BALB/c Mouse Model. Iran. J. Immunol. 2020, 17, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, K.; Wiener-Kronish, J.P.; Sawa, T. Protective effects of affinity-purified antibody and truncated vaccines against Pseudomonas aeruginosa V-antigen in neutropenic mice. Microbiol. Immunol. 2009, 53, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Zhang, J.; Zhao, L.; Cheng, X.; Gao, C.; Wang, Y.; Xu, W.; Zou, Q.; Gu, J. Rational Design of a Chimeric Derivative of PcrV as a Subunit Vaccine against Pseudomonas aeruginosa. Front. Immunol. 2019, 10, 781. [Google Scholar] [CrossRef] [PubMed]
- Golpasha, I.D.; Mousavi, S.F.; Owlia, P.; Siadat, S.D.; Irani, S. Immunization with 3-oxododecanoyl-L-homoserine lactone-r-PcrV conjugate enhances survival of mice against lethal burn infections caused by Pseudomonas aeruginosa. Bosn. J. Basic Med. Sci. 2015, 15, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Gu, J.; Yang, L.; Gao, C.; Jing, H.; Wang, Y.; Zeng, H.; Zou, Q.; Lv, F.; Zhang, J. Protective Efficacy of the Trivalent Pseudomonas aeruginosa Vaccine Candidate PcrV-OprI-Hcp1 in Murine Pneumonia and Burn Models. Sci. Rep. 2017, 7, 3957. [Google Scholar] [CrossRef]
- Lukac, M.; Pier, G.B.; Collier, R.J. Toxoid of Pseudomonas aeruginosa exotoxin A generated by deletion of an active-site residue. Infect. Immun. 1988, 56, 3095–3098. [Google Scholar] [CrossRef]
- Chen, T.Y.; Lin, C.P.; Loa, C.C.; Chen, T.L.; Shang, H.F.; Hwang, J.; Hui, C.F. A nontoxic Pseudomonas exotoxin A induces active immunity and passive protective antibody against Pseudomonas exotoxin A intoxication. J. Biomed. Sci. 1999, 6, 357–363. [Google Scholar] [CrossRef]
- Tanomand, A.; Najar Peerayeh, S.; Farajnia, S.; Majidi, J. Protective Properties of Nontoxic Recombinant Exotoxin A (Domain I-II) against Pseudomonas aeruginosa Infection. Iran. J. Biotechnol. 2013, 11, 193–198. [Google Scholar] [CrossRef]
- Safari-Zanjani, L.; Shapouri, R.; Dezfulian, M.; Mahdavi, M.; Shafiee Ardestani, M. Exotoxin A-PLGA nanoconjugate vaccine against Pseudomonas aeruginosa infection: Protectivity in murine model. World J. Microbiol. Biotechnol. 2019, 35, 94. [Google Scholar] [CrossRef]
- Kawamoto, S.; Shibano, Y.; Fukushima, J.; Ishii, N.; Morihara, K.; Okuda, K. Site-directed mutagenesis of Glu-141 and His-223 in Pseudomonas aeruginosa elastase: Catalytic activity, processing, and protective activity of the elastase against Pseudomonas infection. Infect. Immun. 1993, 61, 1400–1405. [Google Scholar] [CrossRef]
- Sokol, P.A.; Kooi, C.; Hodges, R.F.; Cachia, P.; Woods, D.E. Immunization with a Pseudomonas aeruginosa elastase peptide reduces severity of experimental lung infections due to P. aeruginosa or Burkholderia cepacia. J. Infect. Dis. 2000, 81, 1682–1692. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.D.; Dunkley, M.L.; Moore, R.; Reynolds, S.; Bastin, D.A.; Kyd, J.M.; Cripps, A.W. Catalase immunization from Pseudomonas aeruginosa enhances bacterial clearance in the rat lung. Vaccine 2000, 19, 348–357. [Google Scholar] [CrossRef]
- Thomas, L.D.; Kyd, J.M.; Bastin, D.A.; Dunkley, M.L.; Cripps, A.W. Immunisation with non-integral OMPs promotes pulmonary clearance of Pseudomonas aeruginosa. FEMS Immunol. Med. Microbiol. 2003, 37, 155–160. [Google Scholar] [CrossRef]
- Thomas, L.D.; Cripps, A.W.; Kyd, J.M. Immune response mechanisms against Pseudomonas aeruginosa associated with mucosal immunization with protein antigens in a rat model of acute lung infection. Vaccine 2009, 27, 3324–3330. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miyairi, S.; Tateda, K.; Fuse, E.T.; Ueda, C.; Saito, H.; Takabatake, T.; Ishii, Y.; Horikawa, M.; Ishiguro, M.; Standiford, T.J.; et al. Immunization with 3-oxododecanoyl-L-homoserine lactone-protein conjugate protects mice from lethal Pseudomonas aeruginosa lung infection. J. Med. Microbiol. 2006, 55, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Sen-Kilic, E.; Blackwood, C.B.; Boehm, D.T.; Witt, W.T.; Malkowski, A.C.; Bevere, J.R.; Wong, T.Y.; Hall, J.M.; Bradford, S.D.; Varney, M.E.; et al. Intranasal Peptide-Based FpvA-KLH Conjugate Vaccine Protects Mice from Pseudomonas aeruginosa Acute Murine Pneumonia. Front. Immunol. 2019, 10, 2497. [Google Scholar] [CrossRef] [PubMed]
- Elhosary, M.A.; Bahey-El-Din, M.; AbdelBary, A.; El Guink, N.; Aboushleib, H.M. Immunization with the ferric iron-binding periplasmic protein HitA provides protection against Pseudomonas aeruginosa in the murine infection model. Microb. Pathog. 2019, 131, 181–185. [Google Scholar] [CrossRef]
- Buret, A.; Dunkley, M.; Clancy, R.L.; Cripps, A.W. Effector mechanisms of intestinally induced immunity to Pseudomonas aeruginosa in the rat lung: Role of neutrophils and leukotriene B4. Infect. Immun. 1993, 61, 671–679. [Google Scholar] [CrossRef]
- Buret, A.; Dunkley, M.L.; Pang, G.; Clancy, R.L.; Cripps, A.W. Pulmonary immunity to Pseudomonas aeruginosa in intestinally immunized rats roles of alveolar macrophages, tumor necrosis factor alpha, and interleukin-1 alpha. Infect. Immun. 1994, 62, 5335–5343. [Google Scholar] [CrossRef]
- Cripps, A.W.; Dunkley, M.L.; Clancy, R.L. Mucosal and systemic immunizations with killed Pseudomonas aeruginosa protect against acute respiratory infection in rats. Infect. Immun. 1994, 62, 1427–1436. [Google Scholar] [CrossRef]
- Fan, Y.; Mu, Y.; Lu, L.; Tian, Y.; Yuan, F.; Zhou, B.; Yu, C.; Wang, Z.; Li, X.; Lei, S.; et al. Hydrogen peroxide-inactivated bacteria induces potent humoral and cellular immune responses and releases nucleic acids. Int. Immunopharmacol. 2019, 69, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Meynet, E.; Laurin, D.; Lenormand, J.L.; Camara, B.; Toussaint, B.; Le Gouëllec, A. Killed but metabolically active Pseudomonas aeruginosa-based vaccine induces protective humoral- and cell-mediated immunity against Pseudomonas aeruginosa pulmonary infections. Vaccine 2018, 36, 1893–1900. [Google Scholar] [CrossRef] [PubMed]
- Priebe, G.P.; Brinig, M.M.; Hatano, K.; Grout, M.; Coleman, F.T.; Pier, G.B.; Goldberg, J.B. Construction and characterization of a live, attenuated aroA deletion mutant of Pseudomonas aeruginosa as a candidate intranasal vaccine. Infect. Immun. 2002, 70, 1507–1517. [Google Scholar] [CrossRef] [PubMed]
- Priebe, G.P.; Meluleni, G.J.; Coleman, F.T.; Goldberg, J.B.; Pier, G.B. Protection against fatal Pseudomonas aeruginosa pneumonia in mice after nasal immunization with a live, attenuated aroA deletion mutant. Infect. Immun. 2003, 71, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Priebe, G.P.; Walsh, R.L.; Cederroth, T.A.; Kamei, A.; Coutinho-Sledge, Y.S.; Goldberg, J.B.; Pier, G.B. IL-17 is a critical component of vaccine-induced protection against lung infection by lipopolysaccharide-heterologous strains of Pseudomonas aeruginosa. J. Immunol. 2008, 181, 4965–4975. [Google Scholar] [CrossRef] [PubMed]
- Kamei, A.; Coutinho-Sledge, Y.S.; Goldberg, J.B.; Priebe, G.P.; Pier, G.B. Mucosal vaccination with a multivalent, live-attenuated vaccine induces multifactorial immunity against Pseudomonas aeruginosa acute lung infection. Infect. Immun. 2011, 79, 1289–1299. [Google Scholar] [CrossRef] [PubMed]
- Cabral, M.P.; Correia, A.; Vilanova, M.; Gärtner, F.; Moscoso, M.; García, P.; Vallejo, J.A.; Pérez, A.; Francisco-Tomé, M.; Fuentes-Valverde, V.; et al. A live auxotrophic vaccine confers mucosal immunity and protection against lethal pneumonia caused by Pseudomonas aeruginosa. PLoS Pathog. 2020, 16, e1008311. [Google Scholar] [CrossRef]
- Saha, S.; Takeshita, F.; Matsuda, T.; Jounai, N.; Kobiyama, K.; Matsumoto, T.; Sasaki, S.; Yoshida, A.; Xin, K.Q.; Klinman, D.M.; et al. Blocking of the TLR5 activation domain hampers protective potential of flagellin DNA vaccine. J. Immunol. 2007, 179, 1147–1154. [Google Scholar] [CrossRef]
- Price, B.M.; Galloway, D.R.; Baker, N.R.; Gilleland, L.B.; Staczek, J.; Gilleland, H.E. Protection against Pseudomonas aeruginosa chronic lung infection in mice by genetic immunization against outer membrane protein F (OprF) of P. aeruginosa. Infect. Immun. 2001, 69, 3510–3515. [Google Scholar] [CrossRef]
- Price, B.M.; Barten Legutki, J.; Galloway, D.R.; von Specht, B.U.; Gilleland, L.B.; Gilleland, H.E.; Staczek, J. Enhancement of the protective efficacy of an oprF DNA vaccine against Pseudomonas aeruginosa. FEMS Immunol. Med. Microbiol. 2002, 33, 89–99. [Google Scholar] [CrossRef]
- Staczek, J.; Gilleland, L.B.; van der Heyde, H.C.; Gilleland, H.E. DNA vaccines against chronic lung infections by Pseudomonas aeruginosa. FEMS Immunol. Med. Microbiol. 2003, 37, 147–153. [Google Scholar] [CrossRef]
- Gong, Q.; Ruan, M.D.; Niu, M.F.; Qin, C.L.; Hou, Y.; Guo, J.Z. Immune efficacy of DNA vaccines based on oprL and oprF genes of Pseudomonas aeruginosa in chickens. Poult. Sci. 2018, 97, 4219–4227. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Wang, Y.; Xia, Y.; Zhang, L.; Yang, Q.; Lei, J. A DNA vaccine encoding VP22 of herpes simplex virus type I (HSV-1) and OprF confers enhanced protection from Pseudomonas aeruginosa in mice. Vaccine 2016, 34, 4399–4405. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Takeshita, F.; Sasaki, S.; Matsuda, T.; Tanaka, T.; Tozuka, M.; Takase, K.; Matsumoto, T.; Okuda, K.; Ishii, N.; et al. Multivalent DNA vaccine protects mice against pulmonary infection caused by Pseudomonas aeruginosa. Vaccine 2006, 24, 6240–6249. [Google Scholar] [CrossRef]
- Pennington, J.E. Preliminary investigations of Pseudomonas aeruginosa vaccine in patients with leukemia and cystic fibrosis. J. Infect. Dis. 1974, 130, S159–S162. [Google Scholar] [CrossRef]
- Pennington, J.E.; Reynolds, H.Y.; Wood, R.E.; Robinson, R.A.; Levine, A.S. Use of a Pseudomonas aeruginosa vaccine in pateints with acute leukemia and cystic fibrosis. Am. J. Med. 1975, 58, 629–636. [Google Scholar] [CrossRef]
- Jones, R.J.; Roe, E.A.; Lowbury, E.J.; Miler, J.J.; Spilsbury, J.F. A new Pseudomonas vaccine: Preliminary trial on human volunteers. J. Hyg. (Lond.) 1976, 76, 429–439. [Google Scholar] [CrossRef]
- Langford, D.T.; Hiller, J. Prospective, controlled study of a polyvalent Pseudomonas vaccine in cystic fibrosis-three year results. Arch. Dis. Child. 1984, 59, 1131–1134. [Google Scholar] [CrossRef]
- Cryz, S.J.; Fürer, E.; Sadoff, J.C.; Germanier, R. A polyvalent Pseudomonas aeruginosa O-polysaccharide-toxin A conjugate vaccine. Antibiot. Chemother. (1971) 1987, 39, 249–255. [Google Scholar] [CrossRef]
- Cryz, S.J.; Sadoff, J.C.; Cross, A.S.; Fürer, E. Safety and immunogenicity of a polyvalent Pseudomonas aeruginosa O-polysaccharide-toxin A vaccine in humans. Antibiot. Chemother. (1971) 1989, 42, 177–183. [Google Scholar] [CrossRef]
- Lang, A.B.; Rüdeberg, A.; Schöni, M.H.; Que, J.U.; Fürer, E.; Schaad, U.B. Vaccination of cystic fibrosis patients against Pseudomonas aeruginosa reduces the proportion of patients infected and delays time to infection. Pediatr. Infect. Dis. J. 2004, 23, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Döring, G. Prevention of Pseudomonas aeruginosa infection in cystic fibrosis patients. Int. J. Med. Microbiol. 2010, 300, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Pier, G.B.; DesJardin, D.; Grout, M.; Garner, C.; Bennett, S.E.; Pekoe, G.; Fuller, S.A.; Thornton, M.O.; Harkonen, W.S.; Miller, H.C. Human immune response to Pseudomonas aeruginosa mucoid exopolysaccharide (alginate) vaccine. Infect. Immun. 1994, 62, 3972–3979. [Google Scholar] [CrossRef] [PubMed]
- Crowe, B.A.; Enzersberger, O.; Schober-Bendixen, S.; Mitterer, A.; Mundt, W.; Livey, I.; Pabst, H.; Kaeser, R.; Eibl, M.; Eibl, J. The first clinical trial of immuno’s experimental Pseudomonas aeruginosa flagellar vaccines. Antibiot. Chemother. (1971) 1991, 44, 143–156. [Google Scholar] [CrossRef]
- Döring, G.; Pfeiffer, C.; Weber, U.; Mohr-Pennert, A.; Dorner, F. Parenteral application of a Pseudomonas aeruginosa flagella vaccine elicits specific anti-flagella antibodies in the airways of healthy individuals. Am. J. Respir. Crit. Care Med. 1995, 151, 983–985. [Google Scholar] [CrossRef] [PubMed]
- Döring, G.; Meisner, C.; Stern, M.; Flagella Vaccine Trial Study Group. A double-blind randomized placebo-controlled phase III study of a Pseudomonas aeruginosa flagella vaccine in cystic fibrosis patients. Proc. Natl. Acad. Sci. USA 2007, 104, 11020–11025. [Google Scholar] [CrossRef]
- Jang, I.J.; Kim, I.S.; Park, W.J.; Yoo, K.S.; Yim, D.S.; Kim, H.K.; Shin, S.G.; Chang, W.H.; Lee, N.G.; Jung, S.B.; et al. Human immune response to a Pseudomonas aeruginosa outer membrane protein vaccine. Vaccine 1999, 17, 158–168. [Google Scholar] [CrossRef]
- Lee, N.G.; Jung, S.B.; Ahn, B.Y.; Kim, Y.H.; Kim, J.J.; Kim, D.K.; Kim, I.S.; Yoon, S.M.; Nam, S.W.; Kim, H.S.; et al. Immunization of burn-patients with a Pseudomonas aeruginosa outer membrane protein vaccine elicits antibodies with protective efficacy. Vaccine 2000, 18, 1952–1961. [Google Scholar] [CrossRef]
- Kim, D.K.; Kim, J.J.; Kim, J.H.; Woo, Y.M.; Kim, S.; Yoon, D.W.; Choi, C.S.; Kim, I.; Park, W.J.; Lee, N.; et al. Comparison of two immunization schedules for a Pseudomonas aeruginosa outer membrane proteins vaccine in burn patients. Vaccine 2000, 19, 1274–1283. [Google Scholar] [CrossRef]
- von Specht, B.; Knapp, B.; Hungerer, K.; Lücking, C.; Schmitt, A.; Domdey, H. Outer membrane proteins of Pseudomonas aeruginosa as vaccine candidates. J. Biotechnol. 1996, 44, 145–153. [Google Scholar] [CrossRef]
- Mansouri, E.; Gabelsberger, J.; Knapp, B.; Hundt, E.; Lenz, U.; Hungerer, K.D.; Gilleland, H.E.; Staczek, J.; Domdey, H.; von Specht, B.U. Safety and immunogenicity of a Pseudomonas aeruginosa hybrid outer membrane protein F-I vaccine in human volunteers. Infect. Immun. 1999, 67, 1461–1470. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, E.; Blome-Eberwein, S.; Gabelsberger, J.; Germann, G.; von Specht, B.U. Clinical study to assess the immunogenicity and safety of a recombinant Pseudomonas aeruginosa OprF-OprI vaccine in burn patients. FEMS Immunol. Med. Microbiol. 2003, 37, 161–166. [Google Scholar] [CrossRef]
- Westritschnig, K.; Hochreiter, R.; Wallner, G.; Firbas, C.; Schwameis, M.; Jilma, B. A randomized, placebo-controlled phase I study assessing the safety and immunogenicity of a Pseudomonas aeruginosa hybrid outer membrane protein OprF/I vaccine (IC43) in healthy volunteers. Hum. Vaccines Immunother. 2014, 10, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Rello, J.; Krenn, C.G.; Locker, G.; Pilger, E.; Madl, C.; Balica, L.; Dugernier, T.; Laterre, P.F.; Spapen, H.; Depuydt, P.; et al. A randomized placebo-controlled phase II study of a Pseudomonas vaccine in ventilated ICU patients. Crit. Care 2017, 21, 22. [Google Scholar] [CrossRef] [PubMed]
- Larbig, M.; Mansouri, E.; Freihorst, J.; Tümmler, B.; Köhler, G.; Domdey, H.; Knapp, B.; Hungerer, K.D.; Hundt, E.; Gabelsberger, J.; et al. Safety and immunogenicity of an intranasal Pseudomonas aeruginosa hybrid outer membrane protein F-I vaccine in human volunteers. Vaccine 2001, 19, 2291–2297. [Google Scholar] [CrossRef]
- Baumann, U.; Göcke, K.; Gewecke, B.; Freihorst, J.; von Specht, B.U. Assessment of pulmonary antibodies with induced sputum and bronchoalveolar lavage induced by nasal vaccination against Pseudomonas aeruginosa: A clinical phase I/II study. Respir. Res. 2007, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Cripps, A.W.; Dunkley, M.L.; Clancy, R.L.; Kyd, J. Vaccine strategies against Pseudomonas aeruginosa infection in the lung. Behring Inst. Mitt. 1997, 98, 262–268. [Google Scholar]
- Cripps, A.W.; Peek, K.; Dunkley, M.; Vento, K.; Marjason, J.K.; McIntyre, M.E.; Sizer, P.; Croft, D.; Sedlak-Weinstein, L. Safety and immunogenicity of an oral inactivated whole-cell Pseudomonas aeruginosa vaccine administered to healthy human subjects. Infect. Immun. 2006, 74, 968–974. [Google Scholar] [CrossRef]
- Hanessian, S.; Regan, W.; Watson, D.; Haskell, T.H. Isolation and characterization of antigenic components of a new heptavalent Pseudomonas vaccine. Nat. New Biol. 1971, 229, 209–210. [Google Scholar] [CrossRef]
- Young, L.S.; Meyer, R.D.; Armstrong, D. Pseudomonas aeruginosa vaccine in cancer patients. Ann. Intern. Med. 1973, 79, 518–527. [Google Scholar] [CrossRef]
- Wesley, J.; Fisher, A.; Fisher, M.W. Immunization against Pseudomonas in infection after thermal injury. J. Infect. Dis. 1974, 130, S152–S158. [Google Scholar] [CrossRef] [PubMed]
- MacIntyre, S.; McVeigh, T.; Owen, P. Immunochemical and biochemical analysis of the polyvalent Pseudomonas aeruginosa vaccine PEV. Infect. Immun. 1986, 51, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Erridge, C.; Stewart, J.; Bennett-Guerrero, E.; McIntosh, T.J.; Poxton, I.R. The biological activity of a liposomal complete core lipopolysaccharide vaccine. J. Endotoxin Res. 2002, 8, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Pier, G.B.; Thomas, D.M. Lipopolysaccharide and high-molecular-weight polysaccharide serotypes of Pseudomonas aeruginosa. J. Infect. Dis. 1982, 145, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Pier, G.B. Safety and immunogenicity of high molecular weight polysaccharide vaccine from immunotype 1 Pseudomonas aeruginosa. J. Clin. Invest. 1982, 69, 303–308. [Google Scholar] [CrossRef]
- Hatano, K.; Pier, G.B. Complex serology and immune response of mice to variant high-molecular-weight O polysaccharides isolated from Pseudomonas aeruginosa serogroup O2 strains. Infect. Immun. 1998, 66, 3719–3726. [Google Scholar] [CrossRef]
- Cryz, S.J.; Fürer, E.; Cross, A.S.; Wegmann, A.; Germanier, R.; Sadoff, J.C. Safety and immunogenicity of a Pseudomonas aeruginosa O-polysaccharide toxin A conjugate vaccine in humans. J. Clin. Investig. 1987, 80, 51–56. [Google Scholar] [CrossRef]
- Schaad, U.B.; Lang, A.B.; Wedgwood, J.; Ruedeberg, A.; Que, J.U.; Fürer, E.; Cryz, S.J. Safety and immunogenicity of Pseudomonas aeruginosa conjugate A vaccine in cystic fibrosis. Lancet 1991, 338, 1236–1237. [Google Scholar] [CrossRef]
- Cryz, S.J.; Wedgwood, J.; Lang, A.B.; Ruedeberg, A.; Que, J.U.; Fürer, E.; Schaad, U.B. Immunization of noncolonized cystic fibrosis patients against Pseudomonas aeruginosa. J. Infect. Dis. 1994, 169, 1159–1162. [Google Scholar] [CrossRef][Green Version]
- Lang, A.B.; Schaad, U.B.; Rüdeberg, A.; Wedgwood, J.; Que, J.U.; Fürer, E.; Cryz, S.J. Effect of high-affinity anti-Pseudomonas aeruginosa lipopolysaccharide antibodies induced by immunization on the rate of Pseudomonas aeruginosa infection in patients with cystic fibrosis. J. Pediatr. 1995, 127, 711–717. [Google Scholar] [CrossRef]
- Cryz, S.J.; Lang, A.; Rüdeberg, A.; Wedgwood, J.; Que, J.U.; Fürer, E.; Schaad, U. Immunization of cystic fibrosis patients with a Pseudomonas aeruginosa O-polysaccharide-toxin A conjugate vaccine. Behring Inst. Mitt. 1997, 98, 345–349. [Google Scholar]
- Al-Zeer, M.; Masoud, H. LPS-based conjugate vaccines composed of O-polysaccharide from Pseudomonas aeruginosa IATS 6 and 11 bound to a carrier protein. World J. Microbiol. Biotechnol. 2007, 23, 1541–1549. [Google Scholar] [CrossRef]
- Xie, M.C.; Chen, M.T.; Jiang, X.; Yang, S.; Wu, H.Z.; Wang, X.; Zhao, G.M.; Ge, Y.H. Preparation and immune characteristics of Pseudomonas aeruginosa-IATS 3 O-specific polysaccharide-tetanus toxoid conjugate. Chin. J. Biol. 2012, 25, 1565–1568. [Google Scholar]
- Gavinho, B.; dos Santos, R.R.; Caron, L.F.; Sierakowski, M.R.; Ono, L. Development of a conjugated immunogen of Pseudomonas aeruginosa lipopolysaccharide and tetanus toxoid. Arch. Vet. Sci. 2012, 17, 58–67. [Google Scholar]
- Najafzadeh, F.; Shapouri, R.; Rahnema, M.; Rokhsartalab Azar, S.; Kianmehr, A. Pseudomonas aeruginosa PAO-1 Lipopolysaccharide-Diphtheria Toxoid Conjugate Vaccine: Preparation, Characterization and Immunogenicity. Jundishapur J. Microbiol. 2015, 8, e17712. [Google Scholar] [CrossRef]
- Bryan, L.E.; Kureishi, A.; Rabin, H.R. Detection of antibodies to Pseudomonas aeruginosa alginate extracellular polysaccharide in animals and cystic fibrosis patients by enzyme-linked immunosorbent assay. J. Clin. Microbiol. 1983, 18, 276–282. [Google Scholar] [CrossRef]
- Pier, G.B.; Matthews, W.J.; Eardley, D.D. Immunochemical characterization of the mucoid exopolysaccharide of Pseudomonas aeruginosa. J. Infect. Dis. 1983, 147, 494–503. [Google Scholar] [CrossRef]
- Speert, D.P.; Lawton, D.; Mutharia, L.M. Antibody to Pseudomonas aeruginosa mucoid exopolysaccharide and to sodium alginate in cystic fibrosis serum. Pediatr. Res. 1984, 18, 431–433. [Google Scholar] [CrossRef]
- Pedersen, S.S.; Espersen, F.; Høiby, N.; Shand, G.H. Purification, characterization, and immunological cross-reactivity of alginates produced by mucoid Pseudomonas aeruginosa from patients with cystic fibrosis. J. Clin. Microbiol. 1989, 27, 691–699. [Google Scholar] [CrossRef]
- Ames, P.; DesJardins, D.; Pier, G.B. Opsonophagocytic killing activity of rabbit antibody to Pseudomonas aeruginosa mucoid exopolysaccharide. Infect. Immun. 1985, 49, 281–285. [Google Scholar] [CrossRef]
- Pier, G.B.; Saunders, J.M.; Ames, P.; Edwards, M.S.; Auerbach, H.; Goldfarb, J.; Speert, D.P.; Hurwitch, S. Opsonophagocytic killing antibody to Pseudomonas aeruginosa mucoid exopolysaccharide in older noncolonized patients with cystic fibrosis. N. Engl. J. Med. 1987, 317, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Garner, C.V.; DesJardins, D.; Pier, G.B. Immunogenic properties of Pseudomonas aeruginosa mucoid exopolysaccharide. Infect. Immun. 1990, 58, 1835–1842. [Google Scholar] [CrossRef] [PubMed]
- Pier, G.B.; Coleman, F.; Grout, M.; Franklin, M.; Ohman, D.E. Role of alginate O acetylation in resistance of mucoid Pseudomonas aeruginosa to opsonic phagocytosis. Infect. Immun. 2001, 69, 1895–1901. [Google Scholar] [CrossRef] [PubMed]
- Kashef, N.; Behzadian-Nejad, Q.; Najar-Peerayeh, S.; Mousavi-Hosseini, K.; Moazzeni, M.; Djavid, G.E. Synthesis and characterization of Pseudomonas aeruginosa alginate-tetanus toxoid conjugate. J. Med. Microbiol. 2006, 55, 1441–1446. [Google Scholar] [CrossRef]
- Farjaha, A.; Owlia, P.; Siadat, S.D.; Mousavi, S.F.; Shafieeardestani, M. Conjugation of alginate to a synthetic peptide containing T- and B-cell epitopes as an induction for protective immunity against Pseudomonas aeruginosa. J. Biotechnol. 2014, 192. [Google Scholar] [CrossRef]
- Najafzadeh, F.; Jaberi, G.; Shapouri, R.; Rahnema, M.; Karimi-Nik, A.; Kianmehr, A. Immunogenicity comparison of conjugate vaccines composed of alginate and lipopolysaccharide of Pseudomonas aeruginosa bound to diphtheria toxoid. Iran. J. Microbiol. 2014, 6, 317–323. [Google Scholar]
- Farjah, A.; Owlia, P.; Siadat, S.D.; Mousavi, S.F.; Ardestani, M.S.; Mohammadpour, H.K. Immunological evaluation of an alginate-based conjugate as a vaccine candidate against Pseudomonas aeruginosa. APMIS 2015, 123, 175–183. [Google Scholar] [CrossRef]
- Alikhani, Z.; Salouti, M.; Ardestani, M.S. Synthesis and immunological evaluation of a nanovaccine based on PLGA nanoparticles and alginate antigen against infections caused by Pseudomonas aeruginosa. Biomed. Phys. Eng. Express 2018, 4, 045016. [Google Scholar] [CrossRef]
- Neville, L.F.; Barnea, Y.; Hammer-Munz, O.; Gur, E.; Kuzmenko, B.; Kahel-Raifer, H.; Eren, R.; Elkeles, A.; Murthy, K.G.; Szabó, C.; et al. Antibodies raised against N′-terminal Pseudomonas aeruginosa flagellin prevent mortality in lethal murine models of infection. Int. J. Mol. Med. 2005, 16, 165–171. [Google Scholar] [CrossRef]
- Dakterzada, F.; Mohabati Mobarez, A.; Habibi Roudkenar, M.; Mohsenifar, A. Induction of humoral immune response against Pseudomonas aeruginosa flagellin(1-161) using gold nanoparticles as an adjuvant. Vaccine 2016, 34, 1472–1479. [Google Scholar] [CrossRef]
- Castric, P.A.; Deal, C.D. Differentiation of Pseudomonas aeruginosa pili based on sequence and B-cell epitope analyses. Infect. Immun. 1994, 62, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Sheth, H.B.; Glasier, L.M.; Ellert, N.W.; Cachia, P.; Kohn, W.; Lee, K.K.; Paranchych, W.; Hodges, R.S.; Irvin, R.T. Development of an anti-adhesive vaccine for Pseudomonas aeruginosa targeting the C-terminal region of the pilin structural protein. Biomed. Pept. Proteins Nucleic Acids 1995, 1, 141–148. [Google Scholar] [PubMed]
- Lee, K.K.; Sheth, H.B.; Wong, W.Y.; Sherburne, R.; Paranchych, W.; Hodges, R.S.; Lingwood, C.A.; Krivan, H.; Irvin, R.T. The binding of Pseudomonas aeruginosa pili to glycosphingolipids is a tip-associated event involving the C-terminal region of the structural pilin subunit. Mol. Microbiol. 1994, 11, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Hazes, B.; Sastry, P.A.; Hayakawa, K.; Read, R.J.; Irvin, R.T. Crystal structure of Pseudomonas aeruginosa PAK pilin suggests a main-chain-dominated mode of receptor binding. J. Mol. Biol. 2000, 299, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Keizer, D.W.; Slupsky, C.M.; Kalisiak, M.; Campbell, A.P.; Crump, M.P.; Sastry, P.A.; Hazes, B.; Irvin, R.T.; Sykes, B.D. Structure of a pilin monomer from Pseudomonas aeruginosa: Implications for the assembly of pili. J. Biol. Chem. 2001, 276, 24186–24193. [Google Scholar] [CrossRef]
- Audette, G.F.; Irvin, R.T.; Hazes, B. Crystallographic analysis of the Pseudomonas aeruginosa strain K122-4 monomeric pilin reveals a conserved receptor-binding architecture. Biochemistry 2004, 43, 11427–11435. [Google Scholar] [CrossRef]
- Kus, J.V.; Tullis, E.; Cvitkovitch, D.G.; Burrows, L.L. Significant differences in type IV pilin allele distribution among Pseudomonas aeruginosa isolates from cystic fibrosis (CF) versus non-CF patients. Microbiology 2004, 150, 1315–1326. [Google Scholar] [CrossRef]
- Campbell, A.P.; Wong, W.Y.; Houston, M.; Schweizer, F.; Cachia, P.J.; Irvin, R.T.; Hindsgaul, O.; Hodges, R.S.; Sykes, B.D. Interaction of the receptor binding domains of Pseudomonas aeruginosa pili strains PAK, PAO, KB7 and P1 to a cross-reactive antibody and receptor analog: Implications for synthetic vaccine design. J. Mol. Biol. 1997, 267, 382–402. [Google Scholar] [CrossRef]
- Kao, D.J.; Hodges, R.S. Advantages of a synthetic peptide immunogen over a protein immunogen in the development of an anti-pilus vaccine for Pseudomonas aeruginosa. Chem. Biol. Drug Des. 2009, 74, 33–42. [Google Scholar] [CrossRef]
- Hackbarth, C.; Hodges, R.S. Synthetic peptide vaccine development: Designing dual epitopes into a single pilin peptide immunogen generates antibody cross-reactivity between two strains of Pseudomonas aeruginosa. Chem. Biol. Drug Des. 2010, 76, 293–304. [Google Scholar] [CrossRef][Green Version]
- Faezi, S.; Bahrmand, A.R.; Siadat, S.D.; Nikokar, I.; Sardari, S.; Mahdavi, M. Construction, expression, purification and characterization of secretin domain of PilQ and triple PilA-related disulfide loop peptides fusion protein from Pseudomonas aeruginosa. Iran. J. Basic Med. Sci. 2017, 20, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Faezi, S.; Nikokar, I.; Elmi, A.; Ghasemi, Y.; Farahbakhsh, M.; Salimi Chirani, A.; Mahdavi, M. Molecular characterization and functional analysis of the PilQ380-706: A novel secretin domain in Pseudomonas aeruginosa. Avicenna J. Med. Biotechnol. 2018, 10, 34–40. [Google Scholar] [PubMed]
- Fernandes, P.B.; Kim, C.; Cundy, K.R.; Haung, N.N. Antibodies to cell envelope proteins of Pseudomonas aeruginosa in cystic fibrosis patients. Infect. Immun. 1981, 33, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Mutharia, L.M.; Nicas, T.I.; Hancock, R.E. Outer membrane proteins of Pseudomonas aeruginosa serotype strains. J. Infect. Dis. 1982, 146, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Mutharia, L.M.; Hancock, R.E. Surface localization of Pseudomonas aeruginosa outer membrane porin protein F by using monoclonal antibodies. Infect. Immun. 1983, 42, 1027–1033. [Google Scholar] [CrossRef]
- Gilleland, H.E.; Parker, M.G.; Matthews, J.M.; Berg, R.D. Use of a purified outer membrane protein F (porin) preparation of Pseudomonas aeruginosa as a protective vaccine in mice. Infect. Immun. 1984, 44, 49–54. [Google Scholar] [CrossRef]
- von Specht, B.U.; Strigl, G.; Ehret, W.; Brendel, W. Protective effect of an outer membrane vaccine against Pseudomonas aeruginosa infection. Infection 1987, 15, 408–412. [Google Scholar] [CrossRef]
- Matthews-Greer, J.M.; Gilleland, H.E. Outer membrane protein F (porin) preparation of Pseudomonas aeruginosa as a protective vaccine against heterologous immunotype strains in a burned mouse model. J. Infect. Dis. 1987, 155, 1282–1291. [Google Scholar] [CrossRef]
- Gilleland, H.E.; Gilleland, L.B.; Matthews-Greer, J.M. Outer membrane protein F preparation of Pseudomonas aeruginosa as a vaccine against chronic pulmonary infection with heterologous immunotype strains in a rat model. Infect. Immun. 1988, 56, 1017–1022. [Google Scholar] [CrossRef]
- Fox, C.W.; Campbell, G.D.; Anderson, W.M.; Zavecz, J.H.; Gilleland, L.B.; Gilleland, H.E. Preservation of pulmonary function by an outer membrane protein F vaccine. A study in rats with chronic pulmonary infection caused by Pseudomonas aeruginosa. Chest 1994, 105, 1545–1550. [Google Scholar] [CrossRef]
- Hughes, E.E.; Gilleland, L.B.; Gilleland, H.E. Synthetic peptides representing epitopes of outer membrane protein F of Pseudomonas aeruginosa that elicit antibodies reactive with whole cells of heterologous immunotype strains of P. aeruginosa. Infect. Immun. 1992, 60, 3497–3503. [Google Scholar] [CrossRef] [PubMed]
- Baumann, U.; Mansouri, E.; von Specht, B.U. Recombinant OprF-OprI as a vaccine against Pseudomonas aeruginosa infections. Vaccine 2004, 22, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Göcke, K.; Baumann, U.; Hagemann, H.; Gabelsberger, J.; Hahn, H.; Freihorst, J.; von Specht, B.U. Mucosal vaccination with a recombinant OprF-I vaccine of Pseudomonas aeruginosa in healthy volunteers: Comparison of a systemic vs. a mucosal booster schedule. FEMS Immunol. Med. Microbiol. 2003, 37, 167–171. [Google Scholar] [CrossRef]
- Sorichter, S.; Baumann, U.; Baumgart, A.; Walterspacher, S.; von Specht, B.U. Immune responses in the airways by nasal vaccination with systemic boosting against Pseudomonas aeruginosa in chronic lung disease. Vaccine 2009, 27, 2755–2759. [Google Scholar] [CrossRef] [PubMed]
- Staczek, J.; Gilleland, H.E.; Gilleland, L.B.; Harty, R.N.; García-Sastre, A.; Engelhardt, O.G.; Palese, P. A chimeric influenza virus expressing an epitope of outer membrane protein F of Pseudomonas aeruginosa affords protection against challenge with P. aeruginosa in a murine model of chronic pulmonary infection. Infect. Immun. 1998, 66, 3990–3994. [Google Scholar] [CrossRef]
- Brennan, F.R.; Jones, T.D.; Gilleland, L.B.; Bellaby, T.; Xu, F.; North, P.C.; Thompson, A.; Staczek, J.; Lin, T.; Johnson, J.E.; et al. Pseudomonas aeruginosa outer-membrane protein F epitopes are highly immunogenic in mice when expressed on a plant virus. Microbiology 1999, 145, 211–220. [Google Scholar] [CrossRef]
- Gilleland, H.E.; Gilleland, L.B.; Staczek, J.; Harty, R.N.; García-Sastre, A.; Palese, P.; Brennan, F.R.; Hamilton, W.D.; Bendahmane, M.; Beachy, R.N. Chimeric animal and plant viruses expressing epitopes of outer membrane protein F as a combined vaccine against Pseudomonas aeruginosa lung infection. FEMS Immunol. Med. Microbiol. 2000, 27, 291–297. [Google Scholar] [CrossRef]
- Staczek, J.; Bendahmane, M.; Gilleland, L.B.; Beachy, R.N.; Gilleland, H.E. Immunization with a chimeric tobacco mosaic virus containing an epitope of outer membrane protein F of Pseudomonas aeruginosa provides protection against challenge with P. aeruginosa. Vaccine 2000, 18, 2266–2274. [Google Scholar] [CrossRef]
- Worgall, S.; Krause, A.; Rivara, M.; Hee, K.K.; Vintayen, E.V.; Hackett, N.R.; Roelvink, P.W.; Bruder, J.T.; Wickham, T.J.; Kovesdi, I.; et al. Protection against P. aeruginosa with an adenovirus vector containing an OprF epitope in the capsid. J. Clin. Investig. 2005, 115, 1281–1289. [Google Scholar] [CrossRef]
- Worgall, S.; Krause, A.; Qiu, J.; Joh, J.; Hackett, N.R.; Crystal, R.G. Protective immunity to Pseudomonas aeruginosa induced with a capsid-modified adenovirus expressing P. aeruginosa OprF. J. Virol. 2007, 81, 13801–13808. [Google Scholar] [CrossRef]
- Krause, A.; Whu, W.Z.; Xu, Y.; Joh, J.; Crystal, R.G.; Worgall, S. Protective anti-Pseudomonas aeruginosa humoral and cellular mucosal immunity by AdC7-mediated expression of the P. aeruginosa protein OprF. Vaccine 2011, 29, 2131–2139. [Google Scholar] [CrossRef] [PubMed]
- Gomi, R.; Sharma, A.; Wu, W.; Sung, B.; Worgall, S. Post-exposure immunization by capsid-modified AdC7 vector expressing Pseudomonas aeruginosa OprF clears P. aeruginosa respiratory infection. Vaccine 2017, 35, 7174–7180. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.R.; Laxova, A.; Farrell, P.M.; Barbieri, J.T. Proteomic identification of OprL as a seromarker for initial diagnosis of Pseudomonas aeruginosa infection of patients with cystic fibrosis. J. Clin. Microbiol. 2009, 47, 2483–2488. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.; Kyd, J.M.; Carzino, R.; Armstrong, D.; Grimwood, K.; Otczyk, D.C.; Cripps, A.W. Mucosal and systemic antibody responses to potential Pseudomonas aeruginosa vaccine protein antigens in young children with cystic fibrosis following colonization and infection. Hum. Vaccines Immunother. 2013, 9, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Liu, X. Prokaryotic expression, polyclonal antibody preparation and immunoprotection potential of Pseudomonas aeruginosa outer membrane protein OprH. Acad. J. Second Mil. Med. Univ. 2015, 36, 1092–1096. [Google Scholar] [CrossRef]
- Holder, I.A.; Neely, A.N.; Frank, D.W. PcrV immunization enhances survival of burned Pseudomonas aeruginosa-infected mice. Infect. Immun. 2001, 69, 5908–5910. [Google Scholar] [CrossRef]
- Bjorn, M.J.; Vasil, M.L.; Sadoff, J.C.; Iglewski, B.H. Incidence of exotoxin production by Pseudomonas species. Infect. Immun. 1977, 16, 362–366. [Google Scholar] [CrossRef]
- Pollack, M.; Taylor, N.S.; Callahan, L.T. Exotoxin production by clinical isolates of Pseudomonas aeruginosa. Infect. Immun. 1977, 15, 776–780. [Google Scholar] [CrossRef]
- Pollack, M.; Young, L.S. Protective activity of antibodies to exotoxin A and lipopolysaccharide at the onset of Pseudomonas aeruginosa septicemia in man. J. Clin. Investig. 1979, 63, 276–286. [Google Scholar] [CrossRef]
- Cross, A.S.; Sadoff, J.C.; Iglewski, B.H.; Sokol, P.A. Evidence for the role of toxin A in the pathogenesis of infection with Pseudomonas aeruginosa in humans. J. Infect. Dis. 1980, 142, 538–546. [Google Scholar] [CrossRef]
- Walker, H.L.; McLeod, C.G.; Leppla, S.H.; Mason, A.D. Evaluation of Pseudomonas aeruginosa toxin A in experimental rat burn wound sepsis. Infect. Immun. 1979, 25, 828–830. [Google Scholar] [CrossRef] [PubMed]
- Cryz, S.J.; Friedman, R.L.; Pavlovskis, O.R.; Iglewski, B.H. Effect of formalin toxoiding on Pseudomonas aeruginosa toxin A: Biological, chemical, and immunochemical studies. Infect. Immun. 1981, 32, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Pavlovskis, O.R.; Edman, D.C.; Leppla, S.H.; Wretlind, B.; Lewis, L.R.; Martin, K.E. Protection against experimental Pseudomonas aeruginosa infection in mice by active immunization with exotoxin A toxoids. Infect. Immun. 1981, 32, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Pollack, M.F.; Prescott, R.K. Toxoid from exotoxin A of Pseudomonas aeruginosa: Preparation and characterization. J. Infect. Dis. 1982, 145, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Marburg, S.; Tolman, R.L.; Callahan, L.T. Pseudomonas exotoxin A: Toxoid preparation by photoaffinity inactivation. Proc. Natl. Acad. Sci. USA 1983, 80, 2870–2873. [Google Scholar] [CrossRef] [PubMed]
- Cryz, S.J.; Iglewski, B.H. Toxoids of Pseudomonas aeruginosa toxin A: Production by chemical and genetic means. Rev. Infect. Dis. 1983, 5 (Suppl. 5), S992–S997. [Google Scholar] [CrossRef]
- Manafi, A.; Kohanteb, J.; Mehrabani, D.; Japoni, A.; Amini, M.; Naghmachi, M.; Zaghi, A.H.; Khalili, N. Active immunization using exotoxin A confers protection against Pseudomonas aeruginosa infection in a mouse burn model. BMC Microbiol. 2009, 9, 23. [Google Scholar] [CrossRef]
- Ueda, Y.; Homma, J.Y.; Abe, C. Effects of immunization of horses with common antigen (OEP), protease toxoid, and elastase toxoid on corneal ulceration due to Pseudomonas aeruginosa infection. Jpn. J. Vet. Sci. 1982, 44, 289–300. [Google Scholar] [CrossRef]
- Homma, J.Y.; Abe, C.; Yanagawa, R.; Noda, H. Effectiveness of immunization with multicomponent vaccines in protection against hemorrhagic pneumonia due to Pseudomonas aeruginosa infection in mink. Rev. Infect. Dis. 1983, 5 (Suppl. 5), S858–S866. [Google Scholar] [CrossRef]
- Matsumoto, T.; Tateda, K.; Furuya, N.; Miyazaki, S.; Ohno, A.; Ishii, Y.; Hirakata, Y.; Yamaguchi, K. Efficacies of alkaline protease, elastase and exotoxin A toxoid vaccines against gut-derived Pseudomonas aeruginosa sepsis in mice. J. Med. Microbiol. 1998, 47, 303–308. [Google Scholar] [CrossRef]
- Homma, J.Y.; Tanimoto, H. A multicomponent Pseudomonas aeruginosa vaccine consisting of toxoids of protease, elastase, exotoxin A and a common protective antigen (OEP). Application in patients with diffuse panbronchiolitis. Antibiot. Chemother. (1971) 1987, 39, 215–221. [Google Scholar] [CrossRef]
- Pozuelo, M.J.; Jiménez, P.A.; Valderrey, A.D.; Fernández-Olmos, A.A.; Cantón, R.; Rotger, R. Polimorfismo de los genes mucA y fpvA en Pseudomonas aeruginosa de pacientes con fibrosis quística: Coexistencia de variantes genéticas en el mismo paciente. Enferm. Infecc. Microbiol. Clin. 2011, 29, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, G.F.; Sartorio, R.; Lee, S.H.; Mee, J.M.; Altobell, L.J.; Kujawa, D.P.; Jeffries, E.; Clapham, B.; Meijler, M.M.; Janda, K.D. Antibody interference with N-acyl homoserine lactone-mediated bacterial quorum sensing. J. Am. Chem. Soc. 2006, 128, 2802–2803. [Google Scholar] [CrossRef] [PubMed]
- Freihorst, J.; Merrick, J.M.; Ogra, P.L. Effect of oral immunization with Pseudomonas aeruginosa on the development of specific antibacterial immunity in the lungs. Infect. Immun. 1989, 57, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, T.S.; Priebe, G.P.; Pier, G.B. A live-attenuated Pseudomonas aeruginosa vaccine elicits outer membrane protein-specific active and passive protection against corneal infection. Infect. Immun. 2006, 74, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Krause, A.; Worgall, S. Recent developments for Pseudomonas vaccines. Hum. Vaccines 2011, 7, 999–1011. [Google Scholar] [CrossRef] [PubMed]
- Denis-Mize, K.S.; Price Bm Fau-Baker, N.R.; Baker Nr Fau-Galloway, D.R.; Galloway, D.R. Analysis of immunization with DNA encoding Pseudomonas aeruginosa exotoxin A. FEMS Immunol. Med. Microbiol. 2000, 27, 147–154. [Google Scholar] [CrossRef]
- Shiau, J.W.; Tang, T.K.; Shih, Y.L.; Tai, C.; Sung, Y.Y.; Huang, J.L.; Yang, H.L. Mice immunized with DNA encoding a modified Pseudomonas aeruginosa exotoxin A develop protective immunity against exotoxin intoxication. Vaccine 2000, 19, 1106–1112. [Google Scholar] [CrossRef]
- Jiang, M.; Yao, J.; Feng, G. Protective effect of DNA vaccine encoding Pseudomonas exotoxin A and PcrV against acute pulmonary P. aeruginosa Infection. PLoS ONE 2014, 9, e96609. [Google Scholar] [CrossRef]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Christensen, D. Vaccine adjuvants: Why and how. Hum. Vaccines Immunother. 2016, 12, 2709–2711. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, I.; Garg, R.; van Drunen Littel-van den Hurk, S. Selection of adjuvants for vaccines targeting specific pathogens. Expert Rev. Vaccines 2019, 18, 505–521. [Google Scholar] [CrossRef] [PubMed]
- Apostólico, J.S.; Lunardelli, V.A.S.; Coirada, F.C.; Boscardin, S.B.; Rosa, D.S. Adjuvants: Classification, Modus Operandi, and Licensing. J. Immunol. Res. 2016, 2016, 1459394. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, L.E.; Baker, B.; Perricone, C.; Shoenfeld, Y. Vaccines, adjuvants and autoimmunity. Pharmacol. Res. 2015, 100, 190–209. [Google Scholar] [CrossRef] [PubMed]
- Vetter, V.; Denizer, G.; Friedland, L.R.; Krishnan, J.; Shapiro, M. Understanding modern-day vaccines: What you need to know. Ann. Med. 2018, 50, 110–120. [Google Scholar] [CrossRef]
- Ko, E.J.; Kang, S.M. Immunology and efficacy of MF59-adjuvanted vaccines. Hum. Vaccines Immunother. 2018, 14, 3041–3045. [Google Scholar] [CrossRef]
- He, P.; Zou, Y.; Hu, Z. Advances in aluminum hydroxide-based adjuvant research and its mechanism. Hum. Vaccines Immunother. 2015, 11, 477–488. [Google Scholar] [CrossRef]
- Jazani, N.H.; Parsania, S.; Sohrabpour, M.; Mazloomi, E.; Karimzad, M.; Shahabi, S. Naloxone and alum synergistically augment adjuvant activities of each other in a mouse vaccine model of Salmonella Typhimurium infection. Immunobiology 2011, 216, 744–751. [Google Scholar] [CrossRef]
- Ryu, J.I.; Park, S.A.; Wui, S.R.; Ko, A.; Han, J.E.; Choi, J.A.; Song, M.K.; Kim, K.S.; Cho, Y.J.; Lee, N.G. A de-O-acylated Lipooligosaccharide-Based Adjuvant System Promotes Antibody and Th1-Type Immune Responses to H1N1 Pandemic Influenza Vaccine in Mice. Biomed. Res. Int. 2016, 2016, 3713656. [Google Scholar] [CrossRef]
- Ryu, J.I.; Wui, S.R.; Ko, A.; Lee, Y.J.; Do, H.; Kim, H.J.; Rhee, I.M.; Park, S.A.; Kim, K.S.; Cho, Y.J.; et al. Increased Immunogenicity and Protective Efficacy of a P. aeruginosa Vaccine in Mice Using an Alum and De-O-Acylated Lipooligosaccharide Adjuvant System. J. Microbiol. Biotechnol. 2017, 27, 1539–1548. [Google Scholar] [CrossRef]
- Oberemok, V.V.; Laikova, K.V.; Yurchenko, K.A.; Marochkin, N.A.; Fomochkina, I.I.; Kubyshkin, A.A.-O. SARS-CoV-2 will constantly sweep its tracks: A vaccine containing CpG motifs in ‘lasso’ for the multi-faced virus. Inflamm. Res. 2020, 69, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, G.; Rappuoli, R.; Didierlaurent, A.M. Correlates of adjuvanticity: A review on adjuvants in licensed vaccines. Semin. Immunol. 2018, 39, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Żelechowska, P.; Różalska, S.; Wiktorska, M.; Brzezińska-Błaszczyk, E.; Agier, J. Curdlan stimulates tissue mast cells to synthesize pro-inflammatory mediators, generate ROS, and migrate via Dectin-1 receptor. Cell. Immunol. 2020, 351, 104079. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, H.S.; Wolf, A.J.; Underhill, D.M. Beta-glucan recognition by the innate immune system. Immunol. Rev. 2009, 230, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Clements, J.D.; Norton, E.B. The Mucosal Vaccine Adjuvant LT(R192G/L211A) or dmLT. mSphere 2018, 3. [Google Scholar] [CrossRef]
- Hoggarth, A.; Weaver, A.; Pu, Q.; Huang, T.; Schettler, J.; Chen, F.; Yuan, X.; Wu, M. Mechanistic research holds promise for bacterial vaccines and phage therapies for Pseudomonas aeruginosa. Drug Des. Devel. Ther. 2019, 13, 909–924. [Google Scholar] [CrossRef]
- Toth, A.; Schödel, F.; Duchêne, M.; Massarrat, K.; Blum, B.; Schmitt, A.; Domdey, H.; von Specht, B.U. Protection of immunosuppressed mice against translocation of Pseudomonas aeruginosa from the gut by oral immunization with recombinant Pseudomonas aeruginosa outer membrane protein I expressing Salmonella dublin. Vaccine 1994, 12, 1215–1221. [Google Scholar] [CrossRef]
- Arnold, H.; Bumann, D.; Felies, M.; Gewecke, B.; Sörensen, M.; Gessner, J.E.; Freihorst, J.; von Specht, B.U.; Baumann, U. Enhanced immunogenicity in the murine airway mucosa with an attenuated Salmonella live vaccine expressing OprF-OprI from Pseudomonas aeruginosa. Infect. Immun. 2004, 72, 6546–6553. [Google Scholar] [CrossRef]
- Bumann, D.; Behre, C.; Behre, K.; Herz, S.; Gewecke, B.; Gessner, J.E.; von Specht, B.U.; Baumann, U. Systemic, nasal and oral live vaccines against Pseudomonas aeruginosa: A clinical trial of immunogenicity in lower airways of human volunteers. Vaccine 2010, 28, 707–713. [Google Scholar] [CrossRef]
- Zhang, M.; Sun, C.; Gu, J.; Yan, X.; Wang, B.; Cui, Z.; Sun, X.; Tong, C.; Feng, X.; Lei, L.; et al. Salmonella Typhimurium strain expressing OprF-OprI protects mice against fatal infection by Pseudomonas aeruginosa. Microbiol. Immunol. 2015, 59, 533–544. [Google Scholar] [CrossRef]
- Aguilera-Herce, J.; García-Quintanilla, M.; Romero-Flores, R.; McConnell, M.J.; Ramos-Morales, F. A Live Salmonella Vaccine Delivering PcrV through the Type III Secretion System Protects against Pseudomonas aeruginosa. mSphere 2019, 4, e00116-19. [Google Scholar] [CrossRef] [PubMed]
- Pier, G.B.; Meluleni, G.; Goldberg, J.B. Clearance of Pseudomonas aeruginosa from the murine gastrointestinal tract is effectively mediated by O-antigen-specific circulating antibodies. Infect. Immun. 1995, 63, 2818–2825. [Google Scholar] [CrossRef] [PubMed]
- DiGiandomenico, A.; Rao, J.; Goldberg, J.B. Oral vaccination of BALB/c mice with Salmonella enterica serovar Typhimurium expressing Pseudomonas aeruginosa O antigen promotes increased survival in an acute fatal pneumonia model. Infect. Immun. 2004, 72, 7012–7021. [Google Scholar] [CrossRef] [PubMed]
- DiGiandomenico, A.; Rao, J.; Harcher, K.; Zaidi, T.S.; Gardner, J.; Neely, A.N.; Pier, G.B.; Goldberg, J.B. Intranasal immunization with heterologously expressed polysaccharide protects against multiple Pseudomonas aeruginosa infections. Proc. Natl. Acad. Sci. USA 2007, 104, 4624–4629. [Google Scholar] [CrossRef] [PubMed]
- Bridge, D.R.; Whitmire, J.M.; Makobongo, M.O.; Merrell, D.S. Heterologous Pseudomonas aeruginosa O-antigen delivery using a Salmonella enterica serovar Typhimurium wecA mutant strain. Int. J. Med. Microbiol. 2016, 306, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.M.; Kobe, B.N.; Schmitt, D.M.; Phair, B.; Gilson, T.; Jung, J.Y.; Roberts, L.; Liao, J.; Camerlengo, C.; Chang, B.; et al. Genetic engineering of Francisella tularensis LVS for use as a novel live vaccine platform against Pseudomonas aeruginosa infections. Bioengineered 2015, 6, 82–88. [Google Scholar] [CrossRef]
- Spicer, E.J.; Goldenthal, E.I.; Ikeda, T. A toxicological assessment of Curdlan. Food Chem. Toxicol. 1999, 37, 455–479. [Google Scholar] [CrossRef]
- Molavi, F.; Barzegar-Jalali, M.; Hamishehkar, H. Polyester based polymeric nano and microparticles for pharmaceutical purposes: A review on formulation approaches. J. Control Release 2020, 320, 265–282. [Google Scholar] [CrossRef]
- Jia, Q.; Horwitz, M.A. Live attenuated tularemia vaccines for protection against respiratory challenge with virulent F. tularensis subsp. tularensis. Front. Cell. Infect. Microbiol. 2018, 8, 154. [Google Scholar] [CrossRef]
- Stanaway, J.D.; Atuhebwe, P.L.; Luby, S.P.; Crump, J.A. Assessing the Feasibility of Typhoid Elimination. Clin. Infect. Dis. 2020, 71, S179–S184. [Google Scholar] [CrossRef]
- Miquel-Clopés, A.; Bentley, E.G.; Stewart, J.P.; Carding, S.R. Mucosal vaccines and technology. Clin. Exp. Immunol. 2019, 196, 205–214. [Google Scholar] [CrossRef] [PubMed]




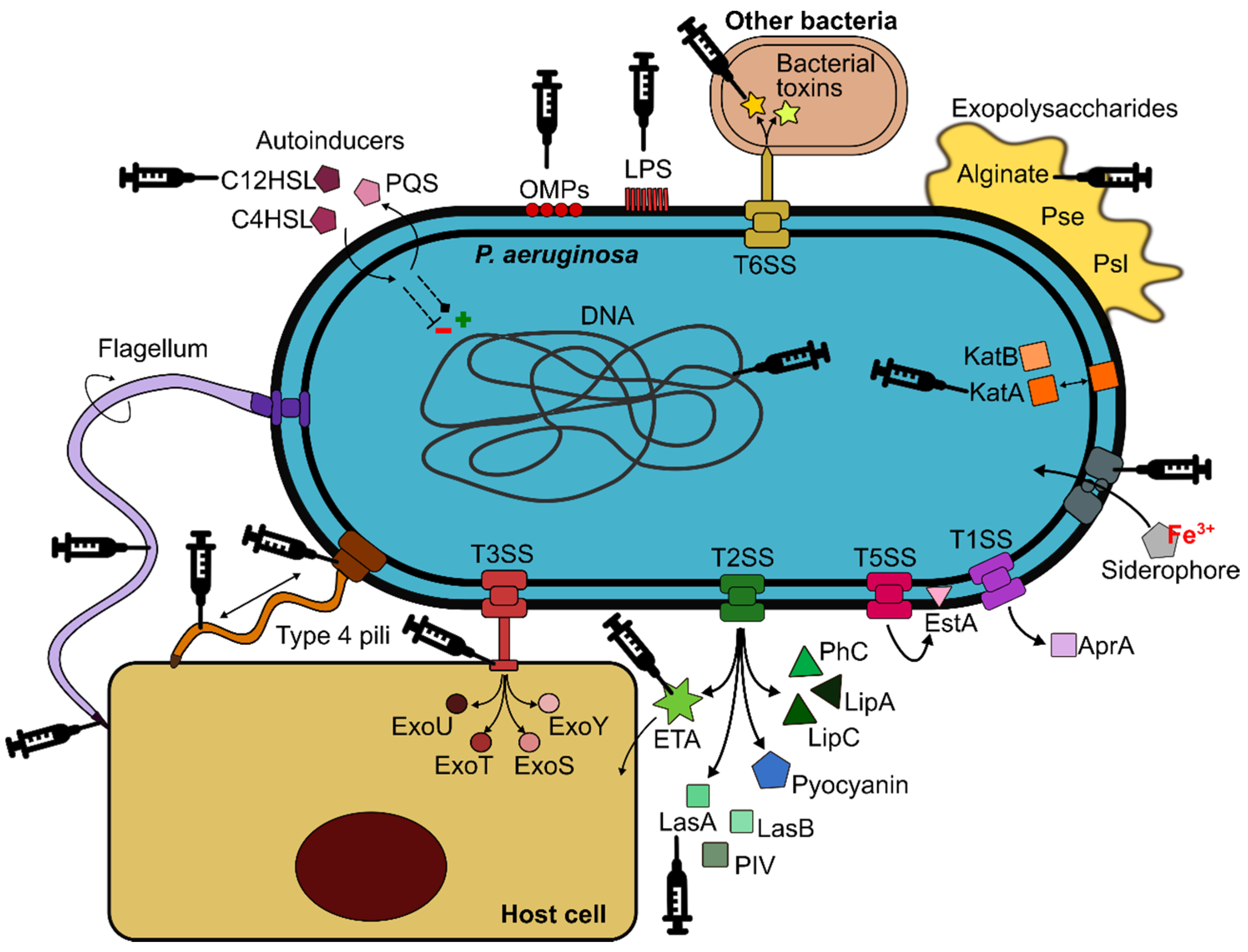{kind=link}
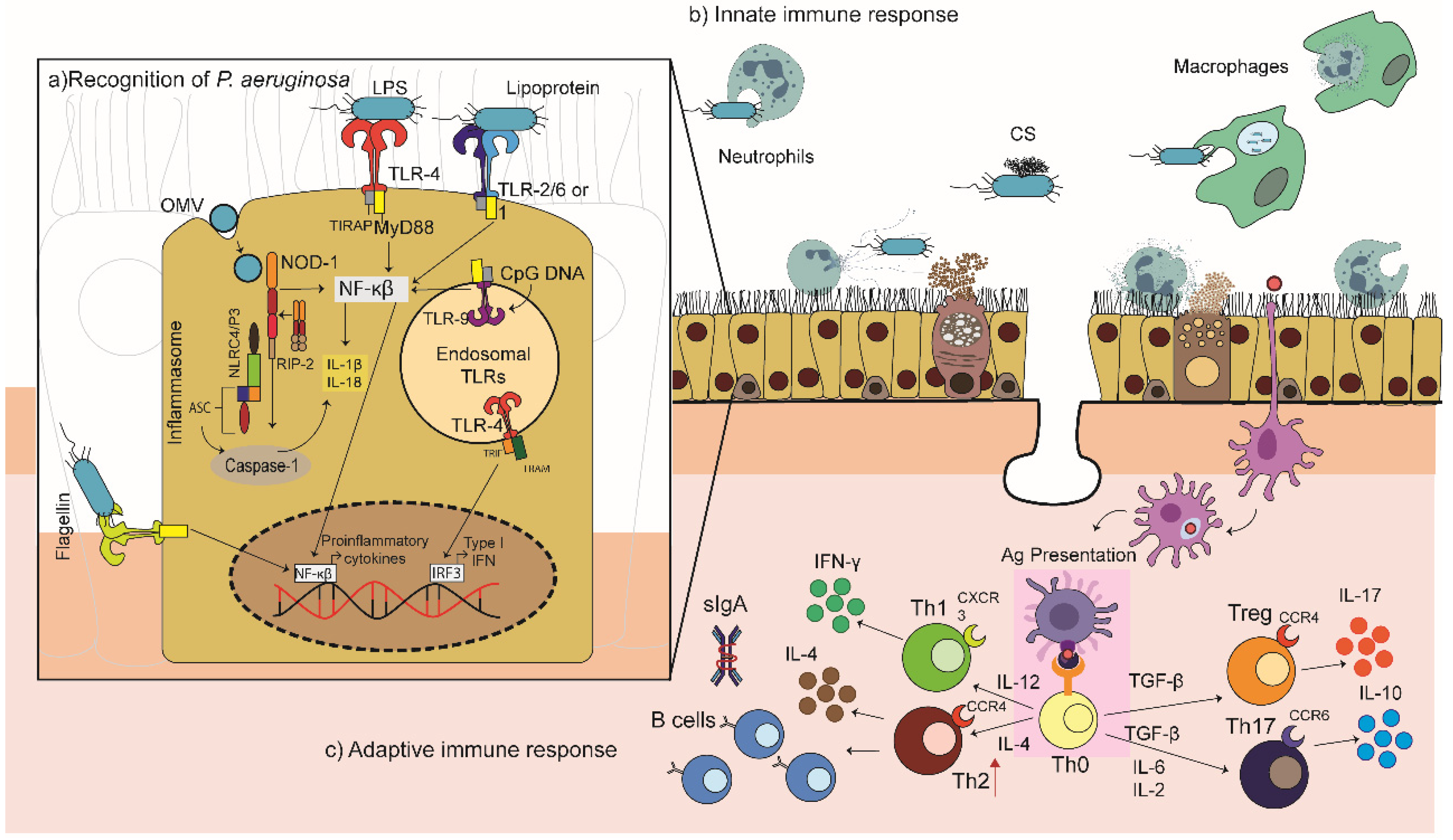{kind=link}
{kind=link}
| Ag | Composition | Animal Model | Strains | Adjuvant | Administration Route; Dose | % Survival and Time *; Immune Response | Ref. |
|---|---|---|---|---|---|---|---|
| Lipopolysaccharide | LPS extracts from 16 strains (PEV-01) | Mice | OPS 1-16 | None | IP; NS | NS | [112] |
| Octavalent OPS-ETA conjugate (Aerugen®) | Rabbits, mice | ATCC 27316, PA53 | AI(OH)3, CT, CpG1826 | IM, IN; 2–50 µg | Neutralising, opsonic IgG, INF-γ, TNF-α | [113,114,115] | |
| Liposomes with core LPS | Rabbits | PAC608 | AI(OH)3 | IM; NS | 70–88%, 48 h; cross-reactive Ab | [116] | |
| Alginate | MEP extracts | Rats, mice, GP | Various | Muramyl dipeptide | IV, IM; 10–100 µg | 33–50%, 4 days; opsonic Ab, IC | [117,118,119] |
| MEP-ETA conjugate | Mice, rabbits | 3064, PA103 | AI(OH)3 | IM; 10–50 µg | Anti-ETA opsonic Ab, anti-MEP IgG | [120] | |
| MEP-KHL conjugate | Mice, rabbits | 2129 | None, Freund’s | SC; 10 µg | Opsonic IgG | [121] | |
| Flagellum | Monovalent FliC | Mice | Various | None | IM; 7–10 µg | 40–100%, 5–10 days; | [122,123] |
| Whole purified flagella | Mice | Various | None | IN; 2 µg | 83,3%, 125 h; protective Ab | [124] | |
| r-flagellins | Mice | PAK, PAO1, BI1, BI2 | Alum, Montanide ISA 70 | SC, ID; 10–20 µg | 25–90%, 7 days; IgG1, IL-4, INF-γ, IL-17, IL-12, IL-10 | [125,126,127,128] | |
| FliC-MEP conjugate | Mice, rabbits | Various | Specol, Freund’s | SC, IN; 3–10 µg | 30–85%, 5 days; opsonic IgG | [129,130] | |
| r-FliC-ETA chimeric protein | Mice | PAO1, 8821M | Freund’s | SC; 20 µg | 80%, 10 days; IgG | [131] | |
| r-FliC-OMP chimeric protein | Mice, monkeys | PAK, PAO1, DM125, DM126 | Freund’s, none | IM, SC; 1–50 µg | 20–80%, 168 h; IgG, C3 complement | [132,133,134] | |
| Type 4 pili | PilA C-terminus | Mice | PAK | Adjuvax™ | IM; 50 µg | IgG1, IgG2 | [135] |
| Whole PilA | Mice | PAO1, clinical isolate | Freund’s, alum, naloxone | IT, SC; 0.004–5 µg | 20–100% 72 h–7 days; IgA, IgG, IL-17 | [136,137] | |
| PilA-ETA chimeric | Rabbits, mice | Various | Freund’s | SC, IN; 1–200 µg | Serum IgG, sIgA | [138,139] | |
| r- PilA and b-FliC | Mice | Various | None | SC; 15–20 µg | 92–100%, 7 days; Th2 response, IgGs, IL-4, INF, IL-17 | [140,141] | |
| PilA-PilQ chimeric | Mice | PAO1 | Alum | SC; 10 µg | Opsonic IgG | [142,143] | |
| PilA-OPS conjugate | Mice | 1244 | None | IN, SC; 5 µg | 40–65%, 225 h; anti-LPS IgG, IgM | [144] | |
| Consensus peptide | Mice | PAK, PAO1, KB7, P1 | Adjuvax™ | IP; 25–50 µg | 20–40%, 48 h; cross-reactive Ab | [145,146] | |
| Low mw fimbrial pili | Mice | PAO1 | Montanide ISA 70 | IP; 50 µg | Th1/Th2 response | [147] | |
| Outer membrane proteins | r-OprI | Mice | Various | AI(OH)3 | IP; 25–27 µg | 85–95.8%, 7–10 days; Ab | [148,149] |
| r-OprF | Mice | Various | None | IM; 10 µg | 36–92%, 10 days; Ab | [150] | |
| His-OprF-OprI protein (systemic formulation, IC43) | Mice | ATCC 33348, PAO1 | AI(OH)3 | IM; 20–100 µg | 80% 7 days; IgG1, IL-3 | [151,152] | |
| His-OprF-OprI protein (mucosal formulation) | Mice | ATCC 27853 | AI(OH)3 | IN; 30 µg | 75%, 7 days; IgG, IgM, IgA | [153] | |
| OprF N-terminus | Mice | ATCC 9027 | BCG, alum | SC; 12 µg | 50%, 48 h; IgG1, IgG2a | [154] | |
| OprF-OprI-ETA chimeric protein | Mice | ATCC 15692, 33349, 29260 | AlPO4 | IP | 60–80%, 10 days; neutralising Ab | [155] | |
| OprF, ETA and LasB | Rats | PAO1 | AI(OH)3 | IM; 25 µg | IgG | [156] | |
| OprF-OprI-PcrV chimeric protein | Mice | PAO1, PAK | Freund’s | SC; 10 µg | 75%, 10 days; IgG | [157] | |
| OprI-OprF-AlgE PHA nanovaccine | Mice | PAO1 | Alum | SC; 20 µg | Th1 response, IFN-γ, IgG2c | [158] | |
| r-OprL | Mice | Various | Freund’s, Curdlan | SC, IN; 50 µg | 60–100%, 7 days; serum IgG, Th17 response | [159,160] | |
| r-OprH | Mice | PAO1, PA14, PA103 | Curdlan | IN; 10 mg/mL | 25–40%, 5 days; IgG | [161] | |
| r-OMP (PA5340, PA3526 and MotY) | Mice | PAO1 | Alum | IP; 10 µg | 20–50%, 5 days; NT | [6] | |
| Type III secretion system | r-PcrV | Mice | PA103, PAO1, PAK | Freund’s, FCA-FIA, alum, CpG ODN | SC, IP, IN; 10 µg | 29–90%, 1–7 days; IgG1, IgG2, IgA | [93,162,163,164] |
| PcrV immunogenic epitopes | Mice | Various | Freund’s, | IM; 30 µg | 50–65% 2–7 days; IgG, IL-17A, IFN-γ | [165,166] | |
| PcrV-C12HSL conjugate | Mice | PAO1 | Freund’s | SC; 10–20 µg | 75%, 14 days; IgG, IgM | [167] | |
| r-PcrV-OprI-Hcp1 chimeric protein | Mice | PAO1 | Freund’s, MF59, Al(OH)3, AlPO4, AS04 | IM; 30 µg | 80%, 7 days; IgG, IL-4, IFN-γ, IL-17A | [168] | |
| r-PopB-PcrH | Mice | PAO1 | Curdlan | IN; 35 µg | 62,5%, 5 days; IL-17 | [92] | |
| PLGA encapsulated PopB-PcrH | Mice | PAO1 | Curdlan | IN; 20 µg | 70%, 6 days; IgG, Th17 response | [94] | |
| Exotoxin A | r-ETA toxoid | Mice | Clinical mucoid strain | Freund’s, AlPO4 | SC, IP; 1–20 µg | 40–100%, 7 days; neutralising Ab | [169,170,171] |
| PLGA encapsulated ETA | Mice | PAO1 | None | IM; 100 µg | IgG, TNF-α, INF-γ, IL-4, IL-17 | [172] | |
| Elastase | LasB toxoid | Mice | IFO 3455 | None | SC; 10 µg | NS | [173] |
| LasB peptides-KLH/TT conjugate | Rats | PAO1, PAO2 | Freund’s | IM, SC; 30 µg | IgG, IgA | [174] | |
| Catalases | Purified KatA and KatE | Rats | 385, 423 | Freund’s | IPP; 10 µg | IgG, IgM, IgA, phagocytic cells | [175,176,177] |
| Auto-inducers | Hapten C12HSL-BSA conjugate | Mice | PAO1 | Freund’s | SC; NS | 36%, 7 days; serum Ab | [178] |
| Iron uptake proteins | r-FpvA, FoxA and HasR | Mice | PAO1, PA14, PA103 | Curdlan | 20 mg/mL | 10–35%, 5 days; NS | [161] |
| FpvA peptide-KLH conjugate | Mice | PAO1 | Curdlan | IN; 35 µg | IgA, IgG, IgM, IL-17 | [179] | |
| r-HitA | Mice | ATCC 9027 | BCG, Freund’s, IFA | SC or IM; 15 µg | IgG | [180] | |
| Whole-cell killed | Killed PA (Pseudostat®) | Rats | CF385 | Freund’s | IPP, SC, IT; 1010 CFU/mL | PMN cells, IgA, IgG, IgM, TNF-α | [181,182,183] |
| H2O2-deactivated PA | Mice | PAO1 | None | 3 × 107 CFU/mL | 100%, 6 days; IgG, IgM | [184] | |
| KBMA+IPTG, OSTAB pEi strain | Mice | CHA | None | SC; 1–2 × 108 CFU/mL | 58.3%, 150h; Ab, IL-17, Th17 cells, | [185] | |
| Live-attenuated bacteria | PAO1ΔaroA and PA14ΔaroA | Mice, rabbits | Clinical isolates and lab strains | None | IN; 1 × 108–2 × 109 CFU/mL | 40–100%, 7 days; IgG, neutrophils, IL-17 | [186,187,188] |
| O2/O5, O6, O10, and O11 serogroups aroA mutants | Mice | Various | None | IN; 1 × 108–1 × 109 CFU/mL | 10–35%, 7 days; opsonic Ab, lung CD4 T cells | [189] | |
| PAO1ΔmurI | Mice | PAO1, PA14, ST235 | None | IN, IM; 3 × 107–2 × 108 CFU/mL | 29–100%, 15 h; IgM, IgG, CD4+ T-cells | [190] | |
| DNA vaccines | Type-a/-b flagellin | Mice | PAO1, PAK | None | IM; 50 µg | 20–90%, 10 days; IgG | [191] |
| OMPs | Mice, chickens | Various | None, Al(OH)3 | IM, IP; 1–200 µg | 40–93.3%, 8 days-6 weeks; opsonic IgG1, IFN-γ, IL-2 | [192,193,194,195] | |
| OprF2-VP22 | Mice | PAO1 | None | IM; 20 µg | 0–40%, 10 days; IgG1, IgG2a | [196] | |
| PilA, OprF-OprI and PcrV | Mice | PAO1, PAK, D4 | None | IM, 100 µg | 30–100%, 10 days; IgG | [197] |
| Ag | Composition | Phase | No. of Patients (Type) | Administration Route; Dose | Adjuvant | Outcomes | Ref |
|---|---|---|---|---|---|---|---|
| Lipopolysaccharide | LPS extracts from seven strains (Pseudogen®) | II | 12 (CF, chronic) | IM; 6–12 µg/Kg | None | Lack of clinical benefit, toxicity in 92% of patients | [198,199] |
| LPS extracts from 16 strains (PEV-01) | I | 15 | SC; 0.5 mL | None | Appearance of protective antibodies | [200] | |
| II | 34 (CF, non-PA colonisation) | SC; 0.25–0.5 mL | None | No reduction in colonisation, rapid clinical deterioration | [201] | ||
| Octavalent OPS-ETA conjugate (Aerugen®) | I | 20 | SC; 0.5 mL (162.5 µg) | None | Safe, neutralising anti-ETA and opsonising anti-LPS IgG | [202,203] | |
| II | 26 (CF paediatric) | IM; 6–12 µg/Kg | None | Only 35% colonisation, better lung preservation | [204] | ||
| III | 476 (CF) | NS | NS | No clinical difference between groups | [205] | ||
| Alginate | Purified MEP extracts | I | 28 | IM, SC; 0.5 mL | None | Poor production of opsonic Ab | [206] |
| Flagellum | Monovalent FliC | I | 220 | IM; 40 µg | Al(OH)3 | IgG and IgA in serum and SIS | [207] |
| II | 10 | IM; 40 µg | None | [208] | |||
| Bivalent FliC | III | 483 (CF non-PA infected) | IM; 40 µg | Al(OH)3, thiomersal | High serum IgG titres to flagella vaccine subtypes, 34% protection | [209] | |
| Outer membrane proteins | OMP extractions from PA strains (CFC-101) | I | 48 | SC, IM; 0.1–1 mg | None | Safe, Immunogenic, TNF-α | [210] |
| II | 48 (burn) | IM; 0.5–1.0 mg | None | No clinical benefit | [211] | ||
| II | 95 (burn) | IM; 0.5–1 mg | None | Adverse reactions | [212] | ||
| r-OprI | I | 28 | IM; 20–500 µg | None | High variability among volunteers | [213] | |
| His-OprF-OprI protein (systemic formulation, IC43) | I | 32 | IM; 20–500 µg | Al(OH)3 | Increased IgG1 titres | [214] | |
| I | 8 (burn) | IM; 100 µg | Al(OH)3 | Well tolerated, increased Ab titres | [215] | ||
| I | 163 | IM; 50–200 µg | Al(OH)3 | Safe, specific IgG response | [216] | ||
| II | 137 (ventilated ICU) | IM; 100–200 µg | Al(OH)3 | IgG response, lower mortality but higher infection in vaccinated group | [217] | ||
| II-III | 799 (medically ill ICU) | IM; 100 µg | None | No difference in mortality and survival rates | [9] | ||
| His-OprF-OprI protein (mucosal formulation, SDS-emugel) | I | 8 | IN; 500 µg | None | Safe, increased IgG and IgA in 6 subjects over 3 months, inter-subject variability | [218] | |
| I-II | 12 | IN, systemic; 1 mg | None | Lasting immunity (IgG and IgA) in serum and airways | [219] | ||
| Whole-cell killed | Killed PA (Pseudostat®) | I | 9 (bronchiectasis) | Oral, NS | NS | Safe | [220] |
| I | 30 | Oral, 150 mg | None | Opsonic Ab, macrophage killing, few adverse events | [221] |
| Adjuvant | Class | Mechanism or Receptor | IR against PA | Use in Humans | Ref |
|---|---|---|---|---|---|
| CpG ODN | IMS | TLR9 | Ab, Th1, CD8 + T cells | Used in the human HBV vaccine (HEPLISAV-B). | [163,313,324,325] |
| Aluminium salts | B | NALP3, ITAM, Ag delivery | Ab, Th2 | Several licensed vaccines (HAB, HBV, DTP, HiB). | [217,313,314,316] |
| Curdlan | DS | Dectin-1; DC activation directs the differentiation of effector Th17 cells | Ab, Th17 | FDA approved (1996) for use as a stabiliser or texturiser in foods. No evidence of any toxicity | [179,326,340] |
| PLGA | DS | Ag delivery | Ab, Th17 | Registered safe by the U.S. FDA for clinical use. | [94,341] |
| dmLT | B | Stimulates IL-17 secretion and increases the transport of secretory IgA (sIgA) | Ab, Th1, Th17 | Phase I and II of ETEC and EVAX vaccines (no adverse events reported). | [95,328] |
| AS04 | B | TLR4 | Ab, Th2 | Used as part of a HBV vaccine (Fendrix, GSK) and an HPV vaccine (Cervarix, GSK). | [325] |
| MF59 | DS | Immune cell recruitment | Ab, Th2 | Currently licensed as part of a flu vaccine (Fluad™, Seqirus) | [316,319,325] |
| AdC7 vector | DS | Ag delivery to mucosal sites | Ab, Th1 | Preclinical studies in mice against P. aeruginosa, Ebola virus and malaria in mice models. | [284,341] |
| CIA06 (alum + dLos) | IMS | TLR4 agonist derived | Ab, Th1, Th17, Th2 | Preclinical studies in mice on the H1N1 pandemic influenza vaccine Greenflu-S®. | [323] |
| NLX + alum | IMS | Opioid receptor antagonists | Ab, Th1, Th17, Th2 | Approved by the FDA as a prescription drug, used to reverse opioid-induced respiratory depression. | [137] |
| Live-attenuated bacteria (S. Typhimurium, F. tularensis LVS) | DS | Ag delivery to mucosal sites | Ab, Th2, Th1 (biased) | Retains toxicity in humans and animals but since 2008, oral live-attenuated Ty21a (S. Typhi) vaccines have been recommended for typhoid control. | [342,343] |
| Ag/Adj | Route | P. aeruginosa Strains | Immunisation (N°x dose, Ag, Adj) | TR | Ref |
|---|---|---|---|---|---|
| PopB+PcrH/ Curdlan | IN | ExoU+ PAO1 and attenuated PAO1ΔaroA (C+) (7 × 105 CFU/mouse) | 3 × 35 µg/dose, 10 mg/mL | Th17 | [92] |
| PopB+PcrH/Curdlan | IN | ExoU+ PAO1 and live-attenuated PAO1ΔaroA (C+) (7 × 105 CFU/mouse) | 3 × 35 µg/dose, 10 mg/mL | Th17 | [94] |
| OMPs/dmLT | ID | PA01 (1.4 × 107 CFU/mouse) | 3 × 1 µg/mouse, 1 µg | Th1/17 | [95] |
| OMP/CIA06 | IM | GN-H3 and PA103 (10 LD50) | 2 × 5 µg/mouse, 0.5 µg dLOS + 25 µg of alum in 100 µL of PBS | Th2/1/17 | [323] |
| r-PilA/alum+NLX | SC | PA01 and a clinical isolate (3–5 × 106 CFU) | 3 × 5 µg/mouse, 6 mg/g body weight (NLX) and 200 mg of alum | Th2/Th1/Th17 | [137] |
| PcrV/ FA, alum, or CpG ODN | IP | PA103 exoS-, exoT+, exoU+ and exoY+ (1.5 × 106 CFU) | 3 × 10 µg/dose, 100µL/dose FCA/FIA, 100 µL/dose alum or 10µg/dose of CpG ODN | FA and CpgODN: Th1/Th2. Alum: Th2 | [93] |
| PcrV/CpG ODN | IN | PA103 strain (ATCC 29260, 1.5 × 106 CFU in 60µL) | 3 × 5 µg/mouse, 6 mg/g body weight and 200 mg of alum | Th2/Th1 | [163] |
| OprF/AdC7 (AdC7OprF) | IM, IT | PA encapsulated in agar beads (5 × 106 CFU, 50μL). | 1 × 109–1011 pu/animal | Th2/Th1 | [309] |
| SseJ-PcrV and SseJ-OprF-I/Attenauated S. Typhimurium SV9699 | IP | PAO1 (9 × 106 bacteria in 0.2 mL) | 1 × 2 × 105 CFU/animal | NS | [334] |
| PilAPa, OprFPa, FliCPa/Attenuated F. tularensis LVS | IN | PA1244 (PilAPa) and PA14 (OprFPa, and FliCPa) | 1 × ~100 CFU/animal | NS | [339] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sainz-Mejías, M.; Jurado-Martín, I.; McClean, S. Understanding Pseudomonas aeruginosa–Host Interactions: The Ongoing Quest for an Efficacious Vaccine. Cells 2020, 9, 2617. https://doi.org/10.3390/cells9122617
Sainz-Mejías M, Jurado-Martín I, McClean S. Understanding Pseudomonas aeruginosa–Host Interactions: The Ongoing Quest for an Efficacious Vaccine. Cells. 2020; 9(12):2617. https://doi.org/10.3390/cells9122617
Chicago/Turabian StyleSainz-Mejías, Maite, Irene Jurado-Martín, and Siobhán McClean. 2020. "Understanding Pseudomonas aeruginosa–Host Interactions: The Ongoing Quest for an Efficacious Vaccine" Cells 9, no. 12: 2617. https://doi.org/10.3390/cells9122617
APA StyleSainz-Mejías, M., Jurado-Martín, I., & McClean, S. (2020). Understanding Pseudomonas aeruginosa–Host Interactions: The Ongoing Quest for an Efficacious Vaccine. Cells, 9(12), 2617. https://doi.org/10.3390/cells9122617