Spontaneous Depolarization-Induced Action Potentials of ON-Starburst Amacrine Cells during Cholinergic and Glutamatergic Retinal Waves



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Immunohistochemistry
2.3. Patch-Clamp Electrophysiological Recording
2.4. Calcium Imaging
2.5. Simultaneous Ca2+ Imaging and Patch-Clamp Recording
2.6. Pharmacology
2.7. Data Analysis
3. Results
3.1. Characterization of Genetically Labeled SACs in Transgenic Mouse Lines
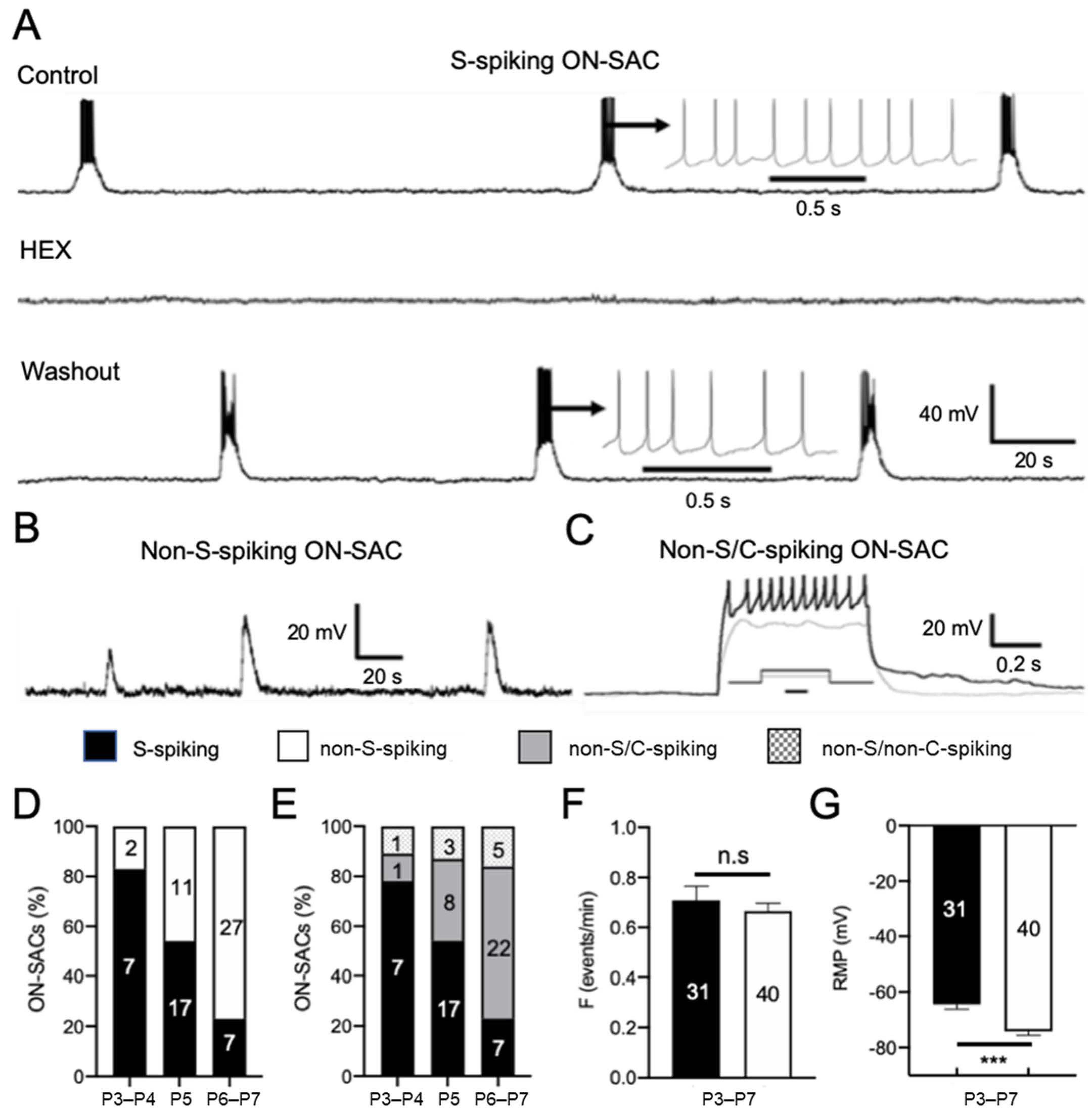
3.2. ON-SACs Exhibit Spontaneous Depolarization Accompanied by APs during Cholinergic Waves
3.3. OFF-SACs Show Spontaneous Depolarization without APs during Cholinergic Waves
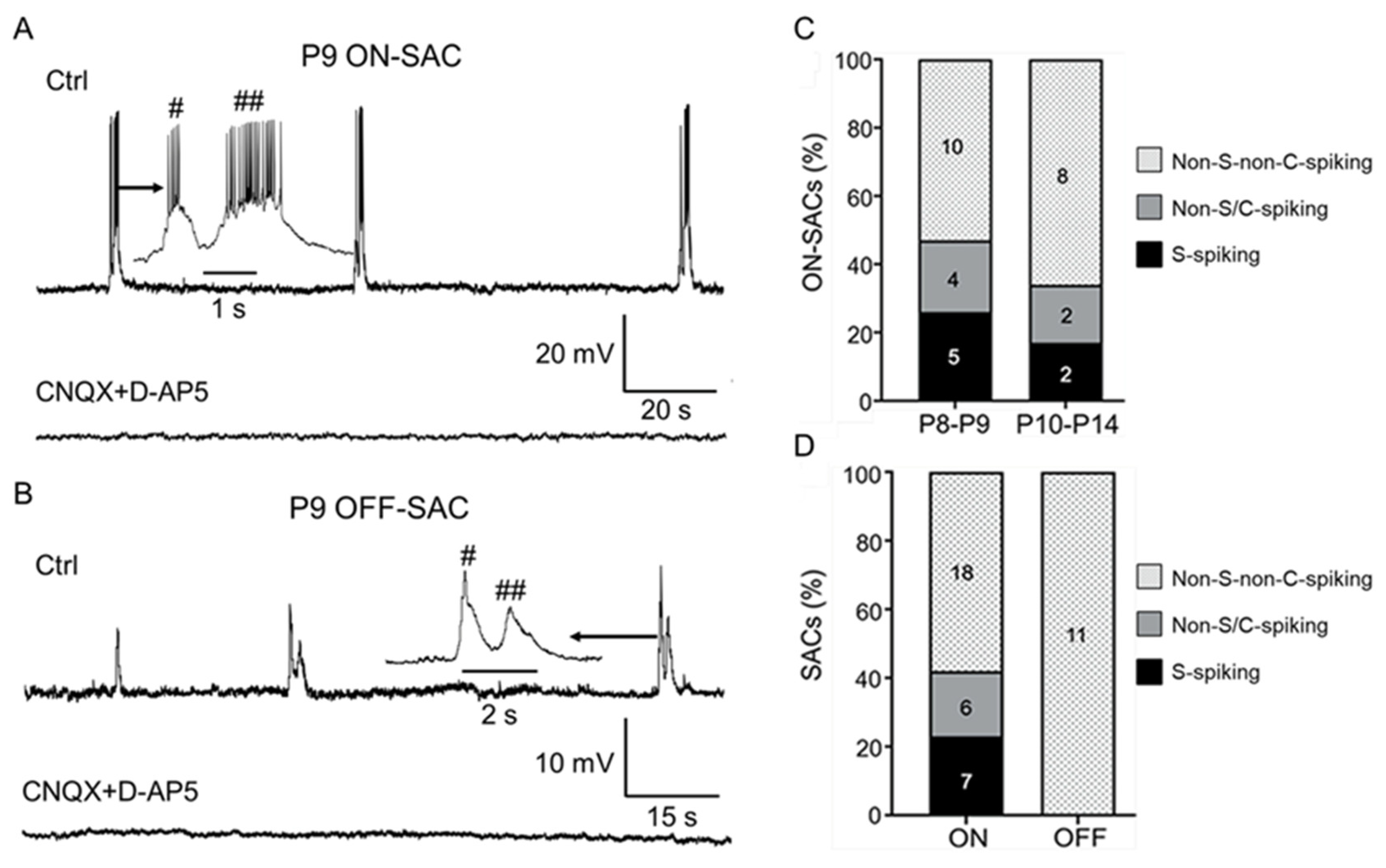
3.4. S-Spiking SACs still only Appear in the ON Layer during Glutamatergic Retinal Waves
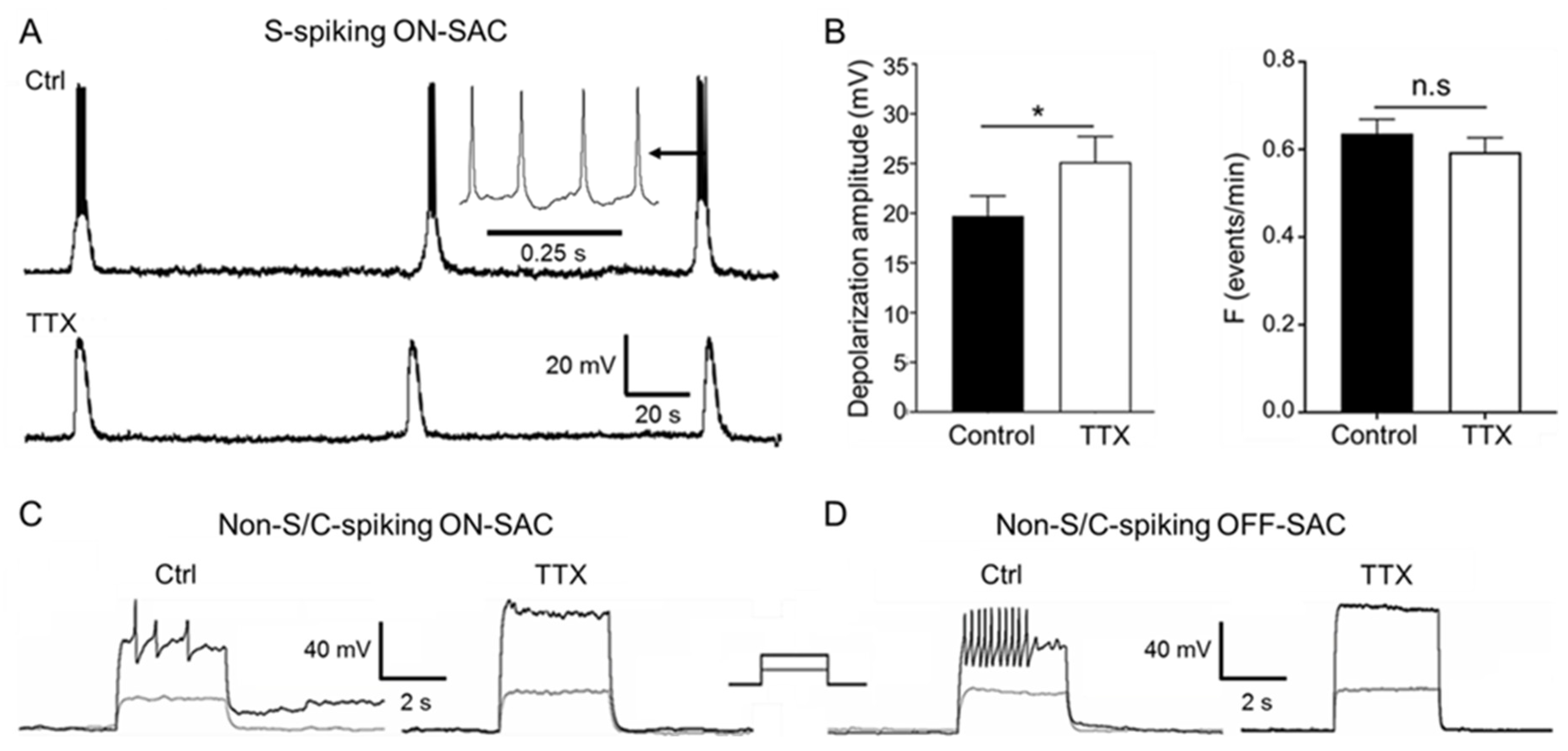
3.5. Blockage of Spontaneous and Current-Induced APs of SACs by TTX
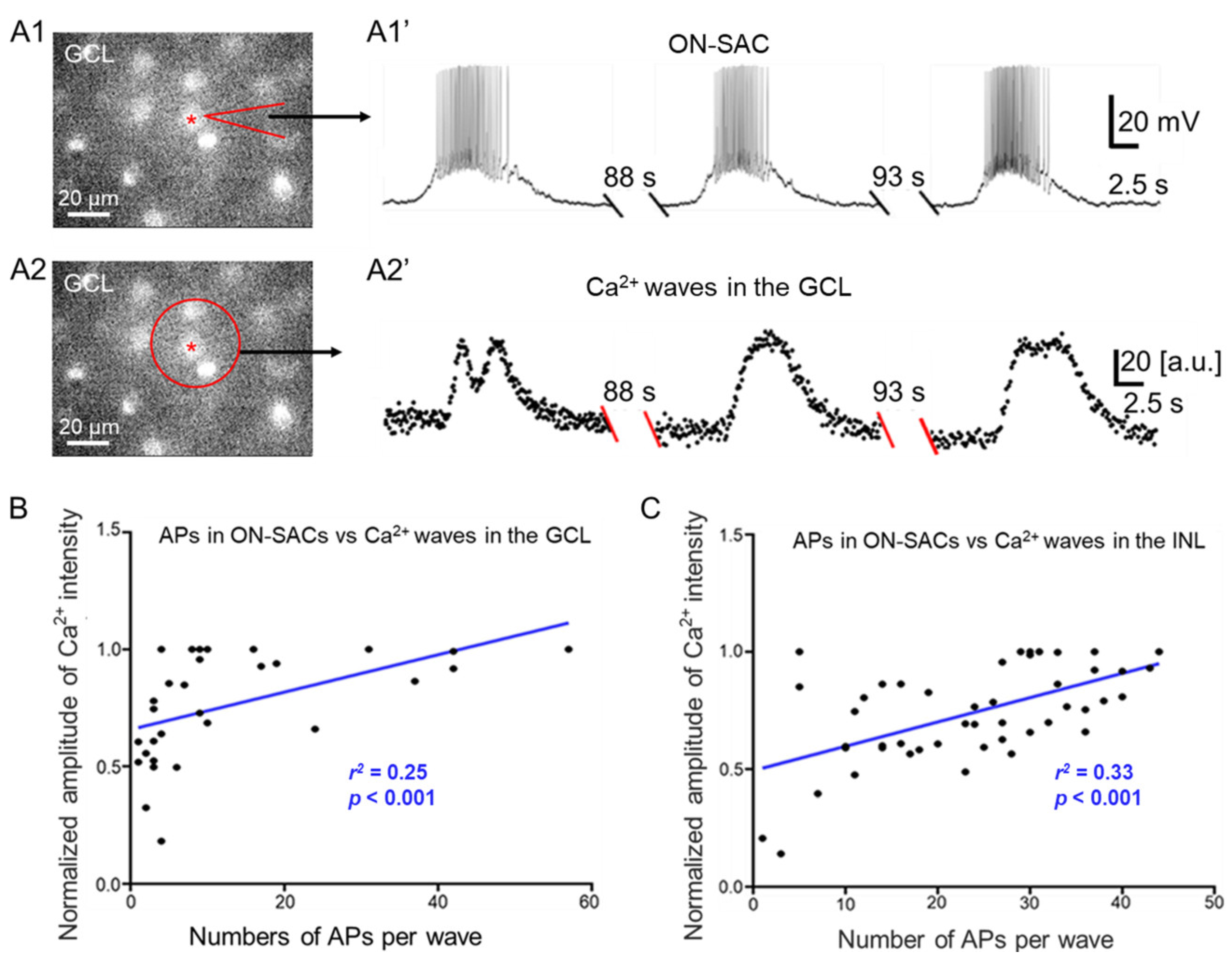
3.6. Relationships between APs of ON-SACs and Ca2+ Waves in the GCL and INL
4. Discussion
4.1. Distinct Electrical Properties of ON- and OFF-SACs during Cholinergic Waves
4.2. Properties and Mechanisms of Glutamatergic Waves in ON- and OFF-SACs
4.3. Possible Roles of ON-SAC’s APs in the Early Development of the Visual System
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Katz, L.C.; Shatz, C.J. Synaptic activity and the construction of cortical circuits. Science 1996, 274, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.O. Retinal waves and visual system development. Annu. Rev. Neurosci. 1999, 22, 29–47. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Lee, S.; Zhou, Z.J. A transient network of intrinsically bursting starburst cells underlies the generation of retinal waves. Nat. Neurosci. 2006, 9, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.P.; Burbridge, T.J.; Ye, M.; Chen, M.; Ge, X.; Zhou, Z.J.; Crair, M.C. Retinal Wave Patterns Are Governed by Mutual Excitation among Starburst Amacrine Cells and Drive the Refinement and Maintenance of Visual Circuits. J. Neurosci. 2016, 36, 3871–3886. [Google Scholar] [CrossRef]
- Kerschensteiner, D. Glutamatergic Retinal Waves. Front. Neural Circuits 2016, 10, 38. [Google Scholar] [CrossRef]
- Feller, M.B. Retinal waves are likely to instruct the formation of eye-specific retinogeniculate projections. Neural Dev. 2009, 4, 24. [Google Scholar] [CrossRef]
- Cang, J.; Renteria, R.C.; Kaneko, M.; Liu, X.; Copenhagen, D.R.; Stryker, M.P. Development of precise maps in visual cortex requires patterned spontaneous activity in the retina. Neuron 2005, 48, 797–809. [Google Scholar] [CrossRef]
- Arroyo, D.A.; Feller, M.B. Spatiotemporal Features of Retinal Waves Instruct the Wiring of the Visual Circuitry. Front. Neural Circuits 2016, 10, 54. [Google Scholar] [CrossRef]
- Firth, S.I.; Wang, C.T.; Feller, M.B. Retinal waves: Mechanisms and function in visual system development. Cell Calcium 2005, 37, 425–432. [Google Scholar] [CrossRef]
- Xu, H.P.; Furman, M.; Mineur, Y.S.; Chen, H.; King, S.L.; Zenisek, D.; Zhou, Z.J.; Butts, D.A.; Tian, N.; Picciotto, M.R.; et al. An instructive role for patterned spontaneous retinal activity in mouse visual map development. Neuron 2011, 70, 1115–1127. [Google Scholar] [CrossRef]
- Ackman, J.B.; Burbridge, T.J.; Crair, M.C. Retinal waves coordinate patterned activity throughout the developing visual system. Nature 2012, 490, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.J. Direct participation of starburst amacrine cells in spontaneous rhythmic activities in the developing mammalian retina. J. Neurosci. 1998, 18, 4155–4165. [Google Scholar] [CrossRef] [PubMed]
- Ford, K.J.; Felix, A.L.; Feller, M.B. Cellular mechanisms underlying spatiotemporal features of cholinergic retinal waves. J. Neurosci. 2012, 32, 850–863. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.J.; Lee, S.; Zhou, Z.J. A developmental switch in the excitability and function of the starburst network in the mammalian retina. Neuron 2004, 44, 851–864. [Google Scholar] [CrossRef]
- Ford, K.J.; Feller, M.B. Assembly and disassembly of a retinal cholinergic network. Vis. Neurosci. 2012, 29, 61–71. [Google Scholar] [CrossRef]
- Renna, J.M.; Chellappa, D.K.; Ross, C.L.; Stabio, M.E.; Berson, D.M. Melanopsin ganglion cells extend dendrites into the outer retina during early postnatal development. Dev. Neurobiol. 2015, 75, 935–946. [Google Scholar] [CrossRef]
- Zhou, Z.J.; Fain, G.L. Starburst amacrine cells change from spiking to nonspiking neurons during retinal development. Proc. Natl. Acad. Sci. USA 1996, 93, 8057–8062. [Google Scholar] [CrossRef]
- Whitney, I.E.; Keeley, P.W.; Raven, M.A.; Reese, B.E. Spatial patterning of cholinergic amacrine cells in the mouse retina. J. Comp. Neurol. 2008, 508, 1–12. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Z.; Wu, S.M. Development of cholinergic amacrine cells is visual activity-dependent in the postnatal mouse retina. J. Comp. Neurol. 2005, 484, 331–343. [Google Scholar] [CrossRef]
- Akrouh, A.; Kerschensteiner, D. Intersecting circuits generate precisely patterned retinal waves. Neuron 2013, 79, 322–334. [Google Scholar] [CrossRef]
- Kerschensteiner, D.; Wong, R.O. A precisely timed asynchronous pattern of ON and OFF retinal ganglion cell activity during propagation of retinal waves. Neuron 2008, 58, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Rossi, J.; Balthasar, N.; Olson, D.; Scott, M.; Berglund, E.; Lee, C.E.; Choi, M.J.; Lauzon, D.; Lowell, B.B.; Elmquist, J.K. Melanocortin-4 receptors expressed by cholinergic neurons regulate energy balance and glucose homeostasis. Cell Metab. 2011, 13, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Madisen, L.; Zwingman, T.A.; Sunkin, S.M.; Oh, S.W.; Zariwala, H.A.; Gu, H.; Ng, L.L.; Palmiter, R.D.; Hawrylycz, M.J.; Jones, A.R.; et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 2010, 13, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Madisen, L.; Garner, A.R.; Shimaoka, D.; Chuong, A.S.; Klapoetke, N.C.; Li, L.; van der Bourg, A.; Niino, Y.; Egolf, L.; Monetti, C.; et al. Transgenic mice for intersectional targeting of neural sensors and effectors with high specificity and performance. Neuron 2015, 85, 942–958. [Google Scholar] [CrossRef] [PubMed]
- Voinescu, P.E.; Kay, J.N.; Sanes, J.R. Birthdays of retinal amacrine cell subtypes are systematically related to their molecular identity and soma position. J. Comp. Neurol. 2009, 517, 737–750. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, A.G.; Ford, K.J.; Johnson, J.; Seal, R.P.; Edwards, R.H.; Copenhagen, D.R.; Feller, M.B. Synaptic and extrasynaptic factors governing glutamatergic retinal waves. Neuron 2009, 62, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Hausselt, S.E.; Euler, T.; Detwiler, P.B.; Denk, W. A dendrite-autonomous mechanism for direction selectivity in retinal starburst amacrine cells. PLoS Biol. 2007, 5, e185. [Google Scholar] [CrossRef]
- Taylor, W.R.; Wassle, H. Receptive field properties of starburst cholinergic amacrine cells in the rabbit retina. Eur. J. Neurosci. 1995, 7, 2308–2321. [Google Scholar] [CrossRef]
- Peters, B.N.; Masland, R.H. Responses to light of starburst amacrine cells. J. Neurophysiol. 1996, 75, 469–480. [Google Scholar] [CrossRef]
- Bloomfield, S.A. Relationship between receptive and dendritic field size of amacrine cells in the rabbit retina. J. Neurophysiol. 1992, 68, 711–725. [Google Scholar] [CrossRef]
- Jensen, R.J. Receptive-field properties of displaced starburst amacrine cells change following axotomy-induced degeneration of ganglion cells. Vis. Neurosci. 1995, 12, 177–184. [Google Scholar] [CrossRef]
- Cohen, E.D. Voltage-gated calcium and sodium currents of starburst amacrine cells in the rabbit retina. Vis. Neurosci. 2001, 18, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, M.; Ito, K.; Morishima, Y.; Shigematsu, Y.; Shimoda, Y. Characterization of voltage-gated ionic channels in cholinergic amacrine cells in the mouse retina. J. Neurophysiol. 2007, 97, 4225–4234. [Google Scholar] [CrossRef] [PubMed]
- Oesch, N.W.; Taylor, W.R. Tetrodotoxin-resistant sodium channels contribute to directional responses in starburst amacrine cells. PLoS ONE 2010, 5, e12447. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.J.; Cote, P.D.; Tremblay, F. Contribution of Nav1.8 sodium channels to retinal function. Neuroscience 2017, 340, 279–290. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, B.J.; Caldwell, J.H.; Ehring, G.R.; Bumsted O’Brien, K.M.; Luo, S.; Levinson, S.R. Tetrodotoxin-resistant voltage-gated sodium channels Na(v)1.8 and Na(v)1.9 are expressed in the retina. J. Comp. Neurol. 2008, 508, 940–951. [Google Scholar] [CrossRef]
- Jeon, C.J.; Strettoi, E.; Masland, R.H. The major cell populations of the mouse retina. J. Neurosci. 1998, 18, 8936–8946. [Google Scholar] [CrossRef]
- Brecha, N.; Johnson, D.; Peichl, L.; Wassle, H. Cholinergic amacrine cells of the rabbit retina contain glutamate decarboxylase and gamma-aminobutyrate immunoreactivity. Proc. Natl. Acad. Sci. USA 1988, 85, 6187–6191. [Google Scholar] [CrossRef]
- O’Malley, D.M.; Sandell, J.H.; Masland, R.H. Co-release of acetylcholine and GABA by the starburst amacrine cells. J. Neurosci. 1992, 12, 1394–1408. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, R.-S.; Yang, X.-L.; Zhong, Y.-M.; Zhang, D.-Q. Spontaneous Depolarization-Induced Action Potentials of ON-Starburst Amacrine Cells during Cholinergic and Glutamatergic Retinal Waves. Cells 2020, 9, 2574. https://doi.org/10.3390/cells9122574
Yan R-S, Yang X-L, Zhong Y-M, Zhang D-Q. Spontaneous Depolarization-Induced Action Potentials of ON-Starburst Amacrine Cells during Cholinergic and Glutamatergic Retinal Waves. Cells. 2020; 9(12):2574. https://doi.org/10.3390/cells9122574
Chicago/Turabian StyleYan, Rong-Shan, Xiong-Li Yang, Yong-Mei Zhong, and Dao-Qi Zhang. 2020. "Spontaneous Depolarization-Induced Action Potentials of ON-Starburst Amacrine Cells during Cholinergic and Glutamatergic Retinal Waves" Cells 9, no. 12: 2574. https://doi.org/10.3390/cells9122574
APA StyleYan, R.-S., Yang, X.-L., Zhong, Y.-M., & Zhang, D.-Q. (2020). Spontaneous Depolarization-Induced Action Potentials of ON-Starburst Amacrine Cells during Cholinergic and Glutamatergic Retinal Waves. Cells, 9(12), 2574. https://doi.org/10.3390/cells9122574