Neuropathic Itch


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
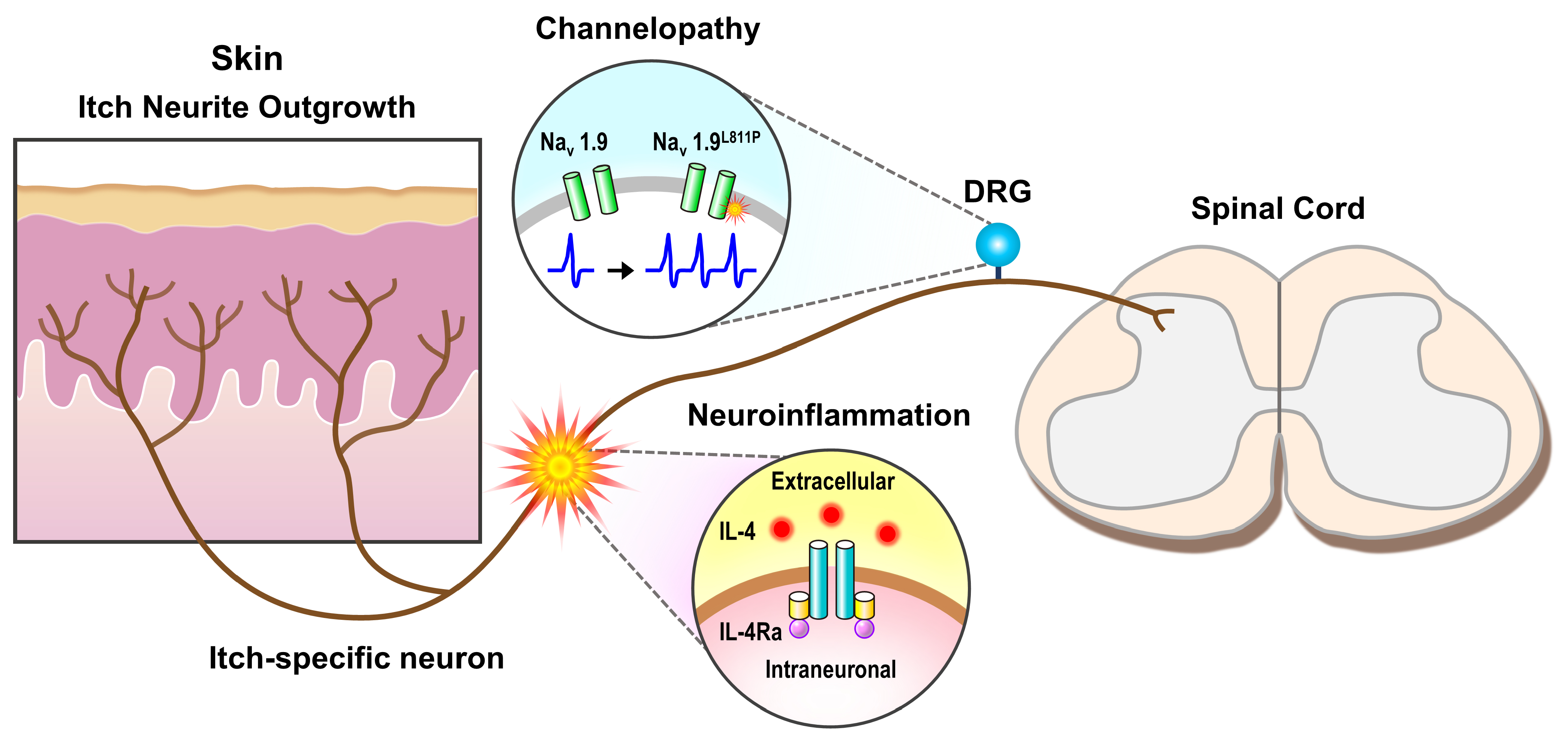
2. Neuropathic Itch and Inflammation
3. Neuropathic Itch and Channelopathies
4. Cellular Mechanisms of Neuropathic Itch
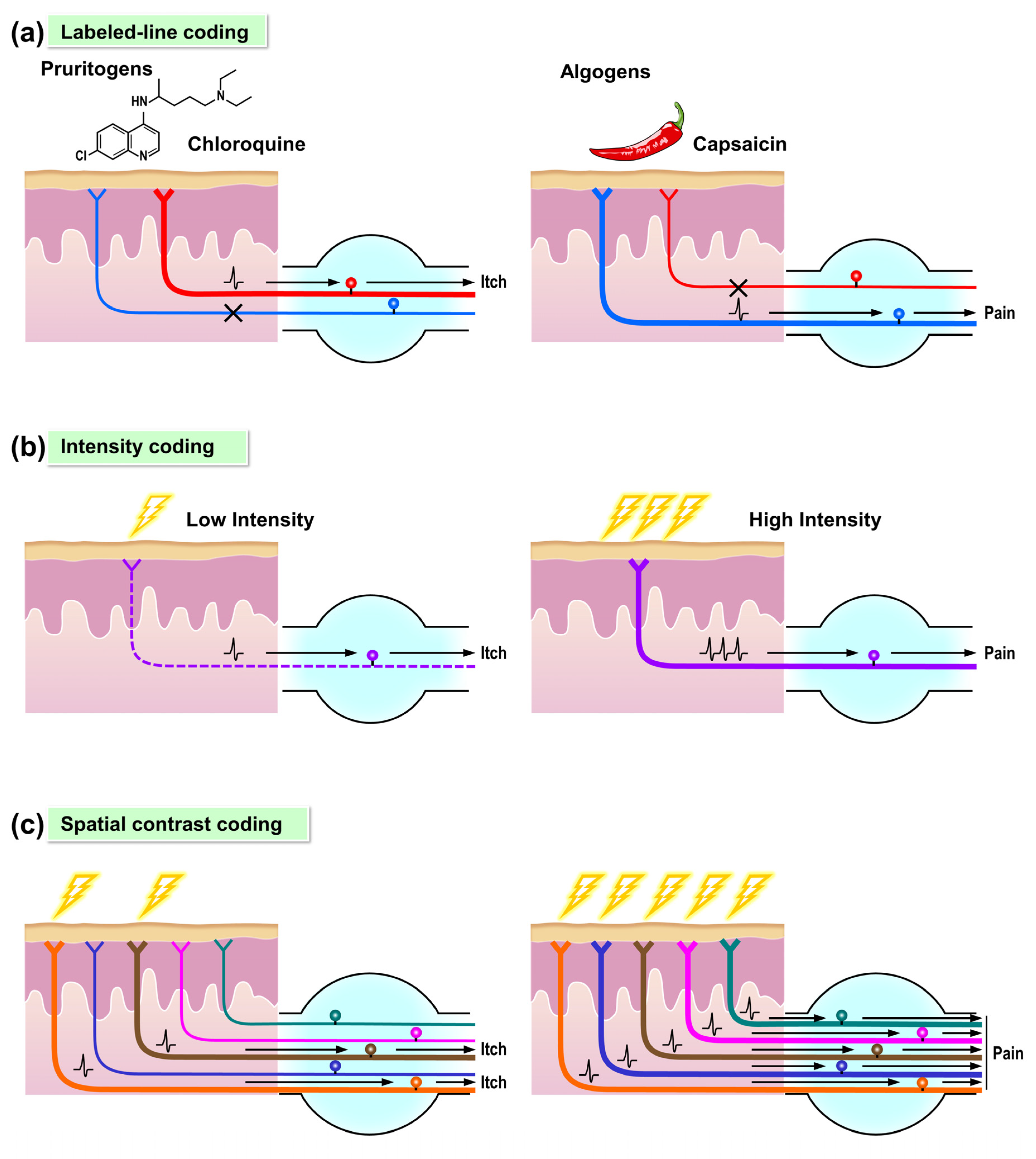
4.1. Molecular Data Support a Labeled-Line for Itch
4.2. Alternative Itch Coding Mechanisms
5. Neuropathic Itch of Peripheral Nerve Neuromas
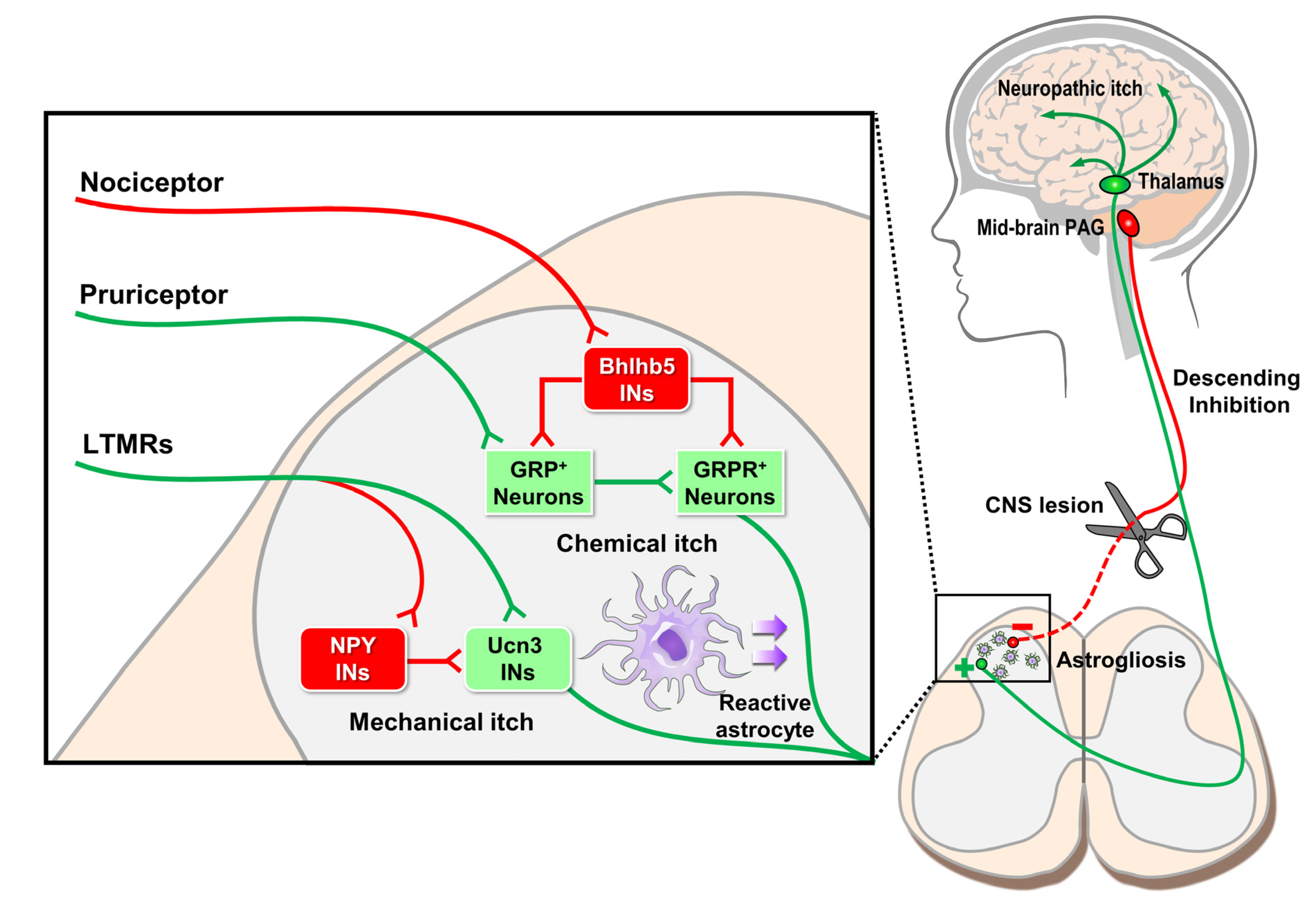
6. Circuit Mechanisms of Neuropathic Itch in the Spinal Cord
7. Descending Supraspinal Control of Itch Sensation
8. Future Therapeutic Directions for Neuropathic Itch
9. Conclusions
Funding
Conflicts of Interest
References
- Oaklander, A.L. Common neuropathic itch syndromes. Acta Derm. Venereol. 2012, 92, 118–125. [Google Scholar] [CrossRef] [PubMed]
- International Association for the Study of Pain Home Page. Available online: https://www.iasp-pain.org/GlobalYear/NeuropathicPain (accessed on 20 September 2020).
- Oaklander, A.L. Neuropathic Itch. In Itch: Mechanisms and Treatment, 1st ed.; Carstens, E., Akiyama, T., Eds.; CRC Press: Boca Raton, FL, USA, 2014; ISBN 978-1466505438. [Google Scholar]
- Cheng, X.Q.; Liang, X.Z.; Wei, S.; Ding, X.; Han, G.H.; Liu, P.; Sun, X.; Quan, Q.; Tang, H.; Zhao, Q.; et al. Protein microarray analysis of cytokine expression changes in distal stumps after sciatic nerve transection. Neural Regen Res. 2020, 15, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, M.; Oaklander, A.L.; Szabo, I.L.; Stander, S.; Schmelz, M. Neuropathic itch. Pain 2019, 160, S11–S16. [Google Scholar] [CrossRef] [PubMed]
- Oaklander, A.L.; Bowsher, D.; Galer, B.; Haanpaa, M.; Jensen, M.P. Herpes zoster itch: Preliminary epidemiologic data. J. Pain 2003, 4, 338–343. [Google Scholar] [CrossRef]
- Binder, A.; Koroschetz, J.; Baron, R. Disease mechanisms in neuropathic itch. Nat. Clin. Pr. Neurol. 2008, 4, 329–337. [Google Scholar] [CrossRef]
- Pereira, M.P.; Derichs, L.; Meyer Zu Horste, G.; Agelopoulos, K.; Stander, S. Generalized chronic itch induced by small-fibre neuropathy: Clinical profile and proposed diagnostic criteria. J. Eur. Acad. Derm. Venereol. 2020, 34, 1795–1802. [Google Scholar] [CrossRef]
- Bellato, E.; Marini, E.; Castoldi, F.; Barbasetti, N.; Mattei, L.; Bonasia, D.E.; Blonna, D. Fibromyalgia syndrome: Etiology, pathogenesis, diagnosis, and treatment. Pain Res. Treat. 2012, 2012, 426130. [Google Scholar] [CrossRef]
- Gracely, R.H.; Schweinhardt, P. Programmed symptoms: Disparate effects united by purpose. Curr. Rheumatol. Rev. 2015, 11, 116–130. [Google Scholar] [CrossRef]
- Laniosz, V.; Wetter, D.A.; Godar, D.A. Dermatologic manifestations of fibromyalgia. Clin. Rheumatol. 2014, 33, 1009–1013. [Google Scholar] [CrossRef]
- Erdogan, H.K.S.; Acer, E.; Bulur, I. Cutaneousfindings infibromyalgia syndrome and their effect on quality of life. Dermatol. Sin. 2016, 34, 131–134. [Google Scholar] [CrossRef]
- Usoskin, D.; Furlan, A.; Islam, S.; Abdo, H.; Lonnerberg, P.; Lou, D.; Hjerling-Leffler, J.; Haeggstrom, J.; Kharchenko, O.; Kharchenko, P.V.; et al. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat. Neurosci. 2015, 18, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.; Torck, A.; Quigley, L.; Wangzhou, A.; Neiman, M.; Rao, C.; Lam, T.; Kim, J.Y.; Kim, T.H.; Zhang, M.Q.; et al. Comparative transcriptome profiling of the human and mouse dorsal root ganglia: An RNA-seq-based resource for pain and sensory neuroscience research. Pain 2018, 159, 1325–1345. [Google Scholar] [CrossRef] [PubMed]
- Nattkemper, L.A.; Tey, H.L.; Valdes-Rodriguez, R.; Lee, H.; Mollanazar, N.K.; Albornoz, C.; Sanders, K.M.; Yosipovitch, G. The Genetics of Chronic Itch: Gene Expression in the Skin of Patients with Atopic Dermatitis and Psoriasis with Severe Itch. J. Investig. Derm. 2018, 138, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Dong, X. Peripheral and Central Mechanisms of Itch. Neuron 2018, 98, 482–494. [Google Scholar] [CrossRef]
- Meixiong, J.; Dong, X. Mas-Related G Protein-Coupled Receptors and the Biology of Itch Sensation. Annu. Rev. Genet. 2017, 51, 103–121. [Google Scholar] [CrossRef]
- Davies, A.J.; Rinaldi, S.; Costigan, M.; Oh, S.B. Cytotoxic Immunity in Peripheral Nerve Injury and Pain. Front. Neurosci. 2020, 14, 142. [Google Scholar] [CrossRef]
- Oetjen, L.K.; Mack, M.R.; Feng, J.; Whelan, T.M.; Niu, H.; Guo, C.J.; Chen, S.; Trier, A.M.; Xu, A.Z.; Tripathi, S.V.; et al. Sensory Neurons Co-opt Classical Immune Signaling Pathways to Mediate Chronic Itch. Cell 2017, 171, 217–228.e213. [Google Scholar] [CrossRef]
- Akiyama, T.; Merrill, A.W.; Carstens, M.I.; Carstens, E. Activation of superficial dorsal horn neurons in the mouse by a PAR-2 agonist and 5-HT: Potential role in itch. J. Neurosci. 2009, 29, 6691–6699. [Google Scholar] [CrossRef]
- Liu, B.; Tai, Y.; Achanta, S.; Kaelberer, M.M.; Caceres, A.I.; Shao, X.; Fang, J.; Jordt, S.E. IL-33/ST2 signaling excites sensory neurons and mediates itch response in a mouse model of poison ivy contact allergy. Proc. Natl. Acad. Sci. USA 2016, 113, E7572–E7579. [Google Scholar] [CrossRef]
- Cevikbas, F.; Wang, X.; Akiyama, T.; Kempkes, C.; Savinko, T.; Antal, A.; Kukova, G.; Buhl, T.; Ikoma, A.; Buddenkotte, J.; et al. A sensory neuron-expressed IL-31 receptor mediates T helper cell-dependent itch: Involvement of TRPV1 and TRPA1. J. Allergy Clin. Immunol. 2014, 133, 448–460. [Google Scholar] [CrossRef]
- Arai, I.; Tsuji, M.; Takeda, H.; Akiyama, N.; Saito, S. A single dose of interleukin-31 (IL-31) causes continuous itch-associated scratching behaviour in mice. Exp. Derm. 2013, 22, 669–671. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Takamori, K. Itch and nerve fibers with special reference to atopic dermatitis: Therapeutic implications. J. Derm. 2014, 41, 205–212. [Google Scholar] [CrossRef]
- Vakharia, P.P.; Chopra, R.; Sacotte, R.; Patel, K.R.; Singam, V.; Patel, N.; Immaneni, S.; White, T.; Kantor, R.; Hsu, D.Y.; et al. Burden of skin pain in atopic dermatitis. Ann. Allergy Asthma Immunol. 2017, 119, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Chen, J.; Hilley, H.; Steele, H.; Yang, J.; Han, L. Molecular Signature of Pruriceptive MrgprA3(+) Neurons. J. Investig. Derm. 2020, 140, 2041–2050. [Google Scholar] [CrossRef]
- Faber, C.G.; Hoeijmakers, J.G.; Ahn, H.S.; Cheng, X.; Han, C.; Choi, J.S.; Estacion, M.; Lauria, G.; Vanhoutte, E.K.; Gerrits, M.M.; et al. Gain of function Nanu1.7 mutations in idiopathic small fiber neuropathy. Ann. Neurol. 2012, 71, 26–39. [Google Scholar] [CrossRef]
- Faber, C.G.; Lauria, G.; Merkies, I.S.; Cheng, X.; Han, C.; Ahn, H.S.; Persson, A.K.; Hoeijmakers, J.G.; Gerrits, M.M.; Pierro, T.; et al. Gain-of-function Nav1.8 mutations in painful neuropathy. Proc. Natl. Acad. Sci. USA 2012, 109, 19444–19449. [Google Scholar] [CrossRef]
- Devigili, G.; Eleopra, R.; Pierro, T.; Lombardi, R.; Rinaldo, S.; Lettieri, C.; Faber, C.G.; Merkies, I.S.; Waxman, S.G.; Lauria, G. Paroxysmal itch caused by gain-of-function Nav1.7 mutation. Pain 2014, 155, 1702–1707. [Google Scholar] [CrossRef]
- Huang, J.; Estacion, M.; Zhao, P.; Dib-Hajj, F.B.; Schulman, B.; Abicht, A.; Kurth, I.; Brockmann, K.; Waxman, S.G.; Dib-Hajj, S.D. A Novel Gain-of-Function Nav1.9 Mutation in a Child With Episodic Pain. Front. Neurosci. 2019, 13, 918. [Google Scholar] [CrossRef]
- Salvatierra, J.; Diaz-Bustamante, M.; Meixiong, J.; Tierney, E.; Dong, X.; Bosmans, F. A disease mutation reveals a role for NaV1.9 in acute itch. J. Clin. Investig. 2018, 128, 5434–5447. [Google Scholar] [CrossRef]
- Han, L.; Ma, C.; Liu, Q.; Weng, H.J.; Cui, Y.; Tang, Z.; Kim, Y.; Nie, H.; Qu, L.; Patel, K.N.; et al. A subpopulation of nociceptors specifically linked to itch. Nat. Neurosci. 2013, 16, 174–182. [Google Scholar] [CrossRef]
- Mishra, S.K.; Hoon, M.A. The cells and circuitry for itch responses in mice. Science 2013, 340, 968–971. [Google Scholar] [CrossRef]
- Sun, Y.G.; Chen, Z.F. A gastrin-releasing peptide receptor mediates the itch sensation in the spinal cord. Nature 2007, 448, 700–703. [Google Scholar] [CrossRef]
- Sun, Y.G.; Zhao, Z.Q.; Meng, X.L.; Yin, J.; Liu, X.Y.; Chen, Z.F. Cellular basis of itch sensation. Science 2009, 325, 1531–1534. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Sikand, P.; Ma, C.; Tang, Z.; Han, L.; Li, Z.; Sun, S.; LaMotte, R.H.; Dong, X. Mechanisms of itch evoked by beta-alanine. J. Neurosci. 2012, 32, 14532–14537. [Google Scholar] [CrossRef] [PubMed]
- Sikand, P.; Dong, X.; LaMotte, R.H. BAM8-22 peptide produces itch and nociceptive sensations in humans independent of histamine release. J. Neurosci. 2011, 31, 7563–7567. [Google Scholar] [CrossRef]
- Sikand, P.; Shimada, S.G.; Green, B.G.; LaMotte, R.H. Sensory responses to injection and punctate application of capsaicin and histamine to the skin. Pain 2011, 152, 2485–2494. [Google Scholar] [CrossRef] [PubMed]
- Sikand, P.; Shimada, S.G.; Green, B.G.; LaMotte, R.H. Similar itch and nociceptive sensations evoked by punctate cutaneous application of capsaicin, histamine and cowhage. Pain 2009, 144, 66–75. [Google Scholar] [CrossRef]
- Andrew, D.; Craig, A.D. Spinothalamic lamina I neurons selectively sensitive to histamine: A central neural pathway for itch. Nat. Neurosci. 2001, 4, 72–77. [Google Scholar] [CrossRef]
- Davidson, S.; Zhang, X.; Khasabov, S.G.; Moser, H.R.; Honda, C.N.; Simone, D.A.; Giesler, G.J., Jr. Pruriceptive spinothalamic tract neurons: Physiological properties and projection targets in the primate. J. Neurophysiol. 2012, 108, 1711–1723. [Google Scholar] [CrossRef]
- Sun, S.; Xu, Q.; Guo, C.; Guan, Y.; Liu, Q.; Dong, X. Leaky gate model: Intensity-dependent coding of pain and itch in the spinal cord. Neuron 2017, 93, 840–853. [Google Scholar] [CrossRef]
- Handwerker, H.O.; Forster, C.; Kirchhoff, C. Discharge patterns of human C-fibers induced by itching and burning stimuli. J. Neurophysiol. 1991, 66, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Tuckett, R.P. Itch evoked by electrical stimulation of the skin. J. Investig. Derm. 1982, 79, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Melzack, R. Phantom limbs. Sci. Am. 1992, 266, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Tey, H.L.; Maddison, B.; Wang, H.; Ishiju, Y.; McMichael, A.; Marks, M.; Willford, P.; Maruzivab, D.; Ferdinando, D.; Dick, J.; et al. Cutaneous innervation and itch in keloids. Acta Derm. Venereol. 2012, 92, 529–531. [Google Scholar] [CrossRef] [PubMed]
- Kwak, I.S.; Choi, Y.H.; Jang, Y.C.; Lee, Y.K. Immunohistochemical analysis of neuropeptides (protein gene product 9.5, substance P and calcitonin gene-related peptide) in hypertrophic burn scar with pain and itching. Burns 2014, 40, 1661–1667. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Yosipovitch, G.; Chan, Y.H.; Goh, C.L. Pruritus, pain, and small nerve fiber function in keloids: A controlled study. J. Am. Acad Derm. 2004, 51, 1002–1006. [Google Scholar] [CrossRef] [PubMed]
- Stella, M.; Calcagni, M.; Teich-Alasia, S.; Ramieri, G.; Cellino, G.; Panzica, G. Sensory endings in skin grafts and scars after extensive burns. Burns 1994, 20, 491–495. [Google Scholar] [CrossRef]
- Giummarra, M.J.; Georgiou-Karistianis, N.; Nicholls, M.E.; Gibson, S.J.; Chou, M.; Bradshaw, J.L. Corporeal awareness and proprioceptive sense of the phantom. Br. J. Psychol. 2010, 101, 791–808. [Google Scholar] [CrossRef]
- Oliveira, K.M.C.; Pindur, L.; Han, Z.; Bhavsar, M.B.; Barker, J.H.; Leppik, L. Time course of traumatic neuroma development. PLoS ONE 2018, 13, e0200548. [Google Scholar] [CrossRef]
- Solorzano, C.; Villafuerte, D.; Meda, K.; Cevikbas, F.; Braz, J.; Sharif-Naeini, R.; Juarez-Salinas, D.; Llewellyn-Smith, I.J.; Guan, Z.; Basbaum, A.I. Primary afferent and spinal cord expression of gastrin-releasing peptide: Message, protein, and antibody concerns. J. Neurosci. 2015, 35, 648–657. [Google Scholar] [CrossRef]
- Fenzi, F.; Benedetti, M.D.; Moretto, G.; Rizzuto, N. Glial cell and macrophage reactions in rat spinal ganglion after peripheral nerve lesions: An immunocytochemical and morphometric study. Arch. Ital. Biol. 2001, 139, 357–365. [Google Scholar] [PubMed]
- Winkelstein, B.A.; Rutkowski, M.D.; Sweitzer, S.M.; Pahl, J.L.; DeLeo, J.A. Nerve injury proximal or distal to the DRG induces similar spinal glial activation and selective cytokine expression but differential behavioral responses to pharmacologic treatment. J. Comp. Neurol. 2001, 439, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Svensson, M.; Eriksson, P.; Persson, J.K.; Molander, C.; Arvidsson, J.; Aldskogius, H. The response of central glia to peripheral nerve injury. Brain Res. Bull. 1993, 30, 499–506. [Google Scholar] [CrossRef]
- Ji, R.R.; Donnelly, C.R.; Nedergaard, M. Astrocytes in chronic pain and itch. Nat. Rev. Neurosci. 2019, 20, 667–685. [Google Scholar] [CrossRef]
- Tsuda, M. Astrocytes in the spinal dorsal horn and chronic itch. Neurosci. Res. 2018, 126, 9–14. [Google Scholar] [CrossRef]
- Shiratori-Hayashi, M.; Koga, K.; Tozaki-Saitoh, H.; Kohro, Y.; Toyonaga, H.; Yamaguchi, C.; Hasegawa, A.; Nakahara, T.; Hachisuka, J.; Akira, S.; et al. STAT3-dependent reactive astrogliosis in the spinal dorsal horn underlies chronic itch. Nat. Med. 2015, 21, 927–931. [Google Scholar] [CrossRef]
- Koga, K.; Yamagata, R.; Kohno, K.; Yamane, T.; Shiratori-Hayashi, M.; Kohro, Y.; Tozaki-Saitoh, H.; Tsuda, M. Sensitization of spinal itch transmission neurons in a mouse model of chronic itch requires an astrocytic factor. J. Allergy Clin. Immunol. 2020, 145, 183–191.e110. [Google Scholar] [CrossRef]
- Netravathi, M.; Saini, J.; Mahadevan, A.; Hari-Krishna, B.; Yadav, R.; Pal, P.K.; Satishchandra, P. Is pruritus an indicator of aquaporin-positive neuromyelitis optica? Mult. Scler. 2017, 23, 810–817. [Google Scholar] [CrossRef]
- Ross, S.E.; Mardinly, A.R.; McCord, A.E.; Zurawski, J.; Cohen, S.; Jung, C.; Hu, L.; Mok, S.I.; Shah, A.; Savner, E.M.; et al. Loss of inhibitory interneurons in the dorsal spinal cord and elevated itch in Bhlhb5 mutant mice. Neuron 2010, 65, 886–898. [Google Scholar] [CrossRef]
- Bourane, S.; Duan, B.; Koch, S.C.; Dalet, A.; Britz, O.; Garcia-Campmany, L.; Kim, E.; Cheng, L.; Ghosh, A.; Ma, Q.; et al. Gate control of mechanical itch by a subpopulation of spinal cord interneurons. Science 2015, 350, 550–554. [Google Scholar] [CrossRef]
- Pan, H.; Fatima, M.; Li, A.; Lee, H.; Cai, W.; Horwitz, L.; Hor, C.C.; Zaher, N.; Cin, M.; Slade, H.; et al. Identification of a Spinal Circuit for Mechanical and Persistent Spontaneous Itch. Neuron 2019, 103, 1135–1149.e1136. [Google Scholar] [CrossRef] [PubMed]
- Oaklander, A.L.; Cohen, S.P.; Raju, S.V. Intractable postherpetic itch and cutaneous deafferentation after facial shingles. Pain 2002, 96, 9–12. [Google Scholar] [CrossRef]
- Lagerstrom, M.C.; Rogoz, K.; Abrahamsen, B.; Persson, E.; Reinius, B.; Nordenankar, K.; Olund, C.; Smith, C.; Mendez, J.A.; Chen, Z.F.; et al. VGLUT2-dependent sensory neurons in the TRPV1 population regulate pain and itch. Neuron 2010, 68, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Heinricher, M.M.; Tavares, I.; Leith, J.L.; Lumb, B.M. Descending control of nociception: Specificity, recruitment and plasticity. Brain Res. Rev. 2009, 60, 214–225. [Google Scholar] [CrossRef]
- Boadas-Vaello, P.; Castany, S.; Homs, J.; Alvarez-Perez, B.; Deulofeu, M.; Verdu, E. Neuroplasticity of ascending and descending pathways after somatosensory system injury: Reviewing knowledge to identify neuropathic pain therapeutic targets. Spinal Cord. 2016, 54, 330–340. [Google Scholar] [CrossRef]
- Kim, Y.S.; Chu, Y.; Han, L.; Li, M.; Li, Z.; LaVinka, P.C.; Sun, S.; Tang, Z.; Park, K.; Caterina, M.J.; et al. Central terminal sensitization of TRPV1 by descending serotonergic facilitation modulates chronic pain. Neuron 2014, 81, 873–887. [Google Scholar] [CrossRef]
- Koeppel, M.C.; Bramont, C.; Ceccaldi, M.; Habib, M.; Sayag, J. Paroxysmal pruritus and multiple sclerosis. Br. J. Derm. 1993, 129, 597–598. [Google Scholar] [CrossRef]
- Pogatzki-Zahn, E.M.; Pereira, M.P.; Cremer, A.; Zeidler, C.; Dreyer, T.; Riepe, C.; Wempe, C.; Lotts, T.; Segelcke, D.; Ringkamp, M.; et al. Peripheral Sensitization and Loss of Descending Inhibition Is a Hallmark of Chronic Pruritus. J. Investig. Derm. 2020, 140, 203–211.e204. [Google Scholar] [CrossRef]
- Samineni, V.K.; Grajales-Reyes, J.G.; Sundaram, S.S.; Yoo, J.J.; Gereau, R.W.t. Cell type-specific modulation of sensory and affective components of itch in the periaqueductal gray. Nat. Commun. 2019, 10, 4356. [Google Scholar] [CrossRef]
- Chapman, J.; Brown, P.; Goldfarb, L.G.; Arlazoroff, A.; Gajdusek, D.C.; Korczyn, A.D. Clinical heterogeneity and unusual presentations of Creutzfeldt-Jakob disease in Jewish patients with the PRNP codon 200 mutation. J. Neurol. Neurosurg. Psychiatry 1993, 56, 1109–1112. [Google Scholar] [CrossRef]
- Ukai, K. Possible effectiveness of low-dose pregabalin in a patient suffering from Lewy body disease and chronic pruritus. Psychogeriatrics 2017, 17, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Leknes, S.G.; Bantick, S.; Willis, C.M.; Wilkinson, J.D.; Wise, R.G.; Tracey, I. Itch and motivation to scratch: An investigation of the central and peripheral correlates of allergen- and histamine-induced itch in humans. J. Neurophysiol. 2007, 97, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Cohen, O.S.; Chapman, J.; Lee, H.; Nitsan, Z.; Appel, S.; Hoffman, C.; Rosenmann, H.; Korczyn, A.D.; Prohovnik, I. Pruritus in familial Creutzfeldt-Jakob disease: A common symptom associated with central nervous system pathology. J. Neurol. 2011, 258, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, P.E.; Braun, C.W. Unilateral pruritus after a stroke. Arch. Derm. 1987, 123, 1527–1530. [Google Scholar] [CrossRef]
- Roncati, L.; Gatti, A.M.; Pusiol, T.; Piscioli, F.; Barbolini, G.; Maiorana, A. The first investigative science-based evidence of Morgellons psychogenesis. Ultrastruct. Pathol. 2016, 40, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Orsolini, L.; Gentilotti, A.; Giordani, M.; Volpe, U. Historical and clinical considerations on Ekbom’s syndrome. Int. Rev. Psychiatry 2020, 32, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Haas, N.L.; Nicholson, A.; Haas, M.R.C. Delusional parasitosis as presenting symptom of occipital lobe cerebrovascular accident. Am. J. Emerg. Med. 2019, 37, 1990.e3–1990.e5. [Google Scholar] [CrossRef] [PubMed]
- Schneider, G.; Grebe, A.; Bruland, P.; Heuft, G.; Stander, S. Criteria Suggestive of Psychological Components of Itch and Soma-toform Itch: Study of a Large Sample of Patients with Chronic Pruritus. Acta Derm. Venereol. 2020, 100, adv00075. [Google Scholar] [CrossRef]
- Meixiong, J.; Anderson, M.; Limjunyawong, N.; Sabbagh, M.F.; Hu, E.; Mack, M.R.; Oetjen, L.K.; Wang, F.; Kim, B.S.; Dong, X. Activation of Mast-Cell-Expressed Mas-Related G-Protein-Coupled Receptors Drives Non-histaminergic Itch. Immunity 2019, 50, 1163–1171.e1165. [Google Scholar] [CrossRef]
- Meixiong, J.; Vasavda, C.; Green, D.; Zheng, Q.; Qi, L.; Kwatra, S.G.; Hamilton, J.P.; Snyder, S.H.; Dong, X. Identification of a bilirubin receptor that may mediate a component of cholestatic itch. Elife 2019, 8. [Google Scholar] [CrossRef]
- Meixiong, J.; Vasavda, C.; Snyder, S.H.; Dong, X. MRGPRX4 is a G protein-coupled receptor activated by bile acids that may contribute to cholestatic pruritus. Proc. Natl. Acad. Sci. USA 2019, 116, 10525–10530. [Google Scholar] [CrossRef] [PubMed]
- Feldman, S.R.; Green, L.; Kimball, A.B.; Siu, K.; Zhao, Y.; Herrera, V.; Nyirady, J.; Alexis, A.F. Secukinumab improves scalp pain, itching, scaling and quality of life in patients with moderate-to-severe scalp psoriasis. J. Dermatol. Treat. 2017, 28, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.L.; Savage, K.T.; Qiu, C.C.; Jin, A.; Valdes-Rodriguez, R.; Mollanazar, N.K. Chronic Pruritus Responding to Dupilumab-A Case Series. Medines 2019, 6, 72. [Google Scholar] [CrossRef]
- Chiricozzi, A.; Maurelli, M.; Gori, N.; Argenziano, G.; De Simone, C.; Calabrese, G.; Girolomoni, G.; Peris, K. Dupilumab improves clinical manifestations, symptoms, and quality of life in adult patients with chronic nodular prurigo. J. Am. Acad. Derm. 2020, 83, 39–45. [Google Scholar] [CrossRef]
- Stander, S.; Yosipovitch, G.; Legat, F.J.; Lacour, J.P.; Paul, C.; Narbutt, J.; Bieber, T.; Misery, L.; Wollenberg, A.; Reich, A.; et al. Trial of Nemolizumab in Moderate-to-Severe Prurigo Nodularis. N. Engl. J. Med. 2020, 382, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Kabashima, K.; Matsumura, T.; Komazaki, H.; Kawashima, M.; Nemolizumab, J.P.S.G. Trial of Nemolizumab and Topical Agents for Atopic Dermatitis with Pruritus. N. Engl. J. Med. 2020, 383, 141–150. [Google Scholar] [CrossRef]
- Stander, S.; Kwon, P.; Hirman, J.; Perlman, A.J.; Weisshaar, E.; Metz, M.; Luger, T.A.; Group, T.C.P.S. Serlopitant reduced pruritus in patients with prurigo nodularis in a phase 2, randomized, placebo-controlled trial. J. Am. Acad. Derm. 2019, 80, 1395–1402. [Google Scholar] [CrossRef]
- Yosipovitch, G.; Stander, S.; Kerby, M.B.; Larrick, J.W.; Perlman, A.J.; Schnipper, E.F.; Zhang, X.; Tang, J.Y.; Luger, T.; Steinhoff, M. Serlopitant for the treatment of chronic pruritus: Results of a randomized, multicenter, placebo-controlled phase 2 clinical trial. J. Am. Acad. Derm. 2018, 78, 882–891.e810. [Google Scholar] [CrossRef]
- Boozalis, E.; Sheu, M.; Selph, J.; Kwatra, S.G. Botulinum toxin type A for the treatment of localized recalcitrant chronic pruritus. J. Am. Acad. Derm. 2018, 78, 192–194. [Google Scholar] [CrossRef]
- Wallengren, J.; Bartosik, J. Botulinum toxin type A for neuropathic itch. Br. J. Derm. 2010, 163, 424–426. [Google Scholar] [CrossRef]
- Shaarawy, E.; Hegazy, R.A.; Abdel Hay, R.M. Intralesional botulinum toxin type A equally effective and better tolerated than intralesional steroid in the treatment of keloids: A randomized controlled trial. J. Cosmet Derm. 2015, 14, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Maari, C.; Marchessault, P.; Bissonnette, R. Treatment of notalgia paresthetica with botulinum toxin A: A double-blind randomized controlled trial. J. Am. Acad. Derm. 2014, 70, 1139–1141. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.G.; Grossman, S.K.; Valdes-Rodriguez, R.; Berenato, F.; Korbutov, J.; Chan, Y.H.; Lavery, M.J.; Yosipovitch, G. Topical ketamine-amitriptyline-lidocaine for chronic pruritus: A retrospective study assessing efficacy and tolerability. J. Am. Acad. Derm. 2017, 76, 760–761. [Google Scholar] [CrossRef] [PubMed]
- Papoiu, A.D.; Valdes-Rodriguez, R.; Nattkemper, L.A.; Chan, Y.H.; Hahn, G.S.; Yosipovitch, G. A novel topical formulation containing strontium chloride significantly reduces the intensity and duration of cowhage-induced itch. Acta Derm. Venereol. 2013, 93, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Kardon, A.P.; Polgar, E.; Hachisuka, J.; Snyder, L.M.; Cameron, D.; Savage, S.; Cai, X.; Karnup, S.; Fan, C.R.; Hemenway, G.M.; et al. Dynorphin acts as a neuromodulator to inhibit itch in the dorsal horn of the spinal cord. Neuron 2014, 82, 573–586. [Google Scholar] [CrossRef]
- Fishbane, S.; Jamal, A.; Munera, C.; Wen, W.; Menzaghi, F.; Investigators, K.-T. A Phase 3 Trial of Difelikefalin in Hemodialysis Patients with Pruritus. N. Engl. J. Med. 2020, 382, 222–232. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meixiong, J.; Dong, X.; Weng, H.-J. Neuropathic Itch. Cells 2020, 9, 2263. https://doi.org/10.3390/cells9102263
Meixiong J, Dong X, Weng H-J. Neuropathic Itch. Cells. 2020; 9(10):2263. https://doi.org/10.3390/cells9102263
Chicago/Turabian StyleMeixiong, James, Xinzhong Dong, and Hao-Jui Weng. 2020. "Neuropathic Itch" Cells 9, no. 10: 2263. https://doi.org/10.3390/cells9102263
APA StyleMeixiong, J., Dong, X., & Weng, H.-J. (2020). Neuropathic Itch. Cells, 9(10), 2263. https://doi.org/10.3390/cells9102263