1. Introduction
In order to cover up the deficit of regenerative potential, new treatment approaches for cardiac diseases are needed. Several studies suggest that approaches based on the application of stem cells may be the future strategy in order to compensate the regenerative deficiency of the heart. As myocardial infarction (MI) occurs due to the absent blood flow in the coronary arteries, the formation of new blood vessels would be an essential prerequisite for the regeneration. Yet, in the search for suitable stem cell subtypes, various other properties are of interest as well. This includes cells that interfere with pathological immune mechanisms as well as cellular mediators and chemokines that cause migration and homing processes. Besides, it is of the tremendous importance whether the cells are proofed to be safe in application. Furthermore, the cells can also be treated in advance in order to develop desirable properties. As the CD133
+ stem cell therapies and other approaches with hematopoietic cells provide promising results, research has to examine more cell types in order to achieve additional effects and thus to further improve the therapeutic measures. Until now, no stem cell-related cardiac complications following intramyocardial transplantation of bone marrow (BM) derived stem cells have been reported so that the therapy is considered to be safe [
1]. In addition, the BM also contains a population named after their ability to differentiate into various cell types of mesenchymal origin. These multipotent stem cells are expandable and modifiable in cell culture and offer several distinctive characteristics, which make them a promising therapeutic approach for damaged heart tissue [
2]. Especially mesenchymal stem cells (MSC) are able to decrease infarct size and myocardial fibrosis [
3]. Thereby, cardiac remodeling in total may be attenuated as well as the heart’s contractility improves [
3]. Furthermore, MSC have paracrine effects that inhibit apoptosis of endothelial cells in hypoxic condition, increase their survival, and thus stimulate angiogenesis processes [
4]. As another advantage, MSC do not express any antigens of histocompatibility and thus can be used in allogenic transplantation as well. Quite the contrary, these cells even have immunomodulatory paracrine effects that counteract inflammatory responses [
5,
6]. As inflammatory processes play a decisive role in the development of chronic heart diseases and have a negative influence on the heart function, these effects may also improve the therapy significantly. After the transplantation, MSC may be able to stimulate the production of reparative growth factors, inhibit local inflammatory response, and replace damaged cells [
7]. Apart from these positive effects, difficulties with the practical application arise from the large heterogeneity of MSC as well. Thus, it is uncertain to predict which exact cell type arises from the MSC population after the application. Indeed, a study by Yoon et al. showed that the direct transplantation of unselected BM cells into the infarcted myocardium induces significant intramyocardial calcification [
8]. This clearly demonstrates the research demands in order to guarantee the safety of MSC as therapeutic option. The development of methods for the identification and isolation of MSC subpopulations is crucial for a determined specific function and a successful clinical application. A currently much examined MSC subtype is characterized by the expression of the surface marker CD271. Several studies showed that this marker is one of the most specific concerning BM derived MSC [
9]. Thus, there may be a particular suitability of CD271
+ MSC for preselected cell products. CD271 enables prospective cell isolation of MSC from BM derived mononuclear cells (MNC) [
10] and have been shown to be essential in the formation of the heart as they influence the survival of endothelial cells, vascular smooth muscle cells, and cardiac myocytes. Furthermore, these proteins regulate angiogenesis and vasculogenesis by autocrine and paracrine mechanisms [
10,
11,
12].
Knowledge of the therapeutic potential attributable to distinct BM derived stem cell subtypes primed due to MI of the donors is still greatly lacking. Therefore, this study intends to examine the benefits of CD271+ and CD133+ stem cells. In this context, we have explored the effects of CD271+ cells alone as well as in combination with CD133+ with the main focus on their angiogenic potential (in vitro examination). Moreover, the therapeutic capacities of both primed BM stem cells were investigated following cardiac ischemia/reperfusion in mice.
2. Materials and Methods
2.1. Bone Marrow Aspiration
Informed donors gave written consent to the aspiration of their BM according to the Declaration of Helsinki. The ethical committee of the University of Rostock has approved the presented study (registered as no. A201023) as of 29 April, 2010. BM samples were obtained by sternal aspiration from patients undergoing coronary artery bypass grafting (CABG) surgery at Rostock University, Germany. Anticoagulation was achieved by heparinization with 250 i.E./mL sodium heparine (B. Braun Melsungen AG, Melsungen Germany).
2.2. Cell Isolation (Single and Dual Method)
Mononuclear cells (MNC) were isolated by density gradient centrifugation using Lymphocyte Separation Medium (LSM; 1.077 g/L, PAA Laboratories GmbH, Pasching, Austria). CD133+ and CD271+ stem cells were enriched by positive magnetic selection using the magnet activated cell sorting (MACS) system (Miltenyi Biotec, Bergisch Gladbach, Germany). In case of CD133+ direct labeling was applied and indirect labeling for CD271+ cells (CD271-APC/anti-APC-microbeads; all Miltenyi Biotec). Purified stem cells were appropriately employed in further in vitro and in vivo experiments. For the dual method, the CD133+ population was isolated from the CD271– fraction and vice versa.
2.3. Phenotypical Characterization by Flow Cytometry
Cell fractions were suspended in cold MACS-buffer containing PBS, 2 mM EDTA and 0.5% BSA. To reduce unspecific binding, FcR blocking reagent (Miltenyi Biotec) was added to all samples. Cells were stained using the mouse anti-human-CD133-phycoerythrin (PE; 293C2), -CD34-fluorescein isothiocyanate (FITC) and -CD271-allophycocyanine (all Miltenyi Biotec), -CD45-allophycocyanin-H7 as well as -CD45-Horizon-V500 (both BD Biosciences, Heidelberg, Germany) following incubation for 10 min in the dark at 4 °C. For optimal multicolor setting and correction of the spectral overlap single stained mouse isotype antibodies were considered as controls. The gating strategy was performed with matched isotype/fluorescence minus one control. After performing antibody staining 15 µM 4′,6-Diamidino-2-phenylindole (DAPI, Thermo Fisher Scientific, Waltham, MA, USA.) was added, cells were incubated for 2 min and then immediately acquired by BD™ LSRII flow cytometer and data were analyzed using FACS-Diva software, version 6.1.2 (both Becton Dickinson, Franklin Lakes, NJ, USA). Purity and viability of all cell isolations were analyzed using near-IR live dead stain (Thermo Fisher).
2.4. Angiogenesis Assay
Freshly isolated stem cells were prepared at 4 °C in an extracellular growth matrix using 100 µL BD MatrigelTM Matrix (BD Biosciences) and 100 µL complete endothelial growth medium (EGM-2, Lonza, Basel, Switzerland) per well of a 24-well-plate (Greiner Bio-One, Kremsmünster, Austria). For the assay 100,000 CD133+ or CD271+ cells (single model) or both cell types in a ratio of 50,000 cells each (co-culture model) were seeded per well. After 20 min polymerization of the matrix at 37 °C the matrix 200 µL EGM-2was added carefully onto each well. The medium was changed every two days for another 14 days.
2.5. Three Dimensional (3D) Microscopy Analysis
After two weeks the tube formation assays were conducted according to the criteria of network length and count of nodal points. For this, z-stack images were acquired using the Zeiss high-resolution microscope ELYRA PS.1 LSM 780 confocal imaging system and corresponding Zen2011 software (both Carl Zeiss AG, Oberkochen, Germany). Z-Stack images are transversal slice images (two-dimensional) of the 3D assay and thus allow representative analysis of their structures. For each well, five z-stacks with ten images were taken and included into a final interpretation. The evaluation was performed using the ImageJ free software (NIH, Bethesda, MD, USA). For better illustration, the different levels of the z-stack were marked in different colors. Likewise, the amount of nodal points was counted, respectively. In total, 900 images (n = 6) were analyzed and measured toward network length and count of nodal points.
2.6. Cell Tracking within Matrigel Matrix
In order to further investigate the cell networks accomplished in Matrigel matrix, immunofluorescence staining was carried out on angiogenesis assay. For better discrimination and alterations within the matrix, freshly isolated CD133+ cells were stained with the lipophilic cell permeable dye CFDA-SE as well as CD271+ cells with the red fluorescent lipophilic tracer PKH26 (both Sigma-Aldrich, Saint Louis, MO, USA). Additionally, both cell types were stained for nuclei discrimination with Hoechst 33324 (Thermo Fisher). Acquisition and analyzes were performed using the Axiovert 40 CFL fluorescence microscope with Axio Cam MRm ZEN software (both Carl Zeiss AG).
2.7. Immunofluorescence Staining within 3D Matrix
Mouse anti-human-CD29 allophycocyanin as well as -CD73-phycoerythrin antibodies (both BD Biosciences) were diluted with EGM-2 in 1:10 ratio and incubated with the cells for 30 min. Afterwards, the assays were washed with EGM-2. For each marker an isotype control was applied in the same way in order to obtain a negative control. Additionally, both cell types were stained with Hoechst 33324. The analysis was performed by means of the Zeiss high-resolution microscope ELYRA PS.1 LSM 780 confocal imaging system and corresponding Zen2011 software Z-stack images were used for 3D reconstructions.
2.8. Gene Expression Analysis by Quantitative Real-Time-PCR
Cells derived from the single and co-culture models were collected after the termination of the angiogenesis assay and have undergone lysis in TRIzol® reagent (Thermo Fisher). RNA was extracted following the manufacturer’s instructions. For reverse transcription of total RNA amount (2 µg) and cDNA synthesis, SuperScript® III Reverse Transcriptase (Thermo Fisher) and oligo (dT)15 Primers (Promega, Fitchburg, WI, USA) were used. Quantitative real time-PCR was performed with StepOnePlusTM Real-Time PCR System (Applied Biosystems, Waltham, MA, USA) in TaqMan® Universal Master Mix (Thermo Fisher) according to the instructions of the manufacturer. The expression of the housekeeping gene ribosomal protein, large, P0 (human RPLP0, TaqManTM VIC® Endogenous Control 4310879E) was used on each cell type. Similarly, human ACTA (alpha smooth muscle actin) (TaqMan® Assay ID: Hs00909449_m1, FAM-MGB), NGFR (nerve growth factor receptor) (TaqMan® Assay ID: Hs00609976_m1, FAM-MGB) and vWF (von Willebrand factor) (TaqMan® Assay ID: Hs01109446_m1, FAM-MGB, all Thermo Fisher) were analyzed in duplicates and normalized to RPLP0. Negative controls were included in each assay. Cycle thresholds (CT) for single reactions were determined with StepOne™ Software 2.0 (formula: ΔCT mean = CT mean − CT mean RPLP0).
2.9. Animals
All animal procedures were in conformity with the guidelines from Directive 2010/63/EU of the European Parliament on the protection of animals used for scientific purposes. The federal animal care committee of LALLF Mecklenburg-Vorpommern (Germany) approved the study protocol (approval number LALLF M-V/TSD/7221.3-1.1-088/11). Severe Combined Immunodeficiency beige mice (SCID beige; strain CB17.Cg-PrkdcscidLystbg-J/Crl, female, 22 ± 2 g, Charles River, Sulzfeld, Germany) were randomly assigned to 4 groups: Healthy control (SHAM, n = 3), two MI groups with implanted human stem cells of the respective source (MI133, MI271 each n = 3) and untreated MI control group (MIC n = 3).
2.10. Generation of Reperfused MI in Mice and Stem Cell Implantation
Mice were anesthetized with 50mg/kg Pentobarbital (Vetmedica GmbH, Ingelheim, Germany) intraperitoneal injection. After thoracotomy and preparation, the left anterior descending coronary artery (LAD) was ligated. After 45 min each mouse received an intramyocardial cell injection. For cell treatment, 100,000 stem cells were suspended in 10 µL PBS containing 0.5% BSA and 2 mM EDTA and mixed with an equal amount of reduced growth-factor BD MatrigelTM Matrix. The same experimental set-up was applied to control groups using cell suspension buffer and MatrigelTM for application. Four injections, 5 µL each were given along the border of the blanched myocardium. Subsequently, the ligation was removed. Healthy control (SHAM)-operated mice underwent identical surgical procedures without left anterior descending coronary artery (LAD)-ligation.
2.11. Organ Harvesting
Mice underwent euthanization 48 h after intervention by cervical dislocation. Each heart was removed, embedded in O.C.T.
TM Compound (Sakura Finetek, Alphen aan den Rijn, Nederlands) and snap-frozen in liquid nitrogen. For further examinations of the infarction area [
13] and RT-PCR, the heart tissue was divided into four horizontal levels from the apex to the base and cut into slices.
2.12. Immunofluorescence Staining within Tissue Sections
For immunofluorescence staining slices of cryosectioned hearts were fixed with 4% paraformaldehyde (Sigma Aldrich) for 20 min at room temperature following treatment with M.O.M.TM Mouse Ig Blocking Reagent and M.O.M.TM protein concentrate according to the instructions of Vector® M.O.M.TM Immunodetection Kit (LINARIS GmbH, Mannheim, Germany). To identify human cells, monoclonal anti-human-nuclei (Merck Millipore, Darmstadt, Germany) primary antibody as first and anti-mouse Alexa-Fluor® 594 as secondary antibody was used. Nuclei were stained with DAPI and the tissue was mounted using coverslip and Dako mounting medium (Dako, Santa Clara, CA, USA).
2.13. Gene Expression Analysis by RT² Profiler PCR Array
Interlayers of the cryosectioned hearts were used for RNA-isolation, which was performed with TRIzol® reagent. Reverse transcription as well as PCR were performed using RT² Profiler PCR Array Mouse Angiogenesis® kit (Qiagen, Germantown, MD, USA). According to the instructions of the manufacturer an RT2 SYBR Green ROX qPCR Mastermix (Qiagen) was used. The expression of the housekeeping genes ActB, B2M, GAPDH, GUSB, and HSP90AB1 as well as the gene panel were analyzed in triplicates and normalized to the mean of all the housekeeping genes. No genomic DNA was found in the samples. Moreover, we observed no apparent inhibition by any impurities that affect the reverse transcription as well as the amplification of the gene-specific products.
2.14. Statistical Analysis
Statistical analysis was performed using SigmaStat (Version 3.5, Systat Software, San Jose, CA, USA) and IBM SPSS Statistics for Windows (Version 22.0., IBM Corp., Armonk, NY, USA). Comparisons of two experimental groups were performed using Mann–Whitney U test, when data were not normally distributed (Shapiro–Wilk’s test) otherwise Student t-test was utilized. p values ≤0.05 were considered as statistically significant.
4. Discussion
Our present study predominantly relied on investigation of angiogenic capabilities of primed human CD133
+ and CD271
+ BM derived stem cells alone and in a co-culture in vitro as well as in mice using combined cell transplantation post MI. Recently, we already have demonstrated the beneficial therapy when a single administration of stem cells was applied [
13].
First, we were able to achieve suitable isolation protocols and storage parameters. The results from the manual immuno-magnetic cell separation in combination with flow cytometric analysis revealed relatively few CD133
+ and CD271
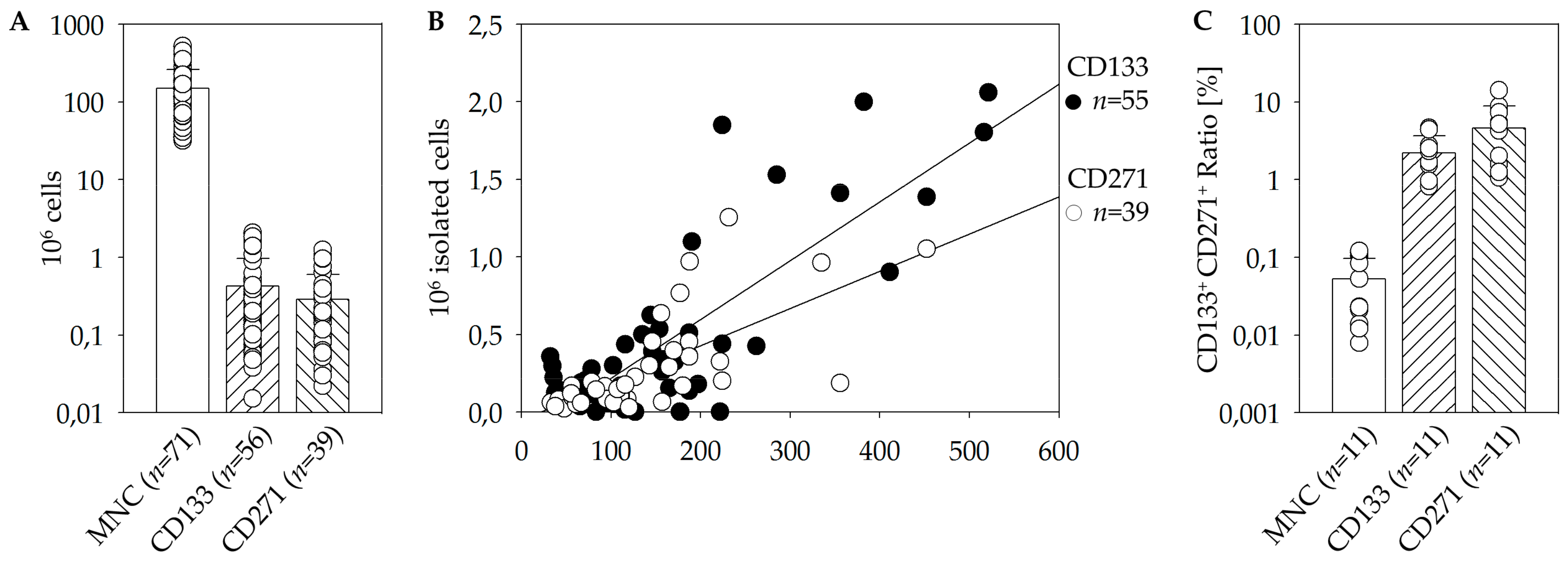
+ cells within BM. Moreover, a linear dependence between target stem cells and the initial amount of MNC has been found (
Figure 1B). However, the initial cell numbers and percentages differed eminently within the patient’s cohort [
17,
18].
Whether CD133 and CD271 are separate subsets or co-expression exists, is an important aspect in order to evaluate the feasibility of a combined therapeutic approach. In conclusion, CD133
+ CD271
+ double-positive cells are represented at a higher proportion within the CD133
+ as well as the CD271
+ cells than within the total MNC fraction (
Figure 1C). Nevertheless, only a small percentage of isolated cells express both surface markers. During magnetic cell separation no selective enrichment or depletion of double or single positive cells took place. Therefore, the separation of the two populations is considered to be sufficient for this study and the investigation of their separate and combined effects.
In clinical setting, autologous stem cells may be transplanted 24 to 30 h after BM aspiration [
19]. A recent study by our laboratory closely investigated the impact of short-term storage on human CD133
+ cells. At this, we used standardized non-freezing storage conditions even for up to 72 h. We have shown that cell number as well as metabolic activity decreased after 30 h, although no significant alterations were observed in cell viability at this point over the time. In line with that, also present results do not indicate a negative influence of storage on cell numbers or quality (
Table 1). This applies to storage of BM as well as for negative fractions intended for dual isolation.
In the present study, we have characterized the phenotype of both stem cells types. The flow cytometric strategy relied on our previous work [
13,
15]. Cultivated MSC are often characterized by the expression of several surface marker including CD73, CD105, and CD44 as well as the absence of others, including CD45 and CD34 [
2,
13]. In our analysis the expression of CD73 and CD105 was significantly increased on CD271
+ cells compared to remaining BM subtypes. We have already detected a favorable survival pattern, improved healing performance, and a more robust preservation of cardiac function for CD105
+ cells in infarcted hearts [
20]. Therefore, the increased expression of CD105 on the CD271
+ cell population can be expected to have a positive influence on the angiogenic potential. Whereas most of cultivated MSC do not express CD45, the largest proportion of freshly isolated CD271
+ cells expressed CD45
dim - a surface marker that has been correlated to mesenchymal colony formation by Cuthbert and coworkers [
16]. The percentage of CD45
- cells was small and showed considerable fluctuations between patients. In conclusion, the most frequent population in purified CD271
+ cells presumably consists of mesenchymal progenitors [
13]. Of note, as our recent flow cytometric observation revealed, the same CD271
+ MSC used for the present study have undergone phenotypic alterations toward a pericytic phenotype [
13]. This is in agreement with the observation by Kutcher and coworkers and Bellagamba and associates [
21,
22].
Proliferative capacity of MSC is associated with their clonogenic properties. In accordance with Quirici and coworkers [
10], colony-formation was only detected in MNC and CD271
+ fraction, but neither in CD271
– nor in CD133
+ population [
13]. The ability to proliferate is a major contributor to the impacts and effectiveness of stem cells in clinical applications. This applies to various functions such as differentiation into specialized mature cells as well as self-renewal or angiogenic processes. Although CD133
+ HSC do not show colony formation in CFU-F assays, they have been demonstrated to proliferate and generate offspring with endothelial characteristics in CFU-EC assays [
19]. CFU-EC acquire the functional phenotype of EC and also show typical gene expression including vWF [
19] and angiogenic factors such as FGF3, PDGFB, and others [
23]. Furthermore, we detected a significant augmentation of EC-colonies after hypoxic preconditioning of CD133
+ cells and thus assumed that hypoxia may lead to the differentiation of CD133
+ cells towards endothelial lineage [
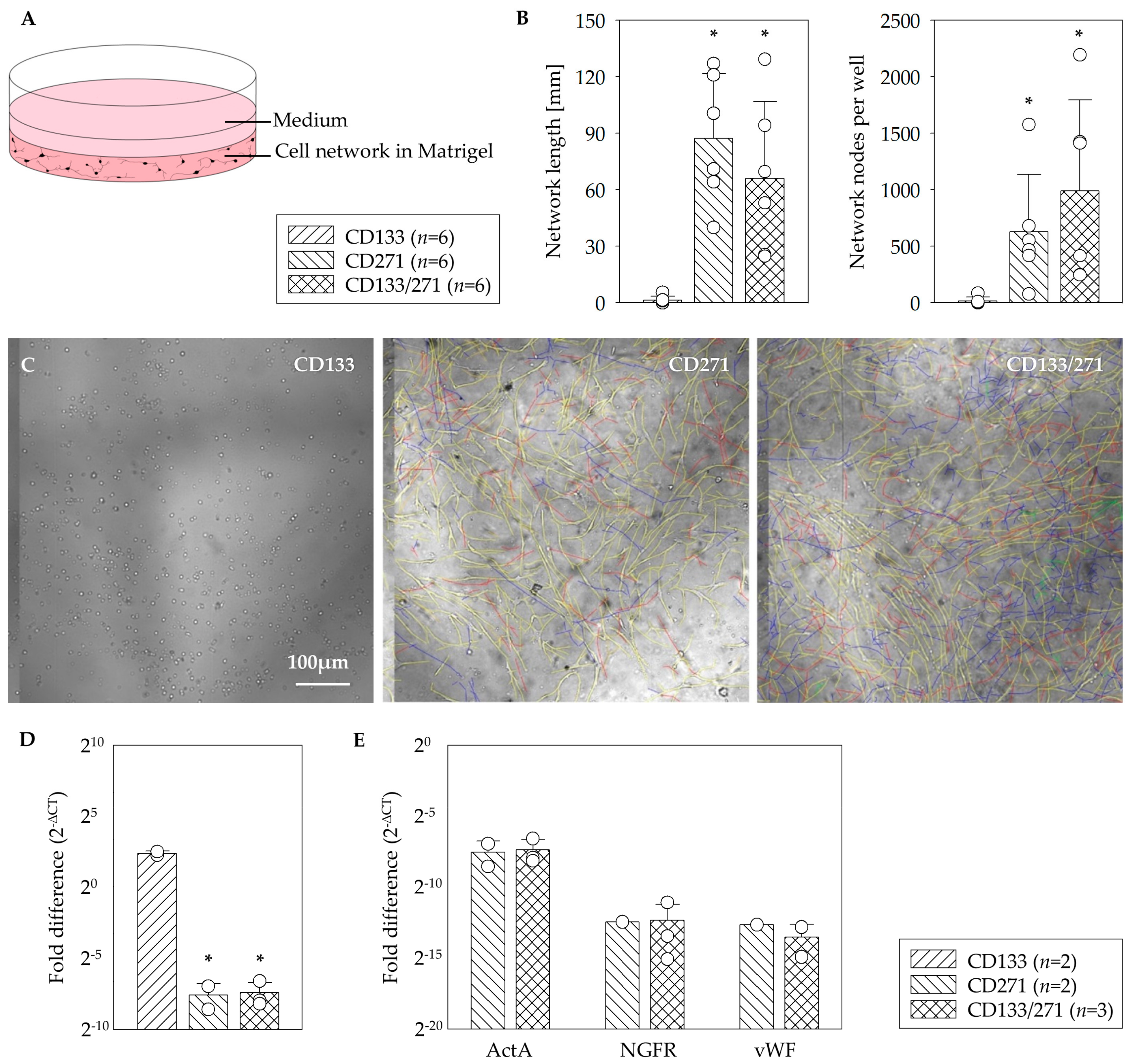
23]. Our results indicate that in vitro network formation relies on CD271
+ cells. Since CD133
+ cells alone rarely build network structures at all, it is remarkable that they will nevertheless contribute to the network, if they are cultivated in combination with CD271
+ cells (
Figure 2). The fluorescence images provided evidence for this contribution as both fluorescent signals were detected from within the networks (
Figure 3). Noteworthy, we previously observed similar network formation also in co-culture when primed cardiac CD45
- CD44
+ DDR2
+ MSC from MI-induced rats were setup together with endothelial cells [
24]. Immunoflorescence staining indicated a strong expression of CD73 marker within the networks built by CD271
+ as well as CD133
+/CD271
+ cells of the angiogenesis assays. CD73 is often referred to us the MSC phenotype [
25,
26]. CD133
+ cell networks also expressed CD73 whereas the maker was not detected on CD133
+ cells that did not form network structures. For this study the involvement of the marker in angiogenic processes is of particular importance. At this, CD73 does not only seem to be associated with cells that are involved in angiogenesis, but appears to play a crucial role in the process itself. A study by Wang and associates found that capillary-like structures were formed more in CD73
+/+ pulmonary microvascular endothelial cells (PMEC) than in CD73
−/− PMEC in vitro. In accordance with that they also observed that the extent of tumor angiogenesis as well as the tumor size was greater in CD73
+/+ mice compared to CD73
−/− mice in vivo. Furthermore, CD73 expression decreased the adhesion of EC to collagen IV and promoted migration. Thus, they concluded that CD73 contributes to EC forming new vessels especially in cancer conditions [
27]. A study by Allard and associates demonstrated that both tumor and host-derived CD73 are involved in these processes. In fact, tumor-derived CD73 enhances the production of vascular endothelial growth factor (VEGF) by tumor cells whereas host-derived CD73 is required for in vivo angiogenic responses. Furthermore, they also confirmed that EC require CD73 expression for tube formation and migration [
28]. Consequently, measures against CD73 have been shown to impair angiogenesis in tumors [
28,
29]. However, Böring and associates stated that CD73 deficiency has no effect on angiogenesis in their mice model of hind limb ischemia [
30]. Thus, it is not certain if the angiogenic effect applies on tumor tissue only and might be different in non-tumor environment. The increased expression of CD73 especially on CD271
+ cells may have positive impact on the angiogenic potential of the stem cell treatment. We were able to show that CD271 network structures express ActA and thus a marker for muscular differentiation. This expression was 10 fold higher in assays consisting of CD133
+ cell cultures (
Figure 2D). To simulate network formation in vitro, we used Matrigel as a 3D framework. Interestingly the CD133
+ cell culture in Matrigel led to a 10-fold higher level of
ActA (
α-SMA) gene in comparison to the CD133
+/271
+ cell co-culture. However, Lu and associates observed that following culture in endothelial cell-promoting environment CD133
+ cells did not produce endothelial-like cells that expressed α-SMA, but only HUVEC and CD34
+ progenitor cells [
31]. As we have already shown recently, flow cytometric analysis of the CD133
+ cell product isolated from sternal BM (similar patient’s disease baseline as in the present study) was characterized by the co-expression of approx. 80% of CD34 marker [
15]. Therefore, detected
ActA mRNA levels in our CD133
+ cell angiogenesis assay seem to be reasonable. Interestingly, CD133
+ αSMA
+ phenotype was found only in association with blood vessels of cancer-associated fibroblasts [
32], confirming the linkage of α-SMA expression to endothelial-like origin and cell activation. Furthermore, the possible reason of less or comparable
ActaA mRNA levels with a CD271
+ cell culture for the CD133
+/CD271
+ cell co-culture might be explained by the phenotypical alterations of CD133
+ hematopoietic stem cells when applied together with MSC and/or due to the maintained undifferentiated state of MSC in co-culture model [
33,
34]. Hence, these in vitro co-culture conditions may have a reducing effect on the
α-SMA mRNA levels in total.
Moreover, we examined mRNA expression profiles using the infarcted area of heart cross sections. In line with recent observations of the proliferative capacities and induction of regenerative pathways, demonstrated by our group [
24], the present study focused on testing of possible angiogenic potential in the early phase post MI. On the one hand in a murine model, the switch between the early inflammatory and the proliferative phase eventuate 2-4 days after MI [
35]. At the same time, neovascularization of surrounding, viable myocardium in the infarct border zone is also crucial during this process of tissue remodeling, although angiogenesis occurs in the granulation tissue that will ultimately form the infarct scar [
36]. On the other hand, a complete engraftment and no significant differences in the retention of both cell subsets were evident in the hearts 48 h following MI as shown in our own previous work [
13]. Indeed, in a recent long term study using the identical human stem cell types as well as the identical mouse model we did observe functional improvement [
13].
Recently, we could show the co-localization of CD133
+ cell derivatives with functional blood vessels 3 weeks post transplantation. This direct impact of CD133
+ stem cells on angiogenesis was not detected for CD271
+ cells, suggesting different cellular mechanisms for cardiac regeneration [
13]. Coherently, indications of the present study 48 h after MI reveal positive effects of the stem cell therapy regarding angiogenesis. The expression of the pro-angiogenic cytokines Cxcl1 and Cxcl2 has been significantly increased in the CD133
+ cell treated animal group, whereas the MI271 group showed less expression (
Figure 4A). However, the same applies to the expression of IL1B and IL6 as well as Timp1 [
37] (
Figure 4B). Al-Amran and co-workers demonstrated that down-regulation of Cxcl2 results in decrement in myocardial neutrophil infiltration, which is associated with less reactive oxygen species and reactive nitrogen species formation after global ischemia reperfusion apoptosis [
38]. Therefore, we assume, that CD133
+ cell therapy may have a valuable impact on a moderate course of the neutrophil-mediated tissue injury [
39], although, we only could observe a significant lower
TGFβ3 mRNA level of this fibrotic marker in MI133 therapy group when compared with MI271 [
40]. Our long term follow up experiments revealed unanticipated data. Subsequently, both cell therapy groups led to a decreased collagen deposition at the infarction border as well as improved vessel infiltration of scar tissue compared to the MIC group [
13,
40,
41,
42]. Again, these data show possible different cellular mechanisms for angiogenic effects mediated via CD133
+ vs. CD271
+. Furthermore, many existing in vitro studies confirmed the involvement of TGFβ in the activation of cardiac fibroblasts. Nevertheless, the in vivo functioning has not completely been explained because of the complexity and the context-dependent signaling cascades [
40]. Interestingly, mRNA levels for pro-inflammatory interleukins (see above) within MI133 therapy group were significantly enhanced, while TGFβ3 levels were rather reduced. Since IL-6 bears pleiotropic properties, a possible switch from pro- into anti-inflammatory or even immunomodulatory signaling cascades, would be in favor for a respective therapy. Importantly, it has been shown, that both IL-6 with TGFβ play a crucial role in the shift between inhibitory Tregs and pro-inflammatory Th17 cells and this is associated with their ratio at all [
43].
Finally, we have observed significantly increased mRNA levels of the pro angiogenic factor VEGFC after CD271
+ stem cell treatment compared to MI133 group (
Figure 4B). It is known that MSC promotes angiogenesis by releasing VEGF [
44,
45,
46]. Accordingly, Liu and associates observed enhanced cardiac function, increased perfusion and angiogenesis in vivo using MSC as a VEGF delivery system [
47]. Collectively, analysis of hearts of the MI271 therapy in the early ischemic phase revealed rather immunomodulatory fate of CD271
+ stem cells.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}