Multiple Sclerosis CD49d+CD154+ As Myelin-Specific Lymphocytes Induced During Remyelination

 ,
,  , ,
, ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Subjects
2.2. EAE Mouse Model
2.3. Human Cell Model of Progenitor Cells Differentiation to Mature Myelin Producing Oligodendrocytes
2.4. RR-MS CD49d+CD154+ Lymphocyte Sorting
2.5. hOPC-Lymphocyte Co-Culture
2.6. Analysis of RR-MS Myelin-Specific CD49d+CD154+ Cell Proliferation using CFSE Method
2.7. Flow Cytometry Analysis of CXCR4, CXCR7, CCR6 Surface Expression on CD49d+CD154+ Lymphocytes, and CD40 on OPCs
2.8. Chemotaxis Assay
2.9. Human Chemokine Multiple Profiling Assays
2.10. Mouse CNS Histopathological Examination
2.11. Immunohistochemistry (IHC)
2.12. Statistics
3. Results
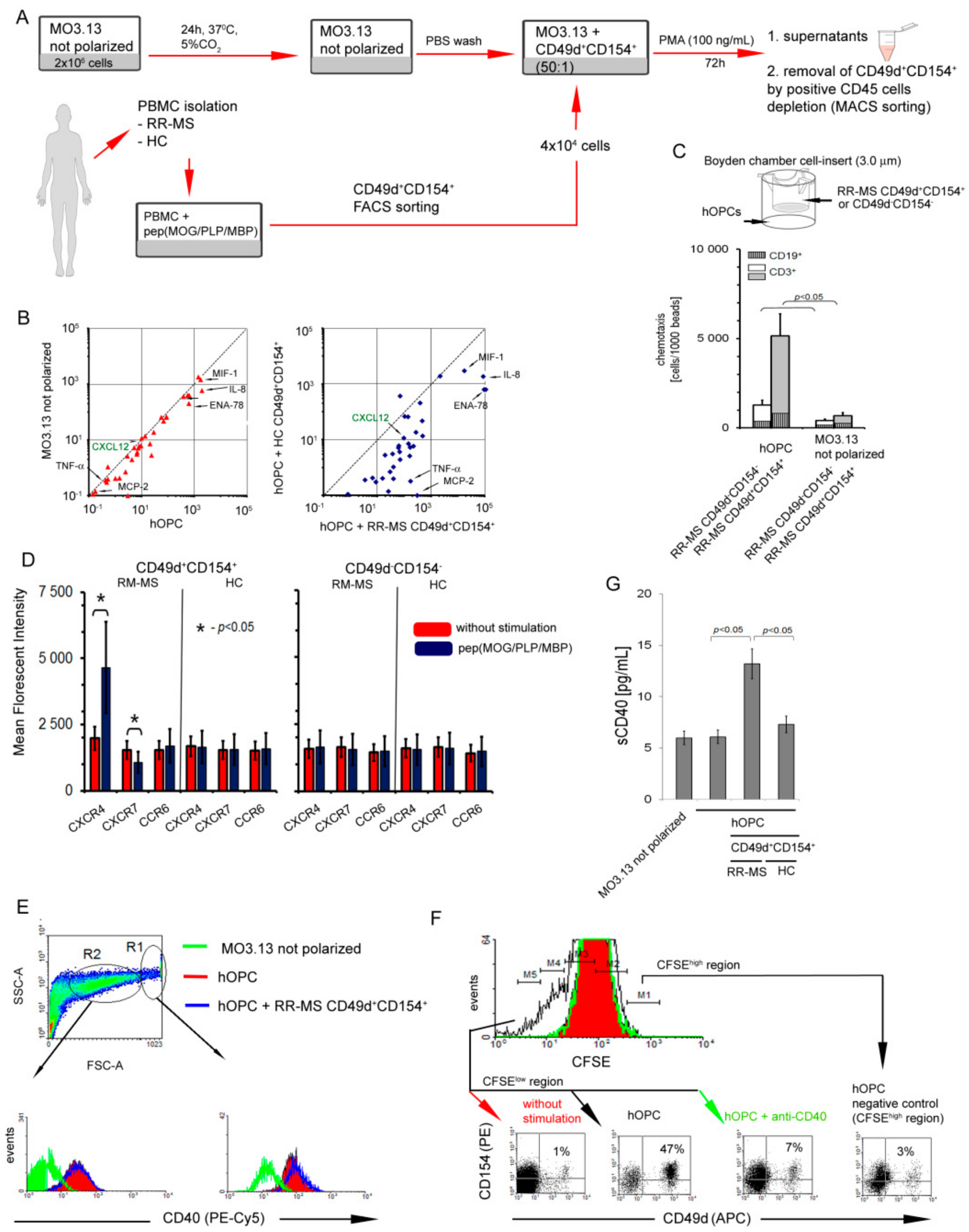
3.1. CD49d+CD154+ Lymphocytes of RR-MS Patients Proliferate in Vitro in Response to Myelin Proteins
3.2. Interaction of RR-MS CD49d+CD154+ Lymphocytes with Maturing Human OPCs (hOPCs) Generates Positive Proinflammatory Feedback Loop
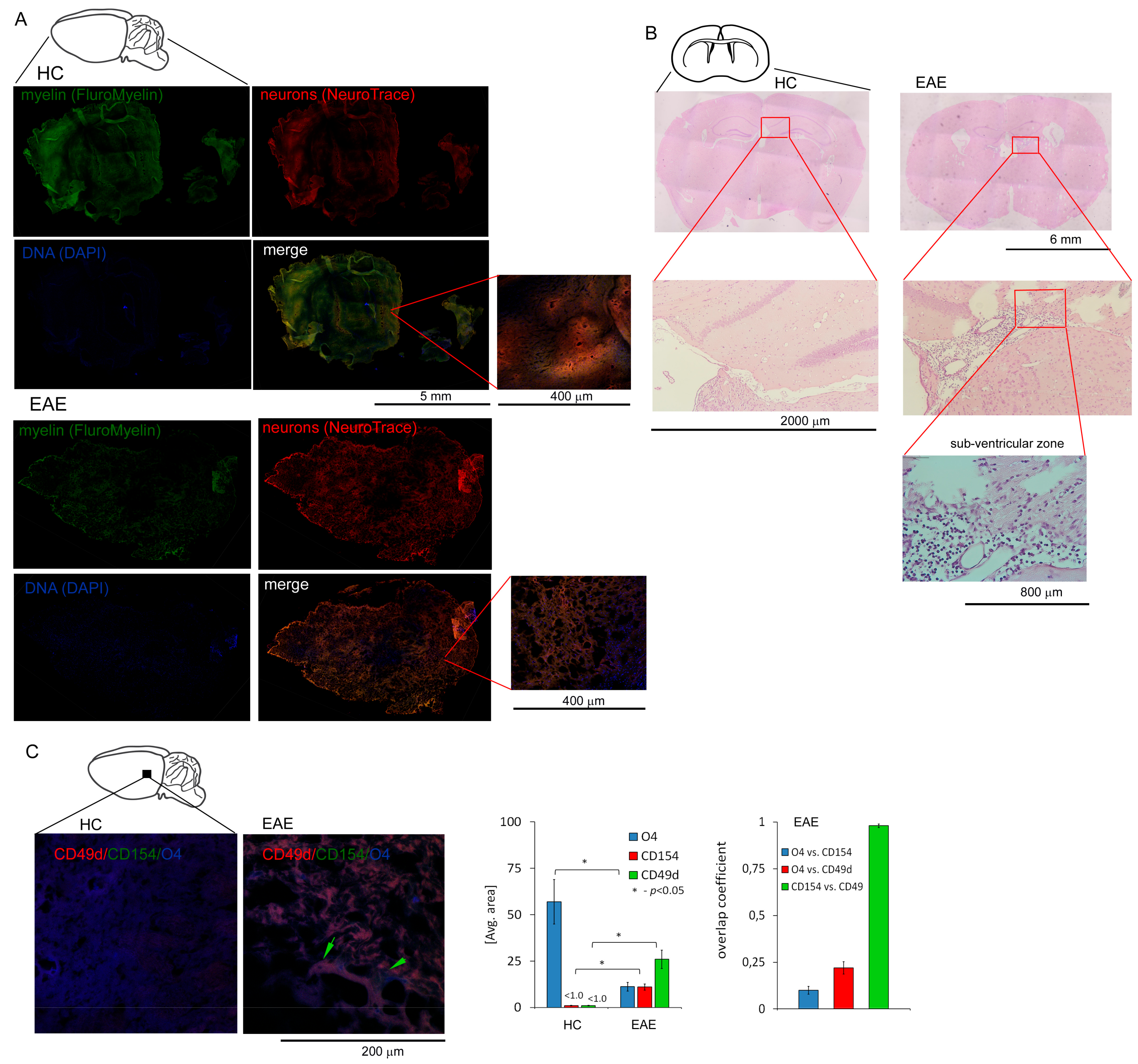
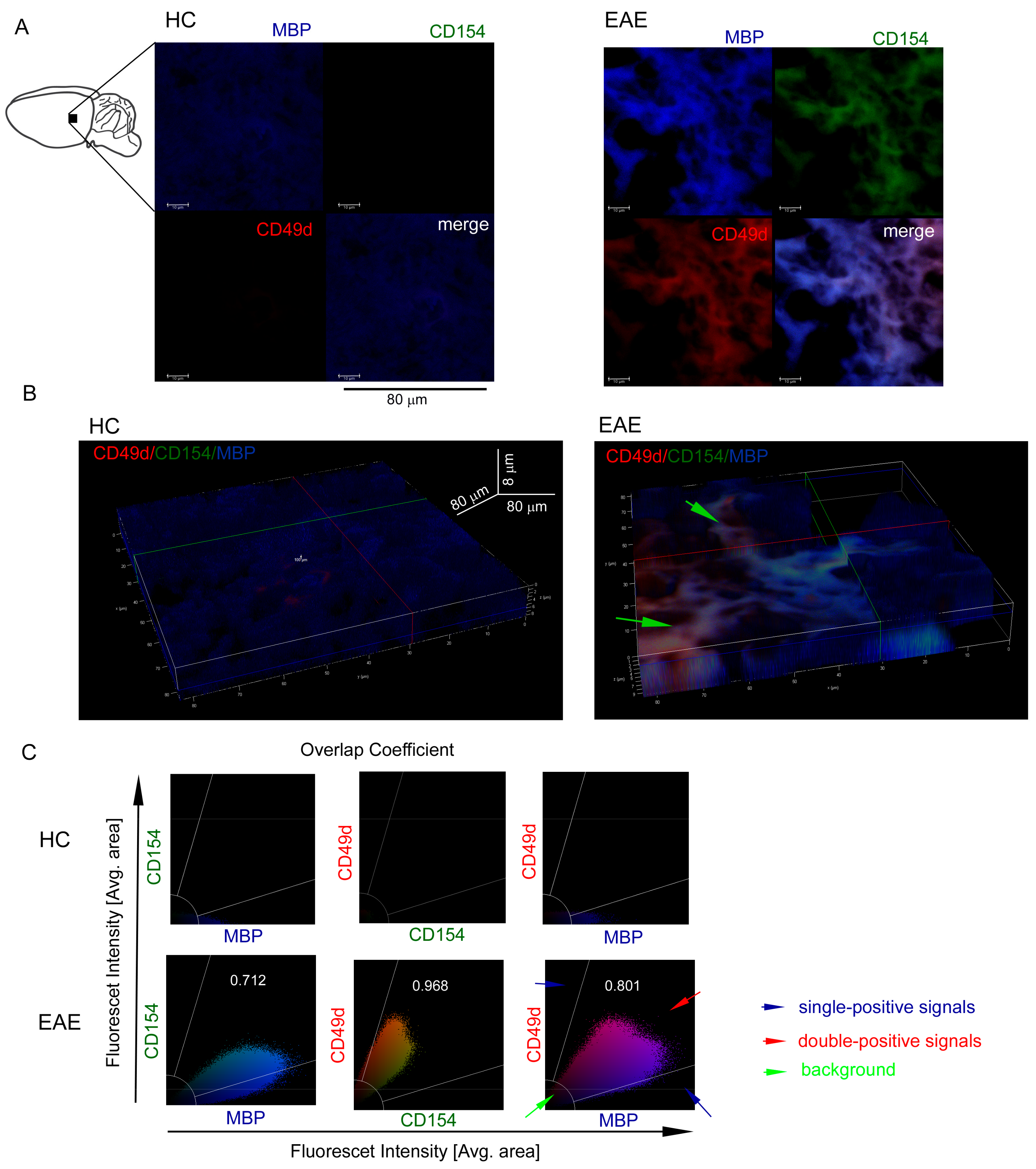
3.3. CD49d+CD154+ Lymphocytes are Present in the EAE Mouse Brain
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McFarland, H.F.; Martin, R. Multiple sclerosis: A complicated picture of autoimmunity. Nat. Immunol. 2007, 8, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Lublin, F.D.; Reingold, S.C.; Cohen, J.A.; Cutter, G.R.; Sørensen, P.S.; Thompson, A.J.; Wolinsky, J.S.; Balcer, L.J.; Banwell, B.; Barkhof, F.; et al. Defining the clinical course of multiple sclerosis: The 2013 revisions. Neurology 2014, 83, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Bramow, S.; Frischer, J.M.; Lassmann, H.; Koch-Henriksen, N.; Lucchinetti, C.F.; Sørensen, P.S.; Laursen, H. Demyelination versus remyelination in progressive multiple sclerosis. Brain 2010, 133, 2983–2998. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.; Tourtellotte, W.W.; Rudick, R.; Trapp, B.D. Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. N. Engl. J. Med. 2002, 346, 165–173. [Google Scholar] [CrossRef]
- Pierson, E.R.; Stromnes, I.M.; Goverman, J.M. B cells promote induction of experimental autoimmune encephalomyelitis by facilitating reactivation of T cells in the central nervous system. J. Immunol. 2014, 192, 929–939. [Google Scholar] [CrossRef]
- Prineas, J.W.; Connell, F. Remyelination in multiple sclerosis. Ann. Neurol. 1979, 5, 22–31. [Google Scholar] [CrossRef]
- Gerritse, K.; Laman, J.D.; Noelle, R.J.; Aruffo, A.; Ledbetter, J.A.; Boersma, W.J.; Claassen, E. CD40-CD40 ligand interactions in experimental allergic encephalomyelitis and multiple sclerosis. Proc. Natl. Acad. Sci. USA 1996, 93, 2499–2504. [Google Scholar] [CrossRef]
- Issazadeh, S.; Navikas, V.; Schaub, M.; Sayegh, M.; Khoury, S. Kinetics of expression of costimulatory molecules and their ligands in murine relapsing experimental autoimmune encephalomyelitis in vivo. J. Immunol. 1998, 161, 1104–1112. [Google Scholar]
- Aarts, S.A.B.M.; Seijkens, T.T.P.; van Dorst, K.J.F.; Dijkstra, C.D.; Kooij, G.; Lutgens, E. The CD40–CD40L dyad in experimental autoimmune encephalomyelitis and multiple sclerosis. Front. Immunol. 2017, 12, 1791. [Google Scholar] [CrossRef]
- Grewal, I.S.; Foellmer, H.G.; Grewal, K.D.; Xu, J.; Hardardottir, F.; Baron, J.L.; Janeway, C.A., Jr.; Flavell, R.A. Requirement for CD40 ligand in costimulation induction, T cell activation, and experimental allergic encephalomyelitis. Science 1996, 273, 1864–1867. [Google Scholar] [CrossRef]
- Abromson-Leeman, S.; Maverakis, E.; Bronson, R.; Dorf, M.E. CD40-mediated activation of T cells accelerates, but is not required for, encephalitogenic potential of myelin basic protein-recognizing T cells in a model of progressive experimental autoimmune encephalomyelitis. Eur. J. Immunol. 2001, 31, 527–538. [Google Scholar] [CrossRef]
- Laman, J.D.; Maassen, C.B.; Schellekens, M.M.; Visser, L.; Kap, M.; de Jong, E.; van Puijenbroek, M.; van Stipdonk, M.J.; van Meurs, M.; Schwärzler, C.; et al. Therapy with antibodies against CD40L (CD154) and CD44-variant isoforms reduces experimental autoimmune encephalomyelitis induced by a proteolipid protein peptide. Mult. Scler. J. 1998, 4, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Howard, L.M.; Miga, A.J.; Vanderlugt, C.L.; Dal Canto, M.C.; Laman, J.D.; Noelle, R.J.; Miller, S.D. Mechanisms of immunotherapeutic intervention by anti-CD40L (CD154) antibody in an animal model of multiple sclerosis. J. Clin. Invest 1999, 103, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Becher, B.; Durell, B.G.; Miga, A.V.; Hickey, W.F.; Noelle, R.J. The clinical course of experimental autoimmune encephalomyelitis and inflammation is controlled by the expression of CD40 within the central nervous system. J. Exp. Med. 2001, 193, 967–974. [Google Scholar] [CrossRef] [PubMed]
- International Multiple Sclerosis Genetics Consortium (IMSGC); Beecham, A.H.; Patsopoulos, N.A.; Xifara, D.K.; Davis, M.F.; Kemppinen, A.; Cotsapas, C.; Shah, T.S.; Spencer, C.; Booth, D.; et al. Wellcome Trust Case Control Consortium 2 (WTCCC2). Analysis of immune-related loci identifies 48 new susceptibility variants for multiple sclerosis. Nat. Genet. 2013, 45, 1353–1360. [Google Scholar]
- Wagner, M.; Wisniewski, A.; Bilinska, M.; Pokryszko-Dragan, A.; Cyrul, M.; Kusnierczyk, P.; Jasek, M. Investigation of gene-gene interactions between CD40 and CD40L in Polish multiple sclerosis patients. Hum. Immunol. 2014, 75, 796–801. [Google Scholar] [CrossRef]
- Ding, C.; Ireland, S.J.; Remington, G.; Alvarez, E.; Racke, M.K.; Greenberg, B.; Frohman, E.M.; Monson, N.L. CD40-mediated NF-κB activation in B cells is increased in multiple sclerosis and modulated by therapeutics. J. Immunol. 2016, 197, 4257–4265. [Google Scholar]
- Rommer, P.S.; Dudesek, A.; Stüve, O.; Zettl, U.K. Monoclonal antibodies in treatment of multiple sclerosis. Clin. Exp. Immunol. 2014, 175, 373–384. [Google Scholar] [CrossRef]
- Elovaara, I.; Ukkonen, M.; Leppäkynnäs, M.; Lehtimäki, T.; Luomala, M.; Peltola, J.; Dastidar, P. Adhesion molecules in multiple sclerosis: Relation to subtypes of disease and methylprednisolone therapy. Arch. Neurol. 2000, 57, 546–551. [Google Scholar] [CrossRef]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: Revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal research: Repoting in vivo experiments: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef] [PubMed]
- Buntinx, M.; Vanderlocht, J.; Hellings, N.; Vandenabeele, F.; Lambrichts, I.; Raus, J.; Ameloot, M.; Stinissen, P.; Steels, P. Characterization of three human oligodendroglial cell lines as a model to study oligodendrocyte injury: Morphology and oligodendrocyte-specific gene expression. J. Neurocytol. 2003, 32, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Givan, A.L. A flow cytometric assay for quantitation of rare antigen-specific T cells: Using cell-tracking dyes to calculate precursor frequencies for proliferation. Immunol. Invest 2007, 36, 563–580. [Google Scholar] [CrossRef] [PubMed]
- Lewkowicz, P.; Cwiklińska, H.; Mycko, M.P.; Cichalewska, M.; Domowicz, M.; Lewkowicz, N.; Jurewicz, A.; Selmaj, K.W. Dysregulated RNA-induced silencing complex (RISC) assembly within CNS corresponds with abnormal miRNA expression during autoimmune demyelination. J. Neurosci. 2015, 35, 7521–7537. [Google Scholar] [CrossRef]
- Clark, E.A.; Ledbetter, J.A. How B and T cells talk to each other. Nature 1994, 367, 425–428. [Google Scholar] [CrossRef]
- Chatzigeorgiou, A.; Lyberi, M.; Chatzilymperis, G.; Nezos, A.; Kamper, E. CD40/CD40L signaling and its implication in health and disease. Biofactors 2009, 35, 474–483. [Google Scholar] [CrossRef]
- Chu, T.; Shields, L.B.E.; Zhang, Y.P.; Feng, S.Q.; Shields, C.B.; Cai, J. CXCL12/CXCR4/CXCR7 chemokine axis in the central nervous system: Therapeutic targets for remyelination in demyelinating diseases. Neuroscientist 2017, 23, 627–648. [Google Scholar] [CrossRef]
- Reboldi, A.; Coisne, C.; Baumjohann, D.; Benvenuto, F.; Bottinelli, D.; Lira, S.; Uccelli, A.; Lanzavecchia, A.; Engelhardt, B.; Sallusto, F. C-C chemokine receptor 6-regulated entry of TH-17 cells into the CNS through the choroid plexus is required for the initiation of EAE. Nat. Immunol. 2009, 10, 514–523. [Google Scholar] [CrossRef]
- Falcão, A.M.; van Bruggen, D.; Marques, S.; Meijer, M.; Jäkel, S.; Agirre, E.; Floriddia, E.M.; Vanichkina, D.P.; ffrench-Constant, C.; Williams, A.; et al. Disease-specific oligodendrocyte lineage cells arise in multiple sclerosis. Nat Med. 2018, 24, 1837–1844. [Google Scholar] [CrossRef]
- Franklin, R.J.M.; ffrench-Constant, C. Regenerating CNS myelin—From mechanisms to experimental medicines. Nat. Rev. Neurosci. 2017, 18, 753–769. [Google Scholar] [CrossRef]
- Goldman, S.A.; Nedergaard, M.; Windrem, M.S. Glial progenitor cell-based treatment and modeling of neurological disease. Science 2012, 338, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Calderon, T.M.; Eugenin, E.A.; Lopez, L.; Kumar, S.S.; Hesselgesser, J.; Raine, C.S.; Berman, J.W. A role for CXCL12 (SDF-1α) in the pathogenesis of multiple sclerosis: Regulation of CXCL12 expression in astrocytes by soluble myelin basic protein. J. Neuroimmunol. 2006, 177, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Moll, N.M.; Cossoy, M.B.; Fisher, E.; Staugaitis, S.M.; Tucky, B.H.; Rietsch, A.M.; Chang, A.; Fox, R.J.; Trapp, B.D.; Ransohoff, R.M. Imaging correlates of leukocyte accumulation and CXCR4/CXCL12 in multiple sclerosis. Arch. Neurol. 2009, 66, 44–53. [Google Scholar] [CrossRef] [PubMed]
- McCandless, E.E.; Zhang, B.; Diamond, M.S.; Klein, R.S. CXCR4 antagonism increases T cell trafficking in the central nervous system and improves survival from West Nile virus encephalitis. Proc. Natl. Acad. Sci. USA 2008, 105, 11270–11275. [Google Scholar] [CrossRef]
- Balabanian, K.; Lagane, B.; Infantino, S.; Chow, K.Y.; Harriague, J.; Moepps, B.; Arenzana-Seisdedos, F.; Thelen, M.; Bachelerie, F. The chemokine SDF-1/CXCL12 binds to and signals through the orphan receptor RDC1 in T lymphocytes. J. Biol. Chem. 2005, 280, 35760–35766. [Google Scholar] [CrossRef]
- Jelcic, I.; Al Nimer, F.; Wang, J.; Lentsch, V.; Planas, R.; Jelcic, I.; Madjovski, A.; Ruhrmann, S.; Faigle, W.; Frauenknecht, K.; et al. Memory B cells activate brain-homing, autoreactive CD4+ T cells in multiple sclerosis. Cell 2018, 175, 85–100. [Google Scholar] [CrossRef]
- Mues, M.; Bartholomaus, I.; Thestrup, T.; Griesbeck, O.; Wekerle, H.; Kawakami, N.; Krishnamoorthy, G. Real-time in vivo analysis of T cell activation in the central nervous system using a genetically encoded calcium indicator. Nat. Med. 2013, 19, 778–783. [Google Scholar] [CrossRef]
- Piatek, P.; Namiecinska, M.; Domowicz, M.; Przygodzka, P.; Wieczorek, M.; Michlewska, S.; Lewkowicz, N.; Tarkowski, M.; Lewkowicz, P. MS CD49d+CD154+ lymphocytes reprogram oligodendrocytes into immune reactive cells affecting CNS regeneration. Cells 2019, 8, 1508. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PBMC (2 × 106 cells/mL) | ||
|---|---|---|
| RR-MS (n = 10) | HC (n = 10) | |
| CCL1 (I-309) | 22 ± 14.2 # | 6.8 ± 2.99 |
| CCL2 (MCP-1) | 552 ± 177.2 # | 66 ± 25.9 |
| CCL3 (MIP-1α) | <3.1 | <3.1 |
| CCL7 (MCP-3) | 134 ± 69.2 # | <3.2 |
| CCL8 (MCP-2) | 57 ± 23.5 # | <0.6 |
| CCL11(Eotaxin) | 34.8 ± 18.66 | 35.4 ± 13.33 |
| CCL13 (MCP-4) | <9.4 | <9.4 |
| CCL15 (MIP-1D) | <3.1 | <3.1 |
| CCL17 (TARC) | 27 ± 10.9 # | <3.4 |
| CCL19 (MIP-3β) | <9.1 | <9.1 |
| CCL20 (MIP-3α) | 26 ± 9.3 # | <8.7 |
| CCL21 (6Ckine) | 69 ± 24.9 # | <1.2 |
| CCL22 (MDC) | 124 ± 58.3 # | <5.9 |
| CCL23 (MPIF-1) | <3.5 | <3.5 |
| CCL24(Eotaxin-2) | 369. ± 81.1 # | 218 ± 67.1 |
| CCL25 (TECK) | 127 ± 39.0 # | 50.8 ± 25.1 |
| CCL26 (Eotaxin-3) | 9.2 ± 2.19 # | <1.4 |
| CCL27 (CTACK) | 39.4 ± 8.31 # | 0.8 ± 0.42 |
| CX3CL1 (Factalkine) | 109 ± 31.7 # | <9.6 |
| CXCL12 (SDFα+β) | 33 ± 12.0 # | <5.3 |
| CXCL1 (Gro-α) | 456 ± 119.34 # | 46.1 ± 27.11 |
| CXCL2 (Gro-β) | 153 ± 33.9 # | 12 ± 7.8 |
| CXCL5 (ENA-78) | 6205 ± 1842.3 # | 676 ± 227.4 |
| CXCL6 (GCP-2) | 18.9 ± 10.34 # | <3.1 |
| CXCL8 (IL-8) | 11206 ± 909.4 # | 867 ± 299.7 |
| CXCL9 (MIG) | 87 ± 44.7 # | <4.9 |
| CXCL10 (IP-10) | <2.6 | <2.6 |
| CXCL11 (I-TAC) | 59 ± 17.9 # | <2.9 |
| CXCL13 (BCA-1) | <0.1 | <0.1 |
| CXCL16 (SCYB16) | 76 ± 17.8 # | <13.4 |
| TNF-α | 39 ± 16.6 # | <3.1 |
| IFN-γ | 28.1 ± 3.2 # | <0.1 |
| IL-1β | 14 ± 3.9 # | 5 ± 3.7 |
| IL-2 | 3.0 ± 1.96 # | <0.4 |
| IL-4 | 7.1 ± 5.77 | 5.9 ± 3.08 |
| IL-6 | 28 ± 8.1 # | 8 ± 3.2 |
| MIF1 | 1071 ± 377.0 # | 408 ± 144.2 |
| IL-10 | <5.1 | <5.1 |
| IL-16 | 219 ± 83.1 | 213 ± 57.2 |
| GM-CSF | <1.5 | <1.5 |
| MO3.13 (2 × 106 cells/mL) | ||||
|---|---|---|---|---|
| Not Polarized | Polarized to OLs | Polarized to OLs + RR-MS CD49d+CD154+ | Polarized to OLs + HC CD49d+CD154+ | |
| CCL1 (I-309)) | 3.6 ± 2.21 | 6.8 ± 2.99 | 68.1 ± 18.06 * § | 10.4 ± 4.94 |
| CCL2 (MCP-1) | 391.6 ± 94.32 | 895 ± 209.9† | 977 ± 494.1 | 927 ± 200.3 |
| CCL3 (MIP-1α) | <3.1 | <3.1 | <3.1 | <3.1 |
| CCL7 (MCP-3) | <3.2 | <3.2 | <3.2 | <3.2 |
| CCL8 (MCP-2) | <0.6 | <0.6 | 62 ± 15.9 * § | <0.6 |
| CCL11(Eotaxin) | 64.7 ± 19.57 | 95.6 ± 26.47 | 137.0 ± 31.59 * § | 81.8 ± 21.60 |
| CCL13 (MCP-4) | <9.4 | <9.4 | <9.4 | <9.4 |
| CCL15 (MIP-1D) | <3.1 | <3.1 | <3.1 | <3.1 |
| CCL17 (TARC) | <3.4 | <3.4 | <3.4 | <3.4 |
| CCL19 (MIP-3β) | <9.1 | <9.1 | <9.1 | <9.1 |
| CCL20 (MIP-3α) | <8.7 | <8.7 | 99 ± 18.7 * § | <8.7 |
| CCL21 (6Ckine) | 18 ± 4.9 | 29 ± 10.7 | 76.5 ± 18.2 * § | 26 ± 12.94 |
| CCL22 (MDC) | <5.9 | <5.9 | 23 ± 12.1 * § | <5.9 |
| CCL23 (MPIF-1) | <3.5 | <3.5 | <3.5 | <3.5 |
| CCL24(Eotaxin-2) | <1.1 | <1.1 | 71 ± 27.1 * § | 12 ± 11.9* |
| CCL25 (TECK) | 46 ± 18.5 | 69 ± 23.3 | 263 ± 83.3 * § | 79. ± 20.8 |
| CCL26 (Eotaxin-3) | <1.4 | <1.4 | 16.8 ± 2.60 * § | <1.4 |
| CCL27 (CTACK) | 0.4 ± 0.33 | 0.5 ± 0.37 | 29.5 ± 15.19 * § | 1.5 ± 1.25 |
| CX3CL1 (Factalkine) | 57 ± 10.48 | 89 ± 11.8† | 506 ± 46.4 * § | 79 ± 30.9 |
| CXCL12 (SDFα+β) | <5.3 | <5.3 | 264 ± 59.2 * § | <5.3 |
| CXCL1 (Gro-α) | 208.1 ± 79.34 | 347.5 ± 127.78† | 1533.4 ± 301.68 * § | 365.9 ± 195.19 |
| CXCL2 (Gro-β) | 7 ± 3.9 | 21 ± 8.8† | 217.9 ± 185.66 * § | 33 ± 13.5 |
| CXCL5 (ENA-78) | 296. ± 164.8 | 862 ± 227.4† | 10,432 ± 1129.3 * § | 985 ± 395.2 |
| CXCL6 (GCP-2) | 64.8 ± 23.95 | 54.9 ± 27.33 | 329.9 ± 105.99 * § | 64.0 ± 28.58 |
| CXCL8 (IL-8) | 913. ± 209.0 | 3235 ± 998.6† | 16,896 ± 5981.1 * § | 3626 ± 672.1 |
| CXCL9 (MIG) | <4.9 | <4.9 | 69 ± 13.9 * § | <4.9 |
| CXCL10 (IP-10) | <2.6 | <2.6 | <2.6 | <2.6 |
| CXCL11 (I-TAC) | <2.9 | <2.9 | <2.9 | <2.9 |
| CXCL13 (BCA-1) | <0.1 | <0.1 | <0.1 | <0.1 |
| CXCL16 (SCYB16) | 360 ± 178.2 | 383 ± 166.1 | 388 ± 258.7 | 342 ± 187.0 |
| TNF-α | <3.1 | <3.1 | 36.5 ± 12.22 * § | <3.1 |
| IFN-γ | <0.1 | <0.1 | <0.1 | <0.1 |
| IL-1β | <0.3 | <0.3 | 25.8 ± 12.33* § | <0.3 |
| IL-2 | <0.4 | <0.4 | <0.4 | <0.4 |
| IL-4 | <0.4 | <0.4 | <0.4 | <0.4 |
| IL-6 | 3 ± 1.8 | 24 ± 9.8† | 259 ± 13.2 * § | 27 ± 10.2 |
| MIF1 | 1931 ± 822.2 | 2735 ± 767.9† | 3687 ± 872.8 * § | 2708. ± 772.4 |
| IL-10 | <5.1 | <5.1 | <5.1 | <5.1 |
| IL-16 | <13.7 | <13.7 | <13.7 | <13.7 |
| GM-CSF | <1.5 | <1.5 | <1.5 | <1.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piatek, P.; Namiecinska, M.; Domowicz, M.; Wieczorek, M.; Michlewska, S.; Matysiak, M.; Lewkowicz, N.; Tarkowski, M.; Lewkowicz, P. Multiple Sclerosis CD49d+CD154+ As Myelin-Specific Lymphocytes Induced During Remyelination. Cells 2020, 9, 15. https://doi.org/10.3390/cells9010015
Piatek P, Namiecinska M, Domowicz M, Wieczorek M, Michlewska S, Matysiak M, Lewkowicz N, Tarkowski M, Lewkowicz P. Multiple Sclerosis CD49d+CD154+ As Myelin-Specific Lymphocytes Induced During Remyelination. Cells. 2020; 9(1):15. https://doi.org/10.3390/cells9010015
Chicago/Turabian StylePiatek, Paweł, Magdalena Namiecinska, Małgorzata Domowicz, Marek Wieczorek, Sylwia Michlewska, Mariola Matysiak, Natalia Lewkowicz, Maciej Tarkowski, and Przemysław Lewkowicz. 2020. "Multiple Sclerosis CD49d+CD154+ As Myelin-Specific Lymphocytes Induced During Remyelination" Cells 9, no. 1: 15. https://doi.org/10.3390/cells9010015
APA StylePiatek, P., Namiecinska, M., Domowicz, M., Wieczorek, M., Michlewska, S., Matysiak, M., Lewkowicz, N., Tarkowski, M., & Lewkowicz, P. (2020). Multiple Sclerosis CD49d+CD154+ As Myelin-Specific Lymphocytes Induced During Remyelination. Cells, 9(1), 15. https://doi.org/10.3390/cells9010015