Hepatocellular Toxicity of Paris Saponins I, II, VI and VII on Two Kinds of Hepatocytes-HL-7702 and HepaRG Cells, and the Underlying Mechanisms

 , ,
, ,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
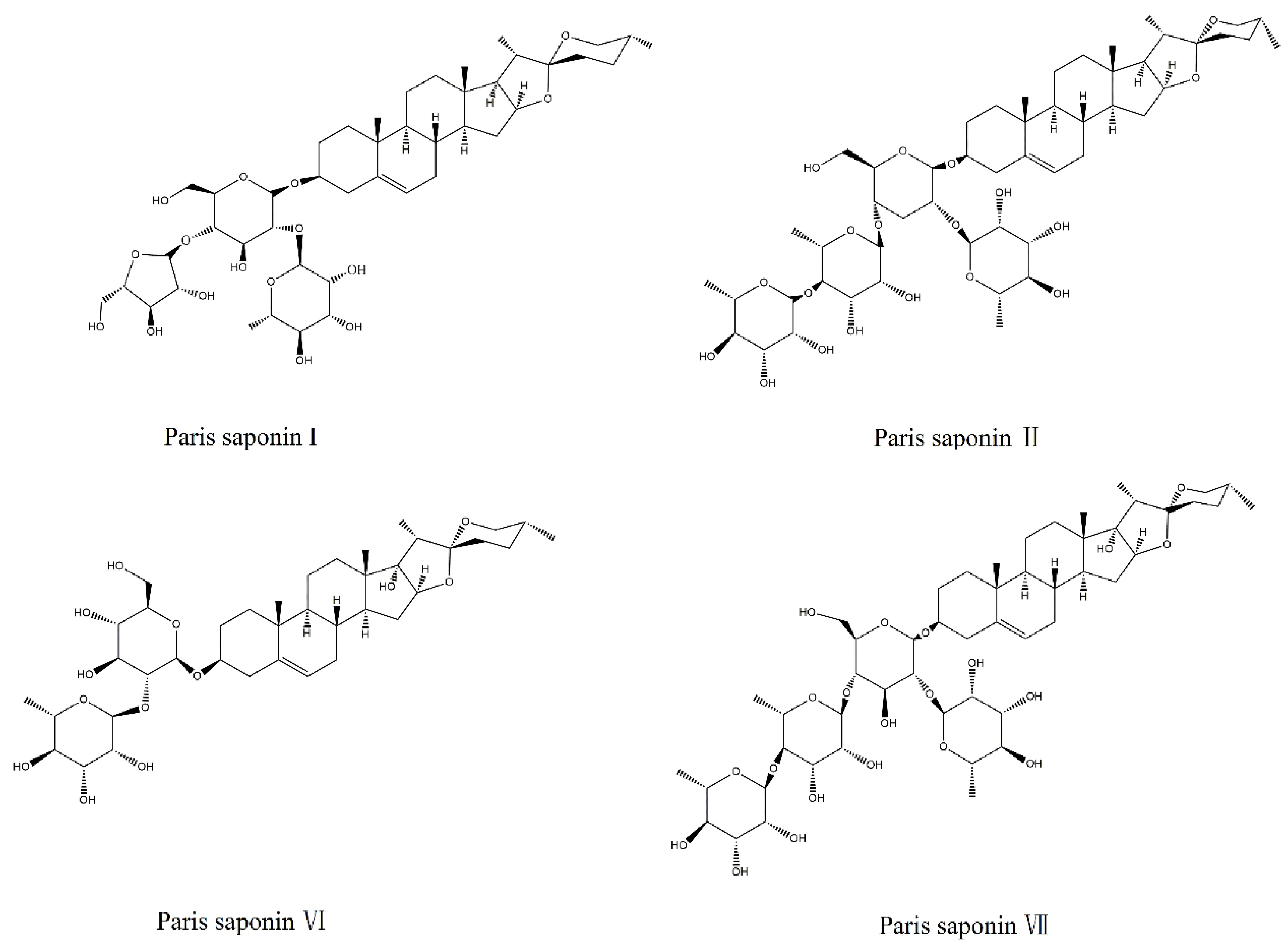
2.1. Drugs and Reagents
2.2. Cell Culture Conditions
2.3. Comparison of Cytotoxicities of the Four Saponins in Rhizoma Paridis on HL-7702 Cells
2.4. Comparison of Cytotoxicities of the Four Saponins in Rhizoma Paridis to HepaRG Cells
2.5. LDH Assay
2.6. DAPI Staining
2.7. Determination of the Effect of PSI on Apoptosis and Necrosis
2.8. Determination of Effect of PSI on Intracellular ROS Levels
2.9. Determination of the Effect of PSI on Mitochondrial Transmembrane Potential (∆ψ)
2.10. Cell Cycle Analysis
2.11. Assay of Effect of PSI on Expressions of Relevant Proteins Using Western-Blot
2.12. Statistical Analysis
3. Results
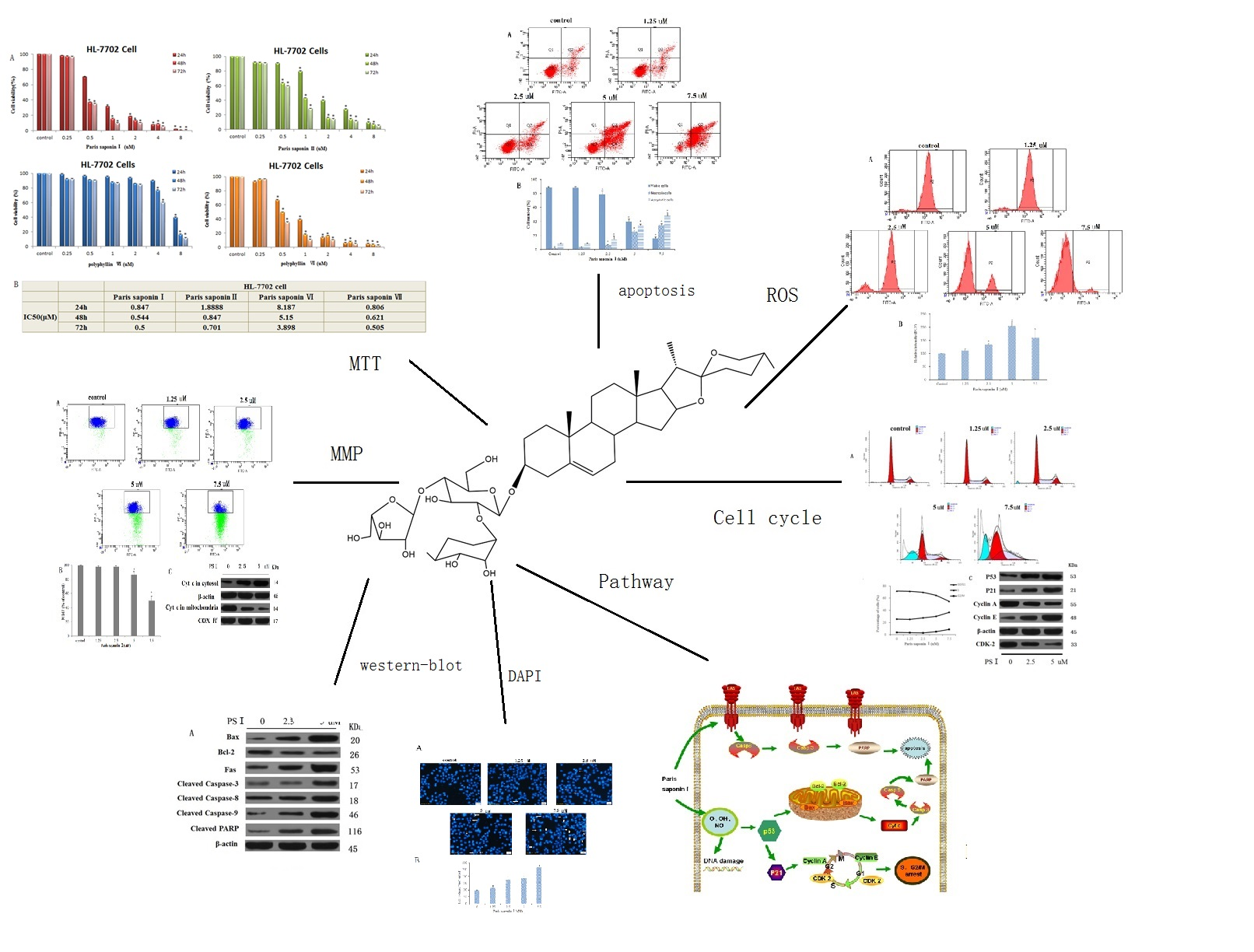
3.1. Comparison of Cytotoxicities of the Four Saponins of Rhizoma Paridis on HL-7702 Cells
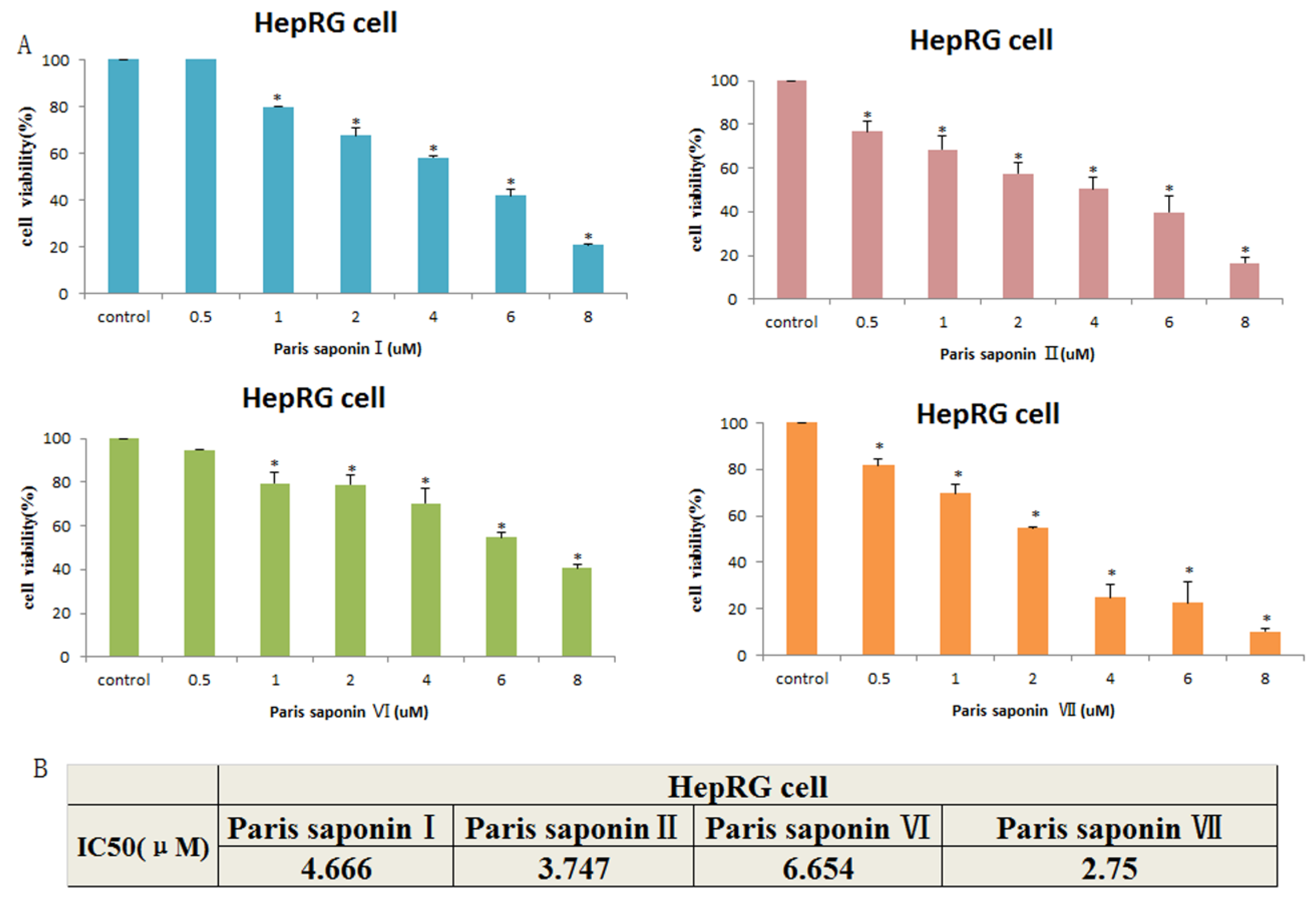
3.2. Cytotoxic Effect of the Four Saponins on HepaRG Cells
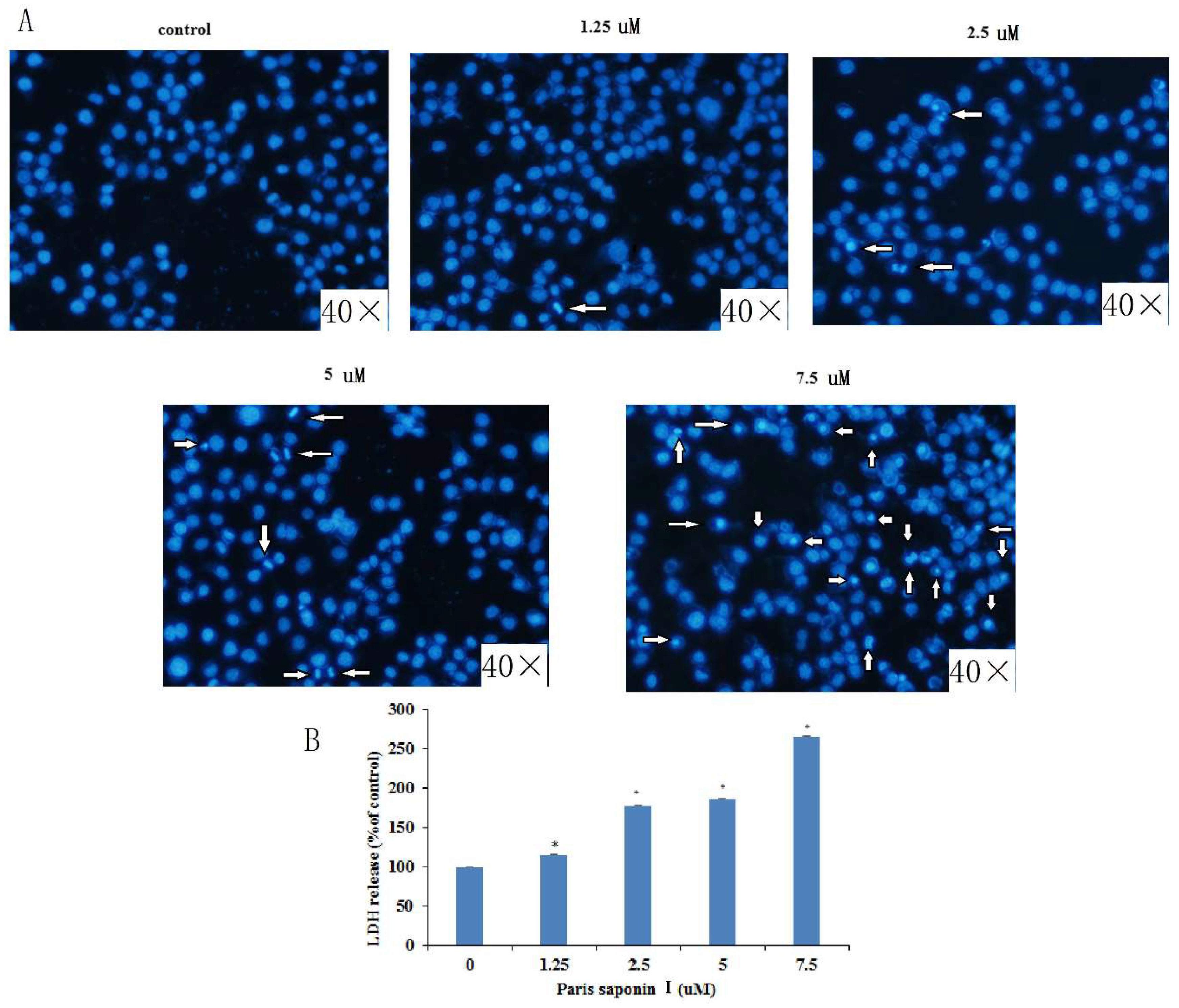
3.3. LDH release and DAPI Staining
3.4. Effect of PSI on the Apoptosis and Necrosis of HepaRG Cells
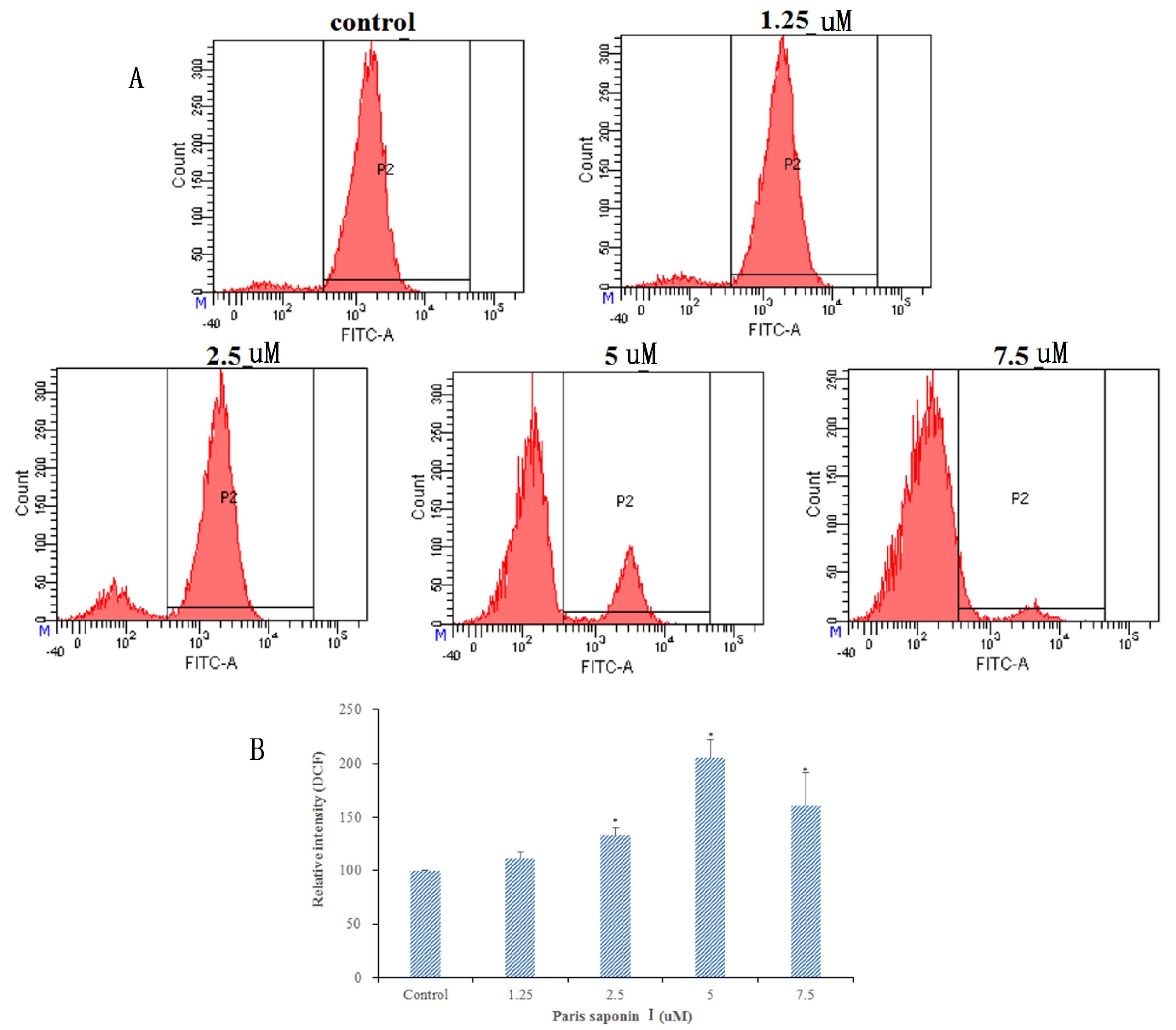
3.5. Effects of PSI on ROS Generation
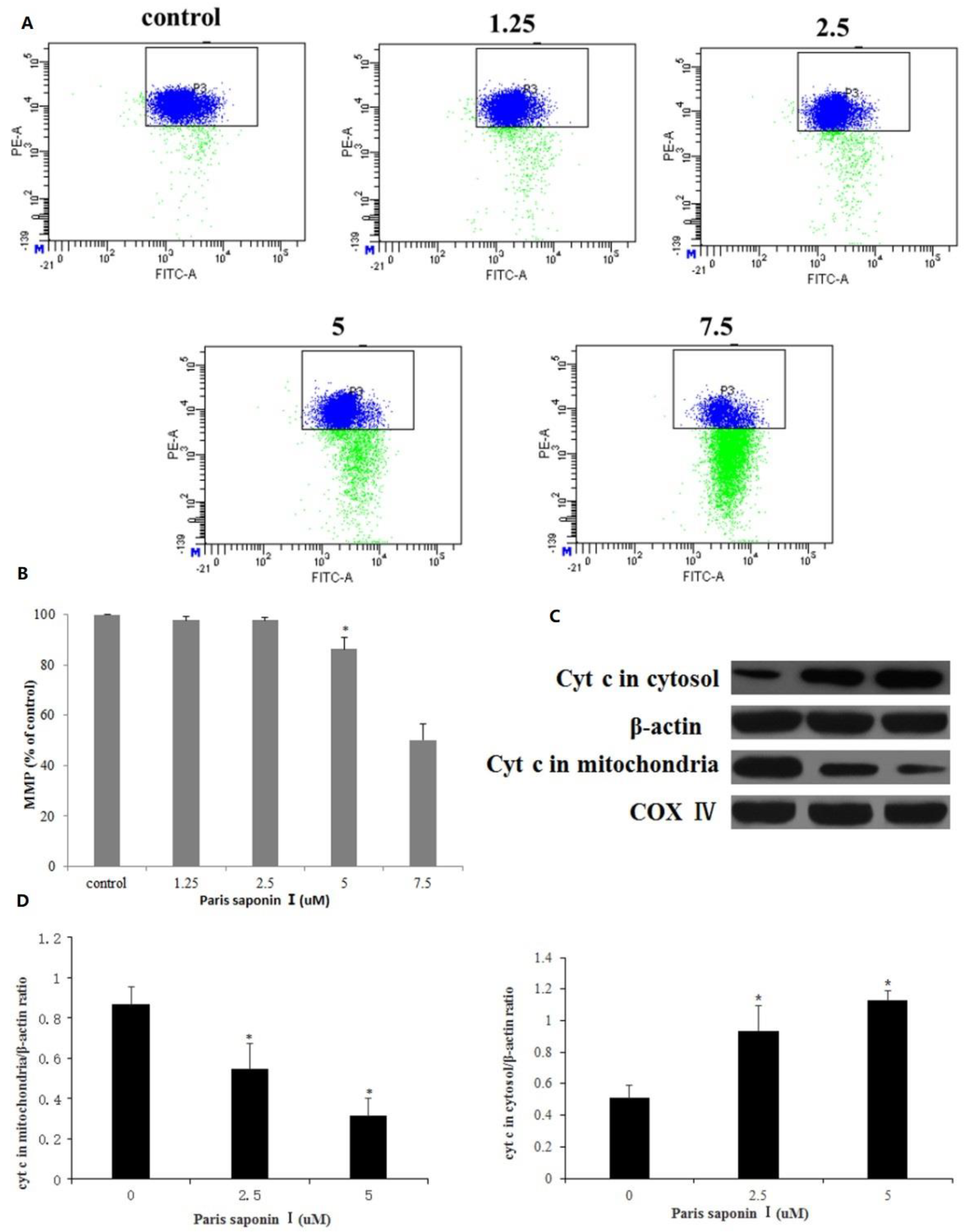
3.6. Effect of PSI on MMP and Cytochrome c Released from HepaRG Cells
3.7. Effect of PSI on Cell Cycle Distribution
3.8. Effect of PSI on the Expression of Apoptosis-Related Proteins in HepaRG Cells
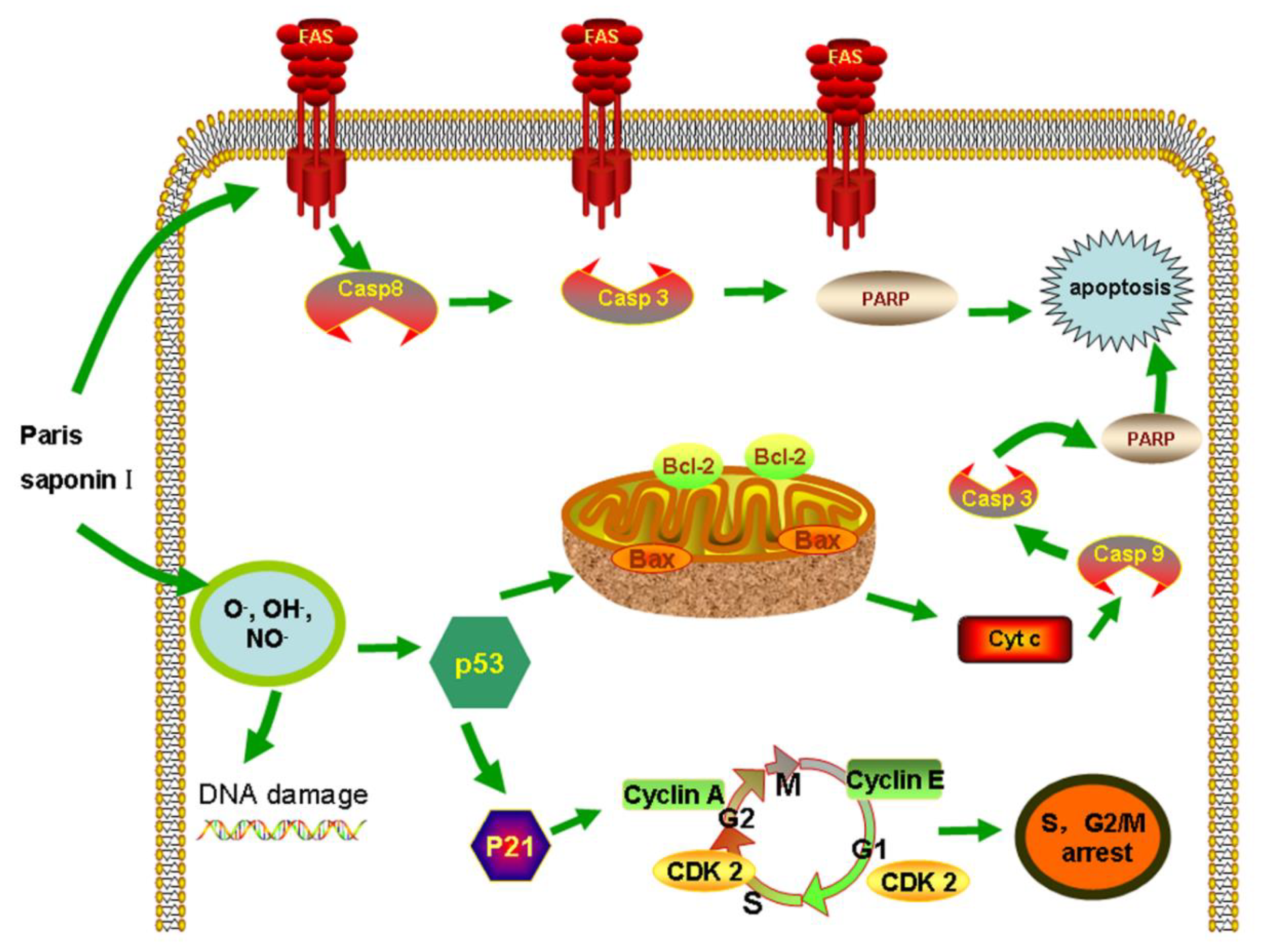
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Negi, J.S.; Bisht, V.K.; Bhandari, A.K.; Bhatt, V.P.; Singh, P.; Singh, N. Paris polyphylla: Chemical and biological prospectives. Anti-Cancer Agents Med. Chem. 2014, 14, 833–839. [Google Scholar] [CrossRef]
- Jing, S.; Wang, Y.; Li, X.; Man, S.; Gao, W. Chemical constituents and antitumor activity from paris polyphylla smith var. yunnanensis. Nat. Product Res. 2016, 31, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gao, W.; Man, S.; Wang, J.; Li, N.; Yin, S.; Liu, C. Pharmacological evaluation of sedative–hypnotic activity and gastro-intestinal toxicity of Rhizoma Paridis saponins. J. Ethnopharmacol. 2012, 144, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.N.; Xv, L.; Shi, L.; Li, Y.; Lin, L.F. Advances in toxicological studies on liver injury caused by common saponins. Cent. South Pharm. 2010, 8, 916–919. [Google Scholar]
- Zhou, Y.L.; Peng, L.L.; Wu, T.K.; Chen, C.X.; Zhou, S.Q. Study on the toxicity of total saponins from Rhizoma Paridis. Sichuan J. Physiol. 1988, 4, 51–52. [Google Scholar]
- Li, J.; Man, S.; Qiu, P.; Fan, W.; Zhang, L.; Gao, W. Toxicological risks of Rhizoma paridis saponins in rats involved NF-κB and Nrf2 signaling. RSC Adv. 2016, 6, 31889–31897. [Google Scholar] [CrossRef]
- Man, S.; Qiu, P.; Li, J.; Zhang, L.; Gao, W. Global metabolic profiling for the study of Rhizoma Paridis saponins-induced hepatotoxicity in rats. Environ. Toxicol. 2017, 32, 99–108. [Google Scholar] [CrossRef]
- Lee, M.S.; Chan, J.Y.W.; Kong, S.K.; Yu, B.; Eng-Choon, V.O.; Nai-Ching, H.W.; Fung, K.P. Effects of polyphyllin D, a steroidal saponin in Paris polyphylla, in growth inhibition of human breast cancer cells and in xenograft. Cancer Biol. Ther. 2005, 4, 1248–1254. [Google Scholar] [CrossRef]
- Xiao, X.; Zou, J.; Bui-Nguyen, T.M.; Bai, P.; Gao, L.; Liu, J.; Wang, H. Paris saponin II of Rhizoma Paridis–a novel inducer of apoptosis in human ovarian cancer cells. Biosci. Trends 2012, 4, 201–211. [Google Scholar] [CrossRef]
- Shi, Y.M.; Yang, L.; Geng, Y.D.; Zhang, C.; Kong, L.Y. Polyphyllin I induced-apoptosis is enhanced by inhibition of autophagy in human hepatocellular carcinoma cells. Phytomedicine 2015, 22, 1139–1149. [Google Scholar] [CrossRef]
- Zhao, P.J.; Song, S.C.; Du, L.W.; Zhou, G.H.; Ma, S.L.; Li, J.H.; Jiang, H. Paris Saponins enhance radiosensitivity in a gefitinib-resistant lung adenocarcinoma cell line by inducing apoptosis and G2/M cell cycle phase arrest. Mol. Med. Reports 2016, 13, 2878–2884. [Google Scholar] [CrossRef] [PubMed]
- Tong, Q.Y.; He, Y.; Zhao, Q.B.; Qing, Y.; Huang, W.; Wu, X.H. Cytotoxicity and apoptosis-inducing effect of steroidal saponins from Dioscorea zingiberensis Wright against cancer cells. Steroids 2012, 77, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Anthérieu, S.; Chesné, C.; Li, R.; Guguen-Guillouzo, C.; Guillouzo, A. Optimization of the HepaRG cell model for drug metabolism and toxicity studies. Toxicol. Vitro 2012, 26, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- McGill, M.R.; Yan, H.M.; Ramachandran, A.; Murray, G.J.; Rollins, D.E.; Jaeschke, H. HepaRG cells: A human model to study mechanisms of acetaminophen hepatotoxicity. Hepatology 2011, 53, 974–982. [Google Scholar] [CrossRef] [PubMed]
- Gerets, H.H.J.; Tilmant, K.; Gerin, B.; Chanteux, H.; Depelchin, B.; Dhalluin, S.; Atienzar, F.A. Characterization of primary human hepatocytes, HepG2 cells, and HepaRG cells at the mRNA level and CYP activity in response to inducers and their predictivity for the detection of human hepatotoxins. Cell Bio. Toxicol. 2012, 28, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.; Rafiq, M.T.; Yang, J.; Liu, D.; Lu, L.; He, Z.; Yang, X. Impact assessment of cadmium toxicity and its bioavailability in human cell lines (Caco-2 and HL-7702). BioMed Res. Int. 2014, 839538. [Google Scholar] [CrossRef]
- Guo, J.; Song, W.; Ding, F.; Zhang, J.; Sun, Z. Study on cytotoxicity and structure–activity relationship of HL-7702 cell exposed to naphthoquinones. Environ. Toxicol. Pharmacol. 2012, 33, 408–413. [Google Scholar] [CrossRef]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef]
- Candé, C.; Cecconi, F.; Dessen, P.; Kroemer, G. Apoptosis-inducing factor (AIF): Key to the conserved caspase-independent pathways of cell death? J. Cell Sci. 2002, 115, 4727–4734. [Google Scholar] [CrossRef]
- Liu, L.; Fu, J.; Li, T.; Cui, R.; Ling, J.; Yu, X.; Zhang, Y. NG, a novel PABA/NO-based oleanolic acid derivative, induces human hepatoma cell apoptosis via a ROS/MAPK-dependent mitochondrial pathway. Eur. J. Pharmacol. 2012, 691, 61–68. [Google Scholar] [CrossRef]
- Ola, M.S.; Nawaz, M.; Ahsan, H. Role of Bcl-2 family proteins and caspases in the regulation of apoptosis. Mol. Cell. Biochem. 2011, 351, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Oltval, Z.N.; Milliman, C.L.; Korsmeyer, S.J. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programed cell death. Cell 1993, 74, 609–619. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. The Bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Kushwaha, H.N.; Srivastava, A.K.; Srivastava, S.; Jamal, N.; Srivastava, K.; Ray, R.S. Piperine attenuates UV-R induced cell damage in human keratinocytes via NF-kB, Bax/Bcl-2 pathway: An application for photoprotection. J. Photochem. Photobiol. B Biol. 2017, 172, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Ong, R.C.; Lei, J.; Lee, R.K.; Cheung, J.Y.; Fung, K.P.; Lin, C.; Kong, S.K. Polyphyllin D induces mitochondrial fragmentation and acts directly on the mitochondria to induce apoptosis in drug-resistant HepG2 cells. Cancer Lett. 2008, 261, 158–164. [Google Scholar] [CrossRef]
- Man, S.; Fan, W.; Liu, Z.; Gao, W.; Li, Y.; Zhang, L.; Liu, C. Antitumor pathway of Rhizoma Paridis Saponins based on the metabolic regulatory network alterations in H22 hepatocarcinoma mice. Steroids 2014, 84, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.J.; Mulholland, D.J.; Valamehr, B.; Mosessian, S.; Sellers, W.R.; Wu, H. PTEN nuclear localization is regulated by oxidative stress and mediates p53-dependent tumor suppression. Mol. Cell. Biol. 2008, 28, 3281–3289. [Google Scholar] [CrossRef]
- Li, H.; Zhu, H.; Xu, C.J.; Yuan, J. Cleavage of BID by caspase 8 mediates the mitochondrial damage in the Fas pathway of apoptosis. Cell 1998, 94, 491–501. [Google Scholar] [CrossRef]
- Ye, W.; Zhu, S.; Liao, C.; Xiao, J.; Wu, Q.; Lin, Z.; Chen, J. Advanced oxidation protein products induce apoptosis of human chondrocyte through reactive oxygen species-mediated mitochondrial dysfunction and endoplasmic reticulum stress pathways. Fundam. Clin. Pharmacol. 2017, 31, 64–74. [Google Scholar] [CrossRef]
- Xiao, X.; Bai, P.; Nguyen, T.M.B.; Xiao, J.; Liu, S.; Yang, G.; Wang, H. The antitumoral effect of Paris Saponin I associated with the induction of apoptosis through the mitochondrial pathway. Mol. Cancer Ther 2009, 8, 1179–1188. [Google Scholar] [CrossRef]
- Zhang, C.; Jia, X.; Bao, J.; Chen, S.; Wang, K.; Zhang, Y.; Mei, Z. Polyphyllin VII induces apoptosis in HepG2 cells through ROS-mediated mitochondrial dysfunction and MAPK pathways. BMC Complement. Altern. Med. 2015, 16, 58. [Google Scholar] [CrossRef] [PubMed]
- Siu, F.M.; Ma, D.L.; Cheung, Y.W.; Lok, C.N.; Yan, K.; Yang, Z.; Che, C.M. Proteomic and transcriptomic study on the action of a cytotoxic saponin (Polyphyllin D): Induction of endoplasmic reticulum stress and mitochondria-mediated apoptotic pathways. Proteomics 2008, 8, 3105–3117. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Du, L.; Jiang, H.; Zhu, X.; Li, J.; Xu, J. Paris saponin I sensitizes gastric cancer cell lines to cisplatin via cell cycle arrest and apoptosis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2016, 22, 3798. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.Y.N.; Ong, R.C.Y.; Suen, Y.K.; Ooi, V.; Wong, H.N.C.; Mak, T.C.W.; Kong, S.K. Polyphyllin D is a potent apoptosis inducer in drug-resistant HepG2 cells. Cancer Lett. 2005, 217, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Suzuki, T.; Hara, F.; Yasui, T.; Uga, N.; Naoe, A. Polyphyllin D, a steroidal saponin in Paris polyphylla, induces apoptosis and necroptosis cell death of neuroblastoma cells. Pediatric Surg. Int. 2017, 33, 713–719. [Google Scholar] [CrossRef]
- Liu, J.; Man, S.; Li, J.; Zhang, Y.; Meng, X.; Gao, W. Inhibition of diethylnitrosamine-induced liver cancer in rats by Rhizoma paridis saponin. Environ. Toxicol. pharmacol. 2016, 46, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Su, D.; Ma, S.L. The Effect of Chonglou Saponin I on Proliferation and Apoptosis in Lung Adenocarcinoma Cell Line PC9. J. Chin. Oncol. 2012, 18, 166–169. [Google Scholar]
- Wu, Z.; Zhang, J.; Xu, F.; Wang, Y.; Zhang, J. Rapid and simple determination of polyphyllin i, ii, vi, and vii in different harvest times of cultivated Paris polyphylla Smith var. yunnanensis (Franch.) Hand.-Mazz by UPLC-MS/MS and FT-IR. J. Nat. Med. 2017, 71, 139–147. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Liu, D.H.; Peng, H.S.; Hao, Q.X.; Chen, M.L.; Zhang, A.Z.; Huang, L.Q. Determination of eight steroidal saponins in 15 kinds of genus Paris. China J. Chin. Mater. Med. 2017, 42, 3443–3451. [Google Scholar]
- Chen, M.; Ye, K.; Zhang, B.; Xin, Q.; Li, P.; Kong, A.N.; Yang, J. Paris Saponin II inhibits colorectal carcinogenesis by regulating mitochondrial fission and NF-κB pathway. Pharmacol. Res. 2019, 139, 273–285. [Google Scholar] [CrossRef]
- Yan, L.L.; Zhang, Y.J.; Gao, W.Y.; Man, S.L.; Wang, Y. In vitro and in vivo anticancer activity of steroid saponins of paris polyphylla var. yunnanensis. Exp. Oncol. 2009, 31, 27–32. [Google Scholar] [PubMed]
- Sun, C.L.; Ni, W.; Yan, H.; Liu, Z.H.; Yang, L.; Si, Y.A.; Liu, H.Y. Steroidal saponins with induced platelet aggregation activity from the aerial parts of Paris verticillata. Steroids 2014, 92, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.J.; Sun, D.J.; Ni, W.; Chen, C.X.; Hua, Y.; He, L. Steroidal saponins with antimicrobial activity from stems and leaves of paris polyphylla var. yunnanensis. Steroids 2012, 77, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhou, X.; Zhao, Y.; Lu, Y.; Shi, Q.; Wang, Y.; Liang, Q.Q. Polyphyllin I ameliorates collagen-induced arthritis by suppressing the inflammation response in macrophages through the NF-κB pathway. Front. Immunol. 2018, 9, 2091. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.Y.W.; Koon, J.C.M.; Liu, X.; Detmar, M.; Yu, B.; Kong, S.K.; Fung, K.P. Polyphyllin D, a steroidal saponin from Paris polyphylla, inhibits endothelial cell functions in vitro and angiogenesis in zebrafish embryos in vivo. J. Ethnopharmacol. 2011, 137, 64–69. [Google Scholar] [CrossRef] [PubMed]
- GuangLie, C.; WeiShi, G.; GaiLing, H.; JianPing, C. Effect of Paris saponin on antitumor and immune function in U14 tumor-bearing mice. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Zou, P.; Tang, Q.; Zheng, F.; Wu, J.; Chen, Z.; Hann, S.S. HOTAIR-mediated reciprocal regulation of EZH2 and DNMT1 contribute to polyphyllin I-inhibited growth of castration-resistant prostate cancer cells in vitro and in vivo. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 589–599. [Google Scholar] [CrossRef]
- Liu, Z.; Zheng, Q.; Chen, W.; Wu, M.; Gao, W. Chemosensitizing effect of paris saponin I on camptothecin and 10-hydroxycamptothecin in lung cancer cells via p38 mapk, erk, and akt signaling pathways. Eur. J. Med. Chem. 2016, 125, 760–769. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. BBA Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Li, S.; Cheng, B.; Hou, L.; Huang, L.; Cui, Y.; Xu, D.; Li, S. Dioscin inhibits colon cancer cells’ growth by reactive oxygen species-mediated mitochondrial dysfunction and p38 and JNK pathways. Anti-cancer Drugs 2018, 29, 234–242. [Google Scholar] [CrossRef]
- Lou, Z.; Li, X.; Zhao, X.; Du, K.; Li, X.; Wang, B. Resveratrol attenuates hydrogen peroxide-induced apoptosis, reactive oxygen species generation, and PSGL-1 and VWF activation in human umbilical vein endothelial cells, potentially via MAPK signalling pathways. Mol. Med. Reports 2018, 17, 2479–2487. [Google Scholar] [CrossRef] [PubMed]
- Wang, X. The expanding role of mitochondria in apoptosis. Genes Dev. 2001, 15, 2922–2933. [Google Scholar] [PubMed]
- Li, H.; Chen, J.; Xiong, C.; Wei, H.; Yin, C.; Ruan, J. Apoptosis induction by the total flavonoids from arachniodes exilis in hepg2 cells through reactive oxygen species-mediated mitochondrial dysfunction involving mapk activation. Evid. Based Complement. Altern. Med. 2014, 2014, 1–11. [Google Scholar]
- Jiang, H.; Zhao, P.J.; Su, D.; Feng, J.; Ma, S.L. Paris saponin I induces apoptosis via increasing the Bax/Bcl-2 ratio and caspase-3 expression in gefitinib-resistant non-small cell lung cancer in vitro and in vivo. Mol. Med. Reports 2014, 9, 2265–2272. [Google Scholar] [CrossRef] [PubMed]










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Liu, Y.; Sun, M.; Sai, N.; You, L.; Dong, X.; Yin, X.; Ni, J. Hepatocellular Toxicity of Paris Saponins I, II, VI and VII on Two Kinds of Hepatocytes-HL-7702 and HepaRG Cells, and the Underlying Mechanisms. Cells 2019, 8, 690. https://doi.org/10.3390/cells8070690
Wang W, Liu Y, Sun M, Sai N, You L, Dong X, Yin X, Ni J. Hepatocellular Toxicity of Paris Saponins I, II, VI and VII on Two Kinds of Hepatocytes-HL-7702 and HepaRG Cells, and the Underlying Mechanisms. Cells. 2019; 8(7):690. https://doi.org/10.3390/cells8070690
Chicago/Turabian StyleWang, Wenping, Yi Liu, Mingyi Sun, Na Sai, Longtai You, Xiaoxv Dong, Xingbin Yin, and Jian Ni. 2019. "Hepatocellular Toxicity of Paris Saponins I, II, VI and VII on Two Kinds of Hepatocytes-HL-7702 and HepaRG Cells, and the Underlying Mechanisms" Cells 8, no. 7: 690. https://doi.org/10.3390/cells8070690
APA StyleWang, W., Liu, Y., Sun, M., Sai, N., You, L., Dong, X., Yin, X., & Ni, J. (2019). Hepatocellular Toxicity of Paris Saponins I, II, VI and VII on Two Kinds of Hepatocytes-HL-7702 and HepaRG Cells, and the Underlying Mechanisms. Cells, 8(7), 690. https://doi.org/10.3390/cells8070690