Defocused Image Changes Signaling of Ganglion Cells in the Mouse Retina


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Ethics Approval
2.2. Retina Preparation
2.3. Full Field Light Stimulation
2.4. Single Cell Patterned Light Stimulation
2.5. Electrical Recording
2.6. Data Analysis
2.7. Immunocytochemistry
3. Results
3.1. Projecting Defocused Images on Photoreceptors under Microscopy
3.2. The Light Intensities of the Defocused Image Activate the Cone Pathway
3.3. RGCs Showed Decreasing Spikes to Smaller Light Spot Stimuli
3.4. Different Spatial Frequencies Image Could Change the Signaling of α-RGCs
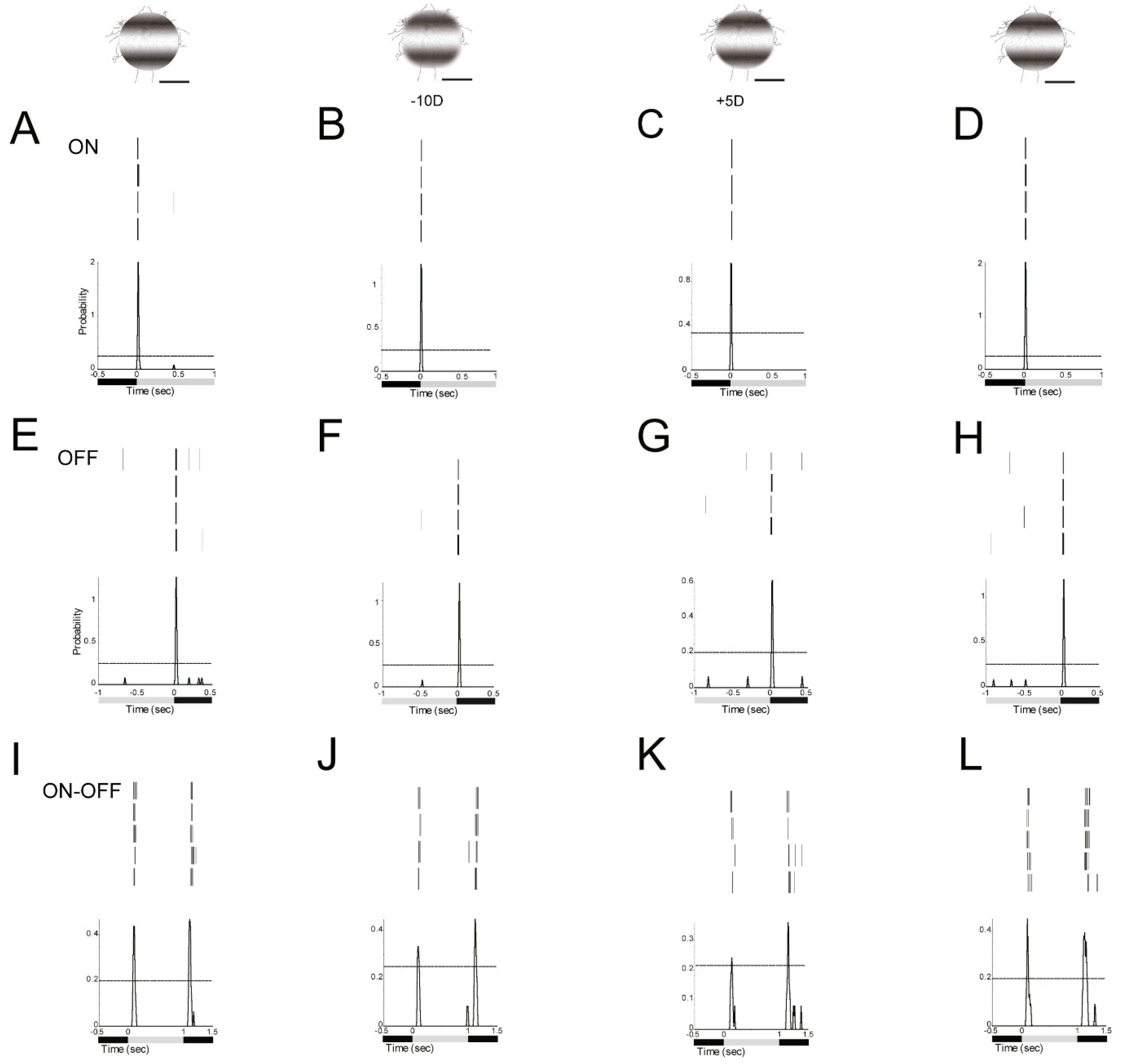
3.5. Defocused Images Altered the Signaling Responses in RGCs
3.6. Some RGCs Responded Differently to Plus and Minus Defocus
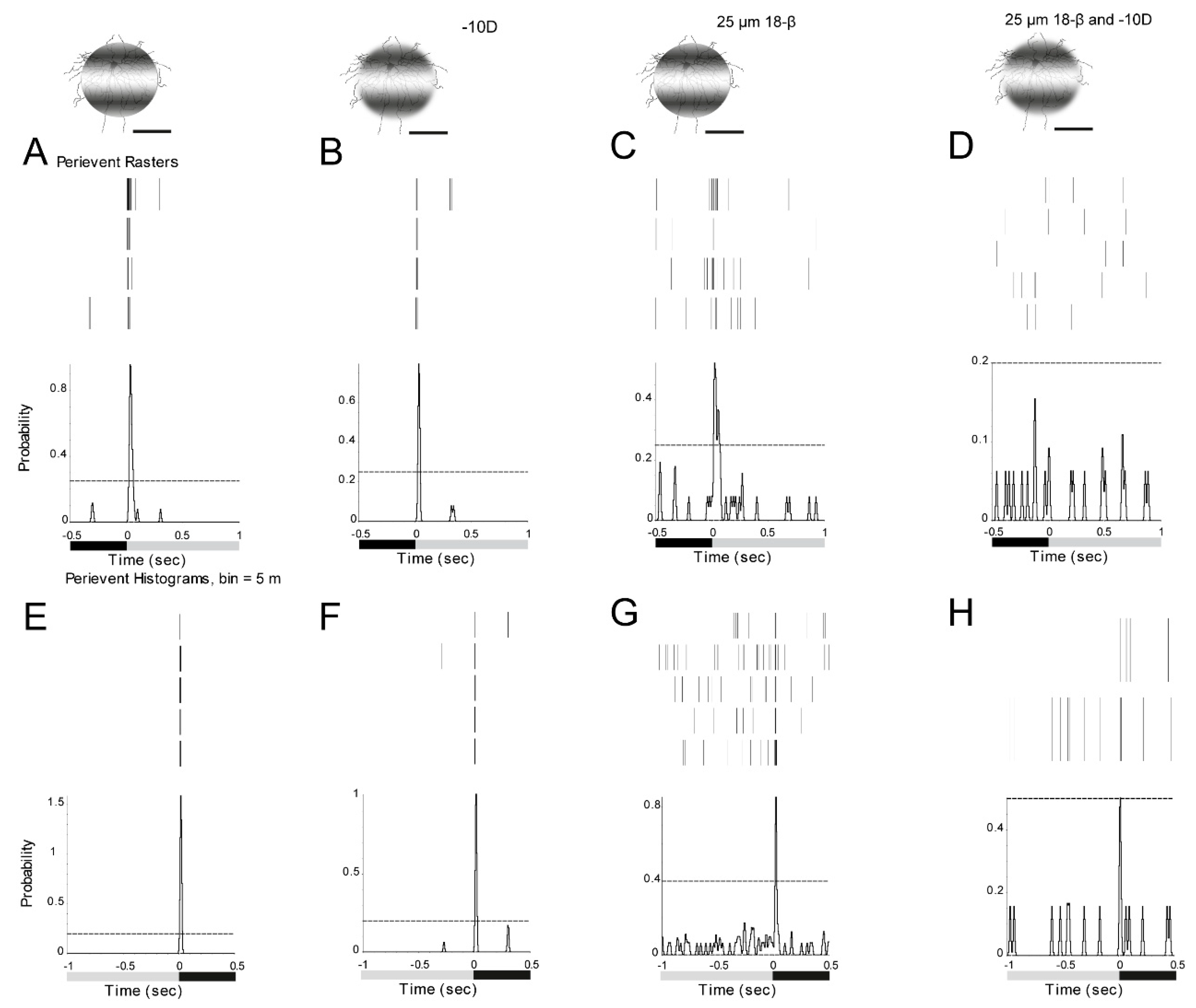
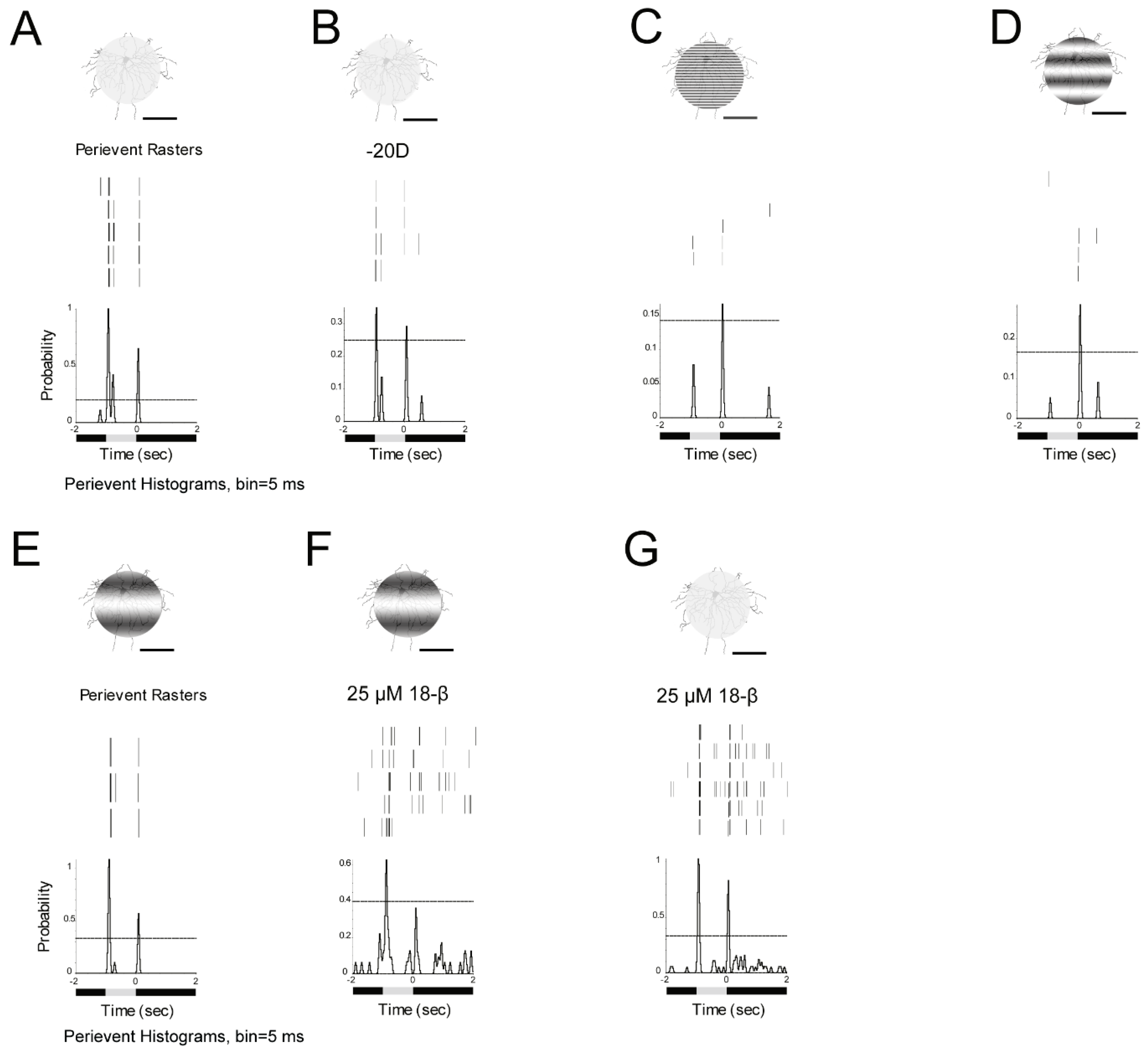
3.7. ON and OFF Alpha RGCs’ Response to Defocused Image Was Affected by Gap Junctions. Amacrine Cells May Contribute to These Effects Through Gap Junctions
3.8. ON Alpha RGCs Showed Decreased Responses to Defocused Images After the Application of Gap Junction Blocker
3.9. ON–OFF RGCs Showed Different Responses to Defocused Images and Spatial Frequencies. Amacrine Cells Might Contribute to the Effects Through Gap Junctions
4. Discussion
4.1. Mouse Eye Growth Models Can Provide a Powerful Means to Study Refractive Development
4.2. Defocused Image Changes the Signaling of Ganglion Cells in the Mouse Retina
4.3. Different Spatial Frequencies Can Also Change the Signaling of Ganglion Cells
4.4. Amacrine Cells Play an Important Function in Increasing Signaling and Decreasing Background Noise
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vitale, S.; Sperduto, R.D.; Ferris, F.L. Increased prevalence of myopia in the United States between 1971-1972 and 1999-2004. Arch. Ophthalmol. 2009, 127, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Goh, W.S.H.; Lam, C.S.Y. Changes in refractive trends and optical components of Hong Kong Chinese aged 19–39 years. Ophthalmic Physiol. Opt. 1994, 14, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Morgan, I.G.; Ohno-Matsui, K.; Saw, S.M. Myopia. Lancet 2012, 379, 1739–1748. [Google Scholar] [CrossRef]
- Morgan, I.G.; French, A.N.; Ashby, R.S.; Guo, X.; Ding, X.; He, M.; Rose, K.A. The epidemics of myopia: Aetiology and prevention. Prog. Retin. Eye Res. 2018, 62, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Ohno-Matsui, K. Pathologic Myopia. Asia Pac. J. Ophthalmol. 2016, 5, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Schaeffel, F. Test systems for measuring ocular parameters and visual function in mice. Front. Biosci. 2008, 13, 4904. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Remtulla, S.; Hallett, P. A schematic eye for the mouse, and comparisons with the rat. Vis. Res. 1985, 25, 21–31. [Google Scholar] [CrossRef]
- I Flitcroft, D. Emmetropisation and the aetiology of refractive errors. Eye 2014, 28, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Smith III, E.L.; Hung, L.F. The role of optical defocus in regulating refractive development in infant monkeys. Vision Res. 1999, 39, 1415–1435. [Google Scholar] [CrossRef]
- Smith III, E.L.; Hung, L.F.; Huang, J.; Arumugam, B. Effects of local myopic defocus on refractive development in monkeys. Optom. Vis. Sci. 2013, 90, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Read, S.A.; Collins, M.J.; Sander, B.P. Human Optical Axial Length and Defocus. Investig. Opthalmology Vis. Sci. 2010, 51, 6262–6269. [Google Scholar] [CrossRef] [PubMed]
- Chiang, S.T.-H.; Chen, T.-L.; Phillips, J.R. Effect of Optical Defocus on Choroidal Thickness in Healthy Adults With Presbyopia. Investig. Opthalmology Vis. Sci. 2018, 59, 5188–5193. [Google Scholar] [CrossRef]
- Hoseini-Yazdi, H.; Vincent, S.J.; Collins, M.J.; A Read, S. Regional alterations in human choroidal thickness in response to short-term monocular hemifield myopic defocus. Ophthalmic Physiol. Opt. 2019, 39, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Ho, W.-C.; Wong, O.-Y.; Chan, Y.-C.; Wong, S.-W.; Kee, C.-S.; Chan, H.H.-L. Sign-dependent changes in retinal electrical activity with positive and negative defocus in the human eye. Vis. Res. 2012, 52, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.R. Monovision slows juvenile myopia progression unilaterally. Br. J. Ophthalmol. 2005, 89, 1196–1200. [Google Scholar] [CrossRef] [PubMed]
- Berntsen, D.A.; Barr, C.D.; Mutti, D.O.; Zadnik, K. Peripheral Defocus and Myopia Progression in Myopic Children Randomly Assigned to Wear Single Vision and Progressive Addition Lenses. Investig. Opthalmology Vis. Sci. 2013, 54, 5761–5770. [Google Scholar] [CrossRef] [PubMed]
- Schaeffel, F.; Wildsoet, C. Can the retina alone detect the sign of defocus? Ophthalmic Physiol. Opt. 2013, 33, 362–367. [Google Scholar] [CrossRef]
- Chin, M.P.; Chu, P.H.W.; Cheong, A.M.Y.; Chan, H.H.L. Human Electroretinal Responses to Grating Patterns and Defocus Changes by Global Flash Multifocal Electroretinogram. PLoS ONE 2015, 10, e0123480. [Google Scholar] [CrossRef] [PubMed]
- Völgyi, B.; Chheda, S.; Bloomfield, S.A. Tracer Coupling Patterns of the Ganglion Cell Subtypes in the Mouse Retina. J. Comp. Neurol. 2009, 512, 664–687. [Google Scholar] [CrossRef]
- Baden, T.; Berens, P.; Franke, K.; Rosón, M.R.; Bethge, M.; Euler, T. The functional diversity of retinal ganglion cells in the mouse. Nature 2016, 529, 345–350. [Google Scholar] [CrossRef]
- Vlasits, A.L.; Euler, T.; Franke, K. Function first: Classifying cell types and circuits of the retina. Curr. Opin. Neurobiol. 2019, 56, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Gjorgjieva, J.; Sompolinsky, H.; Meister, M. Benefits of Pathway Splitting in Sensory Coding. J. Neurosci. 2014, 34, 12127–12144. [Google Scholar] [CrossRef] [PubMed]
- Asari, H.; Meister, M. Divergence of visual channels in the inner retina. Nat. Neurosci. 2012, 15, 1581–1589. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, H.; Liu, W.L.; Massey, S.C.; Mills, S.L. ON inputs to the OFF layer: Bipolar cells that break the stratification rules of the retina. J. Neurosci. 2009, 29, 8875–8883. [Google Scholar] [CrossRef] [PubMed]
- Rodieck, R. The First Steps in Seeing; Sinauer Associates: Sunderland, MA, USA, 1998. [Google Scholar]
- Krieger, B.; Qiao, M.; Rousso, D.L.; Sanes, J.R.; Meister, M. Four alpha ganglion cell types in mouse retina: Function, structure, and molecular signatures. PLoS ONE 2017, 12, e0180091. [Google Scholar] [CrossRef]
- Wässle, H.; Peichl, L.; Boycott, B.B. Morphology and topography of on- and off-alpha cells in the cat retina. Proc. R. Soc. London. Ser. B: Boil. Sci. 1981, 212, 157–175. [Google Scholar]
- Chakraborty, R.; Na Park, H.; Hanif, A.M.; Sidhu, C.S.; Iuvone, P.M.; Pardue, M.T. ON pathway mutations increase susceptibility to form-deprivation myopia. Exp. Eye Res. 2015, 137, 79–83. [Google Scholar] [CrossRef]
- Crewther, S.G.; Crewther, D.P. Inhibition of retinal ON/OFF systems differentially affects refractive compensation to defocus. NeuroReport 2003, 14, 1233–1237. [Google Scholar] [CrossRef]
- Schaeffel, F.; Feldkaemper, M. Animal models in myopia research. Clin. Exp. Optom. 2015, 98, 507–517. [Google Scholar] [CrossRef]
- Duan, X.; Qiao, M.; Bei, F.; Kim, I.J.; He, Z.; Sanes, J.R. Subtype-specific regeneration of retinal ganglion cells following axotomy: Effects of osteopontin and mTOR signaling. Neuron 2015, 85, 1244–1256. [Google Scholar] [CrossRef]
- Bloomfield, S.A.; Miller, R.F. A physiological and morphological study of the horizontal cell types of the rabbit retina. J. Comp. Neurol. 1982, 208, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Jeon, C.-J.; Strettoi, E.; Masland, R.H. The Major Cell Populations of the Mouse Retina. J. Neurosci. 1998, 18, 8936–8946. [Google Scholar] [CrossRef] [PubMed]
- Penn, J.S.; Williams, T.P. A new microspectrophotometric method for measuring absorbance of rat photoreceptors. Vis. Res. 1984, 24, 1673–1676. [Google Scholar] [CrossRef]
- Peirce, J.W. PsychoPy—Psychophysics software in Python. J. Neurosci. Methods 2007, 162, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Borghuis, B.G.; Marvin, J.S.; Looger, L.L.; Demb, J.B. Two-Photon Imaging of Nonlinear Glutamate Release Dynamics at Bipolar Cell Synapses in the Mouse Retina. J. Neurosci. 2013, 33, 10972–10985. [Google Scholar] [CrossRef] [PubMed]
- Della Santina, L.; Inman, D.M.; Lupien, C.B.; Horner, P.J.; Wong, R.O.L. Differential Progression of Structural and Functional Alterations in Distinct Retinal Ganglion Cell Types in a Mouse Model of Glaucoma. J. Neurosci. 2013, 33, 17444–17457. [Google Scholar] [CrossRef]
- Naka, K.I.; Rushton, W.A.H. S-potentials from luminosity units in the retina of fish (Cyprinidae). J. Physiol. 1966, 185, 587–599. [Google Scholar] [CrossRef]
- Baylor, D.A.; Hodgkin, A.L.; Lamb, T.D. Reconstruction of the electrical responses of turtle cones to flashes and steps of light. J. Physiol. 1974, 242, 759–791. [Google Scholar] [CrossRef]
- Thibos, L.N.; Werblin, F.S. The properties of surround antagonism elicited by spinning windmill patterns in the mudpuppy retina. J. Physiol. 1978, 278, 101–116. [Google Scholar] [CrossRef]
- Osterhout, J.A.; Josten, N.; Yamada, J.; Pan, F.; Wu, S.-W.; Nguyen, P.L.; Panagiotakos, G.; Inoue, Y.U.; Egusa, S.F.; Volgyi, B.; et al. Cadherin-6 Mediates Axon-Target Matching in a Non-Image-Forming Visual Circuit. Neuron 2011, 71, 632–639. [Google Scholar] [CrossRef]
- Schmucker, C.; Schaeffel, F. A paraxial schematic eye model for the growing C57BL/6 mouse. Vis. Res. 2004, 44, 1857–1867. [Google Scholar] [CrossRef] [PubMed]
- Deans, M.R.; Volgyi, B.; A Goodenough, D.; A Bloomfield, S.; Paul, D.L. Connexin36 Is Essential for Transmission of Rod-Mediated Visual Signals in the Mammalian Retina. Neuron 2002, 36, 703–712. [Google Scholar] [CrossRef]
- Pan, F.; Toychiev, A.; Zhang, Y.; Atlasz, T.; Ramakrishnan, H.; Roy, K.; Völgyi, B.; Akopian, A.; Bloomfield, S.A. Inhibitory masking controls the threshold sensitivity of retinal ganglion cells. J. Physiol. 2016, 594, 6679–6699. [Google Scholar] [CrossRef] [PubMed]
- Volgyi, B.; Pan, F.; Paul, D.L.; Wang, J.T.; Huberman, A.D.; Bloomfield, S.A. Gap Junctions Are Essential for Generating the Correlated Spike Activity of Neighboring Retinal Ganglion Cells. PLOS ONE 2013, 8, 69426. [Google Scholar] [CrossRef] [PubMed]
- Justice, M.J.; Siracusa, L.D.; Stewart, A.F. Technical approaches for mouse models of human disease. Dis. Model. Mech. 2011, 4, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Seabrook, T.A.; Burbridge, T.J.; Crair, M.C.; Huberman, A.D. Architecture, Function, and Assembly of the Mouse Visual System. Annu. Rev. Neurosci. 2017, 40, 499–538. [Google Scholar] [CrossRef] [PubMed]
- Barathi, V.; Boopathi, V.; Yap, E.P.; Beuerman, R.W. Two models of experimental myopia in the mouse. Vis. Res. 2008, 48, 904–916. [Google Scholar] [CrossRef] [PubMed]
- Pardue, M.T.; Stone, R.A.; Iuvone, P.M. Investigating mechanisms of myopia in mice. Exp. Eye Res. 2013, 114, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Tan, C.C.; Faulkner, A.; Jabbar, S.B.; Schmid, G.; Abey, J.; Iuvone, P.M.; Pardue, M.T. Retinal degeneration increases susceptibility to myopia in mice. Mol. Vis. 2013, 19, 2068–2079. [Google Scholar]
- Stone, R.A.; Pardue, M.T.; Iuvone, P.M.; Khurana, T.S. Pharmacology of Myopia and Potential Role for Intrinsic Retinal Circadian Rhythms. Exp. Eye Res. 2013, 114, 35–47. [Google Scholar] [CrossRef]
- Norton, T.T. Experimental myopia in tree shrews. Ciba Found Symp. 1990, 155, 178–194. [Google Scholar] [PubMed]
- Norton, T.T.; Essinger, J.A.; McBrien, N.A. Lid-suture myopia in tree shrews with retinal ganglion cell blockade. Vis. Neurosci. 1994, 11, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Schmid, K.L.; Wildsoet, C.F. Effects on the compensatory responses to positive and negative lenses of intermittent lens wear and ciliary nerve section in chicks. Vis. Res. 1996, 36, 1023–1036. [Google Scholar] [CrossRef]
- Angle, J.; Wissmann, D.A. The epidemiology of myopia. Am. J. Epidemiol. 1980, 111, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Saw, S.M.; Katz, J.; Schein, O.D.; Chew, S.J.; Chan, T.K. Epidemiology of myopia. Epidemiol. Rev. 1996, 18, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Zadnik, K. The Glenn A. Fry Award Lecture (1995). Myopia development in childhood. Optom. Vis. Sci. 1997, 74, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Saw, S.M.; Chua, W.H.; Wu, H.M.; Yap, E.; Chia, K.S.; A Stone, R. Myopia: Gene-environment interaction. Ann. Acad. Med. Singap. 2000, 29, 290–297. [Google Scholar] [PubMed]
- Goldschmidt, E.; Jacobsen, N. Genetic and environmental effects on myopia development and progression. Eye (Lond) 2014, 28, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Cho, P.; Tan, Q. Myopia and orthokeratology for myopia control. Clin. Exp. Optom. 2018, 102, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Joachimsen, L.; Böhringer, D.; Gross, N.J.; Reich, M.; Stifter, J.; Reinhard, T.; Lagrèze, W.A. A Pilot Study on the Efficacy and Safety of 0.01% Atropine in German Schoolchildren with Progressive Myopia. Ophthalmol. Ther. 2019, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.S.Y.; Tang, W.C.; Tse, D.Y.-Y.; Lee, R.P.K.; Chun, R.K.M.; Hasegawa, K.; Qi, H.; Hatanaka, T.; To, C.H. Defocus Incorporated Multiple Segments (DIMS) spectacle lenses slow myopia progression: a 2-year randomised clinical trial. Br. J. Ophthalmol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Norton, T.T. Animal Models of Myopia: Learning How Vision Controls the Size of the Eye. ILAR J. 1999, 40, 59–77. [Google Scholar] [CrossRef] [PubMed]
- Schaeffel, F.; Glasser, A.; Howland, H.C. Accommodation, refractive error and eye growth in chickens. Vis. Res. 1988, 28, 639–657. [Google Scholar] [CrossRef]
- Jiang, X.; Kurihara, T.; Kunimi, H.; Miyauchi, M.; Ikeda, S.-I.; Mori, K.; Tsubota, K.; Torii, H.; Tsubota, K. A highly efficient murine model of experimental myopia. Sci. Rep. 2018, 8, 2026. [Google Scholar] [CrossRef] [PubMed]
- Norton, T.T.; Siegwart, J.T. Animal models of emmetropization: matching axial length to the focal plane. J. Am. Optom. Assoc. 1995, 66, 405–414. [Google Scholar] [PubMed]
- Graham, B.; Judge, S. The effects of spectacle wear in infancy on eye growth and refractive error in the marmoset (Callithrix jacchus). Vis. Res. 1999, 39, 189–206. [Google Scholar] [CrossRef]
- Hung, L.-F.; Crawford, M.; Smith, E.L. Spectacle lenses alter eye growth and the refractive status of young monkeys. Nat. Med. 1995, 1, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.L. Spectacle lenses and emmetropization: The role of optical defocus in regulating ocular development. Optom. Vis. Sci. 1998, 75, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chun, R.K.M.; Liu, M.; Lee, R.P.K.; Sun, Y.; Zhang, T.; Lam, C.; Liu, Q.; To, C.H. Optical Defocus Rapidly Changes Choroidal Thickness in Schoolchildren. PLOS ONE 2016, 11, e0161535. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; Read, S.A.; Collins, M.J. Hyperopic Defocus and Diurnal Changes in Human Choroid and Axial Length. Optom. Vis. Sci. 2013, 90, 1187–1198. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; Read, S.A.; Collins, M.J. Monocular myopic defocus and daily changes in axial length and choroidal thickness of human eyes. Exp. Eye Res. 2012, 103, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Heidary, G.; Vanderveen, D.; Smith, L.E. Retinopathy of Prematurity: Current Concepts in Molecular Pathogenesis. Semin. Ophthalmol. 2009, 24, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Wallman, J.; Gottlieb, M.; Rajaram, V.; Fugate-Wentzek, L. Local retinal regions control local eye growth and myopia. Sci. 1987, 237, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.A.; Lin, T.; Laties, A.M.; Iuvone, P.M. Retinal dopamine and form-deprivation myopia. Proc. Natl. Acad. Sci. 1989, 86, 704–706. [Google Scholar] [CrossRef] [PubMed]
- Iuvone, P.M.; Haque, R.; Fernandes, A.; Lambert, S.R. Neonatal aphakia is associated with altered levels of dopamine metabolites in the non-human primate retina. Exp. Eye Res. 2015, 140, 187–189. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.A.; Laties, A.M.; Raviola, E.; Wiesel, T.N. Increase in retinal vasoactive intestinal polypeptide after eyelid fusion in primates. Proc. Natl. Acad. Sci. 1988, 85, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Seko, Y.; Shimizu, M.; Tokoro, T. Retinoic Acid Increases in the Retina of the Chick with Form Deprivation Myopia. Ophthalmic Res. 1998, 30, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Mertz, J.R.; Wallman, J. Choroidal Retinoic Acid Synthesis: A Possible Mediator between Refractive Error and Compensatory Eye Growth. Exp. Eye Res. 2000, 70, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.L.; Fox, D.A.; Duncan, G.C. Refractive-error changes in kitten eyes produced by chronic on-channel blockade. Vision Res. 1991, 31, 833–844. [Google Scholar] [CrossRef]
- Chakraborty, R.; Ostrin, L.A.; Nickla, D.L.; Iuvone, P.M.; Pardue, M.T.; Stone, R.A. Circadian rhythms, refractive development, and myopia. Ophthalmic Physiol. Opt. 2018, 38, 217–245. [Google Scholar] [CrossRef] [PubMed]
- Crewther, D.; Crewther, S.; Xie, R. Changes in Eye Growth Produced by Drugs Which Affect Retinal ON or OFF Responses to Light. J. Ocul. Pharmacol. Ther. 1996, 12, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; Na Park, H.; Aung, M.H.; Tan, C.C.; Sidhu, C.S.; Iuvone, P.M.; Pardue, M.T. Comparison of refractive development and retinal dopamine in OFF pathway mutant and C57BL/6J wild-type mice. Mol. Vis. 2014, 20, 1318–1327. [Google Scholar] [PubMed]
- Aleman, A.C.; Wang, M.; Schaeffel, F. Reading and Myopia: Contrast Polarity Matters. Sci. Rep. 2018, 8, 10840. [Google Scholar] [CrossRef] [PubMed]
- Baden, T.; Schaeffel, F.; Berens, P. Visual Neuroscience: A Retinal Ganglion Cell to Report Image Focus? Curr. Biol. 2017, 27, R139–R141. [Google Scholar] [CrossRef] [PubMed]
- Mani, A.; Schwartz, G.W. Circuit mechanisms of a retinal ganglion cell with stimulus dependent response latency and activation beyond its dendrites. Curr. Boil. 2017, 27, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Diedrich, E.; Schaeffel, F. Spatial resolution, contrast sensitivity, and sensitivity to defocus of chicken retinal ganglion cells in vitro. Vis. Neurosci. 2009, 26, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Peichl, L.; Wassle, H. Size, scatter and coverage of ganglion cell receptive field centres in the cat retina. J. Physiol. 1979, 291, 117–141. [Google Scholar] [CrossRef]
- Campbell, F.W.; Robson, J.G. Application of fourier analysis to the visibility of gratings. J. Physiol. 1968, 197, 551–566. [Google Scholar] [CrossRef]









© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, F. Defocused Image Changes Signaling of Ganglion Cells in the Mouse Retina. Cells 2019, 8, 640. https://doi.org/10.3390/cells8070640
Pan F. Defocused Image Changes Signaling of Ganglion Cells in the Mouse Retina. Cells. 2019; 8(7):640. https://doi.org/10.3390/cells8070640
Chicago/Turabian StylePan, Feng. 2019. "Defocused Image Changes Signaling of Ganglion Cells in the Mouse Retina" Cells 8, no. 7: 640. https://doi.org/10.3390/cells8070640
APA StylePan, F. (2019). Defocused Image Changes Signaling of Ganglion Cells in the Mouse Retina. Cells, 8(7), 640. https://doi.org/10.3390/cells8070640