p190RhoGAPs, the ARHGAP35- and ARHGAP5-Encoded Proteins, in Health and Disease


Abstract
1. Introduction
2. p190RhoGAP Architecture
3. p190RhoGAP Cellular Functions
3.1. RhoA-Dependent Cell Mechanisms
3.1.1. Cell–Cell Junctions and Endothelial Permeability
3.1.2. Cell–Cell Junctions and Collective Cell Migration
3.1.3. Cell–Cell Junctions and Entosis
3.1.4. Cell Division
3.1.5. Dendritic Spines and Neuronal Morphogenesis
3.2. RhoA-Independent Functions
3.2.1. RhoC/Rac1 Dependent Cell Migration and Invasion
3.2.2. Gene Expression and Protein Translation
4. p190RhoGAP Function during Development
5. p190RhoGAPs in Human Diseases
5.1. p190RhoGAPs in Neuronal Diseases
5.2. p190RhoGAPs in Cancer
5.2.1. ARHGAP35 as a Significant Mutated Gene in Cancer
5.2.2. ARHGAP35 and ARHGAP5 as Tumor-Suppressor Genes
5.2.3. ARHGAP35 and ARHGAP5 as Potential Oncogenes
5.3. p190A and Other Diseases
6. Conclusions and Future Directions
Funding
Conflicts of Interest
Abbreviations
| Abl2 | v-Abl Abelson murine leukemia viral oncogene homolog 2 |
| ACP1 | acid phosphatase 1 |
| Arg | Abelson related gene |
| Blk | B lymphocyte kinase |
| Brk | breast tumor kinase |
| CRAD | Catenin-RhoGAP-Association Domain |
| EGFR-TKI | epidermal growth factor receptor tyrosine kinase inhibitor |
| FAK | focal adhesion kinase |
| FAS | fetal alcohol spectrum |
| FTD | Frontotemporal dementia |
| GAP | GTPase activating protein |
| GBD | GTP-binding domain |
| GDP | guanosine diphosphate |
| GEF | guanine nucleotide exchange factor |
| GPI | glycosylphosphatidylinositol |
| GRF-1 | glucocorticoid receptor DNA-binding factor 1 |
| GTP | guanosine triphosphate |
| GTPase | guanosine triphosphate hydrolase |
| HCC | hepatocellular carcinoma |
| IPF | idiopathic pulmonary fibrosis |
| MCT1 | multiple copies in T-cell malignancy-1 |
| PBR | polybasic region |
| PDGF | platelet derived growth factor |
| PLS | protrusion localization sequence |
| PrPc | cellular prion protein |
| Ras | Rat sarcoma |
| ROCK | Rho associated coiled-coil-containing protein kinase |
| Rho | Ras homolog |
| TEB | terminal end buds |
| YAP | yes-associated protein |
References
- Toksoz, D.; Merdek, K.D. The Rho small GTPase: Functions in health and disease. Histol. Histopathol. 2002, 17, 915–927. [Google Scholar] [CrossRef]
- Haga, R.B.; Ridley, A.J. Rho GTPases: Regulation and roles in cancer cell biology. Small GTPases 2016, 7, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Hall, A. Rho family GTPases. Biochem. Soc. Trans. 2012, 40, 1378–1382. [Google Scholar] [CrossRef] [PubMed]
- Stiegler, A.L.; Boggon, T.J. The N-Terminal GTPase Domain of p190RhoGAP Proteins Is a PseudoGTPase. Structure 2018, 26, 1451–1461.e1454. [Google Scholar] [CrossRef] [PubMed]
- Burbelo, P.D.; Finegold, A.A.; Kozak, C.A.; Yamada, Y.; Takami, H. Cloning, genomic organization and chromosomal assignment of the mouse p190-B gene. Biochim. Biophys. Acta 1998, 1443, 203–210. [Google Scholar] [CrossRef]
- Burbelo, P.D.; Miyamoto, S.; Utani, A.; Brill, S.; Yamada, K.M.; Hall, A.; Yamada, Y. p190-B, a new member of the Rho GAP family, and Rho are induced to cluster after integrin cross-linking. J. Boil. Chem. 1995, 270, 30919–30926. [Google Scholar]
- Ridley, A.J.; Self, A.J.; Kasmi, F.; Paterson, H.F.; Hall, A.; Marshall, C.J.; Ellis, C. rho family GTPase activating proteins p190, bcr and rhoGAP show distinct specificities in vitro and in vivo. EMBO J. 1993, 12, 5151–5160. [Google Scholar]
- Vincent, S.; Settleman, J. Inhibition of RhoGAP activity is sufficient for the induction of Rho-mediated actin reorganization. Eur. J. Cell Biol. 1999, 78, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, B.; Zheng, Y. Structural determinants required for the interaction between Rho GTPase and the GTPase-activating domain of p190. J. Boil. Chem. 1997, 272, 32830–32835. [Google Scholar]
- Levay, M.; Bartos, B.; Ligeti, E. p190RhoGAP has cellular RacGAP activity regulated by a polybasic region. Cell. Signal. 2013, 25, 1388–1394. [Google Scholar] [CrossRef]
- Ligeti, E.; Dagher, M.C.; Hernandez, S.E.; Koleske, A.J.; Settleman, J. Phospholipids can switch the GTPase substrate preference of a GTPase-activating protein. J. Boil. Chem. 2004, 279, 5055–5058. [Google Scholar] [CrossRef]
- Roof, R.W.; Dukes, B.D.; Chang, J.H.; Parsons, S.J. Phosphorylation of the p190 RhoGAP N-terminal domain by c-Src results in a loss of GTP binding activity. FEBS Lett. 2000, 472, 117–121. [Google Scholar] [CrossRef]
- Tatsis, N.; Lannigan, D.A.; Macara, I.G. The function of the p190 Rho GTPase-activating protein is controlled by its N-terminal GTP binding domain. J. Boil. Chem. 1998, 273, 34631–34638. [Google Scholar] [CrossRef]
- Foster, R.; Hu, K.Q.; Shaywitz, D.A.; Settleman, J. p190 RhoGAP, the major RasGAP-associated protein, binds GTP directly. Mol. Cell. Boil. 1994, 14, 7173–7181. [Google Scholar] [CrossRef]
- Jiang, W.; Sordella, R.; Chen, G.C.; Hakre, S.; Roy, A.L.; Settleman, J. An FF domain-dependent protein interaction mediates a signaling pathway for growth factor-induced gene expression. Mol. Cell 2005, 17, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Parasuraman, P.; Mulligan, P.; Walker, J.A.; Li, B.; Boukhali, M.; Haas, W.; Bernards, A. Interaction of p190A RhoGAP with eIF3A and Other Translation Preinitiation Factors Suggests a Role in Protein Biosynthesis. J. Boil. Chem. 2017, 292, 2679–2689. [Google Scholar] [CrossRef]
- Stiegler, A.L.; Boggon, T.J. p190RhoGAP proteins contain pseudoGTPase domains. Nat. Commun. 2017, 8, 506. [Google Scholar] [CrossRef]
- Stiegler, A.L.; Boggon, T.J. PseudoGTPase domains in p190RhoGAP proteins: A mini-review. Biochem. Soc. Trans. 2018, 46, 1713–1720. [Google Scholar] [CrossRef]
- Bidaud-Meynard, A.; Biname, F.; Lagree, V.; Moreau, V. Regulation of Rho GTPase activity at the leading edge of migrating cells by p190RhoGAP. Small GTPases 2017, 10, 99–110. [Google Scholar] [CrossRef][Green Version]
- Biname, F.; Bidaud-Meynard, A.; Magnan, L.; Piquet, L.; Montibus, B.; Chabadel, A.; Saltel, F.; Lagree, V.; Moreau, V. Cancer-associated mutations in the protrusion-targeting region of p190RhoGAP impact tumor cell migration. J. Cell Boil. 2016, 214, 859–873. [Google Scholar] [CrossRef]
- Wennerberg, K.; Forget, M.A.; Ellerbroek, S.M.; Arthur, W.T.; Burridge, K.; Settleman, J.; Der, C.J.; Hansen, S.H. Rnd proteins function as RhoA antagonists by activating p190 RhoGAP. Curr. Biol. 2003, 13, 1106–1115. [Google Scholar] [CrossRef]
- Bustos, R.I.; Forget, M.A.; Settleman, J.E.; Hansen, S.H. Coordination of Rho and Rac GTPase function via p190B RhoGAP. Curr. Biol. 2008, 18, 1606–1611. [Google Scholar] [CrossRef]
- McGlade, J.; Brunkhorst, B.; Anderson, D.; Mbamalu, G.; Settleman, J.; Dedhar, S.; Rozakis-Adcock, M.; Chen, L.B.; Pawson, T. The N-terminal region of GAP regulates cytoskeletal structure and cell adhesion. EMBO J. 1993, 12, 3073–3081. [Google Scholar] [CrossRef] [PubMed]
- Bryant, S.S.; Briggs, S.; Smithgall, T.E.; Martin, G.A.; McCormick, F.; Chang, J.H.; Parsons, S.J.; Jove, R. Two SH2 domains of p120 Ras GTPase-activating protein bind synergistically to tyrosine phosphorylated p190 Rho GTPase-activating protein. J. Boil. Chem. 1995, 270, 17947–17952. [Google Scholar] [CrossRef]
- Hu, K.Q.; Settleman, J. Tandem SH2 binding sites mediate the RasGAP-RhoGAP interaction: A conformational mechanism for SH3 domain regulation. EMBO J. 1997, 16, 473–483. [Google Scholar] [CrossRef]
- Roof, R.W.; Haskell, M.D.; Dukes, B.D.; Sherman, N.; Kinter, M.; Parsons, S.J. Phosphotyrosine (p-Tyr)-dependent and -independent mechanisms of p190 RhoGAP-p120 RasGAP interaction: Tyr 1105 of p190, a substrate for c-Src, is the sole p-Tyr mediator of complex formation. Mol. Cell. Boil. 1998, 18, 7052–7063. [Google Scholar] [CrossRef]
- Hernandez, S.E.; Settleman, J.; Koleske, A.J. Adhesion-dependent regulation of p190RhoGAP in the developing brain by the Abl-related gene tyrosine kinase. Curr. Biol. 2004, 14, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.H.; Chen, H.Y.; Lin, M.S.; Li, F.Y.; Chang, C.C.; Kuo, M.L.; Settleman, J.; Chen, R.H. Breast tumor kinase phosphorylates p190RhoGAP to regulate rho and ras and promote breast carcinoma growth, migration, and invasion. Cancer Res. 2008, 68, 7779–7787. [Google Scholar] [CrossRef]
- Csepanyi-Komi, R.; Pasztor, M.; Bartos, B.; Ligeti, E. The neglected terminators: Rho family GAPs in neutrophils. Eur. J. Clin. Investig. 2018, 48 (Suppl. 2). [Google Scholar] [CrossRef] [PubMed]
- McCormick, F. ras GTPase activating protein: Signal transmitter and signal terminator. Cell 1989, 56, 5–8. [Google Scholar] [CrossRef]
- O’Connor, K.; Chen, M. Dynamic functions of RhoA in tumor cell migration and invasion. Small GTPases 2013, 4, 141–147. [Google Scholar] [CrossRef]
- Tomar, A.; Lim, S.T.; Lim, Y.; Schlaepfer, D.D. A FAK-p120RasGAP-p190RhoGAP complex regulates polarity in migrating cells. J. Cell Sci. 2009, 122, 1852–1862. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, H.; Mueller, S.C.; Nomizu, M.; Yamada, Y.; Yeh, Y.; Chen, W.T. Activation of beta1 integrin signaling stimulates tyrosine phosphorylation of p190RhoGAP and membrane-protrusive activities at invadopodia. J. Boil. Chem. 1998, 273, 9–12. [Google Scholar] [CrossRef]
- Guegan, F.; Tatin, F.; Leste-Lasserre, T.; Drutel, G.; Genot, E.; Moreau, V. p190B RhoGAP regulates endothelial-cell-associated proteolysis through MT1-MMP and MMP2. J. Cell Sci. 2008, 121, 2054–2061. [Google Scholar] [CrossRef]
- Wildenberg, G.A.; Dohn, M.R.; Carnahan, R.H.; Davis, M.A.; Lobdell, N.A.; Settleman, J.; Reynolds, A.B. p120-catenin and p190RhoGAP regulate cell-cell adhesion by coordinating antagonism between Rac and Rho. Cell 2006, 127, 1027–1039. [Google Scholar] [CrossRef]
- Su, L.; Pertz, O.; Mikawa, M.; Hahn, K.; Parsons, S.J. p190RhoGAP negatively regulates Rho activity at the cleavage furrow of mitotic cells. Exp. Cell Res. 2009, 315, 1347–1359. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Macara, I.G. The PAR-6 polarity protein regulates dendritic spine morphogenesis through p190 RhoGAP and the Rho GTPase. Dev. Cell 2008, 14, 216–226. [Google Scholar] [CrossRef]
- Noren, N.K.; Arthur, W.T.; Burridge, K. Cadherin engagement inhibits RhoA via p190RhoGAP. J. Boil. Chem. 2003, 278, 13615–13618. [Google Scholar] [CrossRef]
- Zebda, N.; Tian, Y.; Tian, X.; Gawlak, G.; Higginbotham, K.; Reynolds, A.B.; Birukova, A.A.; Birukov, K.G. Interaction of p190RhoGAP with C-terminal domain of p120-catenin modulates endothelial cytoskeleton and permeability. J. Boil. Chem. 2013, 288, 18290–18299. [Google Scholar] [CrossRef]
- Hidalgo-Carcedo, C.; Hooper, S.; Chaudhry, S.I.; Williamson, P.; Harrington, K.; Leitinger, B.; Sahai, E. Collective cell migration requires suppression of actomyosin at cell-cell contacts mediated by DDR1 and the cell polarity regulators Par3 and Par6. Nat. Cell Boil. 2011, 13, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Li, C.F.; Chen, J.Y.; Ho, Y.H.; Hsu, W.H.; Wu, L.C.; Lan, H.Y.; Hsu, D.S.; Tai, S.K.; Chang, Y.C.; Yang, M.H. Snail-induced claudin-11 prompts collective migration for tumour progression. Nat. Cell Boil. 2019, 21, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Ponik, S.M.; Trier, S.M.; Wozniak, M.A.; Eliceiri, K.W.; Keely, P.J. RhoA is down-regulated at cell-cell contacts via p190RhoGAP-B in response to tensional homeostasis. Mol. Boil. Cell 2013, 24, 1688–1699. [Google Scholar] [CrossRef]
- Fais, S.; Overholtzer, M. Cell-in-cell phenomena in cancer. Nat. Rev. Cancer 2018, 18, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Overholtzer, M.; Mailleux, A.A.; Mouneimne, G.; Normand, G.; Schnitt, S.J.; King, R.W.; Cibas, E.S.; Brugge, J.S. A nonapoptotic cell death process, entosis, that occurs by cell-in-cell invasion. Cell 2007, 131, 966–979. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Cibas, E.S.; Huang, H.; Hodgson, L.; Overholtzer, M. Induction of entosis by epithelial cadherin expression. Cell Res. 2014, 24, 1288–1298. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Agati, J.M.; Parsons, S.J. p190RhoGAP is cell cycle regulated and affects cytokinesis. J. Cell Boil. 2003, 163, 571–582. [Google Scholar] [CrossRef]
- Manukyan, A.; Ludwig, K.; Sanchez-Manchinelly, S.; Parsons, S.J.; Stukenberg, P.T. A complex of p190RhoGAP-A and anillin modulates RhoA-GTP and the cytokinetic furrow in human cells. J. Cell Sci. 2015, 128, 50–60. [Google Scholar] [CrossRef]
- Mikawa, M.; Su, L.; Parsons, S.J. Opposing roles of p190RhoGAP and Ect2 RhoGEF in regulating cytokinesis. Cell Cycle 2008, 7, 2003–2012. [Google Scholar] [CrossRef] [PubMed]
- Manukyan, A.; Sargsyan, L.; Parsons, S.J.; Stukenberg, P.T. P190RhoGAP prevents mitotic spindle fragmentation and is required to activate Aurora A kinase at acentriolar poles. Chromosoma 2018, 127, 375–386. [Google Scholar] [CrossRef]
- Dupont, H.; Blancq, M. Formation of complexes involving RasGAP and p190 RhoGAP during morphogenetic events of the gastrulation in xenopus. Eur. J. Biochem. 1999, 265, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Brouns, M.R.; Matheson, S.F.; Hu, K.Q.; Delalle, I.; Caviness, V.S.; Silver, J.; Bronson, R.T.; Settleman, J. The adhesion signaling molecule p190 RhoGAP is required for morphogenetic processes in neural development. Development 2000, 127, 4891–4903. [Google Scholar] [PubMed]
- Brouns, M.R.; Matheson, S.F.; Settleman, J. p190 RhoGAP is the principal Src substrate in brain and regulates axon outgrowth, guidance and fasciculation. Nat. Cell Boil. 2001, 3, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Billuart, P.; Winter, C.G.; Maresh, A.; Zhao, X.; Luo, L. Regulating axon branch stability: The role of p190 RhoGAP in repressing a retraction signaling pathway. Cell 2001, 107, 195–207. [Google Scholar] [CrossRef]
- Kim, H.J.; Choi, H.S.; Park, J.H.; Kim, M.J.; Lee, H.G.; Petersen, R.B.; Kim, Y.S.; Park, J.B.; Choi, E.K. Regulation of RhoA activity by the cellular prion protein. Cell Death Dis. 2017, 8, e2668. [Google Scholar] [CrossRef]
- Wheeler, A.P.; Ridley, A.J. Why three Rho proteins? RhoA, RhoB, RhoC, and cell motility. Exp. Cell Res. 2004, 301, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Fingleton, B. Molecular targets in metastasis: Lessons from genomic approaches. Cancer Genom. Proteom. 2007, 4, 211–221. [Google Scholar]
- Vega, F.M.; Fruhwirth, G.; Ng, T.; Ridley, A.J. RhoA and RhoC have distinct roles in migration and invasion by acting through different targets. J. Cell Boil. 2011, 193, 655–665. [Google Scholar] [CrossRef]
- Bravo-Cordero, J.J.; Oser, M.; Chen, X.; Eddy, R.; Hodgson, L.; Condeelis, J. A novel spatiotemporal RhoC activation pathway locally regulates cofilin activity at invadopodia. Curr. Biol. 2011, 21, 635–644. [Google Scholar] [CrossRef]
- Bravo-Cordero, J.J.; Sharma, V.P.; Roh-Johnson, M.; Chen, X.; Eddy, R.; Condeelis, J.; Hodgson, L. Spatial regulation of RhoC activity defines protrusion formation in migrating cells. J. Cell Sci. 2013, 126, 3356–3369. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Cordero, J.J.; Hodgson, L.; Condeelis, J.S. Spatial regulation of tumor cell protrusions by RhoC. Cell Adhes. Migr. 2014, 8, 263–267. [Google Scholar] [CrossRef]
- Ligeti, E.; Settleman, J. Regulation of RhoGAP specificity by phospholipids and prenylation. Methods Enzymol. 2006, 406, 104–117. [Google Scholar] [CrossRef]
- Levay, M.; Settleman, J.; Ligeti, E. Regulation of the substrate preference of p190RhoGAP by protein kinase C-mediated phosphorylation of a phospholipid binding site. Biochemistry 2009, 48, 8615–8623. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.L. Biochemistry and biology of the inducible multifunctional transcription factor TFII-I. Gene 2001, 274, 1–13. [Google Scholar] [CrossRef]
- Mammoto, A.; Connor, K.M.; Mammoto, T.; Yung, C.W.; Huh, D.; Aderman, C.M.; Mostoslavsky, G.; Smith, L.E.; Ingber, D.E. A mechanosensitive transcriptional mechanism that controls angiogenesis. Nature 2009, 457, 1103–1108. [Google Scholar] [CrossRef]
- Sordella, R.; Classon, M.; Hu, K.Q.; Matheson, S.F.; Brouns, M.R.; Fine, B.; Zhang, L.; Takami, H.; Yamada, Y.; Settleman, J. Modulation of CREB activity by the Rho GTPase regulates cell and organism size during mouse embryonic development. Dev. Cell 2002, 2, 553–565. [Google Scholar] [CrossRef]
- Matheson, S.F.; Hu, K.Q.; Brouns, M.R.; Sordella, R.; VanderHeide, J.D.; Settleman, J. Distinct but overlapping functions for the closely related p190 RhoGAPs in neural development. Dev. Neurosci. 2006, 28, 538–550. [Google Scholar] [CrossRef]
- Sordella, R.; Jiang, W.; Chen, G.C.; Curto, M.; Settleman, J. Modulation of Rho GTPase signaling regulates a switch between adipogenesis and myogenesis. Cell 2003, 113, 147–158. [Google Scholar] [CrossRef]
- Chakravarty, G.; Hadsell, D.; Buitrago, W.; Settleman, J.; Rosen, J.M. p190-B RhoGAP regulates mammary ductal morphogenesis. Mol. Endocrinol. 2003, 17, 1054–1065. [Google Scholar] [CrossRef] [PubMed]
- Vargo-Gogola, T.; Heckman, B.M.; Gunther, E.J.; Chodosh, L.A.; Rosen, J.M. P190-B Rho GTPase-activating protein overexpression disrupts ductal morphogenesis and induces hyperplastic lesions in the developing mammary gland. Mol. Endocrinol. 2006, 20, 1391–1405. [Google Scholar] [CrossRef]
- Heckman-Stoddard, B.M.; Vargo-Gogola, T.; Herrick, M.P.; Visbal, A.P.; Lewis, M.T.; Settleman, J.; Rosen, J.M. P190A RhoGAP is required for mammary gland development. Dev. Biol. 2011, 360, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Ferrari, R.; Heutink, P.; Hardy, J.; Pijnenburg, Y.; Posthuma, D.; International, F.T.D.G.C. Gene-based association studies report genetic links for clinical subtypes of frontotemporal dementia. Brain 2017, 140, 1437–1446. [Google Scholar] [CrossRef]
- Gourley, S.L.; Swanson, A.M.; Koleske, A.J. Corticosteroid-induced neural remodeling predicts behavioral vulnerability and resilience. J. Neurosci. 2013, 33, 3107–3112. [Google Scholar] [CrossRef]
- Gourley, S.L.; Olevska, A.; Warren, M.S.; Taylor, J.R.; Koleske, A.J. Arg kinase regulates prefrontal dendritic spine refinement and cocaine-induced plasticity. J. Neurosci. 2012, 32, 2314–2323. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yoshikawa, A.; Meltzer, H.Y. Replication of rs300774, a genetic biomarker near ACP1, associated with suicide attempts in patients with schizophrenia: Relation to brain cholesterol biosynthesis. J. Psychiatr. Res. 2017, 94, 54–61. [Google Scholar] [CrossRef]
- Selva, J.; Egea, G. Ethanol increases p190RhoGAP activity, leading to actin cytoskeleton rearrangements. J. Neurochem. 2011, 119, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Fincham, V.J.; Chudleigh, A.; Frame, M.C. Regulation of p190 Rho-GAP by v-Src is linked to cytoskeletal disruption during transformation. J. Cell Sci. 1999, 112 Pt 6, 947–956. [Google Scholar]
- Settleman, J.; Albright, C.F.; Foster, L.C.; Weinberg, R.A. Association between GTPase activators for Rho and Ras families. Nature 1992, 359, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Settleman, J.; Narasimhan, V.; Foster, L.C.; Weinberg, R.A. Molecular cloning of cDNAs encoding the GAP-associated protein p190: Implications for a signaling pathway from ras to the nucleus. Cell 1992, 69, 539–549. [Google Scholar] [CrossRef]
- Bouton, A.H.; Kanner, S.B.; Vines, R.R.; Wang, H.C.; Gibbs, J.B.; Parsons, J.T. Transformation by pp60src or stimulation of cells with epidermal growth factor induces the stable association of tyrosine-phosphorylated cellular proteins with GTPase-activating protein. Mol. Cell. Boil. 1991, 11, 945–953. [Google Scholar] [CrossRef]
- Ellis, C.; Moran, M.; McCormick, F.; Pawson, T. Phosphorylation of GAP and GAP-associated proteins by transforming and mitogenic tyrosine kinases. Nature 1990, 343, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Z.; Nur, E.K.M.S.; Tikoo, A.; Montague, W.; Maruta, H. The GTPase and Rho GAP domains of p190, a tumor suppressor protein that binds the M(r) 120,000 Ras GAP, independently function as anti-Ras tumor suppressors. Cancer Res. 1997, 57, 2478–2484. [Google Scholar] [PubMed]
- Lawrence, M.S.; Stojanov, P.; Mermel, C.H.; Robinson, J.T.; Garraway, L.A.; Golub, T.R.; Meyerson, M.; Gabriel, S.B.; Lander, E.S.; Getz, G. Discovery and saturation analysis of cancer genes across 21 tumour types. Nature 2014, 505, 495–501. [Google Scholar] [CrossRef]
- Kandoth, C.; McLellan, M.D.; Vandin, F.; Ye, K.; Niu, B.; Lu, C.; Xie, M.; Zhang, Q.; McMichael, J.F.; Wyczalkowski, M.A.; et al. Mutational landscape and significance across 12 major cancer types. Nature 2013, 502, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.D.; Alexandrov, A.; Kim, J.; Wala, J.; Berger, A.H.; Pedamallu, C.S.; Shukla, S.A.; Guo, G.; Brooks, A.N.; Murray, B.A.; et al. Distinct patterns of somatic genome alterations in lung adenocarcinomas and squamous cell carcinomas. Nat. Genet. 2016, 48, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Gopal, R.K.; Kubler, K.; Calvo, S.E.; Polak, P.; Livitz, D.; Rosebrock, D.; Sadow, P.M.; Campbell, B.; Donovan, S.E.; Amin, S.; et al. Widespread Chromosomal Losses and Mitochondrial DNA Alterations as Genetic Drivers in Hurthle Cell Carcinoma. Cancer Cell 2018, 34, 242–255. [Google Scholar] [CrossRef]
- Giannikou, K.; Malinowska, I.A.; Pugh, T.J.; Yan, R.; Tseng, Y.Y.; Oh, C.; Kim, J.; Tyburczy, M.E.; Chekaluk, Y.; Liu, Y.; et al. Whole Exome Sequencing Identifies TSC1/TSC2 Biallelic Loss as the Primary and Sufficient Driver Event for Renal Angiomyolipoma Development. PLoS Genet. 2016, 12, e1006242. [Google Scholar] [CrossRef]
- Horpaopan, S.; Spier, I.; Zink, A.M.; Altmuller, J.; Holzapfel, S.; Laner, A.; Vogt, S.; Uhlhaas, S.; Heilmann, S.; Stienen, D.; et al. Genome-wide CNV analysis in 221 unrelated patients and targeted high-throughput sequencing reveal novel causative candidate genes for colorectal adenomatous polyposis. Int. J. Cancer 2015, 136, E578–E589. [Google Scholar] [CrossRef]
- Kurokawa, Y.; Matoba, R.; Takemasa, I.; Nakamori, S.; Tsujie, M.; Nagano, H.; Dono, K.; Umeshita, K.; Sakon, M.; Ueno, N.; et al. Molecular features of non-B, non-C hepatocellular carcinoma: A PCR-array gene expression profiling study. J. Hepatol. 2003, 39, 1004–1012. [Google Scholar] [CrossRef]
- Earp, M.; Tyrer, J.P.; Winham, S.J.; Lin, H.Y.; Chornokur, G.; Dennis, J.; Aben, K.K.H.; Anton-Culver, H.; Antonenkova, N.; Bandera, E.V.; et al. Variants in genes encoding small GTPases and association with epithelial ovarian cancer susceptibility. PLoS ONE 2018, 13, e0197561. [Google Scholar] [CrossRef]
- Tikoo, A.; Czekay, S.; Viars, C.; White, S.; Heath, J.K.; Arden, K.; Maruta, H. p190-A, a human tumor suppressor gene, maps to the chromosomal region 19q13.3 that is reportedly deleted in some gliomas. Gene 2000, 257, 23–31. [Google Scholar] [CrossRef]
- Zack, T.I.; Schumacher, S.E.; Carter, S.L.; Cherniack, A.D.; Saksena, G.; Tabak, B.; Lawrence, M.S.; Zhsng, C.Z.; Wala, J.; Mermel, C.H.; et al. Pan-cancer patterns of somatic copy number alteration. Nat. Genet. 2013, 45, 1134–1140. [Google Scholar] [CrossRef]
- Wolf, R.M.; Draghi, N.; Liang, X.; Dai, C.; Uhrbom, L.; Eklof, C.; Westermark, B.; Holland, E.C.; Resh, M.D. p190RhoGAP can act to inhibit PDGF-induced gliomas in mice: A putative tumor suppressor encoded on human chromosome 19q13.3. Genes Dev. 2003, 17, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Wolf, R.M.; Wilkes, J.J.; Chao, M.V.; Resh, M.D. Tyrosine phosphorylation of p190 RhoGAP by Fyn regulates oligodendrocyte differentiation. J. Neurobiol. 2001, 49, 62–78. [Google Scholar] [CrossRef] [PubMed]
- Kusama, T.; Mukai, M.; Endo, H.; Ishikawa, O.; Tatsuta, M.; Nakamura, H.; Inoue, M. Inactivation of Rho GTPases by p190 RhoGAP reduces human pancreatic cancer cell invasion and metastasis. Cancer Sci. 2006, 97, 848–853. [Google Scholar] [CrossRef]
- Shimizu, A.; Mammoto, A.; Italiano, J.E., Jr.; Pravda, E.; Dudley, A.C.; Ingber, D.E.; Klagsbrun, M. ABL2/ARG tyrosine kinase mediates SEMA3F-induced RhoA inactivation and cytoskeleton collapse in human glioma cells. J. Boil. Chem. 2008, 283, 27230–27238. [Google Scholar] [CrossRef]
- Qiang, X.F.; Zhang, Z.W.; Liu, Q.; Sun, N.; Pan, L.L.; Shen, J.; Li, T.; Yun, C.; Li, H.; Shi, L.H. miR-20a promotes prostate cancer invasion and migration through targeting ABL2. J. Cell. Biochem. 2014, 115, 1269–1276. [Google Scholar] [CrossRef]
- Bartolome, R.A.; Diaz-Martinez, M.; Colo, G.P.; Arellano-Sanchez, N.; Torres-Ayuso, P.; Kleinovink, J.W.; Merida, I.; Teixido, J. A Blk-p190RhoGAP signaling module downstream of activated Galpha13 functionally opposes CXCL12-stimulated RhoA activation and cell invasion. Cell. Signal. 2014, 26, 2551–2561. [Google Scholar] [CrossRef] [PubMed]
- Bartolome, R.A.; Wright, N.; Molina-Ortiz, I.; Sanchez-Luque, F.J.; Teixido, J. Activated Gα13 impairs cell invasiveness through p190RhoGAP-mediated inhibition of RhoA activity. Cancer Res. 2008, 68, 8221–8230. [Google Scholar] [CrossRef]
- Molina-Ortiz, I.; Bartolome, R.A.; Hernandez-Varas, P.; Colo, G.P.; Teixido, J. Overexpression of E-cadherin on melanoma cells inhibits chemokine-promoted invasion involving p190RhoGAP/p120ctn-dependent inactivation of RhoA. J. Boil. Chem. 2009, 284, 15147–15157. [Google Scholar] [CrossRef]
- Ludwig, K.; Parsons, S.J. The Tumor Suppressor, p190RhoGAP, Differentially Initiates Apoptosis and Confers Docetaxel Sensitivity to Breast Cancer Cells. Genes Cancer 2011, 2, 20–30. [Google Scholar] [CrossRef][Green Version]
- Frank, S.R.; Kollmann, C.P.; Luong, P.; Galli, G.G.; Zou, L.; Bernards, A.; Getz, G.; Calogero, R.A.; Frodin, M.; Hansen, S.H. p190 RhoGAP promotes contact inhibition in epithelial cells by repressing YAP activity. J. Cell Boil. 2018, 217, 3183–3201. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, H.; He, M.; Wu, Y. Glucocorticoid receptor DNA binding factor 1 expression and osteosarcoma prognosis. Tumour Biol. 2014, 35, 12449–12458. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xu, H.; He, M.; Wang, Z.; Wu, Y. Rho GTPase-activating protein 35 rs1052667 polymorphism and osteosarcoma risk and prognosis. Biomed Res. Int. 2014, 2014, 396947. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, Y.M.; Zhou, P.; Wang, X.S.; Wang, G.Y.; Zhao, X.H.; Cui, B.B.; Ren, Y.L.; Dong, X.S.; Chen, Z.Q. Abnormal expression of p190RhoGAP in colorectal cancer patients with poor survival. Am. J. Transl. Res. 2016, 8, 4405–4414. [Google Scholar]
- Organ, S.L.; Hai, J.; Radulovich, N.; Marshall, C.B.; Leung, L.; Sasazuki, T.; Shirasawa, S.; Zhu, C.Q.; Navab, R.; Ikura, M.; et al. p120RasGAP is a mediator of rho pathway activation and tumorigenicity in the DLD1 colorectal cancer cell line. PLoS ONE 2014, 9, e86103. [Google Scholar] [CrossRef] [PubMed]
- Notsuda, H.; Sakurada, A.; Endo, C.; Okada, Y.; Horii, A.; Shima, H.; Kondo, T. p190A RhoGAP is involved in EGFR pathways and promotes proliferation, invasion and migration in lung adenocarcinoma cells. Int. J. Oncol. 2013, 43, 1569–1577. [Google Scholar] [CrossRef] [PubMed]
- Chiu, J.H.; Wen, C.S.; Wang, J.Y.; Hsu, C.Y.; Tsai, Y.F.; Hung, S.C.; Tseng, L.M.; Shyr, Y.M. Role of estrogen receptors and Src signaling in mechanisms of bone metastasis by estrogen receptor positive breast cancers. J. Transl. Med. 2017, 15, 97. [Google Scholar] [CrossRef]
- Harvey, A.J.; Crompton, M.R. Use of RNA interference to validate Brk as a novel therapeutic target in breast cancer: Brk promotes breast carcinoma cell proliferation. Oncogene 2003, 22, 5006–5010. [Google Scholar] [CrossRef][Green Version]
- Chakravarty, G.; Roy, D.; Gonzales, M.; Gay, J.; Contreras, A.; Rosen, J.M. P190-B, a Rho-GTPase-activating protein, is differentially expressed in terminal end buds and breast cancer. Cell Growth Differ. 2000, 11, 343–354. [Google Scholar] [PubMed]
- Wu, M.H.; Chen, Y.A.; Chen, H.H.; Chang, K.W.; Chang, I.S.; Wang, L.H.; Hsu, H.L. MCT-1 expression and PTEN deficiency synergistically promote neoplastic multinucleation through the Src/p190B signaling activation. Oncogene 2014, 33, 5109–5120. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhu, X.; Wang, J.; Li, N.; Li, D.; Sakib, N.; Sha, Z.; Song, W. MiR-744 functions as a proto-oncogene in nasopharyngeal carcinoma progression and metastasis via transcriptional control of ARHGAP5. Oncotarget 2015, 6, 13164–13175. [Google Scholar] [CrossRef]
- Wang, J.; Tian, X.; Han, R.; Zhang, X.; Wang, X.; Shen, H.; Xue, L.; Liu, Y.; Yan, X.; Shen, J.; et al. Downregulation of miR-486-5p contributes to tumor progression and metastasis by targeting protumorigenic ARHGAP5 in lung cancer. Oncogene 2014, 33, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Heckman-Stoddard, B.M.; Vargo-Gogola, T.; McHenry, P.R.; Jiang, V.; Herrick, M.P.; Hilsenbeck, S.G.; Settleman, J.; Rosen, J.M. Haploinsufficiency for p190B RhoGAP inhibits MMTV-Neu tumor progression. Breast Cancer Res. 2009, 11, R61. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Wang, S.J.; Zhang, Y.; Hou, R.; Jiang, J.L.; Cui, H.Y. CD147 promotes cell motility via upregulation of p190-B RhoGAP in hepatocellular carcinoma. Cancer Cell Int. 2016, 16, 69. [Google Scholar] [CrossRef] [PubMed]
- Monaghan-Benson, E.; Wittchen, E.S.; Doerschuk, C.M.; Burridge, K. A Rnd3/p190RhoGAP pathway regulates RhoA activity in idiopathic pulmonary fibrosis fibroblasts. Mol. Boil. Cell 2018, 29, 2165–2175. [Google Scholar] [CrossRef] [PubMed]
- Stewart, K.; Gaitan, Y.; Shafer, M.E.; Aoudjit, L.; Hu, D.; Sharma, R.; Tremblay, M.; Ishii, H.; Marcotte, M.; Stanga, D.; et al. A Point Mutation in p190A RhoGAP Affects Ciliogenesis and Leads to Glomerulocystic Kidney Defects. PLoS Genet. 2016, 12, e1005785. [Google Scholar] [CrossRef]
- Porter, A.P.; Papaioannou, A.; Malliri, A. Deregulation of Rho GTPases in cancer. Small GTPases 2016, 7, 123–138. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
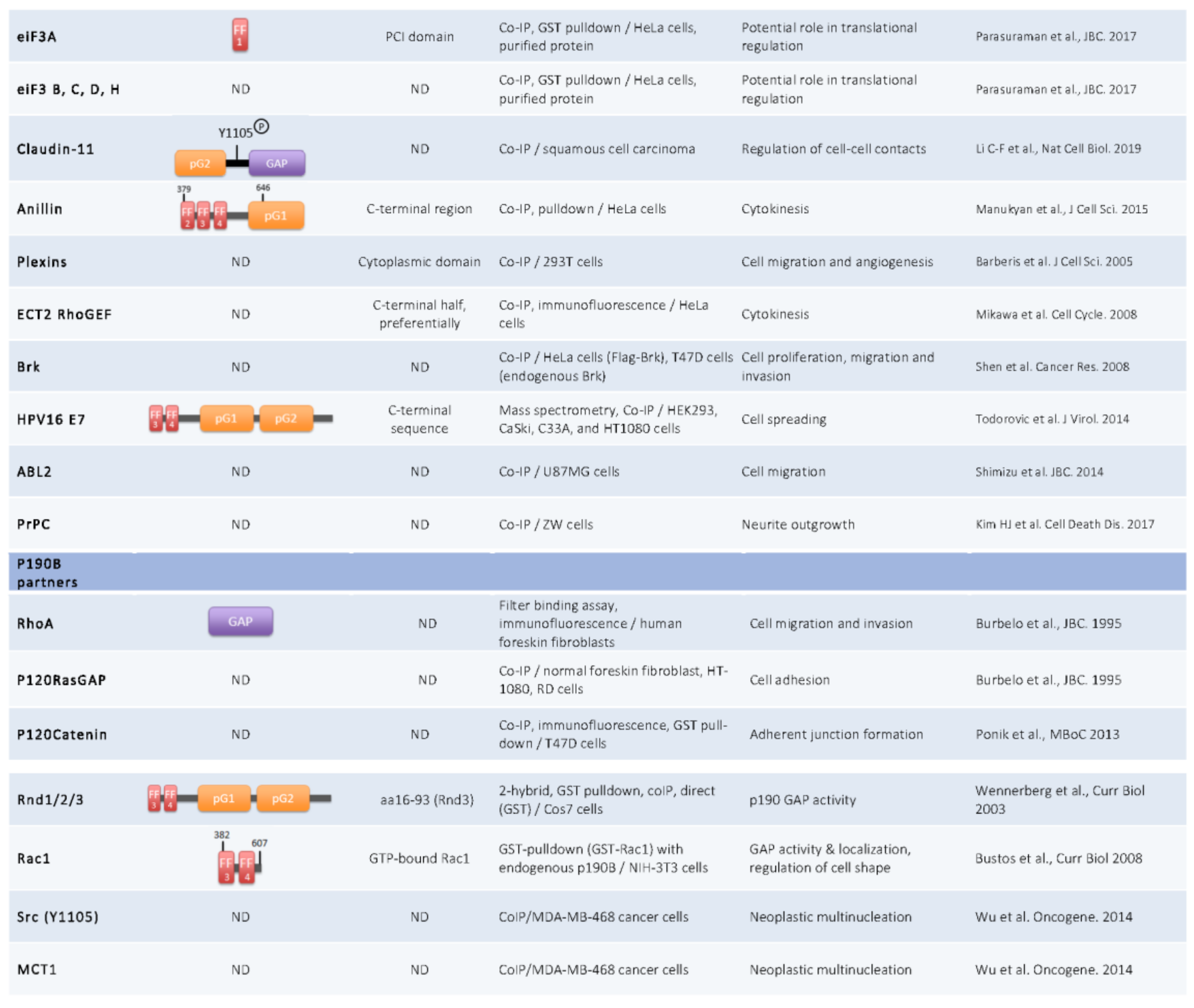{kind=link}
{kind=link}
| Cancer Type | Nb of Tumor/Control | Expression | Detection | Source of Deregulation | Output | References |
|---|---|---|---|---|---|---|
| p190A | ||||||
| Cancer | 3281 T (12 tumor types) | nd | Exome sequencing | Mutations | ARHGAP35 significantly mutated in cancer | Kandoth et al. Nature 2013 |
| 4742 T (21 tumor types) | nd | Exome sequencing | Mutations: 83 missense, 38 nonsense, 16 frameshift and 2 splice site | ARHGAP35 significantly mutated in cancer | Lawrence et al. Nature. 2014 | |
| 191 gliomas + 115 gliomas + 37 ovarian cancers | nd | FISH assay | Locus rearranged/Chromosomal region frequently deleted in solid tumors | Alteration may be responsible of cancerogenesis | Tikoo et al. Gene. 2000 | |
| Osteosarcoma | 247 T | Up | IHC | nd | Poor outcome (larger tumor size, tumor dedifferenciation, high metastatic risk, recurrence-free and overall survival) | Zhao et al. Tumor Biol. 2014 |
| 247 T/428 NT | nd | TaqMan PCR | rs1052667 polymorphism (3′ UTR) | Association with osteosarcoma risk, grade and poor prognosis | Zhao et al. Bio Med Res Int. 2014 | |
| Colorectal cancer | 114 T/NT | Up | qRT-PCR, IHC, WB | nd | Correlation between expression and grade Poor survival outcome | Li et al. Am Transl Res. 2016 |
| 124 T | nd | PCR-SSCP | c.2834dupA > p.Asn946GlufsX11 | Premature stop/Loss-of-function mutation | Ji Choi et al. Pathol. Oncol. Res. 2017 | |
| Lung adenocarcinoma | 133 T/NT | Up | qRT-PCR, IHC | ↗° phospho-p190A | Poor disease-free survival | Notsuda et al. Int J Oncol. 2013 |
| 660 T/NT | nd | Exome sequencing, RNA seq | Missense & frameshift indel or nonsense mutations | ARHGAP35 alteration in oncogene-negative tumors | Campbell et al. Nature Genetics. 2016 | |
| Hepatocellular carcinoma | 37 T/17 NT | Down | PCR array | nd | Potentially responsible of non-viral hepatocarcinogenesis | Kurokawa et al. J Hepatol. 2003 |
| 44 T | nd | IHC | ↗° Grp78 > FAK > phospho-p190A | Link between Grp78 and tumoral invasion | Su et al. BMC Cancer, 2010 | |
| Hürtle Cell Carcinoma | 41 T/NT | nd | Exome sequencing | p.E1273A | Recurrently mutated in HCC (≈5% samples) | Gopal et al. Cancer cell. 2018 |
| Renal angiomyolipoma | 30 T | nd | Sanger sequencing | p.E1273A | Missense variant with uncertain functional effects | Giannikou et al. PLOS Genetics. 2016 |
| p190B | ||||||
| Hepatocellular carcinoma | 47 T | Up | IHC | Positive correlation between the expression levels of CD147 and p190B | Role in HCC progression by regulation of cell movement through p190B and CD147 | Chen et al. Cancer Cell Int. 2016 |
| Nasopharyngeal carcinoma | 52 T | Up | RT-qPCR, WB | ↗°miR-744 > ↗°p190B | Tumor progression | Fang et al. Oncotarget. 2015 |
| Invasive epithelial ovarian cancer | 18,736 T (10,316 of serous histology) / 26,138 NT | Down | eQTL | nd | G allele of AKAP6 rs927062 correlated with reduced expression of ARHGAP5 | Earp et al. PLoS ONE 2018 |
| Lung cancer | 54 T | Up | IHC, qRT-PCR | Inverse correlation between miR-486-5p and ARHGAP5 expression | Positive correlation of p190B expression and cancer stage and lymph node metastasis | Wang et al. Oncogene. 2014 |
| Colorectal adenomatous polyposis | 221 T / 531 NT | Genome-wide analysis of germline CNV, NGS | Somatic point mutations/Partial duplication | One of 11 promising causative candidate genes | Horpaopan et al. Int. J. Cancer. 2015 | |
| Breast cancer | 120 T / 7 NT | Up | qPCR array | Positive correlation between MCT-1 expression and ARHGAP5 expression in tumors | MCT-1 controls cancer cell dissemination and progression through Src/p190B signaling pathway | Wu et al. Oncogene. 2014 |
| Cancer Type | Cell Lines | Loss or Gain of Function Experiment | Output on p190A Activity | Output on Cell Proliferation | Ouput on Cell Migration/Invasion | Other Output | References |
|---|---|---|---|---|---|---|---|
| p190A | |||||||
| Osteosarcoma | MG63 | Loss (si p190A) | nd | Decrease | nd | Increase of apoptosis | Zhao et al. Tumor Biol. 2014 |
| Breast cancer | MDA-MB-231 | Gain (plasmids p190A) | Increase Brk/p190A interaction, phosphorylation of p190A and p190/p120 complex formation | Increase | Increase | Increase tumorigenicity (mice model) | Shen et al. Cancer Res. 2008 |
| MDA-MB-468, MCF10A | Gain (plasmids) | nd | nd | nd | Induction of caspase- and Rho-dependent apoptosis | Ludwig and Parsons. Genes & Cancer. 2011 | |
| Lung adenocarcinoma | A549, LK87, PC-14, H1975 | Loss (si p190A) | Decrease p190/p120 complex formation, Ras inactivation, Rho activation | Cell cycle arrest | Decrease | Inhibition of Ras pathway | Notsuda et al. Int J Oncol. 2013 |
| Glioma | PDGF/Gtv-a cells | Gain (WT GAP domain) | nd | Inhibition of PDGF-induced proliferation | nd | nd | Wolf et al. Genes Dev. 2003 |
| Renal cancer | RCC, 786-O | Loss (si p190A) | nd | nd | nd | Restore fibronectin matrix assembly | Feijoo-Cuaresma. J Biol Chem. 2008 |
| Melanoma | BLM human melanoma cells | Loss (si p190A) | p120ctn allows p190-RhoA interaction and RhoA activation | nd | E-cadherin melanoma transfectant invasion | nd | Molina-Ortiz et al. J Biol Chem. 2009 |
| LOX melanoma cells | Loss (injection of p190 antibody) | nd | nd | Decrease of matrix degradation | No alteration of cell adhesion | Nakahara et al. J Biol Chem. 1998 | |
| Squamous cell carcinoma | 10 SCC cell lines and a primary HNSCC | Loss (sh p190A) | nd | nd | Dissociation of cancer cells | nd | Li et al. Nat Cell Biol. 2019 |
| Pancreatic cancer | AsPC-1, PANC-1 | Gain (p190-RhoA chimera) | Inhibition of RhoA activity (RhoB and RhoC) | No significant effect | Decrease of cell invasion, metastatic formation | nd | Kusama et al. Cancer Sci. 2006 |
| Human Papillomavirus | HaCat, CaSki, C33A, U2OS, HT1080 | Gain (plasmids p190A-HA) | Interaction MD p190A & CR3 of HPV E7 | nd | nd | Decrease actin stress fiber formation, cell spreading | Todorovic et al. J Virology. 2014 |
| Cancer | Huh7, MDA-MB-231, MEF | Gain (plasmids p190A-HA) | ΔPLS, S886F and Δ865-870 increase RhoGAP activity | nd | Alteration of cell migration directionnality | Defect in cell protrusion localisation and lamellipodia persistence | Binamé et al. J Cell Biol. 2016 |
| MDCK cell lines | Loss (sh p190A) | nd | Loss of CIP | nd | Nuclear translocation of YAP, rescue repression of gene transcription of Hippo pathway | Frank et al. J Cell Biol. 2018 | |
| p190B | |||||||
| Breast cancer | MDA-MB-468 | Loss (si p190B) | Interaction between p190B and MCT1 and also Src | Decrease acytokinetic division and neoplastic multinucleation induced by MCT1 | nd | Inhibition of tumor growth | Wu et al. Oncogene. 2014 |
| MCF7-10A, rats | Gain (o/e p190B) | Overexpression of p190B in some murine mammary tumors | nd | nd | Actin cytoskeleton reorganization, loss of adhesion | Chakravarty et al. Cell Growth Differ. 2000 | |
| Neu mice | p190B haploinsufficiency | Affect Rho signaling pathways | nd | nd | Inhibition of tumor initiation and progression, increase tumor free survival | Heckman-Stoddard et al. Breast Cancer Res. 2009 | |
| Hepatocellular carcinoma | Huh7 | Loss (si p190B) | Increase RhoA activity | nd | Increase cell spreading and cell migration | nd | Gen et al. Cancer Lett. 2009 |
| SMMC-7721, Huh7, HepG2 | Loss (si p190B) | Increase of RhoA activity | nd | Decrease migration induced by CD147 | nd | Chen et al. Cancer Cell Int. 2016 | |
| Nasopharyngeal carcinoma | NPC cell lines | Loss (si p190B) | nd | nd | Abrogates migration and invasion induced by miR-744 | nd | Fang et al. Oncotarget. 2015 |
| Cancer | MDCK cell lines | Loss (sh p190B) | nd | Loss of CIP | nd | Nuclear translocation of YAP, rescue repression of gene transcription of Hippo pathway | Frank et al. J Cell Biol. 2018 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Héraud, C.; Pinault, M.; Lagrée, V.; Moreau, V. p190RhoGAPs, the ARHGAP35- and ARHGAP5-Encoded Proteins, in Health and Disease. Cells 2019, 8, 351. https://doi.org/10.3390/cells8040351
Héraud C, Pinault M, Lagrée V, Moreau V. p190RhoGAPs, the ARHGAP35- and ARHGAP5-Encoded Proteins, in Health and Disease. Cells. 2019; 8(4):351. https://doi.org/10.3390/cells8040351
Chicago/Turabian StyleHéraud, Capucine, Mathilde Pinault, Valérie Lagrée, and Violaine Moreau. 2019. "p190RhoGAPs, the ARHGAP35- and ARHGAP5-Encoded Proteins, in Health and Disease" Cells 8, no. 4: 351. https://doi.org/10.3390/cells8040351
APA StyleHéraud, C., Pinault, M., Lagrée, V., & Moreau, V. (2019). p190RhoGAPs, the ARHGAP35- and ARHGAP5-Encoded Proteins, in Health and Disease. Cells, 8(4), 351. https://doi.org/10.3390/cells8040351