Flow Cytometry-Based Characterization of Mast Cells in Human Atherosclerosis

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Methods
2.1. Sample Collection and Processing
2.2. Histology
2.3. Flow Cytometry
2.4. Statistics
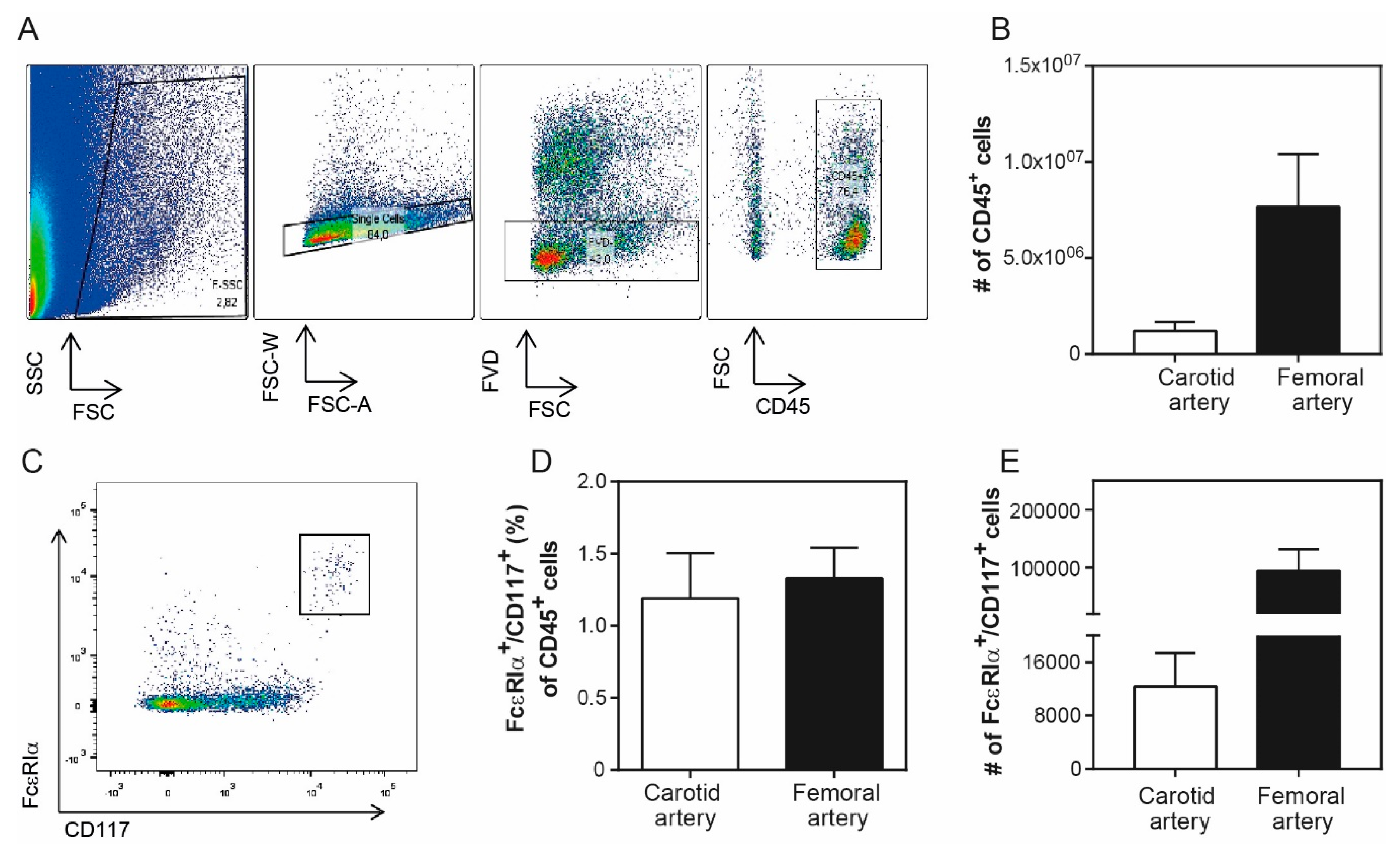
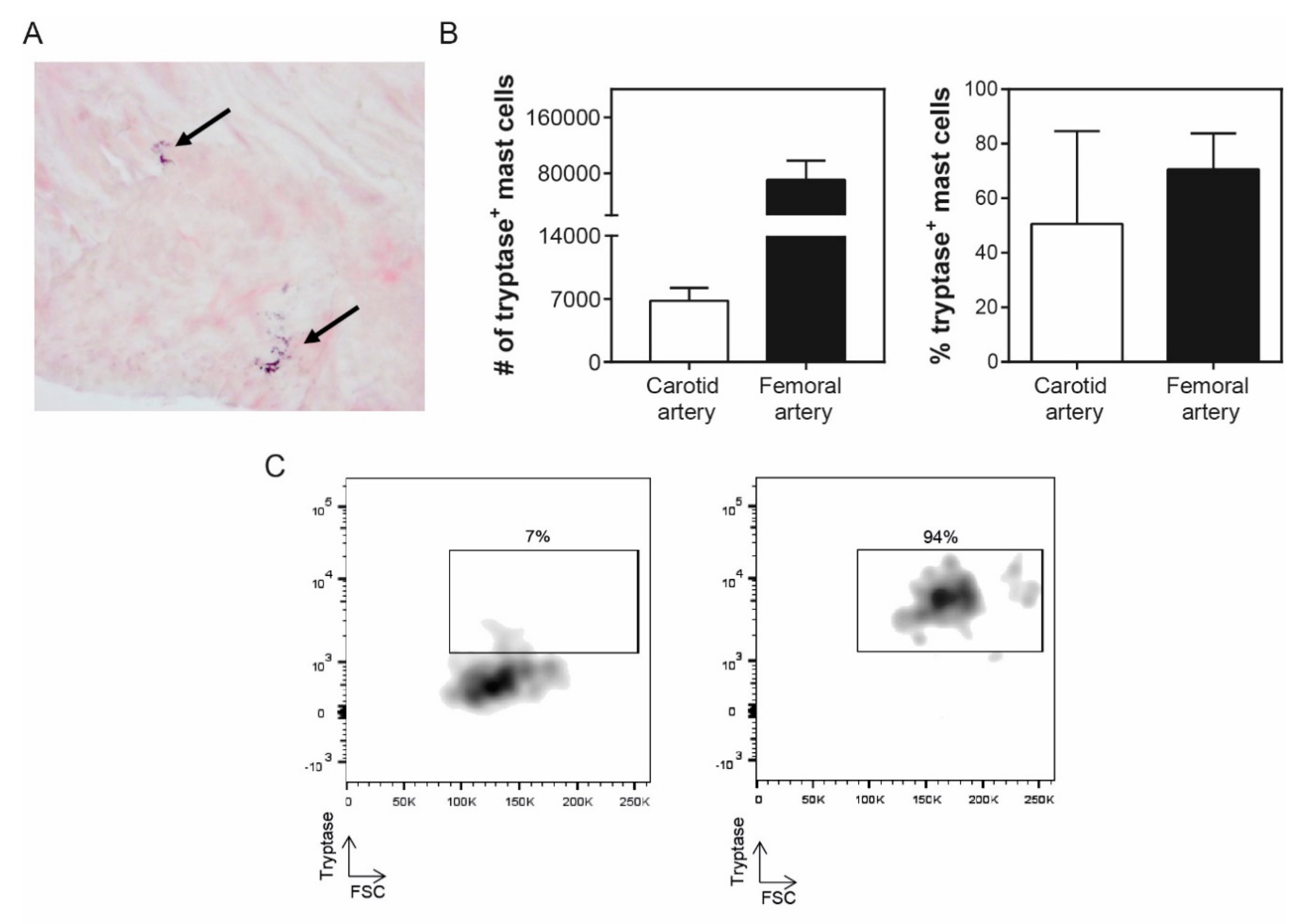
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; de Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart Disease and Stroke Statistics—2017 Update: A Report From the American Heart Association. Circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef]
- Bot, I.; Shi, G.-P.; Kovanen, P.T. Mast cells as effectors in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Amin, K. The role of mast cells in allergic inflammation. Respir. Med. 2012, 106, 9–14. [Google Scholar] [CrossRef]
- Shi, G.-P.; Bot, I.; Kovanen, P.T. Mast cells in human and experimental cardiometabolic diseases. Nat. Rev. Cardiol. 2015, 2, 643–658. [Google Scholar] [CrossRef] [PubMed]
- Kaartinen, M.; Penttila, A.; Kovanen, P.T. Accumulation of activated mast cells in the shoulder region of human coronary atheroma, the predilection site of atheromatous rupture. Circulation 1994, 90, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Kovanen, P.T.; Kaartinen, M.; Paavonen, T. Infiltrates of activated mast cells at the site of coronary atheromatous erosion or rupture in myocardial infarction. Circulation 1995, 92, 1084–1088. [Google Scholar] [CrossRef]
- Bot, I.; Biessen, E.A.L. Mast cells in atherosclerosis. Thromb. Haemost. 2011, 106, 820–826. [Google Scholar]
- Ramalho, L.S.; Oliveira, L.F.; Cavellani, C.L.; Ferraz, M.L.; de Oliveira, F.A.; Miranda Corrêa, R.R.; de Paula Antunes Teixeira, V.; De Lima Pereira, S.A. Role of mast cell chymase and tryptase in the progression of atherosclerosis: Study in 44 autopsied cases. Ann. Diagn. Pathol. 2013, 17, 28–31. [Google Scholar] [CrossRef]
- Mayranpaa, M.I.; Heikkila, H.M.; Lindstedt, K.A.; Walls, A.F.; Kovanen, P.T. Desquamation of human coronary artery endothelium by human mast cell proteases: Implications for plaque erosion. Coron. Artery Dis. 2006, 17, 611–621. [Google Scholar] [PubMed]
- Kaartinen, M.; Penttila, A.; Kovanen, P.T. Mast cells accompany microvessels in human coronary atheromas: Implications for intimal neovascularization and hemorrhage. Atherosclerosis 1996, 123, 123–131. [Google Scholar] [CrossRef]
- Kaartinen, M.; Penttila, A.; Kovanen, P.T. Mast cells of two types differing in neutral protease composition in the human aortic intima. Demonstration of tryptase- and tryptase/chymase-containing mast cells in normal intimas, fatty streaks, and the shoulder region of atheromas. Arterioscler. Thromb. Vasc. Biol. 1994, 14, 966–972. [Google Scholar] [CrossRef]
- Zhi, X.; Xu, C.; Zhang, H.; Tian, D.; Li, X.; Ning, Y.; Yin, L. Tryptase promotes atherosclerotic plaque haemorrhage in ApoE−/− mice. PLoS ONE 2013, 8, e60960. [Google Scholar] [CrossRef] [PubMed]
- Bot, I.; Bot, M.; van Heiningen, S.H.; van Santbrink, P.J.; Lankhuizen, I.M.; Hartman, P.; Gruener, S.; Hilpert, H.; van Berkel, T.J.; Fingerle, J.; et al. Mast cell chymase inhibition reduces atherosclerotic plaque progression and improves plaque stability in ApoE−/− mice. Cardiovasc Res. 2011, 89, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Rohm, I.; Sattler, S.; Atiskova, Y.; Kretzschmar, D.; Pistulli, R.; Franz, M.; Jung, C.; Mall, G.; Kronert, T.; Schulze, P.C.; et al. Increased Number of Mast Cells in Atherosclerotic Lesions Correlates with the Presence of Myeloid but not Plasmacytoid Dendritic Cells as well as Pro-inflammatory T Cells. Clin. Lab. 2016, 62, 2293–2303. [Google Scholar] [CrossRef] [PubMed]
- Willems, S.; Vink, A.; Bot, I.; Quax, P.H.; de Borst, G.J.; de Vries, J.P.; van de Weg, S.M.; Moll, F.L.; Kuiper, J.; Kovanen, P.T.; et al. Mast cells in human carotid atherosclerotic plaques are associated with intraplaque microvessel density and the occurrence of future cardiovascular events. Eur. Heart J. 2013, 34, 3699–3706. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cheng, X.; Xiang, M.X.; Alanne-Kinnunen, M.; Wang, J.A.; Chen, H.; He, A.; Sun, X.; Lin, Y.; Tang, T.T.; et al. IgE stimulates human and mouse arterial cell apoptosis and cytokine expression and promotes atherogenesis in Apoe−/− mice. J. Clin. Investig. 2011, 121, 3564–3577. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Yuan, S.; Liu, Y.; Zeng, Y.; Xie, H.; Liu, Z.; Zhang, S.; Fang, Q.; Wang, J.; Shen, Z. Serum IgE levels are associated with coronary artery disease severity. Atherosclerosis 2016, 251, 355–360. [Google Scholar] [CrossRef]
- Kelley, J.; Hemontolor, G.; Younis, W.; Li, C.; Krishnaswamy, G.; Chi, D.S. Mast cell activation by lipoproteins. Methods Mol. Biol. 2006, 315, 341–348. [Google Scholar]
- Yu, Y.; Blokhuis, B.R.; Garssen, J.; Redegeld, F.A. Non-IgE mediated mast cell activation. Eur. J. Pharmacol. 2016, 778, 33–43. [Google Scholar] [CrossRef]
- Meng, Z.; Yan, C.; Deng, Q.; Dong, X.; Duan, Z.M.; Gao, D.F.; Niu, X.L. Oxidized low-density lipoprotein induces inflammatory responses in cultured human mast cells via Toll-like receptor 4. Cell. Physiol. Biochem. 2013, 31, 842–853. [Google Scholar] [CrossRef]
- de Vries, M.R.; Wezel, A.; Schepers, A.; van Santbrink, P.J.; Woodruff, T.M.; Niessen, H.W.; Hamming, J.F.; Kuiper, J.; Bot, I.; Quax, P.H. Complement factor C5a as mast cell activator mediates vascular remodelling in vein graft disease. Cardiovasc. Res. 2013, 97, 311–320. [Google Scholar] [CrossRef]
- Bot, I.; de Jager, S.C.; Bot, M.; van Heiningen, S.H.; de Groot, P.; Veldhuizen, R.W.; van Berkel, T.J.; von der Thüsen, J.H.; Biessen, E.A. The neuropeptide substance P mediates adventitial mast cell activation and induces intraplaque hemorrhage in advanced atherosclerosis. Circ. Res. 2010, 106, 89–92. [Google Scholar] [CrossRef]
- Verhoeven, B.A.; de Vries, J.P.; Pasterkamp, G.; Ackerstaff, R.G.; Schoneveld, A.H.; Velema, E.; de Kleijn, D.P.; Moll, F.L. Carotid atherosclerotic plaque characteristics are associated with microembolization during carotid endarterectomy and procedural outcome. Stroke 2005, 36, 1735–1740. [Google Scholar] [CrossRef]
- Van Brussel, I.; Ammi, R.; Rombouts, M.; Cools, N.; Vercauteren, S.R.; De Roover, D.; Hendriks, J.M.H.; Lauwers, P.; Van Schil, P.E.; Schrijvers, D.M. Fluorescent activated cell sorting: An effective approach to study dendritic cell subsets in human atherosclerotic plaques. J. Immunol. Methods 2015, 417, 76–85. [Google Scholar] [CrossRef]
- Lovett, J.K.; Gallagher, P.J.; Hands, L.J.; Walton, J.; Rothwell, P.M. Histological correlates of carotid plaque surface morphology on lumen contrast imaging. Circulation 2004, 110, 2190–2197. [Google Scholar] [CrossRef]
- Burd, P.R.; Rogers, H.W.; Gordon, J.R.; Martin, C.A.; Jayaraman, S.; Wilson, S.D.; Dvorak, A.M.; Galli, S.J.; Dorf, M.E. Interleukin 3-dependent and -independent mast cells stimulated with IgE and antigen express multiple cytokines. J. Exp. Med. 1989, 170, 245–257. [Google Scholar] [CrossRef]
- Tsai, M.; Takeishi, T.; Thompson, H.; Langley, K.E.; Zsebo, K.M.; Metcalfe, D.D.; Geissler, E.N.; Galli, S.J. Induction of mast cell proliferation, maturation, and heparin synthesis by the rat c-kit ligand, stem cell factor. Proc. Natl. Acad. Sci. USA 1991, 88, 6382–6386. [Google Scholar] [CrossRef]
- Kraft, S.; Jouvin, M.H.; Kulkarni, N.; Kissing, S.; Morgan, E.S.; Dvorak, A.M.; Schröder, B.; Saftig, P.; Kinet, J.P. The tetraspanin CD63 is required for efficient IgE-mediated mast cell degranulation and anaphylaxis. J. Immunol. 2013, 191, 2871–2878. [Google Scholar] [CrossRef]
- Sinkiewicz, W.; Błazejewski, J.; Bujak, R.; Zekanowska, E.; Sobański, P.; Kubica, J.; Dudziak, J.; Karasek, D.; Małyszka, P.; Balak, W.; et al. Immunoglobulin E as a marker of the atherothrombotic process in patients with acute myocardial infarction. Cardiol. J. 2007, 14, 266–273. [Google Scholar]
- Langer, R.D.; Criqui, M.H.; Feigelson, H.S.; McCann, T.J.; Hamburger, R.N. IgE predicts future nonfatal myocardial infarction in men. J. Clin. Epidemiol. 1996, 49, 203–209. [Google Scholar] [CrossRef]
- Lippi, G.; Cervellin, G.; Sanchis-Gomar, F. Immunoglobulin E (IgE) and ischemic heart disease. Which came first, the chicken or the egg? Ann. Med. 2014, 46, 456–463. [Google Scholar] [CrossRef]
- Insull, W.J. The pathology of atherosclerosis: Plaque development and plaque responses to medical treatment. Am. J. Med. 2009, 122, S3–S14. [Google Scholar] [CrossRef]
- Laske, N.; Bunikowski, R.; Niggemann, B. Extraordinarily high serum IgE levels and consequences for atopic phenotypes. Ann. Allergy. Asthma Immunol. 2003, 91, 202–204. [Google Scholar] [CrossRef]
- Wang, L.; Gao, S.; Xu, W.; Zhao, S.; Zhou, J.; Wang, N.; Yuan, Z. Allergic asthma accelerates atherosclerosis dependent on Th2 and Th17 in apolipoprotein E deficient mice. J. Mol. Cell. Cardiol. 2014, 72, 20–27. [Google Scholar] [CrossRef]
- Abd-Elmoniem, K.Z.; Ramos, N.; Yazdani, S.K.; Ghanem, A.M.; Holland, S.M.; Freeman, A.F.; Gharib, A.M. Coronary atherosclerosis and dilation in hyper IgE syndrome patients: Depiction by magnetic resonance vessel wall imaging and pathological correlation. Atherosclerosis 2017, 258, 20–25. [Google Scholar] [CrossRef][Green Version]
- Indhirajanti, S.; van Daele, P.L.A.; Bos, S.; Mulder, M.T.; Bot, I.; Roeters van Lennep, J.E. Systemic mastocytosis associates with cardiovascular events despite lower plasma lipid levels. Atherosclerosis 2018, 268, 152–156. [Google Scholar] [CrossRef]
- Bot, I.; de Jager, S.C.; Zernecke, A.; Lindstedt, K.A.; van Berkel, T.J.; Weber, C.; Biessen, E.A. Perivascular mast cells promote atherogenesis and induce plaque destabilization in apolipoprotein E-deficient mice. Circulation 2007, 115, 2516–2525. [Google Scholar] [CrossRef]
- Lagraauw, H.M.; Westra, M.M.; Bot, M.; Wezel, A.; van Santbrink, P.J.; Pasterkamp, G.; Biessen, E.A.; Kuiper, J.; Bot, I. Vascular neuropeptide Y contributes to atherosclerotic plaque progression and perivascular mast cell activation. Atherosclerosis 2014, 235, 196–203. [Google Scholar] [CrossRef]
- Finn, D.F.; Walsh, J.J. Twenty-first century mast cell stabilizers. Br. J. Pharmacol. 2013, 170, 23–37. [Google Scholar] [CrossRef]
- Logsdon, S.L.; Oettgen, H.C. Anti-IgE therapy: Clinical utility and mechanistic insights. Curr. Top. Microbiol. Immunol. 2015, 388, 39–61. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | 0 | 1 | 2 | 3 | 4 |
|---|---|---|---|---|---|
| Necrotic Core | No sign of necrotic core | <20% of the arterial area | 20–40% of the arterial area | 40–70% of the arterial area | >70% |
| Calcification | No sign of calcification | <10% of the arterial area | 10–40% of the arterial area | >40% of the arterial area | |
| Foam Cells | No sign of foam cells | Small * A few small sized clusters (~5) of foam cells | Intermediate * Multiple small (~5) or medium sized (~10) clusters of foam cells | Large * At least 1 large sized cluster (~15) of foam cells or foam cells scattered around ~70% of the arterial area | |
| Cholesterol Crystal | No sign of cholesterol crystal | <10% of the necrotic core | 10–40% of the necrotic core | >40% of the necrotic core | |
| Neovasculari-zation | No sign of neovasculari-zation | <25 neovessels in the whole tissue | 25–50 neovessels in whole tissue | >50 neovessels in the whole tissue | |
| Inflammatory Cells | No sign of inflammatory cells | Small * A few (1 or 2) small sized (~50 cells) clusters of inflammatory cells or a few cells occasionally spread throughout the arterial area | Intermediate * Multiple (~5) small sized (~50 cells) clusters of inflammatory cells | Large * At least 1 large sized (~100 cells) cluster of inflammatory cells or cells scattered around ~70% of the arterial area | |
| Shoulder | Not detectable * | No sign of shoulder regions | One sided shoulder region | Two-sided shoulder region | |
| Intraplaque hemorrhage (IPH) | No sign of IPH | <10% IPH of the arterial area | 10–40% IPH of the arterial area | >40% IPH of the arterial area | |
| SMC-rich ECM area of total ECM area | No SMC visible | <20% of the ECM area | 20–40% of the ECM area | >40% of the ECM area | |
| Luminal thrombosis | No | Yes (rupture) |
| Antibody | Fluorochrome | Clone | Concentration | Company |
|---|---|---|---|---|
| Fixable Viability Dye | eFluor 780 | - | 0.1 μg/sample | eBioscience |
| CD45 | PB | 2D1 | 0.25 μg/sample | eBioscience |
| FcεRIα | APC/PE Cy7 | AER-37 | 0.12 μg/sample | eBioscience |
| CD117 | PercP Cy5.5 | 104D2 | 0.1 μg/sample | eBioscience |
| CD63 | PE | H5C6 | 0.1 μg/sample | eBioscience |
| IgE | PE Cy7/FITC | Ige21 | 0.25 μg/sample | eBioscience |
| Tryptase/TPSAB1 | PE | - | 0.1 μg/sample | LSBio |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kritikou, E.; Depuydt, M.A.C.; de Vries, M.R.; Mulder, K.E.; Govaert, A.M.; Smit, M.D.; van Duijn, J.; Foks, A.C.; Wezel, A.; Smeets, H.J.; et al. Flow Cytometry-Based Characterization of Mast Cells in Human Atherosclerosis. Cells 2019, 8, 334. https://doi.org/10.3390/cells8040334
Kritikou E, Depuydt MAC, de Vries MR, Mulder KE, Govaert AM, Smit MD, van Duijn J, Foks AC, Wezel A, Smeets HJ, et al. Flow Cytometry-Based Characterization of Mast Cells in Human Atherosclerosis. Cells. 2019; 8(4):334. https://doi.org/10.3390/cells8040334
Chicago/Turabian StyleKritikou, Eva, Marie A.C. Depuydt, Margreet R. de Vries, Kevin E. Mulder, Arthur M. Govaert, Marrit D. Smit, Janine van Duijn, Amanda C. Foks, Anouk Wezel, Harm J. Smeets, and et al. 2019. "Flow Cytometry-Based Characterization of Mast Cells in Human Atherosclerosis" Cells 8, no. 4: 334. https://doi.org/10.3390/cells8040334
APA StyleKritikou, E., Depuydt, M. A. C., de Vries, M. R., Mulder, K. E., Govaert, A. M., Smit, M. D., van Duijn, J., Foks, A. C., Wezel, A., Smeets, H. J., Slütter, B., Quax, P. H. A., Kuiper, J., & Bot, I. (2019). Flow Cytometry-Based Characterization of Mast Cells in Human Atherosclerosis. Cells, 8(4), 334. https://doi.org/10.3390/cells8040334