Abstract
Hedgehog (Hh) signaling is a critical developmental regulator and its aberrant activation, due to somatic or germline mutations of genes encoding pathway components, causes Basal Cell Carcinoma (BCC) and medulloblastoma (MB). A growing effort has been devoted at the identification of druggable vulnerabilities of the Hedgehog signaling, leading to the identification of various compounds with variable efficacy and/or safety. Emerging evidence shows that an aberrant polyamine metabolism is a hallmark of Hh-dependent tumors and that its pharmacological inhibition elicits relevant therapeutic effects in clinical or preclinical models of BCC and MB. We discuss here the current knowledge of polyamine metabolism, its role in cancer and the available targeting strategies. We review the literature about the connection between polyamines and the Hedgehog signaling, and the potential therapeutic benefit of targeting polyamine metabolism in two malignancies where Hh pathways play a well-established role: BCC and MB.
1. Polyamines: Metabolism, Mechanism of Action and Role in Tumorigenesis
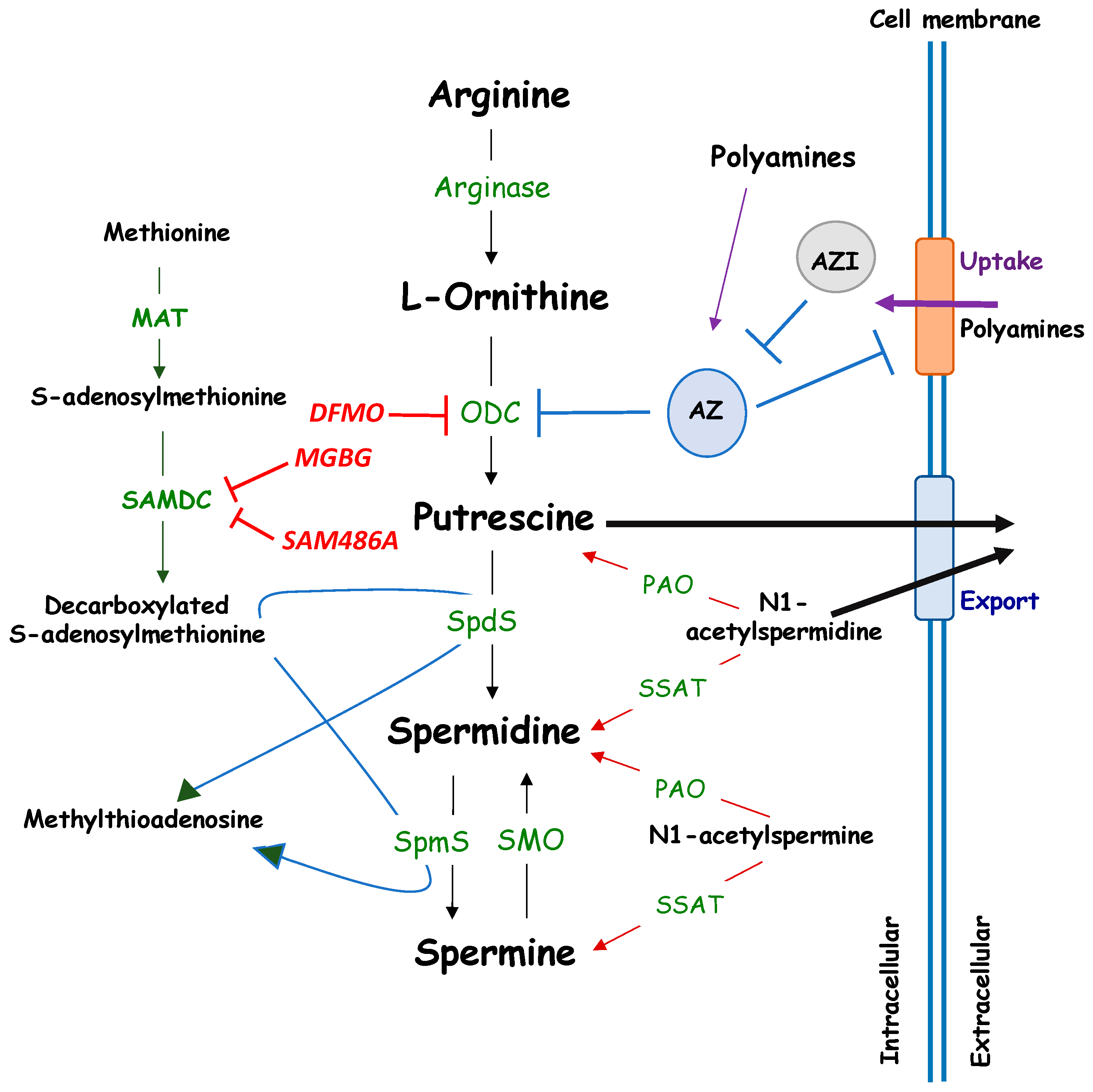
Polyamines are ubiquitous, small, positively charged polycations required for both eukaryotic and prokaryotic cellular growth and differentiation. They are low molecular weight, water soluble, aliphatic amines, with pK values between 8.3 and 10.9. Polyamines bind negatively charged molecules under physiological conditions, including DNA, RNA, ATP, proteins or phospholipids [1,2] and are important regulators of various physiological processes. The naturally occurring polyamines spermine and spermidine are the biosynthetic products of putrescine, which is derived from ornithine by the action of ornithine decarboxylase (ODC), one rate-limiting enzyme, together with S-adenosyl-methionine decarboxylase [3] (SAMDC) [4] (Figure 1).
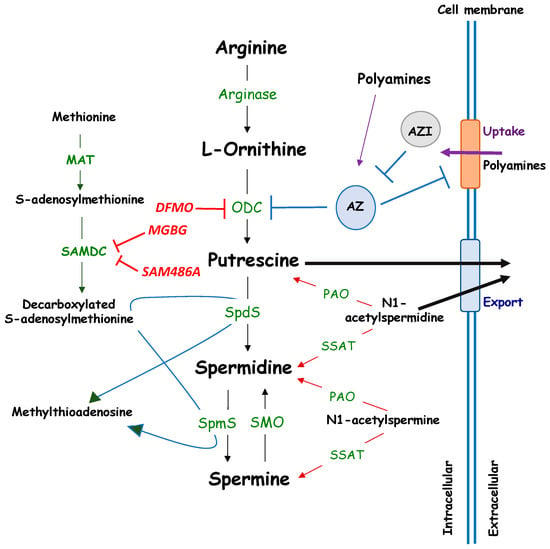
Figure 1.
Metabolism of polyamines. MAT: Methionine adenosyl-transferase; ODC: Ornithine decarboxylase; AZ: Antizyme; AZI: Antizyme Inhibitor; PAO: Polyamine oxidase; SAMDC: S-Adenosyl-l-Methionine Decarboxylase; SMO: Spermine oxidase; SSAT: Spermidine/Spermine-N(1)-acetyltransferase; SpdS: Spermidine Synthase; SpmS: Spermine Synthase. Pharmacological inhibitors of ODC (DFMO) and SAMDC (MGBG, SAM486A) are indicated in red.
During cancer cell growth, the polyamine biosynthetic pathway is very active and polyamine content is increased in tumor cells and tissues, such as breast, colon, skin and prostate cancers [5]. The elevated polyamine content is often linked to increased putrescine synthesis by ornithine decarboxylase, as well as to increased polyamine uptake [6]. Several studies indicate that depletion of polyamines leads to inhibition of tumor growth [7,8,9].
A key role in regulating intracellular polyamine homeostasis is played by the ornithine decarboxylase enzyme, which is tightly regulated at multiple levels in normal and cancer cells, to promptly adjust the levels of polyamines in response to the specific cellular needs [10]. A first level of control of ODC content is through its stability. Indeed, ODC is a very short-lived protein and its rapid turnover is mediated by the proteasome in an ubiquitin-independent manner [11]. A protein called antizyme (AZ) associates with ODC and increases its targeting to the proteasome and consequent degradation [12]. Interestingly, AZ synthesis is induced by polyamines, leading to enhanced association to ODC monomers to form heterodimers and preventing the formation of functional ODC homodimers [11].
A second level of control of ODC content is trough its transcriptional regulation. ODC mRNA levels are regulated by various cues and transcription factors, being the oncogene MYC the best-characterized and more relevant regulator [13,14]. Indeed, the promoter region of the Odc gene contains two canonical E boxes (CACGTG) that bind MYC/Max transcription factors. Consistently, increased ODC expression is observed when MYC is upregulated, such as in cancer [15,16]. A third level of control of ODC expression is via its translation. The ODC mRNA has a long 5′ untranslated region (UTR) of about 300 nucleotides and is enhanced by elevated levels of eIF-4E [17], which binds the cap structure to initiate translation. Alternatively, ODC can be translated independently of cap-mediated initiation, using an internal ribosome entry site (IRES) located in the 5′ UTR [18]. This site would be used only under certain conditions such as in the G2/M phase of the cell cycle, or in response to developmental stimuli (see below).
Both ODC and AZ play an important role in carcinogenesis, as documented by studies in animal models. Targeted expression of an active C-terminally truncated form of ODC, under the control of keratin promoter significantly increased skin tumor development in mice treated with carcinogens or UV radiation or expressing active Ras [19,20,21,22]. Conversely, mice heterozygous for Odc gene (Odc +/−) developed substantially fewer skin papillomas when treated with a tumor-promoting agent [22]. Carcinogenesis was also reduced in mice expressing AZ under the keratin promoter and exposed to chemical of physical carcinogens [23], thus underscoring the relevance of ODC expression during skin carcinogenesis. In addition to skin tumors, Odc haploinsufficiency has been shown to significantly reduce Myc-induced lymphoma development in Eμ-Myc transgenic Odc +/− mice [24].
In agreement with these results, the use of the specific ODC inhibitor, DFMO (d,l-alpha-difluoromethylornithine), led to tumor reduction in animal models of different tumors [25].
Another key regulator of polyamine metabolism with relevance in tumor disease is the SAMDC enzyme, which catalyzes the decarboxylation of S-Adenosylmethionine (SAM) into decarboxylated SAM (dc-SAM). Dc-SAM is the aminopropyl donor for the synthesis of spermidine and sperimine, catalyzed by SpdS and SpmS respectively (Figure 1). SAMDC has been recently found upregulated by mTORC1 in prostate cancer via phosphorylation-mediated stabilization, thus providing an important link between the oncogenic nutrient-sensing machinery and polyamine metabolism and suggesting the potential therapeutic benefit of its targeting [26].
Given the role of the natural polyamines in cancer and growth-related processes, great efforts have been made to synthesize inhibitors for the enzymes involved in polyamine biosynthesis: spermidine and spermine synthase [27] ornithine decarboxylase [28] and S-adenosyl-methionine decarboxylase [29].
Strategies for cancer treatment are currently under development using:
- Inhibitors of polyamine synthesis: (i) DFMO, a specific inhibitor of ornithine decarboxylase; currently, DMFO has been clinically tested in gliomas, neuroblastoma, colon, prostate and non melanoma skin cancer (NMSC, see below) [30]. (ii) methylglyoxal-bis-guanidylhydrazone (MGBG), an inhibitor of S-adenosyl-methionine decarboxylase [3], which reduces spermidine and spermine levels but elevates putrescine levels [31]. Although MGBG is an effective SAMDC inhibitor, its use in chemotherapy is restricted because of its mitochondrial toxicity [4]. (iii) SAM486A (4-amidinoindan-1-one-2′-amidinhydrazone) a derivative of MGBG. Despite it was tested in various cancer cells and animal systems, as well as in phase I and II clinical trials for activity against adult cancers, it resulted ineffective [31] probably because of the induction of compensatory mechanisms, which preserve the intracellular concentrations of polyamines [7].
- Analogues of polyamines [32] which can deplete polyamine content and interfere with polyamine metabolism and/or function.
- Polyamine transport inhibitors which can prevent uptake of exogenous polyamines by blocking membrane transporters [33].
- Polyamine-degrading enzymes such as bovine serum amine oxidase (BSAO: EC 1.4.3.6) [34]. It was observed that the oxidative deamination of spermine by BSAO (bovine serum amine oxidase) generates ammonia and the cytotoxic metabolites hydrogen peroxide and aldehydes. Formation of cytotoxic aldehydes from polyamines or reactive oxygen species (ROS) may have potential in cancer therapy, in analogy to other radical forming processes [35], since these molecules are able to induce stress-activated signal transduction pathways, leading to apoptotic and non-apoptotic cell death, in several cultured tumor cell lines [36]. It has previously been demonstrated that hydrogen peroxide and aldehydes generated by BSAO/spermine enzymatic system were also able to overcome multidrug resistance (MDR) in cancer cells [37]. Therefore, toxic polyamine metabolites are currently explored as probable candidates for a new strategy in tumor therapy [35].
2. Hedgehog-Signaling and Its Targeting in Cancer
Hedgehog signaling regulates embryonic development and stem cell fate and its inappropriate activation causes different forms of cancer [38].
Transmembrane receptors and post-receptor proteins mediate the signal transduction, a process where the primary cilium plays a central role [39]. The receptor apparatus consists in (i) the inhibitory receptor patched (PTCH); and (ii) the transmembrane activator Smoothened (SMO). At the post-receptor level, the cytoplasmic regulator SUFU and the GLI transcription factors (GLI1, GLI2, GLI3) are the key mediators of the execution of the Hh transcriptional program [40] (Figure 2).
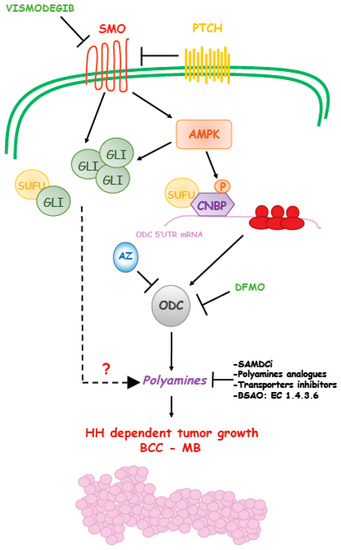
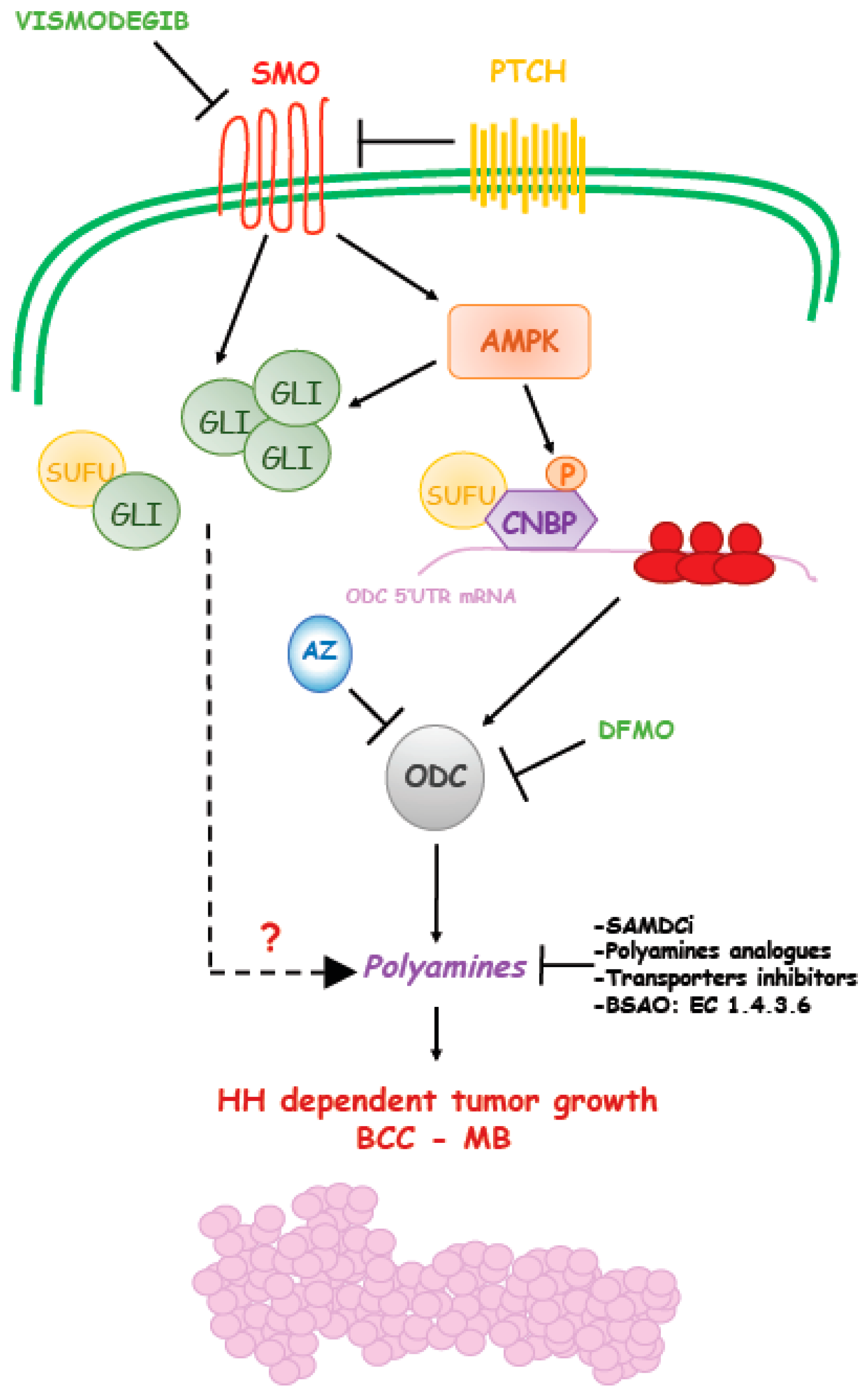
Figure 2.
Schematic representation of Hedgehog-induced polyamines production during cancer formation. Hedgehog signaling induces polyamines levels through upregulation of ODC translation.
Germline loss-of-function mutations of the PTCH gene are found in the Gorlin syndrome, a genetic disease characterized by multiple skin tumors named Basal Cell Carcinoma (BCC) and, with lower frequency, by the brain tumors medulloblastoma (MB) [41].
Sporadic mutations of PTCH1 or other genes encoding Hh pathway components, such as SMO, SUFU, and GLI2, can be found in nearly all BCCs [42,43] and in about one third of MBs (SHH-MB subgroup) [44,45,46] with various frequency (Table 1). In all cases, the overall consequence of these mutations is the constitutive activation of the Hh pathway, which thus represents a key tumorigenic driver for those types of malignancies.

Table 1.
Frequency of mutations in Hh-dependent MB and BCC.
The Hedgehog pathway can be targeted with drugs acting at different levels. Vismodegib has been the first FDA-approved Hh inhibitor and acts by inhibiting the Hh transmembrane activator Smoothened (SMO) [50]. However, despite the good efficacy observed in a subset of patients, results obtained with Vismodegib have been disappointing for two main reasons: (1) the drug is not effective in patients carrying mutations localized downstream of SMO [51,52]; (2) Even in patients that showed good response, tumor cells eventually become drug resistant, due to novel SMO mutations or activation of compensatory mechanisms that restore the function of the downstream effectors [38,53,54,55]. For these reasons, the identification of novel drugs targeting post-receptor Hh components is now a major goal in these malignancies. Several compounds with the ability to directly inhibit GLI transcription factors have been identified, but their efficacy, specificity and safety are still major issues that prevent their immediate use in clinical settings [56]. Also, since GLI activity is modulated by various post-translational modifications, another potential approach would be to use drugs with the ability to affect the different GLI modifiers, such as class I histone deacetylases (HDACs) [57,58,59,60,61,62,63], AMPK [64,65], ERK [66], MEKK1 [67], PIN1 [68].
An alternative strategy would be to target actionable Hh-regulated pathways or metabolisms that are required for tumor growth, using drugs of proven clinical efficacy and safety [38,69]. Among the various options, polyamine metabolism offers a reasonable therapeutic opportunity.
3. Targeting Polyamine Metabolism in Basal Cell Carcinoma
Basal cell carcinoma (BCC) is the most frequent tumor in humans, affecting about 3% of the population [70]. BCC rarely results in death but is often locally destructive, representing a significant economic burden to the health care system [71].
The lesion is typically treated with surgical excision, although there is a significant risk of recurrence. Metastasis occurs rarely, in 0.003–0.1% of patients [72].
The major risk factor of BCC is the prolonged exposure to UV light; hence the main preventive strategy is the reduction of sun exposure and the use of sunscreens. However, despite the risk associated to UV exposure is well known between the population, the incidence of BCC is still increasing over the time, thus raising the interest on the identification of additional preventive strategies [73,74].
Nearly all BCCs display aberrant activation of the Sonic Hedgehog pathway, due to loss of function mutations of PTCH [42,75] or activating mutations of SMO [76] and the Hedgehog inhibitor Vismodegib is approved for the treatment of locally advanced BCC (LaBCC) or metastatic BCC (mBCC) [77]. However, less than 50% of patients with advanced or metastatic BCCs respond to Vismodegib, and an additional 20% acquires secondary resistance during the first year of treatment [77,78,79]. Of note, drug-resistant tumors in patients maintain activation of Hedgehog target genes [55], witnessing an unchanged addiction to the Hedgehog pathway, due to activation of compensatory mechanisms that restore GLI function.
The adverse effects associated with Vismodegib are mild to moderate and include nausea, vomiting, diarrhea, constipation, muscle spasms, fatigue, hair loss, and dysgeusia (distortion of the sense of taste). Thus, the use of Vismodegib for treatment or prevention of BCC raises some legitimate concerns due to the significant risk to develop drug-resistance and the severity of the side effects, especially in case of long-term treatments.
Another key molecular alteration underlying BCC tumorigenesis is an increase of intracellular polyamines, mostly as a consequence of ODC overexpression. Early studies demonstrated that ODC activity is elevated in BCC, compared to normal tissues [80]. It was also shown that tumor promoting agents such as TPA [81] or UV light [82] induce ODC and polyamines, while ODC inhibition significantly limits skin tumor promotion [83].
The first preclinical evidence of a beneficial effect of targeting polyamine metabolism in Hh-induced BCC was documented in mice carrying heterozygous deletion of the Ptch1 gene (Ptch1+/−). When exposed to UVB, these mice develop multiple and severe BCCs with high penetrance by the 20th week of irradiation. Tang et al. [23] demonstrated the importance of ODC and polyamines in this context. They observed that K6 promoter-driven expression of antizyme (AZ), a protein that specifically degrades ODC, in UVB-exposed Ptch1+/− mice prevented BCC formation. Similarly, oral administration of DFMO to UVB-induced Ptch1+/− mice strongly reduced the number and severity of the skin lesions. Thus, these observations demonstrated the relevance of polyamine metabolism in the pathogenesis of Hh-dependent BCC and provided a compelling preclinical evidence of the potential benefit that could result from the inhibition of a key polyamine controller, ODC, with a drug of known clinical efficacy and safety [23].
Following this observation, a few years later it was reported a phase III study on the cancer prevention ability of DFMO in patients with previous history of skin cancer. By evaluating a group of patients with a history of prior non-melanoma skin cancer (NMSC), randomized to 500 mg/m2/day oral DFMO or placebo for 4/5 years, Bailey and colleagues reported the first successful clinical trial of BCC prevention. DFMO significantly reduced new BCC, was well tolerated, with evidence of mild ototoxicity [71,84]. This study was updated two years later, retrospectively assessing the further incidence of skin cancer and adverse events in the same study cohort. The results showed that up to five years of DFMO treatment did not result in latent toxicity or in a rebound increase in rate of BCC after stopping DFMO [85]. Further studies would be appropriate to evaluate the effect of DFMO in combination with other agents.
4. Targeting Polyamine Metabolism in Medulloblastoma
Medulloblastoma represents the most common brain malignancy of the childhood, accounting for almost 20% of all cancers in children aged 0–19 [86]. The current therapeutic approach, based on a combination of surgery, craniospinal radiation and chemotherapy allows about 70% of survivors [87,88], although the majority of patients develop severe long-term side effects, mostly neurological [89,90,91].
Molecular analyses have identified four main molecular subgroups of MB, based on gene expression profiles: WNT, SHH, G3 and G4 [92,93,94]. Tumors of the SHH subgroup (SHH MB) show aberrant activation of the Hh signaling and account for about one third of total MBs. Similar to BCC, they are vulnerable to Hh inhibitors, although trials in patients have shown that they become quickly resistant to Vismodegib. Also, SHH MBs with mutations of SUFU or amplification of GLI2 do not respond at all to this drug [95]. Thus, also for this malignancy there is an emerging need to identify additional drugs targeting post-receptor components of the signaling or regulated metabolic steps. In contrast to BCC, the correlation between SHH MB and polyamine metabolism is less documented. An old report by Scalabrino et al. identified an association between elevated ODC activity and medulloblastoma [96]. However, the distinction of MBs into various molecular subgroups was unknown at that time.
A recent paper showed the correlation of elevated ODC and polyamines with SHH MB molecular subgroup and provided the mechanistic basis of this alteration and its link with Hedgehog signaling [97]. It was observed that SMO activation triggers a non-canonical route that engages the energy sensor AMPK. The key mediator of this response is an RNA binding protein, named CNBP (Cellular Nucleic acid Binding Protein). AMPK phosphorylates CNBP at threonine 173 and this modification increases its association with SUFU and stabilization. The SUFU/CNBP complex binds the 5′ UTR of ODC mRNA and promotes its translation, thereby increasing polyamine metabolism. Thus, SMO activation promotes ODC translation via CNBP. This Hh-regulated translational process is required to support the proliferation of neuronal precursors and it is aberrantly activated in SHH MB, a tumor arising from these cells. Of note, in analogy to what was observed in BCC, targeting this process with the ODC inhibitor DFMO efficiently counteracts the growth of SHH MB in preclinical models. Hence, this report unveiled a novel druggable mechanism of the Hh signaling, where AMPK functions as a key tumor-promoting kinase, which regulates polyamine metabolism and supports tumor growth [97,98] (Figure 2). As a consequence, it would be expected that AMPK blockade would result in inhibition of this non-canonical Hedgehog signaling as well as inhibition of ODC translation and polyamine metabolism and suppression of tumor growth.
In keeping with this evidence, a very recent report showed that the lack of AMPKα2 in mouse cerebellum limits tumorigenesis of SHH MB through downregulation of CNBP and ODC [99]. Thus, in contrast to other reports [65,100,101] it was found that AMPK promotes SHH MB growth in mice, thus raising concerns about the use of agonists of this kinase for therapeutic purposes. Of note, in the same report the authors also found a copy number gain of the genes encoding CNBP and ODC, further supporting the pro-tumorigenic role of this non-canonical axis in SHH MB.
In contrast to BCC, clinical evidence on the efficacy of DFMO on SHH MB patients are still unavailable. A clinical trial of DFMO as a single agent on subjects with relapsed rare tumors, including medulloblastoma, has recently started (ClinicalTrials.gov identifier (NCT number: NCT03581240). Results from this study will provide critical information to understand the efficacy of the polyamine blockade to treat this type of tumor.
Author Contributions
G.C., E.A., S.C. and L.D.M. wrote and edited the paper, Y.K. and S.M.S. revised and edited the paper.
Funding
This work was supported by AIRC grant IG17575, Anna Tramontano Grant, Istituto Pasteur, Fondazione Cenci-Bolognetti, AFM Telethon grant, ‘La Sapienza’ University of Rome and Italian MIUR (Ministero dell’Istruzione, dell’Università e della Ricerca).
Acknowledgments
The authors thank the “International Polyamine Foundation—ONLUS” for the availability to look up in the Polyamines documentation.
Conflicts of Interest
The authors declare no conflict of interest, financial or otherwise.
Abbreviations
Smo: Smoothened; Ptch: Patched; GLI: Glioma-associated oncogene; SUFU: Suppressor of Fused; SHH MB: Sonic Hedgehog type medulloblastoma; HDAC: Histone deacetylase; AMPK: 5′ adenosine monophosphate-activated protein kinase; ERK: extracellular signal–regulated kinases; MEKK1: Mitogen-activated protein kinase kinase kinase; TPA: tetradecanoyl phorbol acetate; WNT: Wingless/Integrated.
References
- Minois, N. Molecular basis of the ‘anti-aging’ effect of spermidine and other natural polyamines—A mini-review. Gerontology 2014, 60, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, K.; Kashiwagi, K. Modulation of cellular function by polyamines. Int. J. Biochem. Cell Biol. 2010, 42, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Bacchi, C.J.; Brun, R.; Croft, S.L.; Alicea, K.; Buhler, Y. In vivo trypanocidal activities of new S-adenosylmethionine decarboxylase inhibitors. Antimicrob. Agents Chemother. 1996, 40, 1448–1453. [Google Scholar] [CrossRef] [PubMed]
- Bae, D.H.; Lane, D.J.R.; Jansson, P.J.; Richardson, D.R. The old and new biochemistry of polyamines. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2053–2068. [Google Scholar] [CrossRef] [PubMed]
- Cervelli, M.; Pietropaoli, S.; Signore, F.; Amendola, R.; Mariottini, P. Polyamines metabolism and breast cancer: State of the art and perspectives. Breast Cancer Res. Treat. 2014, 148, 233–248. [Google Scholar] [CrossRef]
- Heby, O.; Persson, L. Molecular genetics of polyamine synthesis in eukaryotic cells. Trends Biochem. Sci. 1990, 15, 153–158. [Google Scholar] [CrossRef]
- Gamble, L.D.; Hogarty, M.D.; Liu, X.; Ziegler, D.S.; Marshall, G.; Norris, M.D.; Haber, M. Polyamine pathway inhibition as a novel therapeutic approach to treating neuroblastoma. Front. Oncol. 2012, 2, 162. [Google Scholar] [CrossRef]
- Seiler, N.; Atanassov, C.L.; Raul, F. Polyamine metabolism as target for cancer chemoprevention (review). Int. J. Oncol. 1998, 13, 993–1006. [Google Scholar] [CrossRef]
- Devens, B.H.; Weeks, R.S.; Burns, M.R.; Carlson, C.L.; Brawer, M.K. Polyamine depletion therapy in prostate cancer. Prostate Cancer Prostatic Dis. 2000, 3, 275–279. [Google Scholar] [CrossRef]
- Pegg, A.E. Regulation of ornithine decarboxylase. J. Biol. Chem. 2006, 281, 14529–14532. [Google Scholar] [CrossRef]
- Hayashi, S.; Murakami, Y. Rapid and regulated degradation of ornithine decarboxylase. Biochem. J. 1995, 306 Pt 1, 1–10. [Google Scholar] [CrossRef]
- Coffino, P. Regulation of cellular polyamines by antizyme. Nat. Rev. Mol. Cell. Biol. 2001, 2, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E.; Shantz, L.M.; Coleman, C.S. Ornithine decarboxylase as a target for chemoprevention. J. Cell. Biochem. 1995, 22, 132–138. [Google Scholar] [CrossRef]
- Deng, W.; Jiang, X.; Mei, Y.; Sun, J.; Ma, R.; Liu, X.; Sun, H.; Tian, H.; Sun, X. Role of ornithine decarboxylase in breast cancer. Acta Biochim. Biophys. Sin. 2008, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Packham, G.; Cleveland, J.L. Induction of ornithine decarboxylase by IL-3 is mediated by sequential c-Myc-independent and c-Myc-dependent pathways. Oncogene 1997, 15, 1219–1232. [Google Scholar] [CrossRef]
- Nilsson, J.A.; Maclean, K.H.; Keller, U.B.; Pendeville, H.; Baudino, T.A.; Cleveland, J.L. Mnt loss triggers Myc transcription targets, proliferation, apoptosis, and transformation. Mol. Cell. Biol. 2004, 24, 1560–1569. [Google Scholar] [CrossRef]
- Shantz, L.M.; Pegg, A.E. Translational regulation of ornithine decarboxylase and other enzymes of the polyamine pathway. Int. J. Biochem. Cell Biol. 1999, 31, 107–122. [Google Scholar] [CrossRef]
- Pyronnet, S.; Pradayrol, L.; Sonenberg, N. A cell cycle-dependent internal ribosome entry site. Mol. Cell 2000, 5, 607–616. [Google Scholar]
- Pegg, A.E.; Feith, D.J.; Fong, L.Y.; Coleman, C.S.; O’Brien, T.G.; Shantz, L.M. Transgenic mouse models for studies of the role of polyamines in normal, hypertrophic and neoplastic growth. Biochem. Soc. Trans. 2003, 31, 356–360. [Google Scholar] [CrossRef]
- Hayes, C.S.; DeFeo, K.; Lan, L.; Paul, B.; Sell, C.; Gilmour, S.K. Elevated levels of ornithine decarboxylase cooperate with Raf/ERK activation to convert normal keratinocytes into invasive malignant cells. Oncogene 2006, 25, 1543–1553. [Google Scholar] [CrossRef]
- George, K.; Iacobucci, A.; Uitto, J.; O’Brien, T.G. Identification of an X-linked locus modifying mouse skin tumor susceptibility. Mol. Carcinog. 2005, 44, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Cleveland, J.L.; O’Brien, T.G. Haploinsufficiency for odc modifies mouse skin tumor susceptibility. Cancer Res. 2005, 65, 1146–1149. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Kim, A.L.; Feith, D.J.; Pegg, A.E.; Russo, J.; Zhang, H.; Aszterbaum, M.; Kopelovich, L.; Epstein, E.H., Jr.; Bickers, D.R.; et al. Ornithine decarboxylase is a target for chemoprevention of basal and squamous cell carcinomas in Ptch1+/- mice. J. Clin. Investig. 2004, 113, 867–875. [Google Scholar] [CrossRef]
- Nilsson, J.A.; Keller, U.B.; Baudino, T.A.; Yang, C.; Norton, S.; Old, J.A.; Nilsson, L.M.; Neale, G.; Kramer, D.L.; Porter, C.W.; et al. Targeting ornithine decarboxylase in Myc-induced lymphomagenesis prevents tumor formation. Cancer Cell 2005, 7, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Gerner, E.W.; Meyskens, F.L., Jr. Polyamines and cancer: Old molecules, new understanding. Nat. Rev. Cancer 2004, 4, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Zabala-Letona, A.; Arruabarrena-Aristorena, A.; Martin-Martin, N.; Fernandez-Ruiz, S.; Sutherland, J.D.; Clasquin, M.; Tomas-Cortazar, J.; Jimenez, J.; Torres, I.; Quang, P.; et al. mTORC1-dependent AMD1 regulation sustains polyamine metabolism in prostate cancer. Nature 2017, 547, 109–113. [Google Scholar] [CrossRef]
- Shirahata, A.; Takahashi, N.; Beppu, T.; Hosoda, H.; Samejima, K. Effects of inhibitors of spermidine synthase and spermine synthase on polyamine synthesis in rat tissues. Biochem. Pharmacol. 1993, 45, 1897–1903. [Google Scholar] [CrossRef]
- Koomoa, D.L.; Yco, L.P.; Borsics, T.; Wallick, C.J.; Bachmann, A.S. Ornithine decarboxylase inhibition by alpha-difluoromethylornithine activates opposing signaling pathways via phosphorylation of both Akt/protein kinase B and p27Kip1 in neuroblastoma. Cancer Res. 2008, 68, 9825–9831. [Google Scholar] [CrossRef]
- Liao, C.; Wang, Y.; Tan, X.; Sun, L.; Liu, S. Discovery of novel inhibitors of human S-adenosylmethionine decarboxylase based on in silico high-throughput screening and a non-radioactive enzymatic assay. Sci. Rep. 2015, 5, 10754. [Google Scholar] [CrossRef]
- Alexiou, G.A.; Lianos, G.D.; Ragos, V.; Galani, V.; Kyritsis, A.P. Difluoromethylornithine in cancer: New advances. Future Oncol. 2017, 13, 809–819. [Google Scholar] [CrossRef]
- Nowotarski, S.L.; Woster, P.M.; Casero, R.A., Jr. Polyamines and cancer: Implications for chemotherapy and chemoprevention. Expert Rev. Mol. Med. 2013, 15, e3. [Google Scholar] [CrossRef] [PubMed]
- Wallace, H.M.; Fraser, A.V. Polyamine analogues as anticancer drugs. Biochem. Soc. Trans. 2003, 31, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Weeks, R.S.; Vanderwerf, S.M.; Carlson, C.L.; Burns, M.R.; O’Day, C.L.; Cai, F.; Devens, B.H.; Webb, H.K. Novel lysine-spermine conjugate inhibits polyamine transport and inhibits cell growth when given with DFMO. Exp. Cell Res. 2000, 261, 293–302. [Google Scholar] [CrossRef]
- Agostinelli, E.; Przybytkowski, E.; Averill-Bates, D.A. Glucose, glutathione, and cellular response to spermine oxidation products. Free Radic. Biol. Med. 1996, 20, 649–656. [Google Scholar] [CrossRef]
- Agostinelli, E.; Seiler, N. Non-irradiation-derived reactive oxygen species (ROS) and cancer: Therapeutic implications. Amino Acids 2006, 31, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, G.S.; Wallace, H.M. Changes in polyamine catabolism in HL-60 human promyelogenous leukaemic cells in response to etoposide-induced apoptosis. Biochem. J. 1999, 337 Pt 1, 83–87. [Google Scholar] [CrossRef]
- Agostinelli, E.; Condello, M.; Tempera, G.; Macone, A.; Bozzuto, G.; Ohkubo, S.; Calcabrini, A.; Arancia, G.; Molinari, A. The combined treatment with chloroquine and the enzymatic oxidation products of spermine overcomes multidrug resistance of melanoma M14 ADR2 cells: A new therapeutic approach. Int. J. Oncol. 2014, 45, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Di Magno, L.; Coni, S.; Di Marcotullio, L.; Canettieri, G. Digging a hole under Hedgehog: Downstream inhibition as an emerging anticancer strategy. Biochim. Biophys. Acta 2015, 1856, 62–72. [Google Scholar] [CrossRef]
- Rohatgi, R.; Milenkovic, L.; Scott, M.P. Patched1 regulates hedgehog signaling at the primary cilium. Science 2007, 317, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhang, Y.; Sun, B.; McMahon, A.P.; Wang, Y. Hedgehog Signaling: From Basic Biology to Cancer Therapy. Cell Chem. Biol. 2017, 24, 252–280. [Google Scholar] [CrossRef]
- Fujii, K.; Miyashita, T. Gorlin syndrome (nevoid basal cell carcinoma syndrome): Update and literature review. Pediatr. Int. 2014, 56, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Gailani, M.R.; Stahle-Backdahl, M.; Leffell, D.J.; Glynn, M.; Zaphiropoulos, P.G.; Pressman, C.; Unden, A.B.; Dean, M.; Brash, D.E.; Bale, A.E.; et al. The role of the human homologue of Drosophila patched in sporadic basal cell carcinomas. Nat. Genet. 1996, 14, 78–81. [Google Scholar] [CrossRef]
- Epstein, E.H. Basal cell carcinomas: Attack of the hedgehog. Nat. Rev. Cancer 2008, 8, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Amakye, D.; Jagani, Z.; Dorsch, M. Unraveling the therapeutic potential of the Hedgehog pathway in cancer. Nat. Med. 2013, 19, 1410–1422. [Google Scholar] [CrossRef] [PubMed]
- Raffel, C.; Jenkins, R.B.; Frederick, L.; Hebrink, D.; Alderete, B.; Fults, D.W.; James, C.D. Sporadic medulloblastomas contain PTCH mutations. Cancer Res. 1997, 57, 842–845. [Google Scholar] [PubMed]
- Vorechovsky, I.; Tingby, O.; Hartman, M.; Stromberg, B.; Nister, M.; Collins, V.P.; Toftgard, R. Somatic mutations in the human homologue of Drosophila patched in primitive neuroectodermal tumours. Oncogene 1997, 15, 361–366. [Google Scholar] [CrossRef]
- Kool, M.; Jones, D.T.; Jager, N.; Northcott, P.A.; Pugh, T.J.; Hovestadt, V.; Piro, R.M.; Esparza, L.A.; Markant, S.L.; Remke, M.; et al. Genome sequencing of SHH medulloblastoma predicts genotype-related response to smoothened inhibition. Cancer Cell 2014, 25, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Northcott, P.A.; Hielscher, T.; Dubuc, A.; Mack, S.; Shih, D.; Remke, M.; Al-Halabi, H.; Albrecht, S.; Jabado, N.; Eberhart, C.G.; et al. Pediatric and adult sonic hedgehog medulloblastomas are clinically and molecularly distinct. Acta Neuropathol. 2011, 122, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, X.; Parmentier, L.; King, B.; Bezrukov, F.; Kaya, G.; Zoete, V.; Seplyarskiy, V.B.; Sharpe, H.J.; McKee, T.; Letourneau, A.; et al. Genomic analysis identifies new drivers and progression pathways in skin basal cell carcinoma. Nat. Genet. 2016, 48, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Nix, N.M.; Burdine, O.; Walker, M. Vismodegib: First-in-Class Hedgehog Pathway Inhibitor for Metastatic or Locally Advanced Basal Cell Carcinoma. J. Adv. Pract. Oncol. 2014, 5, 294–296. [Google Scholar] [PubMed]
- Dijkgraaf, G.J.; Alicke, B.; Weinmann, L.; Januario, T.; West, K.; Modrusan, Z.; Burdick, D.; Goldsmith, R.; Robarge, K.; Sutherlin, D.; et al. Small molecule inhibition of GDC-0449 refractory smoothened mutants and downstream mechanisms of drug resistance. Cancer Res. 2011, 71, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kawagoe, R.; Sasai, K.; Li, Y.; Russell, H.R.; Curran, T.; McKinnon, P.J. Loss of suppressor-of-fused function promotes tumorigenesis. Oncogene 2007, 26, 6442–6447. [Google Scholar] [CrossRef] [PubMed]
- Yauch, R.L.; Dijkgraaf, G.J.; Alicke, B.; Januario, T.; Ahn, C.P.; Holcomb, T.; Pujara, K.; Stinson, J.; Callahan, C.A.; Tang, T.; et al. Smoothened mutation confers resistance to a Hedgehog pathway inhibitor in medulloblastoma. Science 2009, 326, 572–574. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, H.J.; Pau, G.; Dijkgraaf, G.J.; Basset-Seguin, N.; Modrusan, Z.; Januario, T.; Tsui, V.; Durham, A.B.; Dlugosz, A.A.; Haverty, P.M.; et al. Genomic analysis of smoothened inhibitor resistance in basal cell carcinoma. Cancer Cell 2015, 27, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Atwood, S.X.; Sarin, K.Y.; Whitson, R.J.; Li, J.R.; Kim, G.; Rezaee, M.; Ally, M.S.; Kim, J.; Yao, C.; Chang, A.L.; et al. Smoothened variants explain the majority of drug resistance in basal cell carcinoma. Cancer Cell 2015, 27, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Infante, P.; Alfonsi, R.; Botta, B.; Mori, M.; Di Marcotullio, L. Targeting GLI factors to inhibit the Hedgehog pathway. Trends Pharmacol. Sci. 2015, 36, 547–558. [Google Scholar] [CrossRef]
- Canettieri, G.; Di Marcotullio, L.; Greco, A.; Coni, S.; Antonucci, L.; Infante, P.; Pietrosanti, L.; De Smaele, E.; Ferretti, E.; Miele, E.; et al. Histone deacetylase and Cullin3-REN(KCTD11) ubiquitin ligase interplay regulates Hedgehog signalling through Gli acetylation. Nat. Cell Biol. 2010, 12, 132–142. [Google Scholar] [CrossRef]
- Coni, S.; Antonucci, L.; D’Amico, D.; Di Magno, L.; Infante, P.; De Smaele, E.; Giannini, G.; Di Marcotullio, L.; Screpanti, I.; Gulino, A.; et al. Gli2 acetylation at lysine 757 regulates hedgehog-dependent transcriptional output by preventing its promoter occupancy. PLoS ONE 2013, 8, e65718. [Google Scholar] [CrossRef]
- Canettieri, G.; Di Marcotullio, L.; Coni, S.; Greco, A.; Gulino, A. Turning off the switch in medulloblastoma: The inhibitory acetylation of an oncogene. Cell Cycle 2010, 9, 2047–2048. [Google Scholar] [CrossRef]
- Coni, S.; Mancuso, A.B.; Di Magno, L.; Sdruscia, G.; Manni, S.; Serrao, S.M.; Rotili, D.; Spiombi, E.; Bufalieri, F.; Petroni, M.; et al. Selective targeting of HDAC1/2 elicits anticancer effects through Gli1 acetylation in preclinical models of SHH Medulloblastoma. Sci. Rep. 2017, 7, 44079. [Google Scholar] [CrossRef]
- Gruber, W.; Peer, E.; Elmer, D.P.; Sternberg, C.; Tesanovic, S.; Del Burgo, P.; Coni, S.; Canettieri, G.; Neureiter, D.; Bartz, R.; et al. Targeting class I histone deacetylases by the novel small molecule inhibitor 4SC-202 blocks oncogenic hedgehog-GLI signaling and overcomes smoothened inhibitor resistance. Int. J. Cancer 2018, 142, 968–975. [Google Scholar] [CrossRef] [PubMed]
- Mirza, A.N.; Fry, M.A.; Urman, N.M.; Atwood, S.X.; Roffey, J.; Ott, G.R.; Chen, B.; Lee, A.; Brown, A.S.; Aasi, S.Z.; et al. Combined inhibition of atypical PKC and histone deacetylase 1 is cooperative in basal cell carcinoma treatment. JCI Insight 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Gulino, A.; Di Marcotullio, L.; Canettieri, G.; De Smaele, E.; Screpanti, I. Hedgehog/Gli control by ubiquitination/acetylation interplay. Vitam. Horm. 2012, 88, 211–227. [Google Scholar] [CrossRef] [PubMed]
- Di Magno, L.; Basile, A.; Coni, S.; Manni, S.; Sdruscia, G.; D’Amico, D.; Antonucci, L.; Infante, P.; De Smaele, E.; Cucchi, D.; et al. The energy sensor AMPK regulates Hedgehog signaling in human cells through a unique Gli1 metabolic checkpoint. Oncotarget 2016, 7, 9539–9550. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Luo, J.; Mosley, Y.C.; Hedrick, V.E.; Paul, L.N.; Chang, J.; Zhang, G.; Wang, Y.K.; Banko, M.R.; Brunet, A.; et al. AMP-Activated Protein Kinase Directly Phosphorylates and Destabilizes Hedgehog Pathway Transcription Factor GLI1 in Medulloblastoma. Cell Rep. 2015. [Google Scholar] [CrossRef]
- Po, A.; Silvano, M.; Miele, E.; Capalbo, C.; Eramo, A.; Salvati, V.; Todaro, M.; Besharat, Z.M.; Catanzaro, G.; Cucchi, D.; et al. Noncanonical GLI1 signaling promotes stemness features and in vivo growth in lung adenocarcinoma. Oncogene 2017, 36, 4641–4652. [Google Scholar] [CrossRef] [PubMed]
- Antonucci, L.; Di Magno, L.; D’Amico, D.; Manni, S.; Serrao, S.M.; Di Pastena, F.; Bordone, R.; Yurtsever, Z.N.; Caimano, M.; Petroni, M.; et al. Mitogen-activated kinase kinase kinase 1 inhibits hedgehog signaling and medulloblastoma growth through GLI1 phosphorylation. Int. J. Oncol. 2019, 54, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zhang, H.; Park, S.S.; Venneti, S.; Kuick, R.; Ha, K.; Michael, L.E.; Santi, M.; Uchida, C.; Uchida, T.; et al. Loss of Pin1 Suppresses Hedgehog-Driven Medulloblastoma Tumorigenesis. Neoplasia 2017, 19, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Di Magno, L.; Manzi, D.; D’Amico, D.; Coni, S.; Macone, A.; Infante, P.; Di Marcotullio, L.; De Smaele, E.; Ferretti, E.; Screpanti, I.; et al. Druggable glycolytic requirement for Hedgehog-dependent neuronal and medulloblastoma growth. Cell Cycle 2014, 13, 3404–3413. [Google Scholar] [CrossRef] [PubMed]
- Rubin, A.I.; Chen, E.H.; Ratner, D. Basal-cell carcinoma. N. Engl. J. Med. 2005, 353, 2262–2269. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.T.; Kempton, S.J.; Rao, V.K. The Economics of Skin Cancer: An Analysis of Medicare Payment Data. Plast. Reconstr. Surg. Glob. Open 2016, 4, e868. [Google Scholar] [CrossRef] [PubMed]
- von Domarus, H.; Stevens, P.J. Metastatic basal cell carcinoma. Report of five cases and review of 170 cases in the literature. J. Am. Acad. Dermatol. 1984, 10, 1043–1060. [Google Scholar] [CrossRef]
- Leiter, U.; Eigentler, T.; Garbe, C. Epidemiology of skin cancer. Adv. Exp. Med. Biol. 2014, 810, 120–140. [Google Scholar] [PubMed]
- Mohan, S.V.; Chang, A.L. Advanced Basal Cell Carcinoma: Epidemiology and Therapeutic Innovations. Curr. Dermatol. Rep. 2014, 3, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Aszterbaum, M.; Rothman, A.; Johnson, R.L.; Fisher, M.; Xie, J.; Bonifas, J.M.; Zhang, X.; Scott, M.P.; Epstein, E.H., Jr. Identification of mutations in the human PATCHED gene in sporadic basal cell carcinomas and in patients with the basal cell nevus syndrome. J. Investig. Dermatol. 1998, 110, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Murone, M.; Luoh, S.M.; Ryan, A.; Gu, Q.; Zhang, C.; Bonifas, J.M.; Lam, C.W.; Hynes, M.; Goddard, A.; et al. Activating Smoothened mutations in sporadic basal-cell carcinoma. Nature 1998, 391, 90–92. [Google Scholar] [CrossRef]
- Sekulic, A.; Migden, M.R.; Oro, A.E.; Dirix, L.; Lewis, K.D.; Hainsworth, J.D.; Solomon, J.A.; Yoo, S.; Arron, S.T.; Friedlander, P.A.; et al. Efficacy and safety of vismodegib in advanced basal-cell carcinoma. N. Engl. J. Med. 2012, 366, 2171–2179. [Google Scholar] [CrossRef]
- Basset-Seguin, N.; Sharpe, H.J.; de Sauvage, F.J. Efficacy of Hedgehog pathway inhibitors in Basal cell carcinoma. Mol. Cancer Ther. 2015, 14, 633–641. [Google Scholar] [CrossRef]
- Chang, A.L.; Oro, A.E. Initial assessment of tumor regrowth after vismodegib in advanced Basal cell carcinoma. Arch. Dermatol. 2012, 148, 1324–1325. [Google Scholar] [CrossRef]
- Scalabrino, G.; Pigatto, P.; Ferioli, M.E.; Modena, D.; Puerari, M.; Caru, A. Levels of activity of the polyamine biosynthetic decarboxylases as indicators of degree of malignancy of human cutaneous epitheliomas. J. Investig. Dermatol. 1980, 74, 122–124. [Google Scholar] [CrossRef]
- O’Brien, T.G.; Simsiman, R.C.; Boutwell, R.K. Induction of the polyamine-biosynthetic enzymes in mouse epidermis by tumor-promoting agents. Cancer Res. 1975, 35, 1662–1670. [Google Scholar] [PubMed]
- Verma, A.K.; Lowe, N.J.; Boutwell, R.K. Induction of mouse epidermal ornithine decarboxylase activity and DNA synthesis by ultraviolet light. Cancer Res. 1979, 39, 1035–1040. [Google Scholar] [PubMed]
- Verma, A.K.; Shapas, B.G.; Rice, H.M.; Boutwell, R.K. Correlation of the inhibition by retinoids of tumor promoter-induced mouse epidermal ornithine decarboxylase activity and of skin tumor promotion. Cancer Res. 1979, 39, 419–425. [Google Scholar] [PubMed]
- Elmets, C.A.; Athar, M. Targeting ornithine decarboxylase for the prevention of nonmelanoma skin cancer in humans. Cancer Prev. Res. 2010, 3, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Kreul, S.M.; Havighurst, T.; Kim, K.; Mendonca, E.A.; Wood, G.S.; Snow, S.; Borich, A.; Verma, A.; Bailey, H.H. A phase III skin cancer chemoprevention study of DFMO: Long-term follow-up of skin cancer events and toxicity. Cancer Prev. Res. 2012, 5, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- Khanna, V.; Achey, R.L.; Ostrom, Q.T.; Block-Beach, H.; Kruchko, C.; Barnholtz-Sloan, J.S.; de Blank, P.M. Incidence and survival trends for medulloblastomas in the United States from 2001 to 2013. J. Neurooncol. 2017, 135, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Zeltzer, P.M.; Boyett, J.M.; Finlay, J.L.; Albright, A.L.; Rorke, L.B.; Milstein, J.M.; Allen, J.C.; Stevens, K.R.; Stanley, P.; Li, H.; et al. Metastasis stage, adjuvant treatment, and residual tumor are prognostic factors for medulloblastoma in children: Conclusions from the Children’s Cancer Group 921 randomized phase III study. J. Clin. Oncol. 1999, 17, 832–845. [Google Scholar] [CrossRef]
- Thomas, P.R.; Deutsch, M.; Kepner, J.L.; Boyett, J.M.; Krischer, J.; Aronin, P.; Albright, L.; Allen, J.C.; Packer, R.J.; Linggood, R.; et al. Low-stage medulloblastoma: Final analysis of trial comparing standard-dose with reduced-dose neuraxis irradiation. J. Clin. Oncol. 2000, 18, 3004–3011. [Google Scholar] [CrossRef]
- Wang, J.; Garancher, A.; Ramaswamy, V.; Wechsler-Reya, R.J. Medulloblastoma: From Molecular Subgroups to Molecular Targeted Therapies. Annu. Rev. Neurosci. 2018, 41, 207–232. [Google Scholar] [CrossRef]
- Cox, M.C.; Kusters, J.M.; Gidding, C.E.; Schieving, J.H.; van Lindert, E.J.; Kaanders, J.H.; Janssens, G.O. Acute toxicity profile of craniospinal irradiation with intensity-modulated radiation therapy in children with medulloblastoma: A prospective analysis. Radiat. Oncol. 2015, 10, 241. [Google Scholar] [CrossRef]
- Martin, A.M.; Raabe, E.; Eberhart, C.; Cohen, K.J. Management of pediatric and adult patients with medulloblastoma. Curr. Treat. Options Oncol. 2014, 15, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Schwalbe, E.C.; Lindsey, J.C.; Nakjang, S.; Crosier, S.; Smith, A.J.; Hicks, D.; Rafiee, G.; Hill, R.M.; Iliasova, A.; Stone, T.; et al. Novel molecular subgroups for clinical classification and outcome prediction in childhood medulloblastoma: A cohort study. Lancet Oncol. 2017, 18, 958–971. [Google Scholar] [CrossRef]
- Taylor, M.D.; Northcott, P.A.; Korshunov, A.; Remke, M.; Cho, Y.J.; Clifford, S.C.; Eberhart, C.G.; Parsons, D.W.; Rutkowski, S.; Gajjar, A.; et al. Molecular subgroups of medulloblastoma: The current consensus. Acta Neuropathol. 2012, 123, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Northcott, P.A.; Buchhalter, I.; Morrissy, A.S.; Hovestadt, V.; Weischenfeldt, J.; Ehrenberger, T.; Grobner, S.; Segura-Wang, M.; Zichner, T.; Rudneva, V.A.; et al. The whole-genome landscape of medulloblastoma subtypes. Nature 2017, 547, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.W.; Orr, B.A.; Wu, G.; Gururangan, S.; Lin, T.; Qaddoumi, I.; Packer, R.J.; Goldman, S.; Prados, M.D.; Desjardins, A.; et al. Vismodegib Exerts Targeted Efficacy Against Recurrent Sonic Hedgehog-Subgroup Medulloblastoma: Results From Phase II Pediatric Brain Tumor Consortium Studies PBTC-025B and PBTC-032. J. Clin. Oncol. 2015, 33, 2646–2654. [Google Scholar] [CrossRef]
- Scalabrino, G.; Modena, D.; Ferioli, M.E.; Puerari, M.; Luccarelli, G. Degrees of malignancy in human primary central nervous system tumors: Ornithine decarboxylase levels as better indicators than adenosylmethionine decarboxylase levels. J. Natl. Cancer Inst. 1982, 68, 751–754. [Google Scholar]
- D’Amico, D.; Antonucci, L.; Di Magno, L.; Coni, S.; Sdruscia, G.; Macone, A.; Miele, E.; Infante, P.; Di Marcotullio, L.; De Smaele, E.; et al. Non-canonical Hedgehog/AMPK-Mediated Control of Polyamine Metabolism Supports Neuronal and Medulloblastoma Cell Growth. Dev. Cell 2015, 35, 21–35. [Google Scholar] [CrossRef]
- D’Amico, D.; Canettieri, G. Translating Hedgehog in Cancer: Controlling Protein Synthesis. Trends Mol. Med. 2016, 22, 851–862. [Google Scholar] [CrossRef]
- Zhang, H.; Kuick, R.; Park, S.S.; Peabody, C.; Yoon, J.; Fernandez, E.C.; Wang, J.; Thomas, D.; Viollet, B.; Inoki, K.; et al. Loss of AMPKalpha2 Impairs Hedgehog-Driven Medulloblastoma Tumorigenesis. Int. J. Mol. Sci. 2018, 19, 3287. [Google Scholar] [CrossRef]
- Zhang, R.; Huang, S.Y.; Ka-Wai Li, K.; Li, Y.H.; Hsu, W.H.; Zhang, G.J.; Chang, C.J.; Yang, J.Y. Dual degradation signals destruct GLI1: AMPK inhibits GLI1 through beta-TrCP-mediated proteasome degradation. Oncotarget 2017, 8, 49869–49881. [Google Scholar] [CrossRef]
- Faubert, B.; Vincent, E.E.; Griss, T.; Samborska, B.; Izreig, S.; Svensson, R.U.; Mamer, O.A.; Avizonis, D.; Shackelford, D.B.; Shaw, R.J.; et al. Loss of the tumor suppressor LKB1 promotes metabolic reprogramming of cancer cells via HIF-1alpha. Proc. Natl. Acad. Sci. USA 2014, 111, 2554–2559. [Google Scholar] [CrossRef] [PubMed]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).