Natural Sulfur-Containing Compounds: An Alternative Therapeutic Strategy against Liver Fibrosis

 ,
,  ,
, 
Abstract
1. Introduction
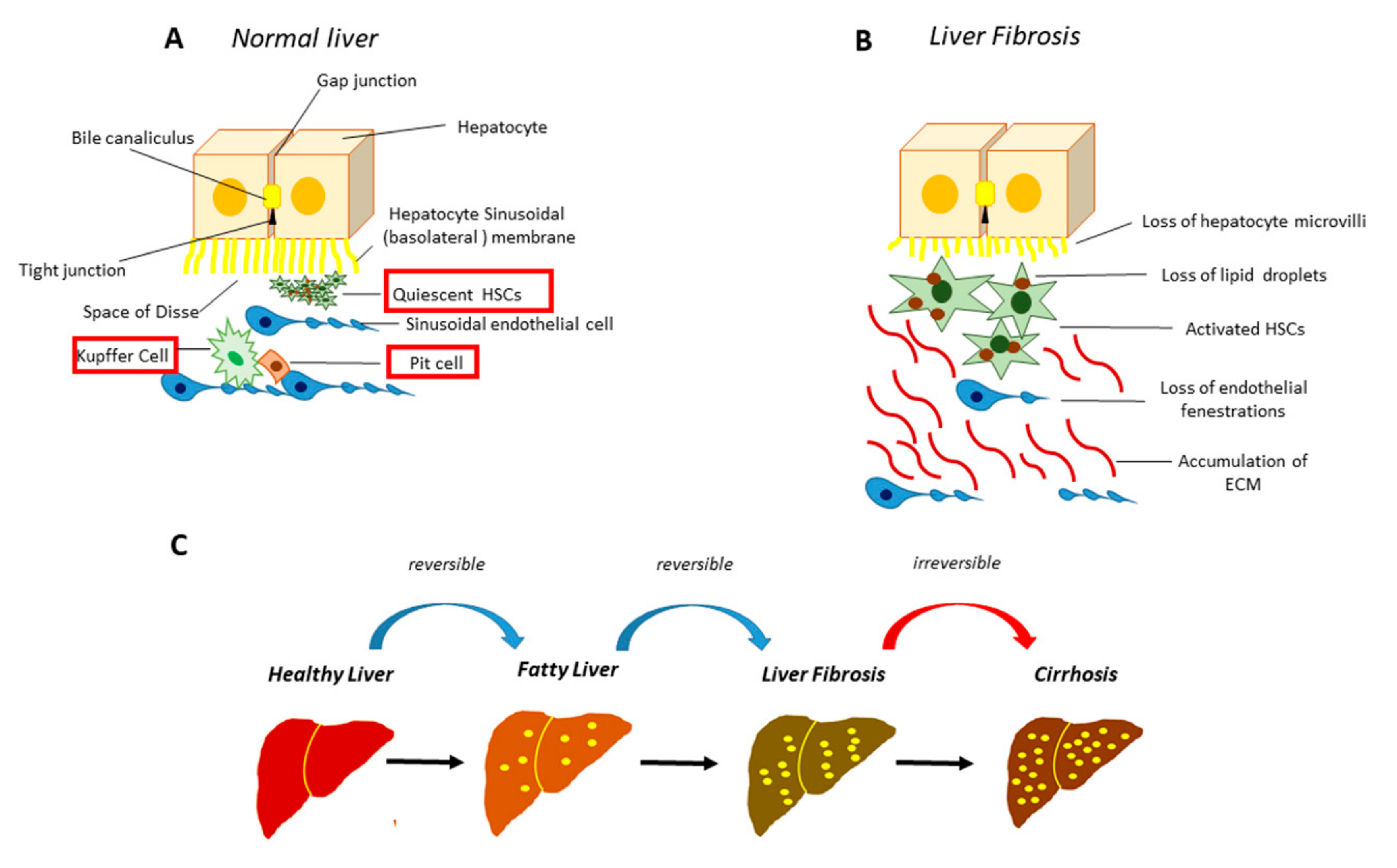
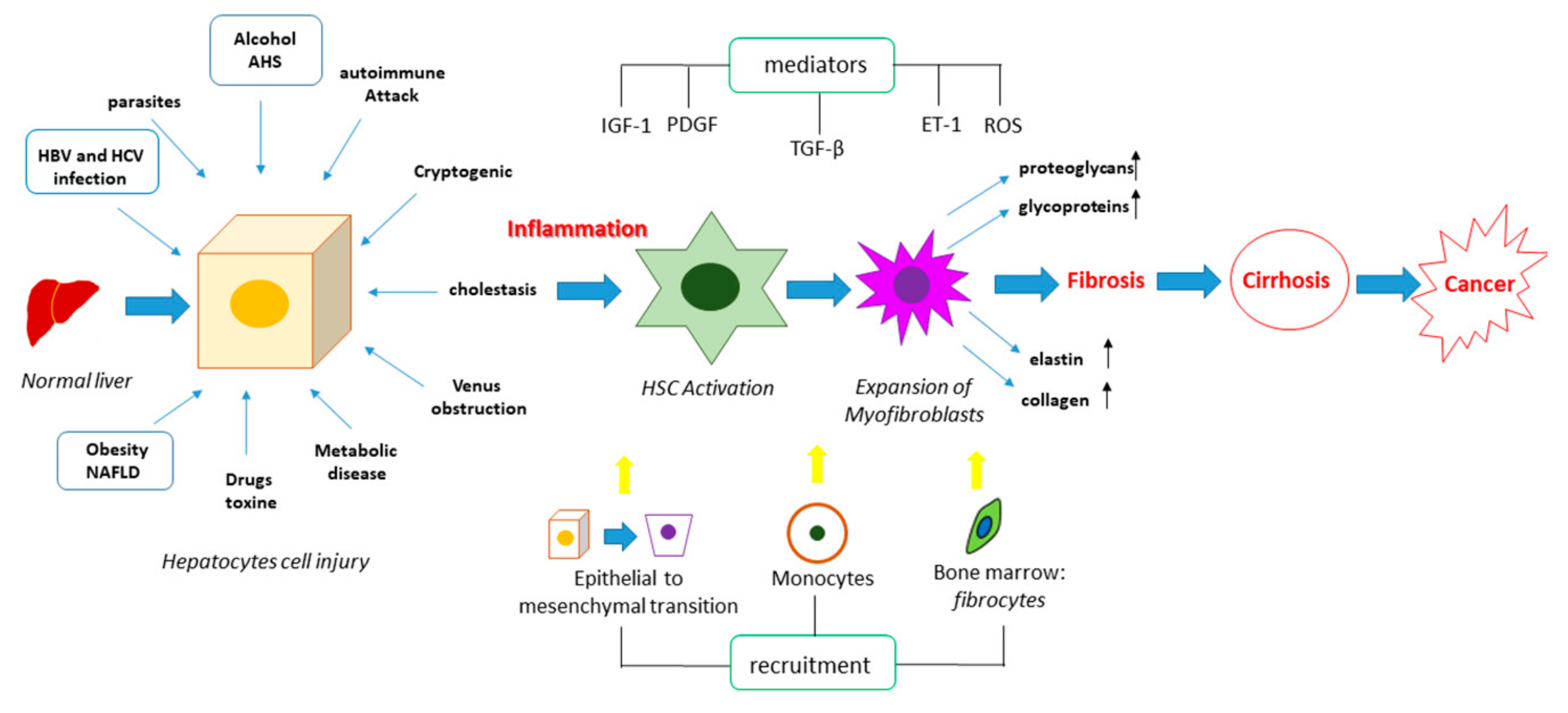
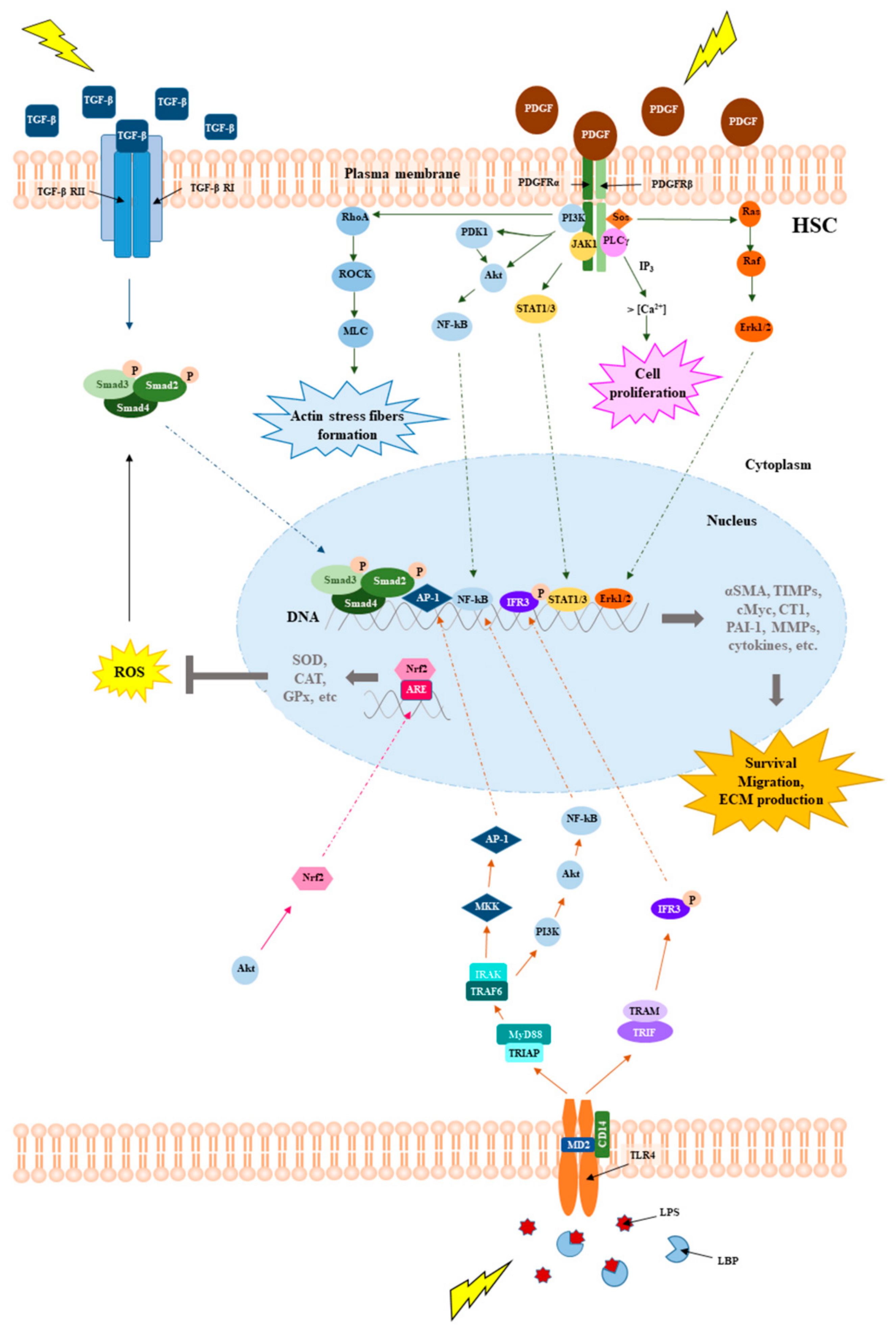
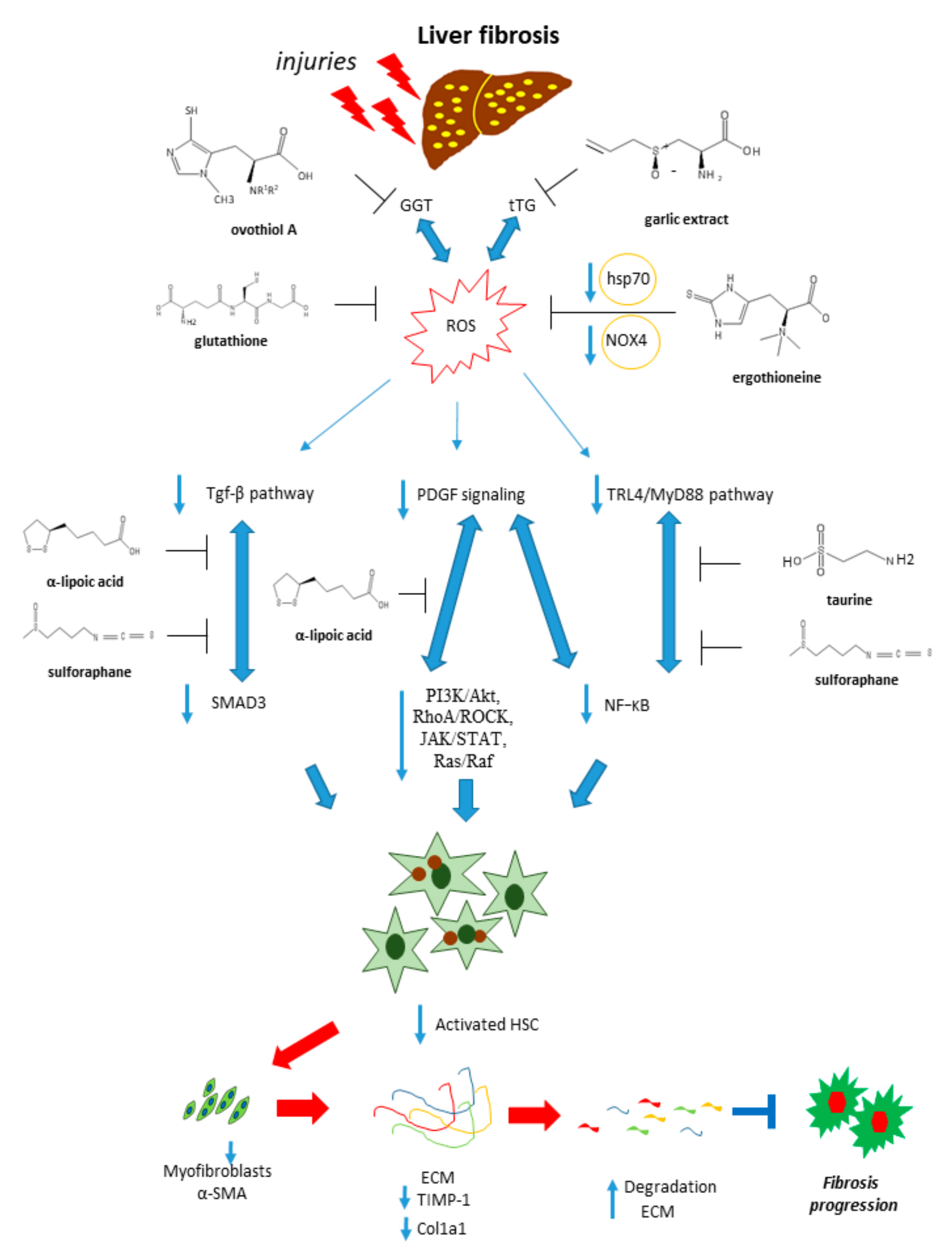
2. Molecular Mechanisms Underpinning Liver Fibrosis
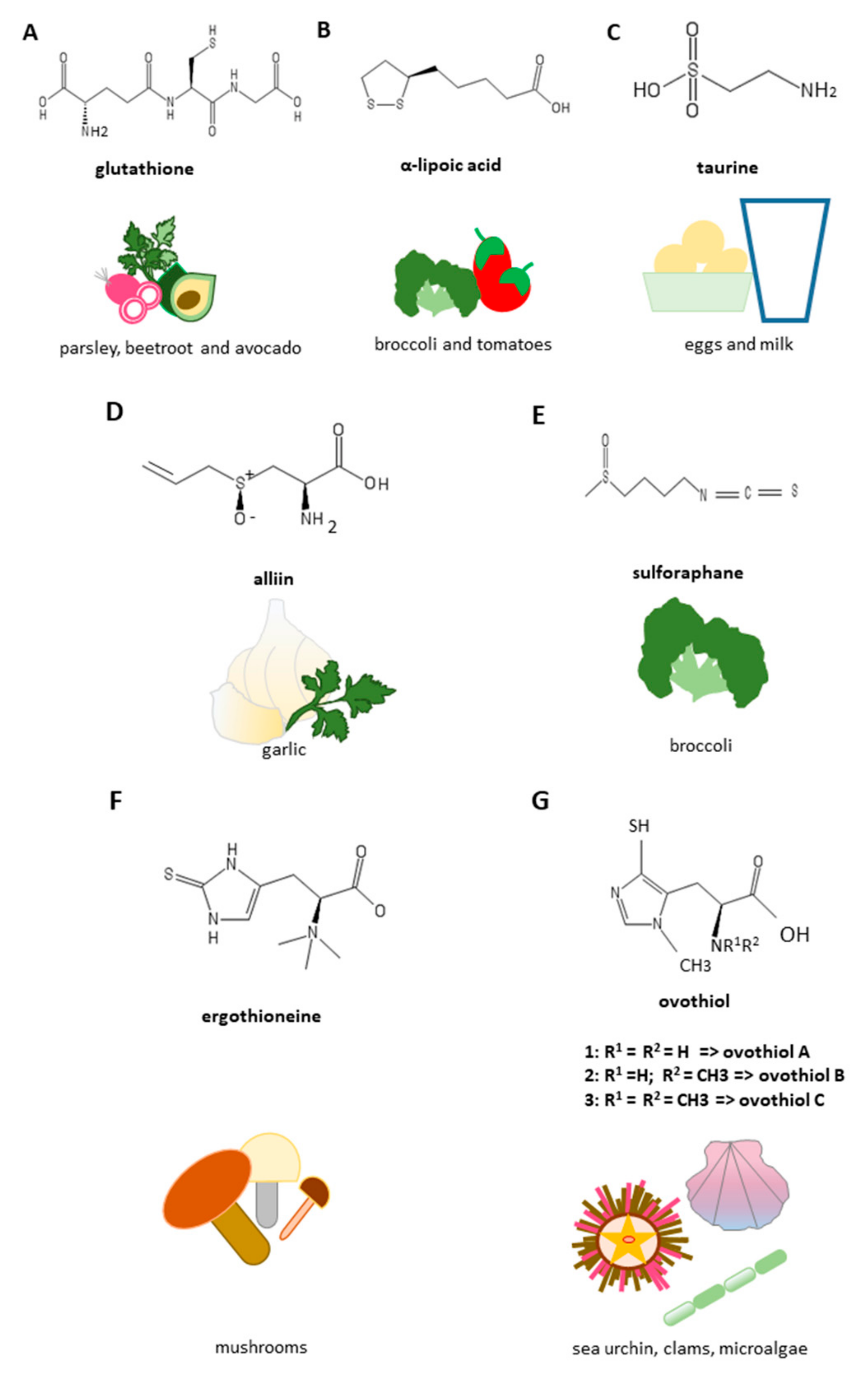
3. Natural Sulfur-Containing Compounds for the Treatment of Liver Fibrosis
3.1. Glutathione
3.2. Lipoic Acid
3.3. Taurine
3.4. Garlic Derived Sulfur Compounds
3.5. Sulforaphane
3.6. Sulfur-Containing Histidines
4. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Akt | Protein Kinase |
| ALA | α-lipoic acid |
| ALT | Alanine transaminase |
| AP-1 | Activator protein-1 |
| AST | Aspartate transaminase |
| BDL | Bile duct ligation |
| cMyc | Cellular myelocytomatosis |
| CPS | Capsaicin |
| DADS | Diallyl disulphide |
| DAS | Diallyl sulphide |
| ECM | Extracellular matrix |
| ERG | Ergothioneine |
| Erk1/2 | Extracellular signal-regulated kinases 1 and 2 |
| ET-1 | Endothelin-1 |
| GGT | γ-glutamyl transpeptidase |
| GPx | Glutathione peroxidase |
| GSH | Glutathione |
| GST | Glutathione S-transferases |
| H2S | Hydrogen sulfide |
| HSC | Hepatic stellate cell |
| Hsp | Heat shock protein |
| IFR3 | Interferon regulatory factor 3 |
| IGF-1 | Insulin-like growth factor 1 |
| IL-6 | Interleukin-6 |
| iNOS | Inducible nitric oxide synthase |
| LBP | LPS-binding protein |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-activated protein kinase |
| MKK | Mitogen-activated protein kinase kinase |
| MMPs | Matrix metalloproteinases |
| mTOR | Mammalian target of rapamycin |
| MyD88 | Myeloid differentiation primary response protein |
| NAC | N-acetyl-L-cysteine |
| NAFLD | Non-alcoholic liver disease |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NOX | NADPH oxidase |
| Nrf-2 | Nuclear factor erythroid 2-related factor 2 |
| OCTN1 | Organic cation transporter 1 |
| PAI-1 | Plasminogen activator inhibitor-1 |
| PDGF | Platelet-derived growth factor |
| PDGFRα/β | Platelet-derived growth factor receptor isoform α/β |
| PDK1 | Phosphoinositide-dependent protein kinase 1 |
| PI3K | Phosphatidylinositol 3-kinase |
| PLCγ | Phospholipase C γ |
| PPARγ | Peroxisome proliferator-activated receptor γ |
| Raf | Rapidly accelerated fibrosarcoma |
| RhoA | Ras homolog gene family, member A |
| ROCK | Rho-associated protein kinase |
| ROS | reactive oxygen species |
| SAC | S-allylcysteine |
| SAG | S-allyl-glutathione |
| SAM | S-adenosyl-L-methionine |
| SECs | Sinusoidal endothelial cells |
| SFN | Sulforaphane |
| SIRT1 | Silent information regulator 1 |
| SMAD2/3/4 | Small mother against decapentaplegic protein 2/3/4 |
| SNAC | S-Nitroso-N-acetylcysteine |
| SOD | Superoxide dismutase |
| Sp1 | Specificity protein 1 |
| STAT1/3 | Signal Transducers of Activated Transcription 1/3 |
| TGF-β | Transforming growth factor beta |
| TGF-β RI/II | Transforming growth factor beta receptor type I/II |
| TIMPs | Tissue inhibitors of metalloproteinases |
| TIRAP | TIR-domain containing adapter protein |
| TLR4 | Toll-like receptor 4 |
| TRAF6 | Tumor necrosis factor receptor-associated factor 6 |
| TRAM | Translocating chain-associating membrane protein |
| TRIF | TIR-domain-containing adapter-inducing interferon-β |
| tTG | Tissue transglutaminase |
| Wnt | Wingless/integrated |
| αSMA | α-smooth muscle actin |
References
- Bedossa, P.; Paradis, V. Liver extracellular matrix in health and disease. J. Pathol. 2003, 200, 504–515. [Google Scholar] [CrossRef]
- Jiang, C.; Zhou, Q.; Jin, M.; Niu, J.; Gao, Y. Congenital hepatic fibrosis with polycystic kidney disease: Two case reports. Medicine 2019, 98, e15600. [Google Scholar] [CrossRef] [PubMed]
- Povero, D.; Busletta, C.; Novo, E.; Di Bonzo, L.V.; Cannito, S.; Paternostro, C.; Parola, M. Liver fibrosis: A dynamic and potentially reversible process. Histol. Histopathol. 2010, 25, 1075–1091. [Google Scholar] [PubMed]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Pinzani, M.; Rombouts, K.; Colagrande, S. Fibrosis in chronic liver diseases: Diagnosis and management. J. Hepatol. 2005, 42, S22–S36. [Google Scholar] [CrossRef]
- Pinter, M.; Trauner, M.; Peck-Radosavljevic, M.; Sieghart, W. Cancer and liver cirrhosis: Implications on prognosis and management. ESMO Open 2016, 1, e000042. [Google Scholar] [CrossRef]
- Weiskirchen, R.; Tacke, F. Liver Fibrosis: From Pathogenesis to Novel Therapies. Dig. Dis. 2016, 34, 410–422. [Google Scholar] [CrossRef]
- Pinzani, M. Pathophysiology of Liver Fibrosis. Dig. Dis. 2015, 33, 492–497. [Google Scholar] [CrossRef]
- Elpek, G.Ö. Cellular and molecular mechanisms in the pathogenesis of liver fibrosis: An update. World J. Gastroenterol. 2014, 20, 7260–7276. [Google Scholar] [CrossRef]
- Arauz, J.; Ramos-Tovar, E.; Muriel, P. Redox state and methods to evaluate oxidative stress in liver damage: From bench to bedside. Ann. Hepatol. 2016, 15, 160–173. [Google Scholar]
- Safadi, R.; Friedman, S.L. Hepatic fibrosis-role of hepatic stellate cell activation. Med. Gen. Med. 2002, 4, 27. [Google Scholar]
- Zhang, C.-Y.; Yuan, W.-G.; He, P.; Lei, J.-H.; Wang, C.-X. Liver fibrosis and hepatic stellate cells: Etiology, pathological hallmarks and therapeutic targets. World J. Gastroenterol. 2016, 22, 10512–10522. [Google Scholar] [CrossRef] [PubMed]
- Sacco, R.; Eggenhoffner, R.; Giacomelli, L. Glutathione in the treatment of liver diseases: Insights from clinical practice. Minerva Gastroenterol. Dietol. 2016, 62, 316–324. [Google Scholar] [PubMed]
- Castellano, I.; Merlino, A. γ-Glutamyltranspeptidases: Sequence, structure, biochemical properties, and biotechnological applications. Cell Mol. Life Sci. 2012, 69, 3381–3394. [Google Scholar] [CrossRef] [PubMed]
- Castellano, I.; Merlino, A. Gamma-Glutamyl Transpeptidases: Structure and Function. In Springer Briefs in Biochemistry and Molecular Biology; Springer: Basel, Switzerland, 2013. [Google Scholar]
- Aitken, A.E.; Richardson, T.A.; Morgan, E.T. Regulation of drug-metabolizing enzymes and transporters in inflammation. Ann. Rev. Pharmacol. Toxicol. 2006, 46, 123–149. [Google Scholar] [CrossRef]
- Li, H.-Y.; Ju, D.; Zhang, D.-W.; Li, H.; Kong, L.-M.; Guo, Y.; Li, C.; Wang, X.-L.; Chen, Z.-N.; Bian, H. Activation of TGF-β1-CD147 positive feedback loop in hepatic stellate cells promotes liver fibrosis. Sci. Rep. 2015, 5, 16552. [Google Scholar] [CrossRef]
- Weng, H.-L.; Liu, Y.; Chen, J.-L.; Huang, T.; Xu, L.-J.; Godoy, P.; Hu, J.-H.; Zhou, C.; Stickel, F.; Marx, A.; et al. The etiology of liver damage imparts cytokines transforming growth factor β1 or interleukin-13 as driving forces in fibrogenesis. Hepatology 2009, 50, 230–243. [Google Scholar] [CrossRef]
- Vallée, A.; Lecarpentier, Y.; Guillevin, R.; Vallée, J.N. Interactions between TGF-β1, canonical WNT/β-catenin pathway and PPAR γ in radiation-induced fibrosis. Oncotarget 2017, 8, 90579–90604. [Google Scholar] [CrossRef]
- Vallée, A.; Lecarpentier, Y.; Vallée, J.N. Thermodynamic Aspects and Reprogramming Cellular Energy Metabolism during the Fibrosis Process. Int. J. Mol. Sci. 2017, 18, 2537. [Google Scholar] [CrossRef]
- Ying, H.-Z.; Chen, Q.; Zhang, W.-Y.; Zhang, H.-H.; Ma, Y.; Zhang, S.-Z.; Fang, J.; Yu, C.-H. PDGF signaling pathway in hepatic fibrosis pathogenesis and therapeutics (Review). Mol. Med. Rep. 2017, 16, 7879–7889. [Google Scholar] [CrossRef]
- Guo, J.; Friedman, S.L. Toll-like receptor 4 signaling in liver injury and hepatic fibrogenesis. Fibrogenesis Tissue Repair 2010, 3, 21. [Google Scholar] [CrossRef] [PubMed]
- Novo, E.; Cannito, S.; Morello, E.; Paternostro, C.; Bocca, C.; Miglietta, A.; Parola, M. Hepatic myofibroblasts and fibrogenic progression of chronic liver diseases. Histol. Histopathol. 2015, 30, 1011–1032. [Google Scholar] [PubMed]
- Hemmann, S.; Graf, J.; Roderfeld, M.; Roeb, E. Expression of MMPs and TIMPs in liver fibrosis 2013; A systematic review with special emphasis on anti-fibrotic strategies. J. Hepatol. 2007, 46, 955–975. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.-Y.; Wu, J.-Y.; Wu, Y.-B.; Zhong, M.-Z.; Lu, H.-M. Effects of endothelin-1 on hepatic stellate cell proliferation, collagen synthesis and secretion, intracellular free calcium concentration. World J. Gastroenterol. 2004, 10, 2697–2700. [Google Scholar] [CrossRef]
- Adamek, A.; Kasprzak, A. Insulin-Like Growth Factor (IGF) System in Liver Diseases. Int. J. Mol. Sci. 2018, 19, 1308. [Google Scholar] [CrossRef]
- Weiskirchen, R.; Weiskirchen, S.; Tacke, F. Recent advances in understanding liver fibrosis: Bridging basic science and individualized treatment concepts. F1000Research 2018, 7. [Google Scholar] [CrossRef]
- Friedman, S.L.; Sheppard, D.; Duffield, J.S.; Violette, S. Therapy for Fibrotic Diseases: Nearing the Starting Line. Sci. Transl. Med. 2013, 5, sr1–sr167. [Google Scholar] [CrossRef]
- Murphy, F.; Arthur, M.; Iredale, J. Developing strategies for liver fibrosis treatment. Expert Opin. Investig. Drug 2002, 11, 1575–1585. [Google Scholar] [CrossRef]
- Weiskirchen, R. Hepatoprotective and Anti-fibrotic Agents: It’s Time to Take the Next Step. Front. Pharmacol. 2016, 6, 303. [Google Scholar] [CrossRef]
- Nair, D.G.; Weiskirchen, R.; Al-Musharafi, S.K. The use of marine-derived bioactive compounds as potential hepatoprotective agents. Acta Pharmacol. Sin. 2015, 36, 158–170. [Google Scholar] [CrossRef]
- D’Argenio, G.; Amoruso, D.C.; Mazzone, G.; Vitaglione, P.; Romano, A.; Ribecco, M.T.; D’Armiento, M.R.; Mezza, E.; Morisco, F.; Fogliano, V.; et al. Garlic extract prevents CCl4-induced liver fibrosis in rats: The role of tissue transglutaminase. Dig. Liver Dis. 2010, 42, 571–577. [Google Scholar] [CrossRef] [PubMed]
- D’Argenio, G.; Mazzone, G.; Ribecco, M.T.; Lembo, V.; Vitaglione, P.; Guarino, M.; Morisco, F.; Napolitano, M.; Fogliano, V.; Caporaso, N. Garlic extract attenuating rat liver fibrosis by inhibiting TGF-β1. Clin. Nutr. 2013, 32, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.J.; Kim, J.-Y.; Min, A.-K.; Park, K.-G.; Harris, R.A.; Kim, H.-J.; Lee, I.-K. Sulforaphane attenuates hepatic fibrosis via NF-E2-related factor 2-mediated inhibition of transforming growth factor-β/Smad signaling. Free Radic. Biol. Med. 2012, 52, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Vitaglione, P.; Morisco, F.; Mazzone, G.; Amoruso, D.C.; Ribecco, M.T.; Romano, A.; Fogliano, V.; Caporaso, N.; D’Argenio, G. Coffee reduces liver damage in a rat model of steatohepatitis: The underlying mechanisms and the role of polyphenols and melanoidins. Hepatology 2010, 52, 1652–1661. [Google Scholar] [CrossRef] [PubMed]
- Shinkawa, H.; Takemura, S.; Minamiyama, Y.; Kodai, S.; Tsukioka, T.; Osada-Oka, M.; Kubo, S.; Okada, S.; Suehiro, S. S-allylcysteine is effective as a chemopreventive agent against porcine serum-induced hepatic fibrosis in rats. Osaka City Med. J. 2009, 55, 61–69. [Google Scholar]
- Shin, M.-O.; Moon, J.-O. Effect of dietary supplementation of grape skin and seeds on liver fibrosis induced by dimethylnitrosamine in rats. Nutr. Res. Pract. 2010, 4, 369–374. [Google Scholar] [CrossRef][Green Version]
- Brancaccio, M.; D’Argenio, G.; Lembo, V.; Palumbo, A.; Castellano, I. Antifibrotic Effect of Marine Ovothiol in an In Vivo Model of Liver Fibrosis. Oxid. Med. Cell. Longev. 2018, 2018, 5045734. [Google Scholar] [CrossRef]
- Umbreen, S.; Lubega, J.; Loake, G.J. Sulphur: The heart of nitric oxide-dependent redox signalling. J. Exp. Bot. 2019, 70, 4279–4286. [Google Scholar] [CrossRef]
- Iwakiri, Y.; Kim, M.Y. Nitric oxide in liver diseases. Trends Pharmacol. Sci. 2015, 36, 524–536. [Google Scholar] [CrossRef]
- Jacob, C. A scent of therapy: Pharmacological implications of natural products containing redox-active sulfur atoms. Nat. Prod. Rep. 2006, 23, 851–863. [Google Scholar] [CrossRef]
- Dunbar, K.L.; Scharf, D.H.; Litomska, A.; Hertweck, C. Enzymatic Carbon-Sulfur Bond Formation in Natural Product Biosynthesis. Chem. Rev. 2017, 117, 5521–5577. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.-D.; Wang, D.-Y.; Li, H.-M.; Guo, J.-C.; Duan, S.-F.; Ji, X.-Y. Hydrogen Sulfide as a Novel Regulatory Factor in Liver Health and Disease. Oxid. Med. Cell. Longev. 2019, 2019, 3831713. [Google Scholar] [CrossRef] [PubMed]
- Giustarini, D.; Colombo, G.; Garavaglia, M.L.; Astori, E.; Portinaro, N.M.; Reggiani, F.; Badalamenti, S.; Aloisi, A.M.; Santucci, A.; Rossi, R.; et al. Assessment of glutathione/glutathione disulphide ratio and S-glutathionylated proteins in human blood, solid tissues, and cultured cells. Free Radic. Biol. Med. 2017, 112, 360–375. [Google Scholar] [CrossRef] [PubMed]
- Castellano, I.; Ruocco, M.R.; Cecere, F.; Di Maro, A.; Chambery, A.; Michniewicz, A.; Parlato, G.; Masullo, M.; De Vendittis, E. Glutathionylation of the iron superoxide dismutase from the psychrophilic eubacterium Pseudoalteromonas haloplanktis. Biochim. Biophys. Acta Proteins Proteomics 2008, 1784, 816–826. [Google Scholar] [CrossRef]
- Floryszak-Wieczorek, J.; Milczarek, G.; Arasimowicz, M.; Ciszewski, A. Do nitric oxide donors mimic endogenous NO-related response in plants? Planta 2006, 224, 1363–1372. [Google Scholar] [CrossRef]
- Honda, Y.; Kessoku, T.; Sumida, Y.; Kobayashi, T.; Kato, T.; Ogawa, Y.; Tomeno, W.; Imajo, K.; Fujita, K.; Yoneda, M.; et al. Efficacy of glutathione for the treatment of nonalcoholic fatty liver disease: An open-label, single-arm, multicenter, pilot study. BMC Gastroenterol. 2017, 17, 96. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Q.; Hesketh, J.; Huang, D.; Gan, F.; Hao, S.; Tang, S.; Guo, Y.; Huang, K. Protective effects of selenium-glutathione-enriched probiotics on CCl4-induced liver fibrosis. J. Nutr. Biochem. 2018, 58, 138–149. [Google Scholar] [CrossRef]
- Durrani, A.I.; Schwartz, H.; Nagl, M.; Sontag, G. Determination of free α-lipoic acid in foodstuffs by HPLC coupled with CEAD and ESI-MS. Food Chem. 2010, 120, 1143–1148. [Google Scholar] [CrossRef]
- Foo, N.-P.; Lin, S.-H.; Lee, Y.-H.; Wu, M.-J.; Wang, Y.-J. α-Lipoic acid inhibits liver fibrosis through the attenuation of ROS-triggered signaling in hepatic stellate cells activated by PDGF and TGF-β. Toxicology 2011, 282, 39–46. [Google Scholar] [CrossRef]
- Min, A.-K.; Kim, M.-K.; Seo, H.-Y.; Kim, H.-S.; Jang, B.K.; Hwang, J.S.; Choi, H.-S.; Lee, K.-U.; Park, K.-G.; Lee, I.-K. Alpha-lipoic acid inhibits hepatic PAI-1 expression and fibrosis by inhibiting the TGF-β signaling pathway. Biochem. Biophys. Res. Commun. 2010, 393, 536–541. [Google Scholar] [CrossRef]
- Sadek, K.M.; Saleh, E.A.; Nasr, S.M. Molecular hepatoprotective effects of lipoic acid against carbon tetrachloride-induced liver fibrosis in rats: Hepatoprotection at molecular level. Hum. Exp. Toxicol. 2017, 37, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Alban, F.T.E.; Gyamfi, D.; Van Golen, R.F.; Heger, M. Chapter 10—Reactive Oxygen and Nitrogen Species and Liver Ischemia-Reperfusion Injury: Role of Lipoic Acid. In The Liver; Patel, V.B., Rajendram, R., Preedy, V.R., Eds.; Academic Press: Boston, MA, USA, 2018; pp. 109–119. [Google Scholar]
- Çimen, B.; Çimen, L.; Çetin, I.; Çetin, A. Alpha-lipoic acid alleviates lipopolysaccharide-induced liver damage in rats via antioxidant effect. Dicle Med. J. 2019, 46, 125–132. [Google Scholar] [CrossRef][Green Version]
- El-Feki, M.A.; Amin, H.M.; Abdalla, A.A.; Fesal, M. Immunomodulatory and Anti-oxidant Effects of Alpha-Lipoic Acid and Vitamin E on Lipopolysaccharide-induced Liver Injury in Rats. Middle East J. Appl. Sci. 2016, 6, 460–467. [Google Scholar]
- Gorąca, A.; Huk-Kolega, H.; Kowalczyk, A.; Skibska, B. Anti-oxidative and anti-inflammatory effects of lipoic acid in rat liver. Postep Hig. Med. Dosw. 2015, 69, 270–276. [Google Scholar] [CrossRef]
- Tanaka, Y.; Kaibori, M.; Miki, H.; Nakatake, R.; Tokuhara, K.; Nishizawa, M.; Okumura, T.; Kwon, A.H. Alpha-lipoic acid exerts a liver-protective effect in acute liver injury rats. J. Surg. Res. 2015, 193, 675–683. [Google Scholar] [CrossRef]
- Armagan, I.; Bayram, D.; Candan, I.A.; Yigit, A.; Celik, E.; Armagan, H.H.; Uğuz, A.C. Effects of pentoxifylline and alpha lipoic acid on methotrexate-induced damage in liver and kidney of rats. Environ. Toxicol. Pharmacol. 2015, 39, 1122–1131. [Google Scholar] [CrossRef]
- Çakir, T.; Baştürk, A.; Polat, C.; Aslaner, A.; Durgut, H.; Şehirli, A.Ö.; Gül, M.; Öğünç, A.V.; Gül, S.; Sabuncuoglu, M.Z.; et al. Does alfa lipoic acid prevent liver from methotrexate induced oxidative injury in rats? Acta Cir. Bras. 2015, 30, 247–252. [Google Scholar] [CrossRef]
- Avci, B.; Bahadir, A.; Tuncel, O.K.; Bilgici, B. Influence of α-tocopherol and α-lipoic acid on bisphenol-A-induced oxidative damage in liver and ovarian tissue of rats. Toxicol. Ind. Health 2014, 32, 1381–1390. [Google Scholar] [CrossRef]
- Sena, M.C.; Cipriano, A.M.; Botelho, F.M.; Seiça, M.R. Lipoic Acid Prevents High-Fat Diet-Induced Hepatic Steatosis in Goto Kakizaki Rats by Reducing Oxidative Stress Through Nrf2 Activation. Int. J. Mol. Sci. 2018, 19, 2706. [Google Scholar] [CrossRef]
- Castro, M.C.; Massa, M.L.; Arbeláez, L.G.; Schinella, G.; Gagliardino, J.J.; Francini, F. Fructose-induced inflammation, insulin resistance and oxidative stress: A liver pathological triad effectively disrupted by lipoic acid. Life Sci. 2015, 137, 1–6. [Google Scholar] [CrossRef]
- Kaya-Dagistanli, F.; Tanriverdi, G.; Altinok, A.; Ozyazgan, S.; Ozturk, M. The effects of alpha lipoic acid on liver cells damages and apoptosis induced by polyunsaturated fatty acids. Food Chem. Toxicol. 2013, 53, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Stanković, M.N.; Mladenović, D.; Ninković, M.; Ethuričić, I.; Sobajić, S.; Jorgačević, B.; de Luka, S.; Vukicevic, R.J.; Radosavljević, T.S. The effects of α-lipoic acid on liver oxidative stress and free fatty acid composition in methionine-choline deficient diet-induced NAFLD. J. Med. Food 2014, 17, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Liu, J.; Pian, L.; Gui, S.; Lu, B. α-lipoic acid protects against carbon tetrachloride-induced liver cirrhosis through the suppression of the TGF-β/Smad3 pathway and autophagy. Mol. Med. Rep. 2019, 19, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Amirkhizi, F.; Hamedi-Shahraki, S.; Hosseinpour-Arjmand, S.; Vaghef-Mehrabany, E.; Ebrahimi-Mameghani, M. Effects of Alpha-Lipoic Acid Supplementation on Oxidative Stress Status in Patients with Non-Alcoholic Fatty Liver Disease: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Iran. Red Crescent Med. J. 2018, 20, e67615. [Google Scholar] [CrossRef]
- Rahmanabadi, A.; Mahboob, S.; Amirkhizi, F.; Hosseinpour-Arjmand, S.; Ebrahimi-Mameghani, M. Oral α-lipoic acid supplementation in patients with non-alcoholic fatty liver disease: Effects on adipokines and liver histology features. Food Funct. 2019, 10, 4941–4952. [Google Scholar] [CrossRef]
- Schaffer, S.; Kim, H.W. Effects and Mechanisms of Taurine as a Therapeutic Agent. Biomol. Ther. 2018, 26, 225–241. [Google Scholar] [CrossRef]
- Miyazaki, T.; Bouscarel, B.; Ikegami, T.; Honda, A.; Matsuzaki, Y. The Protective Effect of Taurine against Hepatic Damage in a Model of Liver Disease and Hepatic Stellate Cells; Taurine 7; Azuma, J., Schaffer, S.W., Ito, T., Eds.; Springer: New York, NY, USA, 2009; pp. 293–303. [Google Scholar]
- Abdel-Moneim, A.M.; Al-Kahtani, M.A.; El-Kersh, M.A.; Al-Omair, M.A. Free Radical-Scavenging, Anti-Inflammatory/Anti-Fibrotic and Hepatoprotective Actions of Taurine and Silymarin against CCl4 Induced Rat Liver Damage. PLoS ONE 2015, 10, e0144509. [Google Scholar] [CrossRef]
- Miyazaki, T.; Matsuzaki, Y. Taurine and liver diseases: A focus on the heterogeneous protective properties of taurine. Amino Acids 2014, 46, 101–110. [Google Scholar] [CrossRef]
- Wu, G.; Yang, J.; Sun, C.; Luan, X.; Shi, J.; Hu, J. Effect of Taurine on Alcoholic Liver Disease in Rats; Springer: New York, NY, USA, 2009; pp. 313–322. [Google Scholar]
- Devi, S.L.; Viswanathan, P.; Anuradha, C.V. Regression of liver fibrosis by taurine in rats fed alcohol: Effects on collagen accumulation, selected cytokines and stellate cell activation. Eur. J. Pharmacol. 2010, 647, 161–170. [Google Scholar] [CrossRef]
- Lin, C.-J.; Chiu, C.-C.; Chen, Y.-C.; Chen, M.-L.; Hsu, T.-C.; Tzang, B.-S. Taurine Attenuates Hepatic Inflammation in Chronic Alcohol-Fed Rats Through Inhibition of TLR4/MyD88 Signaling. J. Med. Food 2015, 18, 1291–1298. [Google Scholar] [CrossRef]
- Wu, G.; Tang, R.; Yang, J.; Tao, Y.; Liu, Z.; Feng, Y.; Lin, S.; Yang, Q.; Lv, Q.; Hu, J. Taurine accelerates alcohol and fat metabolism of rats with alcoholic Fatty liver disease. Adv. Exp. Med. Biol. 2015, 803, 793–805. [Google Scholar] [PubMed]
- Zhang, Z.; Liu, D.; Yi, B.; Liao, Z.; Tang, L.; Yin, D.; He, M. Taurine supplementation reduces oxidative stress and protects the liver in an iron-overload murine model. Mol. Med. Rep. 2014, 10, 2255–2262. [Google Scholar] [CrossRef] [PubMed]
- Gabr, S.A.; Gabr, N.S.; Elsaed, W.M. Protective activity of taurine and molecular fibrogenesis in iron overloaded hepatic tissues. Int. J. Pharmacol. 2019, 15, 418–427. [Google Scholar]
- Liang, J.; Deng, X.; Lin, Z.X.; Zhao, L.C.; Zhang, X.L. Attenuation of portal hypertension by natural taurine in rats with liver cirrhosis. World J. Gastroenterol. 2009, 15, 4529–4537. [Google Scholar] [CrossRef]
- Schwarzer, R.; Kivaranovic, D.; Mandorfer, M.; Paternostro, R.; Wolrab, D.; Heinisch, B.; Reiberger, T.; Ferlitsch, M.; Gerner, C.; Trauner, M.; et al. Randomised clinical study: The effects of oral taurine 6g/day vs. placebo on portal hypertension. Aliment. Pharmacol. Ther. 2018, 47, 86–94. [Google Scholar] [CrossRef]
- Gentile, C.L.; Nivala, A.M.; Gonzales, J.C.; Pfaffenbach, K.T.; Wang, D.; Wei, Y.; Jiang, H.; Orlicky, D.J.; Petersen, D.R.; Pagliassotti, M.J.; et al. Experimental evidence for therapeutic potential of taurine in the treatment of nonalcoholic fatty liver disease. Am. J. Physiol. Reg. I 2011, 301, R1710–R1722. [Google Scholar] [CrossRef]
- Charron, C.S.; Dawson, H.D.; Novotny, J.A. Garlic Influences Gene Expression In Vivo and In Vitro. J. Nutr. 2016, 146, 444S–449S. [Google Scholar] [CrossRef]
- Gong, Z.; Ye, H.; Huo, Y.; Wang, L.; Huang, Y.; Huang, M.; Yuan, X. S-allyl-cysteine attenuates carbon tetrachloride-induced liver fibrosis in rats by targeting STAT3/SMAD3 pathway. Am. J. Transl. Res. 2018, 10, 1337–1346. [Google Scholar]
- Somade, O.T.; Ugbaja, R.N.; Alli, A.A.; Odubote, O.T.; Yusuf, T.S.; Busari, B.T. Diallyl disulfide, an organo-sulfur compound in garlic and onion attenuates trichloromethane-induced hepatic oxidative stress, activation of NFkB and apoptosis in rats. J. Nutr. Intermed. Metab. 2018, 13, 10–19. [Google Scholar] [CrossRef]
- Takemura, S.; Azuma, H.; Osada-Oka, M.; Kubo, S.; Shibata, T.; Minamiyama, Y. S-allyl-glutathione improves experimental liver fibrosis by regulating Kupffer cell activation in rats. Am. J. Physiol. Gastr. L 2018, 314, G150–G163. [Google Scholar] [CrossRef]
- Wu, C.-C.; Sheen, L.-Y.; Chen, H.-W.; Kuo, W.-W.; Tsai, S.-J.; Lii, C.-K. Differential Effects of Garlic Oil and Its Three Major Organosulfur Components on the Hepatic Detoxification System in Rats. J. Agric. Food Chem. 2002, 50, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Zhang, R.; Gao, H.; Xiao, J.; Tipoe, G.L. Chapter 28—Garlic and Liver Diseases. In The Liver; Patel, V.B., Rajendram, R., Preedy, V.R., Eds.; Academic Press: Boston, MA, USA, 2018; pp. 337–347. [Google Scholar]
- Baek, S.-H.; Park, M.; Suh, J.-H.; Choi, H.-S. Protective Effects of an Extract of Young Radish (Raphanus sativus L) Cultivated with Sulfur (Sulfur-Radish Extract) and of Sulforaphane on Carbon Tetrachloride-Induced Hepatotoxicity. Biosci. Biotechnol. Biochem. 2008, 72, 1176–1182. [Google Scholar] [CrossRef] [PubMed]
- Gaona-Gaona, L.; Molina-Jijón, E.; Tapia, E.; Zazueta, C.; Hernández-Pando, R.; Calderón-Oliver, M.; Zarco-Márquez, G.; Pinzón, E.; Pedraza-Chaverri, J. Protective effect of sulforaphane pretreatment against cisplatin-induced liver and mitochondrial oxidant damage in rats. Toxicology 2011, 286, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Yang, S.; Lee, B.-S.; Jeong, S.Y.; Kim, K.-M.; Ku, S.-K.; Bae, J.-S. Hepatic protective effects of sulforaphane through the modulation of inflammatory pathways. J. Asian Nat. Prod. Res. 2019, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.-H.; Li, J.-W.; Sun, H.-T.; He, S.-Q.; Pang, J. Sulforaphane inhibits the activation of hepatic stellate cell by miRNA-423-5p targeting suppressor of fused. Hum. Cell 2019, 32, 403–410. [Google Scholar] [CrossRef]
- Mendivil, E.J.; Sandoval-Rodriguez, A.; Zuñiga-Ramos, L.M.; Santos-Garcia, A.; Armendariz-Borunda, J. Capsaicin and sulforaphane prevent experimental liver fibrosis via upregulation of peroxisome proliferator-activated receptor gamma and nuclear factor (erythroid-derived 2)-like 2. J. Funct. Foods 2019, 52, 382–388. [Google Scholar] [CrossRef]
- Ey, J.; Schömig, E.; Taubert, D. Dietary Sources and Antioxidant Effects of Ergothioneine. J. Agric. Food Chem. 2007, 55, 6466–6474. [Google Scholar] [CrossRef]
- Tang, R.M.Y.; Cheah, I.K.-M.; Yew, T.S.K.; Halliwell, B. Distribution and accumulation of dietary ergothioneine and its metabolites in mouse tissues. Sci. Rep. 2018, 8, 1601. [Google Scholar] [CrossRef]
- Gründemann, D.; Harlfinger, S.; Golz, S.; Geerts, A.; Lazar, A.; Berkels, R.; Jung, N.; Rubbert, A.; Schömig, E. Discovery of the ergothioneine transporter. Proc. Natl. Acad. Sci. USA 2005, 102, 5256–5261. [Google Scholar] [CrossRef]
- Kato, Y.; Kubo, Y.; Iwata, D.; Kato, S.; Sudo, T.; Sugiura, T.; Kagaya, T.; Wakayama, T.; Hirayama, A.; Sugimoto, M.; et al. Gene Knockout and Metabolome Analysis of Carnitine/Organic Cation Transporter OCTN1. J. Pharm. Res. 2010, 27, 832–840. [Google Scholar] [CrossRef]
- Franzoni, F.; Colognato, R.; Galetta, F.; Laurenza, I.; Barsotti, M.; Di Stefano, R.; Bocchetti, R.; Regoli, F.; Carpi, A.; Balbarini, A.; et al. An in vitro study on the free radical scavenging capacity of ergothioneine: Comparison with reduced glutathione, uric acid and trolox. Biomed. Pharmacother. 2006, 60, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Rougee, M.; Bensasson, R.V.; Land, E.J.; Pariente, R. Deactivation of singlet molecular oxygen by thiols and related compounds, possible protectors against skin photosensitivity. Photochem. Photobiol. 1988, 47, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Colognato, R.; Laurenza, I.; Fontana, I.; Coppedé, F.; Siciliano, G.; Coecke, S.; Aruoma, O.I.; Benzi, L.; Migliore, L. Modulation of hydrogen peroxide-induced DNA damage, MAPKs activation and cell death in PC12 by ergothioneine. Clin. Nutr. 2006, 25, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Laurenza, I.; Prato, S.D.; Benzi, L.; Colognato, R.; Migliore, L. Modulation of palmitic acid-induced cell death by ergothioneine: Evidence of an anti-inflammatory action. BioFactors 2008, 33, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Gilmour, P.S.; Jimenez, L.A.; Biswas, S.K.; Antonicelli, F.; Aruoma, O.I. Ergothioneine inhibits oxidative stress- and TNF-α-induced NF-κB activation and interleukin-8 release in alveolar epithelial cells. Biochem. Biophys. Res. Commun. 2003, 302, 860–864. [Google Scholar] [CrossRef]
- Hseu, Y.-C.; Lo, H.-W.; Korivi, M.; Tsai, Y.-C.; Tang, M.-J.; Yang, H.-L. Dermato-protective properties of ergothioneine through induction of Nrf2/ARE-mediated antioxidant genes in UVA-irradiated Human keratinocytes. Free Radic. Biol. Med. 2015, 86, 102–117. [Google Scholar] [CrossRef]
- Cheah, I.K.; Tang, R.; Ye, P.; Yew, T.S.Z.; Lim, K.H.C.; Halliwell, B. Liver ergothioneine accumulation in a guinea pig model of non-alcoholic fatty liver disease. A possible mechanism of defence? Free Radic. Res. 2016, 50, 14–25. [Google Scholar] [CrossRef]
- Bedirli, A.; Sakrak, O.; Muhtaroglu, S.; Soyuer, I.; Guler, I.; Riza Erdogan, A.; Sozuer, E.M. Ergothioneine pretreatment protects the liver from ischemia-reperfusion injury caused by increasing hepatic heat shock protein 70. J. Surg. Res. 2004, 122, 96–102. [Google Scholar] [CrossRef]
- Tang, Y.; Masuo, Y.; Sakai, Y.; Wakayama, T.; Sugiura, T.; Harada, R.; Futatsugi, A.; Komura, T.; Nakamichi, N.; Sekiguchi, H.; et al. Localization of Xenobiotic Transporter OCTN1/SLC22A4 in Hepatic Stellate Cells and Its Protective Role in Liver Fibrosis. J. Pharm. Sci. 2016, 105, 1779–1789. [Google Scholar] [CrossRef]
- Palumbo, A.; Castellano, I.; Napolitano, A. Ovothiol: A Potent Natural Antioxidant from Marine Organisms. In Blue Biotechnology; La Barre, S.S., Bates, S., Eds.; Wiley VCH: Weinheim, Germany, 2018; pp. 583–610. [Google Scholar]
- Braunshausen, A.; Seebeck, F.P. Identification and Characterization of the First Ovothiol Biosynthetic Enzyme. J. Am. Chem. Soc. 2011, 133, 1757–1759. [Google Scholar] [CrossRef]
- Naowarojna, N.; Cheng, R.; Chen, L.; Quill, M.; Xu, M.; Zhao, C.; Liu, P. Mini-Review: Ergothioneine and Ovothiol Biosyntheses, an Unprecedented Trans-Sulfur Strategy in Natural Product Biosynthesis. Biochemistry 2018, 57, 3309–3325. [Google Scholar] [CrossRef] [PubMed]
- Castellano, I.; Seebeck, F.P. On ovothiol biosynthesis and biological roles: From life in the ocean to therapeutic potential. Nat. Prod. Rep. 2018, 35, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Castellano, I.; Migliaccio, O.; D’Aniello, S.; Merlino, A.; Napolitano, A.; Palumbo, A. Shedding light on ovothiol biosynthesis in marine metazoans. Sci. Rep. 2016, 6, 21506. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.L.; Russo, M.; Castellano, I.; Napolitano, A.; Palumbo, A. Ovothiol isolated from sea urchin oocytes induces autophagy in the Hep-G2 cell line. Mar. Drugs 2014, 12, 4069–4085. [Google Scholar] [CrossRef]
- Brancaccio, M.; Russo, M.; Masullo, M.; Palumbo, A.; Russo, G.L.; Castellano, I. Sulfur-containing histidine compounds inhibit gamma-glutamyl transpeptidase activity in human cancer cells. J. Biol. Chem. 2019, 294, 14603–14614. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Model System | Concentration | Patent |
|---|---|---|---|
| Glutathione (GSH) | Wistar rats with CCl4-induced liver fibrosis | 34.1 mg/g [43] | n = 5 US201816232747 CN201811444979 CN201710216398 WO2011US23434 KR19920005534 |
| Lipoic acid (ALA) | C57BL/6 mice with BDL-induced liver fibrosis | 100 mg/kg [46] | n = 1 WO2017KR15370 |
| Wistar rats with CCl4-induced liver fibrosis | 30 mg/kg [47] | ||
| Garlic derived sulfur compounds (SAC, DADS, SAG) | Rat with CCl4-induced liver fibrosis | 200 mg/kg (SAC) [30,31] | n = 1 JP20050270344 |
| Wistar rats with porcine serum-induced liver fibrosis | 0.15% of diet (SAC) [34] | ||
| Rat with CCl4-induced liver fibrosis | 50 mg/kg (SAC) [77] | ||
| Rat with trichloromethane (TCM) induced liver fibrosis | 50 mg/kg (DADS) [78] | ||
| Rat with CCl4-induced liver fibrosis | 200 mg/kg (SAG) [79] | ||
| Ergothioneine (erg) | Guinea pig model of NAFLD | 4%–10% of diet [97] | n = 1 USOO6555141B1 |
| Ovothiol (ovo) | Male balb-c CCl4-induced liver fibrosis | 50 mg/kg [36] | n = 2 WO2019/08247A1 RBI15865-IT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milito, A.; Brancaccio, M.; D’Argenio, G.; Castellano, I. Natural Sulfur-Containing Compounds: An Alternative Therapeutic Strategy against Liver Fibrosis. Cells 2019, 8, 1356. https://doi.org/10.3390/cells8111356
Milito A, Brancaccio M, D’Argenio G, Castellano I. Natural Sulfur-Containing Compounds: An Alternative Therapeutic Strategy against Liver Fibrosis. Cells. 2019; 8(11):1356. https://doi.org/10.3390/cells8111356
Chicago/Turabian StyleMilito, Alfonsina, Mariarita Brancaccio, Giuseppe D’Argenio, and Immacolata Castellano. 2019. "Natural Sulfur-Containing Compounds: An Alternative Therapeutic Strategy against Liver Fibrosis" Cells 8, no. 11: 1356. https://doi.org/10.3390/cells8111356
APA StyleMilito, A., Brancaccio, M., D’Argenio, G., & Castellano, I. (2019). Natural Sulfur-Containing Compounds: An Alternative Therapeutic Strategy against Liver Fibrosis. Cells, 8(11), 1356. https://doi.org/10.3390/cells8111356