Cell Adhesion Molecules Are Mediated by Photobiomodulation at 660 nm in Diabetic Wounded Fibroblast Cells




Abstract
1. Introduction
1.1. Wound Healing and Diabetes
1.2. Cell Adhesion Molecules
1.3. Photobiomodulation (PBM)
2. Materials and Methods
2.1. Cell Isolation and Culure
2.2. Laser Irradiation
2.3. PCR Array
2.4. Statistical Analysis
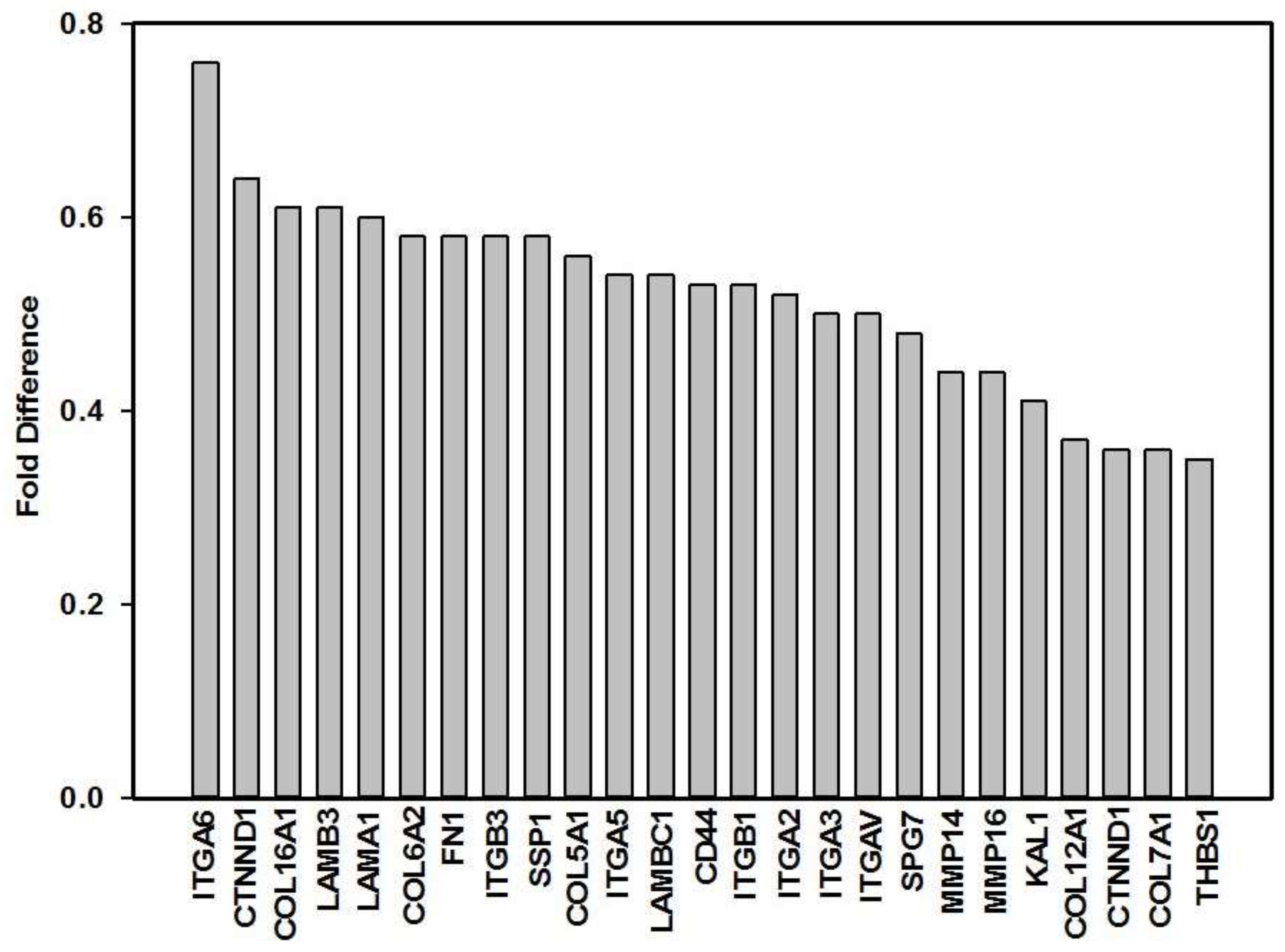
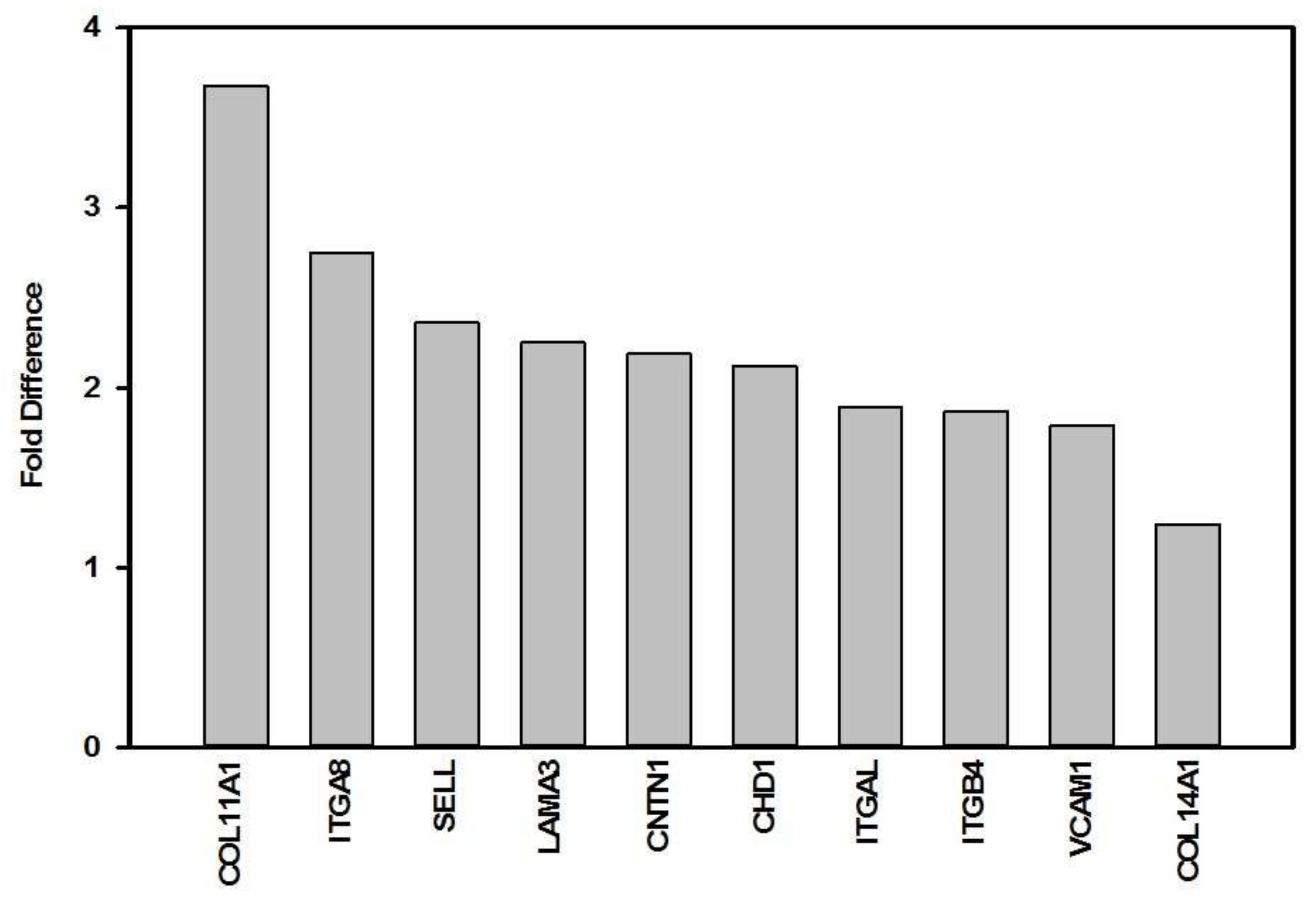
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ayuk, S.M.; Houreld, N.N.; Abrahamse, H. Collagen production in diabetic wounded fibroblasts in response to low-intensity laser irradiation at 660 nm. Diabetes Technol. Ther. 2012, 14, 1110–1117. [Google Scholar] [CrossRef] [PubMed]
- Caley, M.P.; Martins, V.L.C.; O’Toole, E.A. Metalloproteinases and wound healing. Adv. Wound Care 2015, 4, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Lan, C.-C.E.; Wu, C.-S.; Huang, S.-M.; Wu, I.H.; Chen, G.-S. High-glucose environment enhanced oxidative stress and increased interleukin-8 secretion from keratinocytes. Diabetes 2013, 62, 2530–2538. [Google Scholar] [CrossRef] [PubMed]
- McCarty, S.M.; Percival, S.L. Proteases and delayed wound healing. Adv. Wound Care 2013, 2, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wu, T.; Wu, J.; Zhao, L.; Li, Q.; Varghese, Z.; Moorhead, J.F.; Powis, S.H.; Chen, Y.; Ruan, X.Z. Chronic inflammation exacerbates glucose metabolism disorders in C57BL/6J mice fed with high-fat diet. J. Endocrinol. 2013, 219, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Köppel, H.; Riedl, E.; Braunagel, M.; Sauerhoefer, S.; Ehnert, S.; Godoy, P.; Sternik, P.; Dooley, S.; Yard, B.A. L-carnosine inhibits high-glucose-mediated matrix accumulation in human mesangial cells by interfering with TGF-β production and signalling. Nephrol. Dial. Transplant. 2011, 26, 3852–3858. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Liu, W.; Lan, T.; Xie, X.; Peng, J.; Huang, J.; Wang, S.; Shen, X.; Liu, P.; Huang, H. Berberine reduces fibronectin expression by suppressing the S1P-S1P2 receptor pathway in experimental diabetic nephropathy models. PLoS ONE 2012, 7, e43874. [Google Scholar] [CrossRef]
- Taniguchi, K.; Xia, L.; Goldberg, H.J.; Lee, K.W.; Shah, A.; Stavar, L.; Masson, E.; Momen, A.; Shikatani, E.A.; John, R. Inhibition of Src kinase blocks high glucose–induced EGFR transactivation and collagen synthesis in mesangial cells and prevents diabetic nephropathy in mice. Diabetes 2013, 62, 3874–3886. [Google Scholar] [CrossRef] [PubMed]
- IDF, International Diabetes Federation in Diabetes Atlas, 2015. Available online: https://www.idf.org/diabetes atlas (accessed on 8 March 2018).
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed]
- Rozario, T.; DeSimone, D.W. The extracellular matrix in development and morphogenesis: A dynamic view. Dev. Biol. 2010, 341, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Wang, N. Cell adhesion Molecules (CAMs). In Encyclopedia of the Neurological Sciences, 2nd ed.; Daroff, R., Aminoff, M.J., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 628–629. ISBN 9780123851574. [Google Scholar]
- Crawford, M.; Dagnino, L. Cadherins as Central Modulators of Wound Repair. In Wound Healing: Stem Cells Repair and Restorations, Basic and Clinical Aspects; Turksen, K., Ed.; Wiley & Sons Inc.: Hoboken, NJ, USA, 2018; p. 15. ISBN 1119282497. [Google Scholar]
- Boettiger, D. Mechanical control of integrin-mediated adhesion and signaling. Curr. Opin. Cell Biol. 2012, 24, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Jakhu, H.; Gill, G.; Singh, A. Role of integrins in wound repair and its periodontal implications. J. Oral. Biol. Craniofac. Res. 2018, in press. [Google Scholar] [CrossRef]
- Koivisto, L.; Heino, J.; Lari Häkkinen, L.; Larjava, H. Integrins in Wound Healing. Adv. Wound Care 2014, 3, 762–783. [Google Scholar] [CrossRef] [PubMed]
- Schnittert, J.; Bansal, R.; Storm, G.; Prakash, J. Integrins in wound healing, fibrosis and tumor stroma: High potential targets for therapeutics and drug delivery. Adv. Drug Del. Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Shi-Wen, X.; Blumbach, K.; Eastwood, M.; Denton, C.P.; Eckes, B.; Krieg, T.; Abraham, D.J.; Leask, A. Expression of integrin β1 by fibroblasts is required for tissue repair in vivo. J. Cell Sci. 2010, 123, 3674–3682. [Google Scholar] [CrossRef] [PubMed]
- Leckband, D.; de Rooij, J. Cadherin adhesion and mechanotransduction. Annu Rev. Cell Dev. Biol. 2014, 30, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, A.; Rao, M.V.; Wu, Y.; Zaidel-Bar, R. Jack of all trades: Functional modularity in the adherens junction. Curr. Opin. Cell Biol. 2015, 36, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Sanders, A.J.; Jiang, D.G.; Jiang, W.G.; Harding, K.G.; Patel, G.K. Activated leukocyte cell adhesion molecule impacts on clinical wound healing and inhibits HaCaT migration. Int. Wound J. 2011, 8, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Miner, J.J.; Yago, T.; Yao, L.; Lupu, F.; Xia, L.; McEver, R.P. Differential regulation of human and murine P-selectin expression and function in vivo. J. Exp. Med. 2010, 207, 2975–2987. [Google Scholar] [CrossRef] [PubMed]
- McEver, R.P. Selectins: Initiators of leucocyte adhesion and signalling at the vascular wall. Cardiovasc. Res. 2015, 107, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Anders, J.J.; Lanzafame, R.J.; Arany, P.R. Low-level light/laser therapy versus photobiomodulation therapy. Photomed. Laser Surg. 2015, 33, 183–184. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Li, Y.-X.; Chen, H.; Kang, M.; Cheng-Yi, L.T. Effects of Low-Intensity Laser Irradiation on Wound Healing in Diabetic Rats. Int. J. Photoenergy 2012, 2012. [Google Scholar] [CrossRef]
- Farivar, S.; Malekshahabi, T.; Shiari, R. Biological effects of low level laser therapy. Lasers Med. Sci. 2014, 2, 58–62. [Google Scholar]
- Guptaa, A.; Avcia, P.; Sadasivam, M.; Chandran, R.; Parizotto, N.; Vecchio, D.; de Melo, W.C.; Dai, T.; Chiang, L.Y.; Hamblin, M.R. Shining light on nanotechnology to help repair and regeneration. Biotechnol. Adv. 2013, 31, 607–631. [Google Scholar] [CrossRef] [PubMed]
- Ayuk, S.M.; Abrahamse, H.; Houreld, N.N. The role of photobiomodulation on gene expression of cell adhesion molecules in diabetic wounded fibroblasts in vitro. J. Photochem. Photobiol. B Biol. 2016, 161, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.S.; Salman, S.D. Low level diode laser accelerates wound healing. Lasers Med. Sci. 2013, 28, 941–945. [Google Scholar] [CrossRef] [PubMed]
- Masha, R.T.; Houreld, N.N.; Abrahamse, H. Low-intensity laser irradiation at 660 nm stimulates transcription of genes involved in the electron transport chain. Photomed. Laser Surg. 2013, 31, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, N.C.; Brunelli, R.; de Araújo, H.S.S.; Parizotto, N.A.; Renno, A.C.M. Low-level laser therapy (LLLT)(660nm) alters gene expression during muscle healing in rats. J. Photochem. Photobiol. B Biol. 2013, 120, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Tschon, M.; Incerti-Parenti, S.; Cepollaro, S.; Checchi, L.; Fini, M. Photobiomodulation with low-level diode laser promotes osteoblast migration in an in vitro micro wound model. J. Biomed. Opt. 2015, 20, 78002. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Dai, T.; Sharma, S.K.; Huang, Y.-Y.; Carroll, J.D.; Hamblin, M.R. The nuts and bolts of low-level laser (light) therapy. Ann. Biomed. Eng. 2012, 40, 516–533. [Google Scholar] [CrossRef] [PubMed]
- Peplow, P.V.; Chung, T.-Y.; Ryan, B.; Baxter, G.D. Laser photobiostimulation of wound healing: Reciprocity of irradiance and exposure time on energy density for splinted wounds in diabetic mice. Lasers Surg. Med. 2011, 43, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Primo, F.; De Paula, L.; de Siqueira-Moura, M.; Tedesco, A. Photobiostimulation on wound healing treatment by ClAlPc-nanoemulsion from a multiple-wavelength portable light source on a 3D-human stem cell dermal equivalent. Curr. Med. Chem. 2012, 19, 5157–5163. [Google Scholar] [CrossRef] [PubMed]
- Feitosa, M.C.P.; Carvalho, A.F.M.D.; Feitosa, V.C.; Coelho, I.M.; Oliveira, R.A.D.; Arisawa, E.Â.L. Effects of the low-level laser therapy (LLLT) in the process of healing diabetic foot ulcers. Acta Cir. Bras. 2015, 30, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Fekrazad, R.; Asefi, S.; Allahdadi, M.; Kalhori, K.A. Effect of Photobiomodulation on Mesenchymal Stem Cells. Photomed. Laser Surg. 2016, 34, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Lau, P.; Bidin, N.; Krishnan, G.; AnaybBaleg, S.M.; Sum, M.B.M.; Bakhtiar, H.; Nassir, Z.; Hamid, A. Photobiostimulation effect on diabetic wound at different power density of near infrared laser. J. Photochem. Photobiol. B Biol. 2015, 151, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Oton-Leite, A.F.; Silva, G.B.L.; Morais, M.O.; Silva, T.A.; Leles, C.R.; Valadares, M.C.; Pinezi, J.C.D.; Batista, A.C.; Mendonça, E.F. Effect of low-level laser therapy on chemoradiotherapy-induced oral mucositis and salivary inflammatory mediators in head and neck cancer patients. Lasers Surg. Med. 2015, 47, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Peplow, P.V.; Baxter, G.D. Defining a therapeutic window for laser irradiation (810 nm) applied to the inguinal region to ameliorate diabetes in diabetic mice. Photomed. Laser Surg. 2014, 32, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.D.U.E.; Paraguassu, G.M.; De Aquino Xavier, F.C.; Ramalho, M.J.P.; Rodrigues, T.T.; Dos Santos, J.N.; Ramalho, L.M.P. Effect of Laser Phototherapy on Wound Healing in Rats Exposed to a Hyperlipidic Diet. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2015, 120, e84. [Google Scholar] [CrossRef]
- Houreld, N.; Abrahamse, H. In vitro exposure of wounded diabetic fibroblast cells to a helium-neon laser at 5 and 16 J/cm2. Photomed. Laser Surg. 2007, 25, 78–84. [Google Scholar] [CrossRef] [PubMed]
- McDermott, A.M.; Kern, T.S.; Murphy, C.J. The effect of elevated extracellular glucose on migration, adhesion and proliferation of SV40 transformed human corneal epithelial cells. Curr. Eye Res. 1998, 17, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Houreld, N.; Abrahamse, H. Laser light influences cellular viability and proliferation in diabetic-wounded fibroblast cells in a dose- and wavelength-dependent manner. Lasers Med. Sci. 2008, 23, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Ponomareva, L.; Veeranki, S.; Choubey, D. IFI16 induction by glucose restriction in human fibroblasts contributes to autophagy through activation of the ATM/AMPK/p53 pathway. PLoS ONE 2011, 6, e19532. [Google Scholar] [CrossRef] [PubMed]
- Esmaeelinejad, M.; Bayat, M.; Darbandi, H.; Bayat, M.; Mosaffa, N. The effects of low-level laser irradiation on cellular viability and proliferation of human skin fibroblasts cultured in high glucose mediums. Lasers Med. Sci. 2014, 29, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Góralczyk, K.; Szymanska, J.; Szot, K.; Fisz, J.; Rosc, D. Low-level laser irradiation effect on endothelial cells under conditions of hyperglycemia. Lasers Med. Sci. 2016, 31, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Li, M.F.; Zhang, R.; Li, T.T.; Chen, M.Y.; Li, L.X.; Lu, J.X.; Jia, W.P. High glucose increases the expression of inflammatory cytokine genes in macrophages through H3K9 methyltransferase mechanism. J. Interferon Cytokine Res. 2016, 36, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.-C.; Park, A.P.; Guan, J.-L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Cory, G. Scratch-wound assay. Methods Mol. Biol. 2011, 769, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, H.; Chen, J. Human epidermal growth factor coupled to different structural classes of cell penetrating peptides: A comparative study. Int. J. Biol. Macromol. 2017, 105, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Houreld, N.N.; Masha, R.T.; Abrahamse, H. Low-intensity laser irradiation at 660 nm stimulates cytochrome c oxidase in stressed fibroblast cells. Lasers Surg. Med. 2012, 44, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Houreld, N.N.; Ayuk, S.M.; Abrahamse, H. Expression of genes in normal fibroblast cells (WS1) in response to irradiation at 660 nm. J. Photochem. Photobiol. B Biol. 2014, 120, 146–152. [Google Scholar] [CrossRef] [PubMed]
- National Centre for Biotechnology Information, Genes. Available online: https://www.ncbi.nlm.nih.gov/genome/ (accessed on 26 February 2018).
- LeBlanc, S.; Arabzadeh, A.; Benlolo, S.; Breton, V.; Turbide, C.; Beauchemin, N.; Nouvion, A.L. CEACAM1 deficiency delays important wound healing processes. Wound Repair. Regen. 2011, 19, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Keshri, G.K.; Gupta, A.; Yadav, A.; Sharma, S.K.; Singh, S.B. Photobiomodulation with Pulsed and Continuous Wave Near-Infrared Laser (810 nm, Al-Ga-As) Augments Dermal Wound Healing in Immunosuppressed Rats. PLoS ONE 2016, 11, e0166705. [Google Scholar] [CrossRef] [PubMed]
- Jere, S.W.; Houreld, N.N.; Abrahamse, H. Photobiomodulation at 660 nm stimulates proliferation and migration of diabetic wounded cells via the expression of epidermal growth factor and the JAK/STAT pathway. J. Photochem. Photobiol. B Biol. 2018, 179, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Tatmatsu Rocha, J.C.; Ferraresi, C.; Hamblin, M.R.; Damasceno, F.M.; Falcão do Nascimento, N.R.; Driusso, P.; Parizotto, N.A. Low-Level Laser Therapy (904nm) Can increase collagen and reduce oxidative and nitrosative stress in diabetic wounded mouse skin. J. Photochem. Photobiol. B Biol. 2016, 164, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, K.H.; Meyer-Hamme, G.; Schröder, S. Low Level Laser Therapy for the Treatment of Diabetic Foot Ulcers: A Critical Survey. Evid.-Based Complement. Altern. Med. 2014. [Google Scholar] [CrossRef] [PubMed]
- Ruh, A.C.; Frigo, L.; Cavalcanti, M.F.X.B.; Svidnicki, P.; Vicari, V.N.; Lopes-Martins, R.A.B.; Pinto Leal, E.C.; De Isla, N.; Diomede, F.; Trubiani, O.; et al. Laser photobiomodulation in pressure ulcer healing of human diabetic patients: Gene expression analysis of inflammatory biochemical markers. Lasers Med. Sci. 2018, 33, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Ayuk, S.M.; Abrahamse, H.; Houreld, N.N. Photobiomodulation alters matrix protein activity in stressed fibroblast cells in vitro. J. Biophotonics 2018, 11, e201700127. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, P.D.T.C.D.; Silva, I.S.D.; Reis, F.A.D.; Perreira, D.M.; Aydos, R.D. Influence of ingaalp laser (660 nm) on the healing of skin wounds in diabetic rats. Acta Cir. Bras. 2010, 25, 71–79. [Google Scholar] [CrossRef]
- Colombo, F.; Neto, A.D.A.P.V.; Sousa, A.P.C.D.; Marchionni, A.M.T.; Pinheiro, A.L.B.; Reis, S.R.D.A. Effect of low-level laser therapy (660 nm) on angiogenesis in wound healing: A immunohistochemical study in a rodent model. Braz. Dent. J. 2013, 24, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Gharagozlian, S.; Svennevig, K.; Bangstad, H.-J.; Winberg, J.-O.; Kolset, S.O. Matrix metalloproteinases in subjects with type 1 diabetes. BMC Clin. Pathol. 2009, 9. [Google Scholar] [CrossRef] [PubMed]
- Casalechi, H.L.; Leal-Junior, E.C.P.; Xavier, M.; Silva, J.A., Jr.; de Carvalho Pde, T.; Aimbire, F.; Albertini, R. Low-level laser therapy in experimental model of collagenase-induced tendinitis in rats: Effects in acute and chronic inflammatory phases. Lasers Med. Sci. 2013, 28, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2010, 339, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.; Lorenzini, L.; Alessandri, M.; Torricella, R.; Baldassarro, V.A.; Giardino, L.; Calzà, L. In vitro exposure to very low-level laser modifies expression level of extracellular matrix protein RNAs and mitochondria dynamics in mouse embryonic fibroblasts. BMC Complement. Altern. Med. 2015, 15, 78. [Google Scholar] [CrossRef] [PubMed]
- Shih, W.; Yamada, S. N-cadherin-mediated cell-cell adhesion promotes cell migration in a three-dimensional matrix. J. Cell Sci. 2012, 125, 3661–3670. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Parameters | |
|---|---|
| Wavelength | 660 nm |
| Output power | 92.8 mW |
| Spot size | 9.1 cm2 |
| Output power density | 10.22 mW/cm2 |
| Fluence | 5 J/cm2 |
| Duration of irradiation | 8 min 9 s |
| Pathway | Gene |
|---|---|
| Transmembrane Molecules | CD44, CDH1, HAS1, ICAM1, ITGA1, ITGA2, ITGA3, ITGA4, ITGA5, ITGA6, ITGA7, ITGA8, ITGAL, ITGAM, ITGAV, ITGB1, ITGB2, ITGB3, ITGB4, ITGB5, MMP14, MMP15, MMP16, NCAM1, PECAM1, SELE, SELL, SELP, SGCE, SPG7, VCAM1 |
| Cell-Cell Adhesion | CD44, CDH1, COL6A2, COL11A1, COL14A1, CTNND1, ICAM1, ITGA8, VCAM1 |
| Cell-Matrix Adhesion | ADAMTS13, CD44, ITGA1, ITGA2, ITGA3, ITGA4, ITGA5, ITGA6, ITGA7, ITGA8, ITGAL, ITGAM, ITGAV, ITGB1, ITGB2, ITGB3, ITGB4, ITGB5, SGCE, SPP1, THBS3 |
| Other Adhesion Molecules | CLEC3B, CNTN1, COL5A1, COL6A1, COL7A1, COL8A1, COL12A1, COL15A1, COL16A1, CTGF, CTNNA1, CTNNB1, CTNND2, FN1, KAL1, LAMA1, LAMA2, LAMA3, LAMB1, LAMB3, LAMC1, THBS1, THBS2, TNC, VCAN, VTN |
| Housekeeping genes | B2M, HPRT1, RPL13A, GAPDH, ACTB |
| Gene Symbol | Description | Gene ID | Fold Difference 1 | p-Value | Gene Function [54] |
|---|---|---|---|---|---|
| ADAMTS13 | ADAM metallopeptidase with thrombospondin type 1 motif, 13 | 11093 | 0.91 | 0.589 | The enzyme encoded by this gene specifically cleaves von Willebrand Factor (vWF). |
| CD44 | CD44 molecule (Indian blood group) | 960 | 0.53 | 0.049 * | The protein encoded by this gene is a cell-surface glycoprotein involved in cell-cell interactions, cell adhesion and migration. It is a receptor for hyaluronic acid (HA) and can also interact with other ligands, such as osteopontin, collagens, and matrix metalloproteinases (MMPs). |
| CDH1 | Cadherin 1, type 1, E-cadherin | 999 | 2.12 | 0.050 * | This gene encodes a classical cadherin of the cadherin superfamily, and is a cell-cell adhesion protein. |
| CTNNA1 | Catenin (cadherin-associated protein), alpha 1, 102 kDa | 1495 | 0.83 | 0.340 | This gene encodes a member of the catenin family of proteins that play an important role in cell adhesion process by connecting cadherins located on the plasma membrane to the actin filaments inside the cell. |
| CTNNB1 | Catenin (cadherin-associated protein), beta 1, 88 kDa | 1499 | 0.64 | 0.084 | The protein encoded by this gene is part of a complex of proteins that constitute adherens junctions, which are necessary for the creation and maintenance of epithelial cell layers by regulating cell growth and adhesion between cells. They also anchor the actin cytoskeleton and may be responsible for transmitting the contact inhibition signal that causes cells to stop dividing once the epithelial sheet is complete. |
| CTNND1 | Catenin (cadherin-associated protein), delta 1 | 1500 | 0.36 | 0.005 ** | This gene encodes a member of the Armadillo protein family, which function in adhesion between cells and signal transduction. |
| CTNND2 | Catenin (cadherin-associated protein), delta 2 (neural plakophilin-related arm-repeat protein) | 1501 | 2.72 | 0.052 | This gene encodes an adhesive junction associated protein of the armadillo/beta-catenin superfamily. The protein also promotes the disruption of E-cadherin based adherens junction to favour cell spreading upon stimulation by hepatocyte growth factor. |
| CLEC3B | C-type lectin domain family 3, member B | 7123 | 1.14 | 0.749 | This gene codes for the protein tetranectin which may be involved in the packaging of molecules destined for exocytosis. |
| CNTN1 | Contactin 1 | 1272 | 2.19 | 0.041 * | The protein encoded by this gene is a member of the immunoglobulin superfamily. It is a glycosyl-phosphatidylinositol (GPI)-anchored neuronal membrane protein that functions as a cell adhesion molecule. |
| CTGF | Connective tissue growth factor | 1490 | 1.10 | 0.521 | The protein encoded by this gene is a mitogen that is secreted by vascular endothelial cells. The encoded protein plays a role in chondrocyte proliferation and differentiation, cell adhesion in many cell types, and is related to platelet-derived growth factor. |
| COL5A1 | Collagen, type V, alpha 1 | 1289 | 0.56 | 0.036 * | This gene encodes an alpha chain for one of the low abundance fibrillar collagens. Type V collagen is found in tissues containing type I collagen and appears to regulate the assembly of heterotypic fibbers composed of both type I and type V collagen. This gene product is closely related to type XI collagen and it is possible that the collagen chains of types V and XI constitute a single collagen type with tissue-specific chain combinations. |
| COL6A1 | Collagen, type VI, alpha 1 | 1291 | 0.64 | 0.015 * | The basic structural unit of collagen VI is a heterotrimer of the alpha1(VI), alpha2(VI), and alpha3(VI) chains. The protein encoded by this gene is the alpha 1 subunit of type VI collagen (alpha1(VI) chain). Collagen VI is a major structural component of microfibrils. |
| COL6A2 | Collagen, type VI, alpha 2 | 1292 | 0.58 | 0.014 * | The protein encoded by this gene is the alpha 2 subunit of type VI collagen (alpha2(VI) chain). Type VI collagen is a beaded filament collagen found in most connective tissues. The product of this gene contains several domains similar to von Willebrand Factor type A domains. These domains have been shown to bind ECM proteins, which explains the importance of this collagen in organizing matrix components. |
| COL7A1 | Collagen, type VII, alpha 1 | 1294 | 0.36 | 0.017 * | This gene encodes the alpha 1 subunit of type VII collagen. Type VII collagen fibril, composed of three identical alpha collagen chains, is restricted to the basement zone and functions as an anchoring fibril between the external epithelia and the underlying stroma. |
| COL8A1 | Collagen, type VIII, alpha 1 | 1295 | 0.71 | 0.271 | This gene encodes the alpha 1 subunit (one of two alpha chains) of type VIII collagen. The gene product is a short chain collagen and a major component of the basement membrane of the corneal endothelium. |
| COL11A1 | Collagen, type XI, alpha 1 | 1301 | 3.68 | 0.002 ** | This gene encodes the alpha 1 subunit (one of two chains) of type XI collagen, a minor fibrillar collagen. |
| COL12A1 | Collagen, type XII, alpha 1 | 1303 | 0.37 | 0.003 ** | This gene encodes the alpha 1 subunit of type XII collagen, a member of the FACIT (fibril-associated collagens with interrupted triple helices) collagen family. Type XII collagen is a homotrimer found in association with type I collagen, an association that is thought to modify the interactions between collagen I fibrils and the surrounding matrix. |
| COL14A1 | Collagen, type XIV, alpha 1 | 7373 | 1.24 | 0.023 * | This gene encodes the alpha 1 subunit of type XIV collagen, a member of the FACIT collagen family. Type XIV collagen interacts with the fibril surface and is involved in the regulation of fibrillogenesis. |
| COL15A1 | Collagen, type XV, alpha 1 | 1306 | 0.67 | 0.082 | This gene encodes the alpha 1 subunit of type XV collagen, a member of the FACIT collagen family. Type XV collagen has a wide tissue distribution but the strongest expression is localized to basement membrane zones so it may function to adhere basement membranes to underlying connective tissue stroma. |
| COL16A1 | Collagen, type XVI, alpha 1 | 1307 | 0.61 | 0.001 ** | This gene encodes the alpha 1 subunit of type XVI collagen, a member of the FACIT collagen family. Members of this collagen family are found in association with fibril-forming collagens such as type I and II, and serve to maintain the integrity of the ECM. |
| FN1 | Fibronectin 1 | 2335 | 0.58 | 0.008 ** | This gene encodes fibronectin, a glycoprotein present in a dimeric or multimeric form at the cell surface and in ECM. The encoded preproprotein is proteolytically processed to generate the mature protein. Fibronectin is involved in cell adhesion and migration processes including wound healing. |
| HAS1 | Hyaluronan synthase 1 | 3036 | 0.87 | 0.544 | Hyaluronan or hyaluronic acid (HA) is a polysaccharide and is a constituent of the ECM. It serves a variety of functions, including space filling, lubrication of joints, and provision of a matrix through which cells can migrate. HA is actively produced during wound healing and tissue repair to provide a framework for ingrowth of blood vessels and fibroblasts. HA is synthesized by membrane-bound synthase at the inner surface of the plasma membrane. |
| ICAM1 | Intercellular adhesion molecule 1 | 3383 | 0.97 | 0.843 | This gene encodes a cell surface glycoprotein which is typically expressed on endothelial cells and cells of the immune system. It binds to integrins of type CD11a/CD18, or CD11b/CD18. |
| ITGA1 | Integrin, alpha 1 | 3672 | 0.97 | 0.721 | This gene encodes the alpha 1 subunit of integrin receptors. This protein heterodimerizes with the beta 1 subunit to form a cell-surface receptor for collagen and laminin. The heterodimeric receptor is involved in cell-cell adhesion and may play a role in inflammation and fibrosis. The alpha 1 subunit contains an inserted (I) von Willebrand factor type I domain which is thought to be involved in collagen binding. |
| ITGA2 | Integrin, alpha 2 (CD49B, alpha 2 subunit of VLA-2 receptor) | 3673 | 0.52 | 0.015 * | This gene encodes the alpha 2 subunit of a transmembrane receptor for collagens and related proteins. The encoded protein forms a heterodimer with a beta subunit and mediates the adhesion of platelets and other cell types to the ECM. |
| ITGA3 | Integrin, alpha 3 (antigen CD49C, alpha 3 subunit of VLA-3 receptor) | 3675 | 0.50 | 0.010 ** | This gene encodes the alpha 3 subunit of integrin. Integrins are heterodimeric integral membrane proteins that function in cell surface adhesion, cytoskeletal rearrangement and cell signalling. This subunit joins with a beta 1 subunit to form an integrin that interacts with ECM proteins, including members of the laminin family. |
| ITGA4 | Integrin, alpha 4 (antigen CD49D, alpha 4 subunit of VLA-4 receptor) | 3676 | 0.86 | 0.459 | This gene encodes for the alpha 4 subunit of the integrin alpha chain family. This subunit associates with a beta 1 or beta 7 subunit to form an integrin that may play a role in cell motility and migration. |
| ITGA5 | Integrin, alpha 5 (fibronectin receptor, alpha polypeptide) | 3678 | 0.54 | 0.004 ** | This gene encodes for the alpha 5 subunit of the integrin alpha chain family. This subunit associates with the beta 1 subunit to form a fibronectin receptor. |
| ITGA6 | Integrin, alpha 6 | 3655 | 0.76 | 0.012 * | This gene encodes for the alpha 6 subunit of the integrin alpha chain family. This subunit may associate with a beta 1 or beta 4 subunit to form an integrin that interacts with ECM proteins including members of the laminin family. |
| ITGA7 | Integrin, alpha 7 | 3679 | 1.11 | 0.513 | This gene encodes for the alpha 7 subunit of the integrin alpha chain family. This protein functions as a receptor for the basement membrane protein laminin-1. It is mainly expressed in skeletal and cardiac muscles and may be involved in differentiation and migration processes during myogenesis. |
| ITGA8 | Integrin, alpha 8 | 8516 | 2.75 | 0.050 * | This gene encodes for the alpha 8 subunit of the heterodimeric integrin alpha8beta1 protein. The encoded protein is a single-pass type 1 membrane protein. This gene regulates the recruitment of mesenchymal cells into epithelial structures, mediates cell-cell interactions, and regulates neurite outgrowth of sensory and motor neurons. The integrin alpha8beta1 protein plays an important role in wound-healing and organogenesis. |
| ITGAL | Integrin, alpha L (antigen CD11A (p180), lymphocyte function-associated antigen 1; alpha polypeptide) | 3683 | 1.89 | 0.003 ** | This gene encodes the integrin alpha L chain. This I-domain containing alpha integrin combines with the beta 2 chain (ITGB2) to form the integrin lymphocyte function-associated antigen-1 (LFA-1). LFA-1 plays a central role in leukocyte intercellular adhesion through interactions with its ligands, ICAMs 1-3, and also functions in lymphocyte costimulatory signalling. |
| ITGAM | Integrin, alpha M (complement component 3 receptor 3 subunit) | 3684 | 1.77 | 0.065 | This gene encodes the integrin alpha M chain. This I-domain containing alpha integrin combines with the beta 2 chain (ITGB2) to form a leukocyte-specific integrin known as macrophage receptor 1 (‘Mac-1’), or inactivated-C3b (iC3b) receptor 3 (‘CR3’). The alpha M beta 2 integrin is important in the adherence of neutrophils and monocytes to stimulated endothelium, and also in the phagocytosis of complement coated particles. |
| ITGAV | Integrin, alpha V (vitronectin receptor, alpha polypeptide, antigen CD51) | 3685 | 0.50 | 0.002 ** | The product of this gene belongs to the integrin alpha chain family. This subunit associates with beta 1, beta 3, beta 5, beta 6 and beta 8 subunits. The heterodimer consisting of alpha V and beta 3 subunits is also known as the vitronectin receptor. This integrin may regulate angiogenesis. |
| ITGB1 | Integrin, beta 1 (fibronectin receptor, beta polypeptide, antigen CD29 includes MDF2, MSK12) | 3688 | 0.53 | 0.005 ** | The product of this gene belongs to the integrin beta chain family; integrin beta chains combine with multiple different alpha chains to form different integrin heterodimers. The ITGB1 protein product is the integrin beta chain beta 1. Integrin family members are noncovalently associated transmembrane glycoprotein receptors involved in cell adhesion and recognition in a variety of processes including embryogenesis, hemostasis, tissue repair, and immune response. |
| ITGB2 | Integrin, beta 2 (complement component 3 receptor 3 and 4 subunit) | 3689 | 1.58 | 0.083 | The ITGB2 protein product is the integrin beta chain beta 2. The encoded protein plays an important role in immune response. |
| ITGB3 | Integrin, beta 3 (platelet glycoprotein IIIa, antigen CD61) | 3690 | 0.58 | 0.006 ** | The ITGB3 protein product is the integrin beta chain beta 3. Integrin beta 3 is found along with the alpha IIb chain in platelets. |
| ITGB4 | Integrin, beta 4 | 3691 | 1.87 | 0.041 * | This gene encodes the integrin beta 4 subunit, a receptor for the laminins. |
| ITGB5 | Integrin, beta 5 | 3693 | 0.97 | 0.879 | This gene encodes the integrin subunit beta 5, and is involved in adhesion to vitronectin. |
| KAL1 | Kallmann syndrome 1 sequence | 3730 | 0.41 | 0.001 *** | Also known as the ANOS1 gene. The protein encoded for by this gene is a cell surface protein, which is N-glycosylated and may have anti-protease activity. |
| LAMA1 | Laminin, alpha 1 | 284217 | 0.60 | 0.014 * | This gene encodes one of the alpha 1 subunits of laminin. Laminins are a family of ECM glycoproteins that have a heterotrimeric structure consisting of an alpha, beta and gamma chain which are bound to each other by disulphide bonds into a cross-shaped molecule. These proteins make up a major component of the basement membrane and have been implicated in a variety of biological processes including cell adhesion, differentiation, migration, and signalling. |
| LAMA2 | Laminin, alpha 2 | 3908 | 1.17 | 0.396 | This gene encodes the alpha 2 chain, which constitutes one of the subunits of laminin 2 (merosin) and laminin 4 (s-merosin). |
| LAMA3 | Laminin, alpha 3 | 3909 | 2.25 | 0.031 * | This gene encodes the alpha 3 chain of laminins. Laminins are essential for formation and function of the basement membrane. This gene is responsive to several epithelial-mesenchymal regulators including keratinocyte growth factor, epidermal growth factor and insulin-like growth factor. |
| LAMB1 | Laminin, beta 1 | 3912 | 0.95 | 0.609 | This gene encodes the beta chain isoform laminin, beta 1. |
| LAMB3 | Laminin, beta 3 | 3914 | 0.61 | 0.010 ** | The product encoded by this gene is a beta subunit laminin, which together with an alpha and a gamma subunit, forms laminin-5 |
| LAMC1 | Laminin, gamma 1 (formerly LAMB2) | 3915 | 0.54 | 0.008 ** | This gene encodes the gamma chain isoform laminin, gamma 1. Embryos of transgenic mice in which both alleles of the gamma 1 chain gene were inactivated by homologous recombination, lacked basement membranes, indicating that laminin, gamma 1 chain is necessary for laminin heterotrimer assembly. |
| MMP14 | Matrix metallopeptidase 14 (membrane-inserted) | 4323 | 0.44 | 0.016 * | Proteins of the matrix metalloproteinase (MMP) family are involved in the breakdown of ECM in normal physiological processes, such as tissue remodelling. The protein encoded by this gene is a member of the membrane-type MMP (MT-MMP) subfamily; each member of this subfamily contains a potential transmembrane domain suggesting that these proteins are expressed at the cell surface rather than secreted. |
| MMP15 | Matrix metallopeptidase 15 (membrane-inserted) | 4324 | 1.24 | 0.157 | This gene encodes a member of the peptidase M10 family and membrane-type subfamily of MMPs. |
| MMP16 | Matrix metallopeptidase 16 (membrane-inserted) | 4325 | 0.44 | 0.003 ** | Most MMP’s are secreted as inactive pro-proteins which are activated when cleaved by extracellular proteinases. The protein encoded by this gene is a member of the MT-MMP subfamily and activates MMP2 by cleavage. |
| NCAM1 | Neural cell adhesion molecule 1 | 4684 | 0.97 | 0.811 | This gene encodes a cell adhesion protein which is a member of the immunoglobulin superfamily. The encoded protein is involved in cell-to-cell interactions as well as cell-matrix interactions during development and differentiation. The encoded protein has been shown to be involved in development of the nervous system, and for cells involved in the expansion of T cells and dendritic cells which play an important role in immune surveillance. |
| PECAM1 | Platelet/endothelial cell adhesion molecule | 5175 | 3.66 | 0.213 | The protein encoded by this gene is found on the surface of platelets, monocytes, neutrophils, and some types of T-cells, and makes up a large portion of endothelial cell intercellular junctions. The encoded protein is a member of the immunoglobulin superfamily and is likely involved in leukocyte migration, angiogenesis, and integrin activation. |
| SELE | Selectin E | 6401 | 1.50 | 0.078 | The protein encoded by this gene is part of the selectin family of cell adhesion molecules, and is found in cytokine-stimulated endothelial cells and is thought to be responsible for the accumulation of blood leukocytes at sites of inflammation by mediating the adhesion of cells to the vascular lining. |
| SELL | Selectin L | 6402 | 2.36 | 0.013 * | This gene encodes a cell surface adhesion molecule that belongs to a family of adhesion/homing receptors. |
| SELP | Selectin P (granule membrane protein 140 kDa, antigen CD62) | 6403 | 1.90 | 0.249 | This protein redistributes to the plasma membrane during platelet activation and degranulation and mediates the interaction of activated endothelial cells or platelets with leukocytes. |
| SGCE | Sarcoglycan, epsilon | 8910 | 0.93 | 0.657 | This gene encodes the epsilon member of the sarcoglycan family. Sarcoglycans are transmembrane proteins that are components of the dystrophin-glycoprotein complex, which link the actin cytoskeleton to the ECM. |
| SPG7 | Spastic paraplegia 7 (pure and complicated autosomal recessive) | 6687 | 0.48 | 0.005 ** | This gene encodes a mitochondrial metalloprotease protein that is a member of the AAA family. Members of this protein family share an ATPase domain and have roles in diverse cellular processes including membrane trafficking, intracellular motility, organelle biogenesis, protein folding, and proteolysis. |
| SPP1 | Secreted phosphoprotein 1 | 6696 | 0.58 | 0.009 ** | The protein encoded by this gene is involved in the attachment of osteoclasts to the mineralized bone matrix. This protein is also a cytokine that upregulates expression of interferon-gamma and interleukin-12. |
| THBS1 | Thrombospondin 1 | 7057 | 0.35 | 0.001 *** | The protein encoded by this gene is a subunit of a disulphide-linked homotrimeric protein, and belongs to the thrombospondin family. This protein is an adhesive glycoprotein that mediates cell-to-cell and cell-to-matrix interactions. This protein can bind to fibrinogen, fibronectin, laminin, type V collagen and integrins alpha-V/beta-1. |
| THBS2 | Thrombospondin 2 | 7058 | 0.81 | 0.088 | The protein encoded by this gene belongs to the thrombo-spondin family. |
| THBS3 | Thrombospondin 3 | 7059 | 1.03 | 0.794 | The protein encoded by this gene belongs to the thrombo-spondin family. |
| TNC | Tenascin C | 3371 | 1.18 | 0.492 | This gene encodes for an ECM protein. |
| VCAM1 | Vascular cell adhesion molecule 1 | 7412 | 1.79 | 0.002 ** | This gene is a member of the Ig superfamily and encodes a cell surface sialoglycoprotein expressed by cytokine-activated endothelium. This type I membrane protein mediates leukocyte-endothelial cell adhesion and signal transduction. |
| VCAN | Versican | 1462 | 0.96 | 0.775 | This gene is a member of the aggrecan/versican proteoglycan family. The protein encoded is a large chondroitin sulphate proteoglycan and is a major component of the ECM. This protein is involved in cell adhesion, proliferation, migration and angiogenesis and plays a central role in tissue morphogenesis and maintenance. |
| VTN | Vitronectin | 7448 | 1.20 | 0.240 | The protein encoded by this gene is a member of the pexin family. It is found in serum and tissues and promotes cell adhesion and spreading, inhibits the membrane-damaging effect of the terminal cytolytic complement pathway, and binds to several serpin serine protease inhibitors. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Houreld, N.N.; Ayuk, S.M.; Abrahamse, H. Cell Adhesion Molecules Are Mediated by Photobiomodulation at 660 nm in Diabetic Wounded Fibroblast Cells. Cells 2018, 7, 30. https://doi.org/10.3390/cells7040030
Houreld NN, Ayuk SM, Abrahamse H. Cell Adhesion Molecules Are Mediated by Photobiomodulation at 660 nm in Diabetic Wounded Fibroblast Cells. Cells. 2018; 7(4):30. https://doi.org/10.3390/cells7040030
Chicago/Turabian StyleHoureld, Nicolette N., Sandra M. Ayuk, and Heidi Abrahamse. 2018. "Cell Adhesion Molecules Are Mediated by Photobiomodulation at 660 nm in Diabetic Wounded Fibroblast Cells" Cells 7, no. 4: 30. https://doi.org/10.3390/cells7040030
APA StyleHoureld, N. N., Ayuk, S. M., & Abrahamse, H. (2018). Cell Adhesion Molecules Are Mediated by Photobiomodulation at 660 nm in Diabetic Wounded Fibroblast Cells. Cells, 7(4), 30. https://doi.org/10.3390/cells7040030