.png)

A Theoretical Hypothesis on How Immune Cells May Transmit Acquired Traits: A Macrophage–piRNA Pathway for Transgenerational Inheritance


Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods
3. Results
3.1. piRNAs as Mediators of Epigenetic Inheritance
3.1.1. Canonical Functions of piRNAs in Genome Defense
3.1.2. Transgenerational piRNA Inheritance in C. elegans
3.1.3. Small-RNA Inheritance Systems in Plants and Other Eukaryotes
3.2. Environmental Stress, Hsp90, and Transposon Activation
3.2.1. Hsp90 as a Capacitor for Phenotypic Variation
3.2.2. Environmental Stress and Small-RNA Responses
3.3. Macrophages as Regulators of Tissue Remodeling and Stress Responses
3.3.1. Evolutionary Origins of Macrophage-Like Cells
3.3.2. Macrophage Polarization and Functional Plasticity
3.4. Macrophage Extracellular Vesicles Containing Small RNAs
3.5. The ELBO Phenotype as a Model for Stress-Induced Morphological Inheritance
The ELBO Phenotype in Drosophila as a Model of Stress-Induced Morphological Plasticity

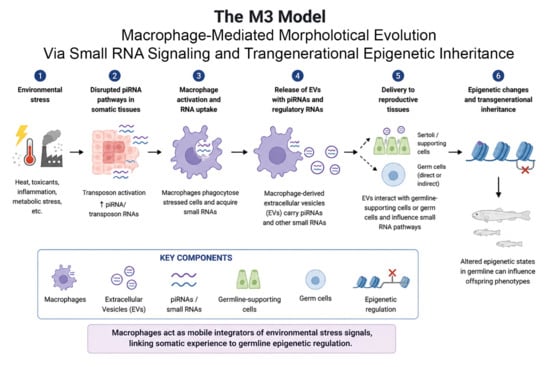
3.6. The Macrophage-Mediated Morphological (M3) Evolution Model
3.6.1. Proposed Mechanism of Macrophage-Mediated Inheritance
3.6.2. Experimental Predictions of the M3 Model
3.6.3. Limitations and Evolutionary Constraints of the M3 Model
4. Discussion and Summary
5. Testable Hypotheses, Alternative Mechanisms, and Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ELBO | Ectopic large bristle outgrowth (epigenetic phenotype in Drosophila; aka EBO) |
| Hsp90 | Heat-shock protein 90 |
| RdRP | RND dependent RNA polymerase |
| PIWI | P-element-induced WImpy testis |
| piRNA | PIWI RNA |
| miRNA | Micro RNA |
| siRNA | Small interfering RNA |
| piRISC | piRNA PIWI complex |
| M1 | M1 macrophage (generally pro-inflammatory) |
| M2 | M2 macrophage (generally anti-inflammatory/pro-fibrotic) |
| M3 | Macrophage-mediated morphological evolution model |
| tRF | tRNA fragment |
| RDDM | RNA-directed DNA methylation |
| RNAe | RNA-induced epigenetic silencing |
| H3K9me | Histone 3 lysine 9 methylation (repressive mark) |
| scnRNA | Scan RNA (ciliates use these for somatic nucleus DNA elimination) |
| LPS | Lipopolysaccharide |
| EV | Extracellular vesicle |
| Kr | Krueppel (Drosophila segmentation gene) |
References
- McLaren, A. Too late for the midwife toad: Stress, variability and Hsp90. Trends Genet. 1999, 15, 169–171. [Google Scholar] [CrossRef]
- Huxley, J. Evolution, the Modern Synthesis; Harper & Brothers: New York, NY, USA; London, UK, 1943. [Google Scholar]
- Dobzhansky, T. Genetics and the Origin of Species; Columbia University Press: New York, NY, USA, 1937. [Google Scholar]
- Gould, S.J. The Structure of Evolutionary Theory; Belknap Press of Harvard University Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Lee, Y.H.; Petkova, A.P.; Granneman, J.G. Identification of an adipogenic niche for adipose tissue remodeling and restoration. Cell Metab. 2013, 18, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Korolenko, A.A.; Nilsson, E.E.; De Santos, S.; Skinner, M.K. Generational stability of environmentally induced epigenetic transgenerational inheritance of adult-onset disease over ten mammalian generations. Environ. Epigenet. 2025, 11, dvaf033. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.E.; Sadler-Riggleman, I.; Skinner, M.K. Environmentally induced epigenetic transgenerational inheritance of disease. Environ. Epigenet. 2018, 4, dvy016. [Google Scholar] [CrossRef] [PubMed]
- Crisostomo, L.; Oliveira, P.F.; Alves, M.G. A systematic scientometric review of paternal inheritance of acquired metabolic traits. BMC Biol. 2023, 21, 255. [Google Scholar] [CrossRef]
- Lismer, A.; Kimmins, S. Emerging evidence that the mammalian sperm epigenome serves as a template for embryo development. Nat. Commun. 2023, 14, 2142. [Google Scholar] [CrossRef]
- Jawaid, A.; Jehle, K.L.; Mansuy, I.M. Impact of Parental Exposure on Offspring Health in Humans. Trends Genet. 2021, 37, 373–388. [Google Scholar] [CrossRef]
- Champroux, A.; Cocquet, J.; Henry-Berger, J.; Drevet, J.R.; Kocer, A. A Decade of Exploring the Mammalian Sperm Epigenome: Paternal Epigenetic and Transgenerational Inheritance. Front. Cell Dev. Biol. 2018, 6, 50. [Google Scholar] [CrossRef]
- Larriba, E.; del Mazo, J. Role of Non-Coding RNAs in the Transgenerational Epigenetic Transmission of the Effects of Reprotoxicants. Int. J. Mol. Sci. 2016, 17, 452. [Google Scholar] [CrossRef]
- Frias-Lasserre, D.; Villagra, C.A. The Importance of ncRNAs as Epigenetic Mechanisms in Phenotypic Variation and Organic Evolution. Front. Microbiol. 2017, 8, 2483. [Google Scholar] [CrossRef]
- Shang, R.; Lee, S.; Senavirathne, G.; Lai, E.C. microRNAs in action: Biogenesis, function and regulation. Nat. Rev. Genet. 2023, 24, 816–833. [Google Scholar] [CrossRef]
- Ow, M.C.; Hall, S.E. Inheritance of Stress Responses via Small Non-Coding RNAs in Invertebrates and Mammals. Epigenomes 2023, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Yap, Y.S.; Patrizio, P.; Cimmino, L.; Sdrimas, K.; Telonis, A.G. The Small Non-Coding RNA Profile of Human and Mouse Sperm. Noncoding RNA 2025, 11, 15. [Google Scholar] [CrossRef]
- Hernandez, R.; Shi, J.; Liu, J.; Li, X.; Wu, J.; Zhao, L.; Zhou, T.; Chen, Q.; Zhou, C. PANDORA-Seq unveils the hidden small noncoding RNA landscape in atherosclerosis of LDL receptor-deficient mice. J. Lipid Res. 2023, 64, 100352. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Yang, Y.; Jiang, W.; Cao, Z.; Shi, J.; Zhang, X.O.; Zhang, Y. PANDORA-seq reveals human sperm sncRNA signature endowed with sperm quality assessment. Genes. Dis. 2026, 13, 101807. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, Y.; Tan, D.; Zhang, X.; Yan, M.; Zhang, Y.; Franklin, R.; Shahbazi, M.; Mackinlay, K.; Liu, S.; et al. PANDORA-seq expands the repertoire of regulatory small RNAs by overcoming RNA modifications. Nat. Cell Biol. 2021, 23, 424–436, Erratum in Nat. Cell Biol. 2021, 23, 676. https://doi.org/10.1038/s41556-021-00687-w. [Google Scholar] [CrossRef]
- Le Thomas, A.; Stuwe, E.; Li, S.; Du, J.; Marinov, G.; Rozhkov, N.; Chen, Y.C.; Luo, Y.; Sachidanandam, R.; Toth, K.F.; et al. Transgenerationally inherited piRNAs trigger piRNA biogenesis by changing the chromatin of piRNA clusters and inducing precursor processing. Genes. Dev. 2014, 28, 1667–1680. [Google Scholar] [CrossRef]
- Czech, B.; Munafo, M.; Ciabrelli, F.; Eastwood, E.L.; Fabry, M.H.; Kneuss, E.; Hannon, G.J. piRNA-Guided Genome Defense: From Biogenesis to Silencing. Annu. Rev. Genet. 2018, 52, 131–157. [Google Scholar] [CrossRef]
- Wu, Y.; Imai, R.; Kosako, H.; Nakahara, T.; Solberg, T.; Siomi, H.; Murano, K. The Piwi-piRNA complex initiates transposon silencing via transcription termination factors PNUTS and Senataxin. Mol. Cell 2025, 85, 3999–4015.e8. [Google Scholar] [CrossRef]
- Czech, B.; Hannon, G.J. One Loop to Rule Them All: The Ping-Pong Cycle and piRNA-Guided Silencing. Trends Biochem. Sci. 2016, 41, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Pastore, B.; Hertz, H.L.; Price, I.F.; Tang, W. pre-piRNA trimming and 2′-O-methylation protect piRNAs from 3′ tailing and degradation in C. elegans. Cell Rep. 2021, 36, 109640. [Google Scholar] [CrossRef]
- Webster, A.; Li, S.; Hur, J.K.; Wachsmuth, M.; Bois, J.S.; Perkins, E.M.; Patel, D.J.; Aravin, A.A. Aub and Ago3 Are Recruited to Nuage through Two Mechanisms to Form a Ping-Pong Complex Assembled by Krimper. Mol. Cell 2015, 59, 564–575. [Google Scholar] [CrossRef]
- Weiser, N.E.; Kim, J.K. Multigenerational Regulation of the Caenorhabditis elegans Chromatin Landscape by Germline Small RNAs. Annu. Rev. Genet. 2019, 53, 289–311. [Google Scholar] [CrossRef]
- Ashe, A.; Sapetschnig, A.; Weick, E.M.; Mitchell, J.; Bagijn, M.P.; Cording, A.C.; Doebley, A.L.; Goldstein, L.D.; Lehrbach, N.J.; Le Pen, J.; et al. piRNAs can trigger a multigenerational epigenetic memory in the germline of C. elegans. Cell 2012, 150, 88–99. [Google Scholar] [CrossRef]
- Shirayama, M.; Seth, M.; Lee, H.C.; Gu, W.; Ishidate, T.; Conte, D., Jr.; Mello, C.C. piRNAs initiate an epigenetic memory of nonself RNA in the C. elegans germline. Cell 2012, 150, 65–77. [Google Scholar] [CrossRef]
- Zhang, Q.; Dang, W.; Wang, M.C. Lysosomes signal through the epigenome to regulate longevity across generations. Science 2025, 389, 1353–1360. [Google Scholar] [CrossRef]
- Erdmann, R.M.; Picard, C.L. RNA-directed DNA Methylation. PLoS Genet. 2020, 16, e1009034. [Google Scholar] [CrossRef] [PubMed]
- Neeb, Z.T.; Nowacki, M. RNA-mediated transgenerational inheritance in ciliates and plants. Chromosoma 2018, 127, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Chalker, D.L.; Meyer, E.; Mochizuki, K. Epigenetics of ciliates. Cold Spring Harb. Perspect. Biol. 2013, 5, a017764. [Google Scholar] [CrossRef] [PubMed]
- Pilling, O.A.; Rogers, A.J.; Gulla-Devaney, B.; Katz, L.A. Insights into transgenerational epigenetics from studies of ciliates. Eur. J. Protistol. 2017, 61, 366–375. [Google Scholar] [CrossRef]
- Ali, W.; Deng, K.; Bian, Y.; Liu, Z.; Zou, H. Spectacular role of epididymis and bio-active cargo of nano-scale exosome in sperm maturation: A review. Biomed. Pharmacother. 2023, 164, 114889. [Google Scholar] [CrossRef]
- Estill, M.; Hauser, R.; Nassan, F.L.; Moss, A.; Krawetz, S.A. The effects of di-butyl phthalate exposure from medications on human sperm RNA among men. Sci. Rep. 2019, 9, 12397. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, D.L.; Petriello, M.C.; Pilsner, J.R. PFAS Exposure and Male Reproductive Health: Implications for Sperm Epigenetics. Semin. Reprod. Med. 2024, 42, 288–301. [Google Scholar] [CrossRef]
- Sharma, U. Paternal Contributions to Offspring Health: Role of Sperm Small RNAs in Intergenerational Transmission of Epigenetic Information. Front. Cell Dev. Biol. 2019, 7, 215. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.M.; Li, C.J.; Vagbo, C.B.; Shi, Y.; Wang, W.L.; Song, S.H.; et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef]
- Gapp, K.; Miska, E.A. tRNA fragments: Novel players in intergenerational inheritance. Cell Res. 2016, 26, 395–396. [Google Scholar] [CrossRef]
- Pantazi, P.; Clements, T.; Veno, M.; Abrahams, V.M.; Holder, B. Distinct non-coding RNA cargo of extracellular vesicles from M1 and M2 human primary macrophages. J. Extracell. Vesicles 2022, 11, e12293. [Google Scholar] [CrossRef] [PubMed]
- Sharma, U.; Conine, C.C.; Shea, J.M.; Boskovic, A.; Derr, A.G.; Bing, X.Y.; Belleannee, C.; Kucukural, A.; Serra, R.W.; Sun, F.; et al. Biogenesis and function of tRNA fragments during sperm maturation and fertilization in mammals. Science 2016, 351, 391–396. [Google Scholar] [CrossRef]
- Park, J.; Ahn, S.H.; Shin, M.G.; Kim, H.K.; Chang, S. tRNA-Derived Small RNAs: Novel Epigenetic Regulators. Cancers 2020, 12, 2773. [Google Scholar] [CrossRef]
- Casier, K.; Boivin, A.; Carre, C.; Teysset, L. Environmentally-Induced Transgenerational Epigenetic Inheritance: Implication of PIWI Interacting RNAs. Cells 2019, 8, 1108. [Google Scholar] [CrossRef]
- Kim, S.Y.; Nair, M.G. Macrophages in wound healing: Activation and plasticity. Immunol. Cell Biol. 2019, 97, 258–267. [Google Scholar] [CrossRef]
- Krzyszczyk, P.; Schloss, R.; Palmer, A.; Berthiaume, F. The Role of Macrophages in Acute and Chronic Wound Healing and Interventions to Promote Pro-wound Healing Phenotypes. Front. Physiol. 2018, 9, 419. [Google Scholar] [CrossRef] [PubMed]
- Strizova, Z.; Benesova, I.; Bartolini, R.; Novysedlak, R.; Cecrdlova, E.; Foley, L.K.; Striz, I. M1/M2 macrophages and their overlaps-myth or reality? Clin. Sci. 2023, 137, 1067–1093. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Zhou, X.; Wang, Y.; Dai, L.; Yuan, J.; Peng, J.; Zhang, X.; Wang, C. Changes in the Small Noncoding RNAome During M1 and M2 Macrophage Polarization. Front. Immunol. 2022, 13, 799733. [Google Scholar] [CrossRef]
- Liu, K.; Peng, J.; Chen, K.; Han, X.; Ma, D.; Liao, X.; Chen, X.; Ding, Q.; Xiao, F. DQ690018 regulated HIF1alpha expression and subsequently mediates macrophage polarization by inhibiting Taf11 mRNA 2′-O-methylation modification. Int. Immunopharmacol. 2026, 168, 115849. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Yu, X.; Zhang, S.; He, Y.; Guo, W. Novel roles of PIWI proteins and PIWI-interacting RNAs in human health and diseases. Cell Commun. Signal. 2023, 21, 343. [Google Scholar] [CrossRef]
- Jo, H.Y.; Kim, M.K.; Kim, K.T.; Choi, C.; Rhee, W.J. Extracellular vesicles for macrophage reprogramming: An emerging paradigm in immunomodulatory therapeutics. J. Biol. Eng. 2025, 20, 13. [Google Scholar] [CrossRef]
- Kumar, M.A.; Baba, S.K.; Sadida, H.Q.; Marzooqi, S.A.; Jerobin, J.; Altemani, F.H.; Algehainy, N.; Alanazi, M.A.; Abou-Samra, A.B.; Kumar, R.; et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct. Target. Ther. 2024, 9, 27. [Google Scholar] [CrossRef]
- Hu, Q.; Lyon, C.J.; Fletcher, J.K.; Tang, W.; Wan, M.; Hu, T.Y. Extracellular vesicle activities regulating macrophage- and tissue-mediated injury and repair responses. Acta Pharm. Sin. B 2021, 11, 1493–1512. [Google Scholar] [CrossRef]
- Gangaraju, V.K.; Yin, H.; Weiner, M.M.; Wang, J.; Huang, X.A.; Lin, H. Drosophila Piwi functions in Hsp90-mediated suppression of phenotypic variation. Nat. Genet. 2011, 43, 153–158. [Google Scholar] [CrossRef]
- Ruden, D.M. The (new) new synthesis and epigenetic capacitors of morphological evolution. Nat. Genet. 2011, 43, 88–89. [Google Scholar] [CrossRef]
- Ruden, D.M.; Garfinkel, M.D.; Sollars, V.E.; Lu, X. Waddington’s widget: Hsp90 and the inheritance of acquired characters. Semin. Cell Dev. Biol. 2003, 14, 301–310. [Google Scholar] [CrossRef]
- Sollars, V.; Lu, X.; Xiao, L.; Wang, X.; Garfinkel, M.D.; Ruden, D.M. Evidence for an epigenetic mechanism by which Hsp90 acts as a capacitor for morphological evolution. Nat. Genet. 2003, 33, 70–74. [Google Scholar] [CrossRef]
- Ku, H.Y.; Lin, H. PIWI proteins and their interactors in piRNA biogenesis, germline development and gene expression. Natl. Sci. Rev. 2014, 1, 205–218. [Google Scholar] [CrossRef]
- Huang, S.; Yoshitake, K.; Asakawa, S. A Review of Discovery Profiling of PIWI-Interacting RNAs and Their Diverse Functions in Metazoans. Int. J. Mol. Sci. 2021, 22, 11166. [Google Scholar] [CrossRef] [PubMed]
- Gainetdinov, I.; Vega-Badillo, J.; Cecchini, K.; Bagci, A.; Colpan, C.; De, D.; Bailey, S.; Arif, A.; Wu, P.H.; MacRae, I.J.; et al. Relaxed targeting rules help PIWI proteins silence transposons. Nature 2023, 619, 394–402. [Google Scholar] [CrossRef]
- Ernst, C.; Odom, D.T.; Kutter, C. The emergence of piRNAs against transposon invasion to preserve mammalian genome integrity. Nat. Commun. 2017, 8, 1411. [Google Scholar] [CrossRef]
- Zawalska, M.; Tarnowski, M. piRNA-Mediated Maintenance of Genome Stability in Gametogenesis and Cancer. Genes 2025, 16, 722. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.D.; Jiang, H.; Zhang, Y.F.; Zhang, Y.; Qian, L.L.; Zhang, Y.F. The regulatory function of piRNA/PIWI complex in cancer and other human diseases: The role of DNA methylation. Int. J. Biol. Sci. 2022, 18, 3358–3373. [Google Scholar] [CrossRef] [PubMed]
- Pezic, D.; Manakov, S.A.; Sachidanandam, R.; Aravin, A.A. piRNA pathway targets active LINE1 elements to establish the repressive H3K9me3 mark in germ cells. Genes. Dev. 2014, 28, 1410–1428. [Google Scholar] [CrossRef]
- Greenberg, M.V.C.; Bourc’his, D. The diverse roles of DNA methylation in mammalian development and disease. Nat. Rev. Mol. Cell Biol. 2019, 20, 590–607. [Google Scholar] [CrossRef]
- Walter, M.; Teissandier, A.; Perez-Palacios, R.; Bourc’his, D. An epigenetic switch ensures transposon repression upon dynamic loss of DNA methylation in embryonic stem cells. eLife 2016, 5, e11418. [Google Scholar] [CrossRef]
- Watanabe, T.; Lin, H. Posttranscriptional regulation of gene expression by Piwi proteins and piRNAs. Mol. Cell 2014, 56, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 2007, 128, 1089–1103. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhu, Y.; Cao, X.; Tan, W.; Yu, J.; Lu, Y.; Kang, R.; Wang, X.; Li, E. The epigenetic regulatory mechanism of PIWI/piRNAs in human cancers. Mol. Cancer 2023, 22, 45. [Google Scholar] [CrossRef] [PubMed]
- Seth, M.; Shirayama, M.; Gu, W.; Ishidate, T.; Conte, D., Jr.; Mello, C.C. The C. elegans CSR-1 argonaute pathway counteracts epigenetic silencing to promote germline gene expression. Dev. Cell 2013, 27, 656–663. [Google Scholar] [CrossRef]
- Kasper, D.M.; Gardner, K.E.; Reinke, V. Homeland security in the C. elegans germ line: Insights into the biogenesis and function of piRNAs. Epigenetics 2014, 9, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Stoeckius, M.; Grun, D.; Rajewsky, N. Paternal RNA contributions in the Caenorhabditis elegans zygote. EMBO J. 2014, 33, 1740–1750. [Google Scholar] [CrossRef]
- Minkina, O.; Hunter, C.P. Stable Heritable Germline Silencing Directs Somatic Silencing at an Endogenous Locus. Mol. Cell 2017, 65, 659–670.e5. [Google Scholar] [CrossRef]
- Svendsen, J.M.; Reed, K.J.; Vijayasarathy, T.; Montgomery, B.E.; Tucci, R.M.; Brown, K.C.; Marks, T.N.; Nguyen, D.A.H.; Phillips, C.M.; Montgomery, T.A. henn-1/HEN1 Promotes Germline Immortality in Caenorhabditis elegans. Cell Rep. 2019, 29, 3187–3199.e4. [Google Scholar] [CrossRef]
- Barucci, G.; Cornes, E.; Singh, M.; Li, B.; Ugolini, M.; Samolygo, A.; Didier, C.; Dingli, F.; Loew, D.; Quarato, P.; et al. Small-RNA-mediated transgenerational silencing of histone genes impairs fertility in piRNA mutants. Nat. Cell Biol. 2020, 22, 235–245. [Google Scholar] [CrossRef]
- Frolows, N.; Ashe, A. Small RNAs and chromatin in the multigenerational epigenetic landscape of Caenorhabditis elegans. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20200112. [Google Scholar] [CrossRef] [PubMed]
- Grishok, A. Small RNAs Worm Up Transgenerational Epigenetics Research. DNA 2021, 1, 37–48. [Google Scholar] [CrossRef]
- Filipowicz, A.; Allard, P. Caenorhabditis elegans as a Model for Environmental Epigenetics. Curr. Environ. Health Rep. 2025, 12, 6. [Google Scholar] [CrossRef]
- Batista, P.J.; Ruby, J.G.; Claycomb, J.M.; Chiang, R.; Fahlgren, N.; Kasschau, K.D.; Chaves, D.A.; Gu, W.; Vasale, J.J.; Duan, S.; et al. PRG-1 and 21U-RNAs interact to form the piRNA complex required for fertility in C. elegans. Mol. Cell 2008, 31, 67–78. [Google Scholar] [CrossRef]
- Pastore, B.; Hertz, H.L.; Tang, W. Pre-piRNA trimming safeguards piRNAs against erroneous targeting by RNA-dependent RNA polymerase. Cell Rep. 2024, 43, 113692. [Google Scholar] [CrossRef] [PubMed]
- Shen, E.Z.; Chen, H.; Ozturk, A.R.; Tu, S.; Shirayama, M.; Tang, W.; Ding, Y.H.; Dai, S.Y.; Weng, Z.; Mello, C.C. Identification of piRNA Binding Sites Reveals the Argonaute Regulatory Landscape of the C. elegans Germline. Cell 2018, 172, 937–951.e18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tu, S.; Stubna, M.; Wu, W.S.; Huang, W.C.; Weng, Z.; Lee, H.C. The piRNA targeting rules and the resistance to piRNA silencing in endogenous genes. Science 2018, 359, 587–592. [Google Scholar] [CrossRef]
- Placentino, M.; de Jesus Domingues, A.M.; Schreier, J.; Dietz, S.; Hellmann, S.; de Albuquerque, B.F.; Butter, F.; Ketting, R.F. Intrinsically disordered protein PID-2 modulates Z granules and is required for heritable piRNA-induced silencing in the Caenorhabditis elegans embryo. EMBO J. 2021, 40, e105280. [Google Scholar] [CrossRef]
- Cornes, E.; Bourdon, L.; Singh, M.; Mueller, F.; Quarato, P.; Wernersson, E.; Bienko, M.; Li, B.; Cecere, G. piRNAs initiate transcriptional silencing of spermatogenic genes during C. elegans germline development. Dev. Cell 2022, 57, 180–196.e7. [Google Scholar] [CrossRef]
- Baduel, P.; De Oliveira, L.; Caillieux, E.; Bohl-Viallefond, G.; Xu, C.; El Messaoudi, M.; Petit, A.; Drai, M.; Barois, M.; Singh, V.; et al. Transposable elements are vectors of recurrent transgenerational epigenetic inheritance. Science 2025, 390, eady3475. [Google Scholar] [CrossRef]
- Bond, D.M.; Baulcombe, D.C. Small RNAs and heritable epigenetic variation in plants. Trends Cell Biol. 2014, 24, 100–107. [Google Scholar] [CrossRef]
- Cao, S.; Chen, Z.J. Transgenerational epigenetic inheritance during plant evolution and breeding. Trends Plant Sci. 2024, 29, 1203–1223. [Google Scholar] [CrossRef]
- Cao, S.; Wang, L.; Han, T.; Ye, W.; Liu, Y.; Sun, Y.; Moose, S.P.; Song, Q.; Chen, Z.J. Small RNAs mediate transgenerational inheritance of genome-wide trans-acting epialleles in maize. Genome Biol. 2022, 23, 53. [Google Scholar] [CrossRef] [PubMed]
- Cheong, J.J. Transgenerational Transmission of Epigenetic Marks During Reproduction in Arabidopsis. Yale J. Biol. Med. 2025, 98, 245–251. [Google Scholar] [CrossRef]
- Erhard, K.F., Jr.; Parkinson, S.E.; Gross, S.M.; Barbour, J.E.; Lim, J.P.; Hollick, J.B. Maize RNA polymerase IV defines trans-generational epigenetic variation. Plant Cell 2013, 25, 808–819. [Google Scholar] [CrossRef]
- Fei, Y.; Nyiko, T.; Molnar, A. Non-perfectly matching small RNAs can induce stable and heritable epigenetic modifications and can be used as molecular markers to trace the origin and fate of silencing RNAs. Nucleic Acids Res. 2021, 49, 1900–1913. [Google Scholar] [CrossRef]
- Karalija, E.; Ibragic, S.; Dahija, S.; Samec, D. Transgenerational Memory of Phenotypic Traits in Plants: Epigenetic Regulation of Growth, Hormonal Balance, and Stress Adaptation. Curr. Issues Mol. Biol. 2025, 47, 404. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, W.; Kobayashi, A.; Kato, A.; Ito, H. The effects of heat induction and the siRNA biogenesis pathway on the transgenerational transposition of ONSEN, a copia-like retrotransposon in Arabidopsis thaliana. Plant Cell Physiol. 2012, 53, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Pachamuthu, K.; Simon, M.; Borges, F. Targeted suppression of siRNA biogenesis in Arabidopsis pollen promotes triploid seed viability. Nat. Commun. 2024, 15, 4612. [Google Scholar] [CrossRef]
- Quarato, P.; Singh, M.; Bourdon, L.; Cecere, G. Inheritance and maintenance of small RNA-mediated epigenetic effects. Bioessays 2022, 44, e2100284. [Google Scholar] [CrossRef] [PubMed]
- Sintaha, M. Molecular Mechanisms of Plant Stress Memory: Roles of Non-Coding RNAs and Alternative Splicing. Plants 2025, 14, 2021. [Google Scholar] [CrossRef] [PubMed]
- Toker, I.A.; Lev, I.; Mor, Y.; Gurevich, Y.; Fisher, D.; Houri-Zeevi, L.; Antonova, O.; Doron, H.; Anava, S.; Gingold, H.; et al. Transgenerational inheritance of sexual attractiveness via small RNAs enhances evolvability in C. elegans. Dev. Cell 2022, 57, 298–309.e9. [Google Scholar] [CrossRef]
- John, U.P.; Spangenberg, G.C. Xenogenomics: Genomic bioprospecting in indigenous and exotic plants through EST discovery, cDNA microarray-based expression profiling and functional genomics. Comp. Funct. Genom. 2005, 6, 230–235. [Google Scholar] [CrossRef]
- Youngson, N.A.; Lin, P.C.; Lin, S.S. The convergence of autophagy, small RNA and the stress response-implications for transgenerational epigenetic inheritance in plants. Biomol. Concepts 2014, 5, 1–8. [Google Scholar] [CrossRef]
- Chang, S.S.; Zhang, Z.; Liu, Y. RNA interference pathways in fungi: Mechanisms and functions. Annu. Rev. Microbiol. 2012, 66, 305–323. [Google Scholar] [CrossRef]
- Grypioti, E.; Richard, H.; Kryovrysanaki, N.; Jaubert, M.; Falciatore, A.; Verret, F.; Kalantidis, K. Dicer-dependent heterochromatic small RNAs in the model diatom species Phaeodactylum tricornutum. New Phytol. 2024, 241, 811–826. [Google Scholar] [CrossRef]
- Holoch, D.; Moazed, D. RNA-mediated epigenetic regulation of gene expression. Nat. Rev. Genet. 2015, 16, 71–84. [Google Scholar] [CrossRef]
- Colonna Romano, N.; Fanti, L. Transposable Elements: Major Players in Shaping Genomic and Evolutionary Patterns. Cells 2022, 11, 1048. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.; Ghatage, T.; Bhan, S.; Lahane, G.P.; Dhar, A.; Kumar, R.; Pandita, R.K.; Bhat, K.M.; Ramos, K.S.; Pandita, T.K. Role of Transposable Elements in Genome Stability: Implications for Health and Disease. Int. J. Mol. Sci. 2022, 23, 7802. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Feder, M.E.; Kang, L. Evolution of heat-shock protein expression underlying adaptive responses to environmental stress. Mol. Ecol. 2018, 27, 3040–3054. [Google Scholar] [CrossRef] [PubMed]
- Coculla, A.; Feldmann, C.; Ogueta, M.; Mews, S.; Langrock, R.; Stanewsky, R. Hsp90 buffers behavioral variability by regulating Pdf transcription in clock neurons of Drosophila melanogaster. PLoS Genet. 2026, 22, e1012044. [Google Scholar] [CrossRef]
- Hummel, B.; Hansen, E.C.; Yoveva, A.; Aprile-Garcia, F.; Hussong, R.; Sawarkar, R. The evolutionary capacitor HSP90 buffers the regulatory effects of mammalian endogenous retroviruses. Nat. Struct. Mol. Biol. 2017, 24, 234–242. [Google Scholar] [CrossRef]
- Hung, H.C.; Kay, S.A.; Weber, F. HSP90, a capacitor of behavioral variation. J. Biol. Rhythm. 2009, 24, 183–192. [Google Scholar] [CrossRef]
- Jarosz, D. Hsp90: A Global Regulator of the Genotype-to-Phenotype Map in Cancers. Adv. Cancer Res. 2016, 129, 225–247. [Google Scholar]
- Lachowiec, J.; Lemus, T.; Borenstein, E.; Queitsch, C. Hsp90 promotes kinase evolution. Mol. Biol. Evol. 2015, 32, 91–99. [Google Scholar] [CrossRef]
- Lemus, T.; Mason, G.A.; Bubb, K.L.; Alexandre, C.M.; Queitsch, C.; Cuperus, J.T. AGO1 and HSP90 buffer different genetic variants in Arabidopsis thaliana. Genetics 2023, 223, iyac163. [Google Scholar] [CrossRef] [PubMed]
- Queitsch, C.; Sangster, T.A.; Lindquist, S. Hsp90 as a capacitor of phenotypic variation. Nature 2002, 417, 618–624. [Google Scholar] [CrossRef]
- Ruden, D.M.; Xiao, L.; Garfinkel, M.D.; Lu, X. Hsp90 and environmental impacts on epigenetic states: A model for the trans-generational effects of diethylstibesterol on uterine development and cancer. Hum. Mol. Genet. 2005, 14, R149–R155. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.; Hirate, Y.; Swalla, B.J. The Hsp90 capacitor, developmental remodeling, and evolution: The robustness of gene networks and the curious evolvability of metamorphosis. Crit. Rev. Biochem. Mol. Biol. 2007, 42, 355–372. [Google Scholar] [CrossRef]
- Rutherford, S.L.; Lindquist, S. Hsp90 as a capacitor for morphological evolution. Nature 1998, 396, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Sawarkar, R.; Paro, R. Heat shock protein 90: A capacitor or a mutator? J. Biosci. 2010, 35, 163–165. [Google Scholar] [CrossRef]
- Sayed, R.; Sahin, O.; Errbii, M.; R, R.; Peuss, R.; Pruser, T.; Schrader, L.; Schulz, N.K.E.; Kurtz, J. HSP90 as an evolutionary capacitor drives adaptive eye size reduction via atonal. Nat. Commun. 2025, 16, 9277. [Google Scholar] [CrossRef]
- Takahashi, K.H. Multiple capacitors for natural genetic variation in Drosophila melanogaster. Mol. Ecol. 2013, 22, 1356–1365. [Google Scholar] [CrossRef]
- Takahashi, K.H.; Ishimori, M.; Iwata, H. HSP90 as a global genetic modifier for male genital morphology in Drosophila melanogaster. Evolution 2018, 72, 2419–2434. [Google Scholar] [CrossRef]
- Zabinsky, R.A.; Mason, G.A.; Queitsch, C.; Jarosz, D.F. It’s not magic-Hsp90 and its effects on genetic and epigenetic variation. Semin. Cell Dev. Biol. 2019, 88, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Zohn, I.E. Hsp90 and complex birth defects: A plausible mechanism for the interaction of genes and environment. Neurosci. Lett. 2020, 716, 134680. [Google Scholar] [CrossRef]
- Rutherford, S.L.; Henikoff, S. Quantitative epigenetics. Nat. Genet. 2003, 33, 6–8. [Google Scholar] [CrossRef]
- Wei, H.; Zhang, Y.; Jia, Y.; Chen, X.; Niu, T.; Chatterjee, A.; He, P.; Hou, G. Heat shock protein 90: Biological functions, diseases, and therapeutic targets. MedComm (2020) 2024, 5, e470. [Google Scholar] [CrossRef]
- Izumi, N.; Kawaoka, S.; Yasuhara, S.; Suzuki, Y.; Sugano, S.; Katsuma, S.; Tomari, Y. Hsp90 facilitates accurate loading of precursor piRNAs into PIWI proteins. RNA 2013, 19, 896–901, Erratum in RNA 2015, 21, 1217. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, M.; Chaudhry, M.T.; Xue, H.; Zhang, L.; Huang, Y.; Liu, C. Translation, ribosome biogenesis, and oxidative damage caused by chlorpyrifos exposure to common carp (Cyprinus carpio L.) liver: Application of combined RNA-seq with sRNA-seq in risk evaluation of environmental toxicant chlorpyrifos. Environ. Sci. Pollut. Res. Int. 2021, 28, 56117–56125. [Google Scholar] [CrossRef]
- Park, S.H.; Butcher, B.G.; Anderson, Z.; Pellegrini, N.; Bao, Z.; D’Amico, K.; Filiatrault, M.J. Analysis of the small RNA P16/RgsA in the plant pathogen Pseudomonas syringae pv. tomato strain DC3000. Microbiology 2013, 159, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Vanderpool, C.K. Physiological consequences of multiple-target regulation by the small RNA SgrS in Escherichia coli. J. Bacteriol. 2013, 195, 4804–4815. [Google Scholar] [CrossRef]
- Yue, E.; Rong, F.; Liu, Z.; Ruan, S.; Lu, T.; Qian, H. Cadmium induced a non-coding RNA microRNA535 mediates Cd accumulation in rice. J. Environ. Sci. 2023, 130, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Qi, Z.; Dong, Z.; Liu, W.; Xu, M.; Gao, M.; Liu, S. LncRNA MT1DP promotes cadmium-induced DNA replication stress by inhibiting chromatin recruitment of SMARCAL1. Sci. Total Environ. 2022, 807, 151078. [Google Scholar] [PubMed]
- Hernandez-Saavedra, D.; Strakovsky, R.S.; Ostrosky-Wegman, P.; Pan, Y.X. Epigenetic Regulation of Centromere Chromatin Stability by Dietary and Environmental Factors. Adv. Nutr. 2017, 8, 889–904. [Google Scholar]
- McCullough, S.D.; Bowers, E.C.; On, D.M.; Morgan, D.S.; Dailey, L.A.; Hines, R.N.; Devlin, R.B.; Diaz-Sanchez, D. Baseline Chromatin Modification Levels May Predict Interindividual Variability in Ozone-Induced Gene Expression. Toxicol. Sci. 2016, 150, 216–224. [Google Scholar]
- Weinhouse, C. The roles of inducible chromatin and transcriptional memory in cellular defense system responses to redox-active pollutants. Free Radic. Biol. Med. 2021, 170, 85–108. [Google Scholar] [CrossRef]
- Alimba, C.G.; Adesida, S.O.; Owonikoko, W.M.; Oteyola, A.O. Micro (Nano)plastics Vitiation of Genetic Material: Systematic Review of Genotoxic Biomarkers and Model Bioindicators. J. Appl. Toxicol. 2026, 46, 110–140. [Google Scholar]
- Bajinka, O.; Barrow, A.; Mendy, S.; Jallow, B.J.J.; Jallow, J.; Barrow, S.; Bah, O.; Camara, S.; Colley, M.L.; Nyabally, S.; et al. The Influence of Parental Environmental Exposure and Nutrient Restriction on the Early Life of Offspring Growth in Gambia-A Pilot Study. Int. J. Environ. Res. Public Health 2022, 19, 13045. [Google Scholar] [CrossRef]
- Bulut, B.; Rigano, L.; Doria, H.B.; Gemuth, G.; Pfenninger, M. A multigenerational study can detect the evolutionary response to BaP exposure in the non-biting freshwater midge Chironomus riparius. Chemosphere 2024, 358, 142242. [Google Scholar] [CrossRef] [PubMed]
- Guedon, R.; Maremonti, E.; Armant, O.; Galas, S.; Brede, D.A.; Lecomte-Pradines, C. A systems biology analysis of reproductive toxicity effects induced by multigenerational exposure to ionizing radiation in C. elegans. Ecotoxicol. Environ. Saf. 2021, 225, 112793. [Google Scholar] [CrossRef] [PubMed]
- Henderson, A.L.; Colaiacovo, M.P. Exposure to phthalates: Germline dysfunction and aneuploidy. Prenat. Diagn. 2021, 41, 610–619. [Google Scholar] [CrossRef]
- Kang, Y.; Lee, Y.; Oh, S.Y.; Hong, K.; Do, J.T. Bisphenol A impairs developmental potential of mouse blastoids through oxidative stress. Environ. Int. 2025, 204, 109820. [Google Scholar] [CrossRef]
- Lei, J.; Du, H.; Bai, M.; Li, K.; Fu, H.; Zhu, Y. Low-Level Exposure to Dinotefuran May Induce Damage to Ovarian and Female Germ Stem Cells of Adolescent Mice via ZBP1- and RIPK1-PANoptosome. Environ. Sci. Technol. 2026, 60, 2337–2352. [Google Scholar] [CrossRef]
- Liu, X.; Ge, P.; Lu, Z.; Yang, R.; Liu, Z.; Zhao, F.; Chen, M. Reproductive toxicity and underlying mechanisms of fine particulate matter (PM(2.5)) on Caenorhabditis elegans in different seasons. Ecotoxicol. Environ. Saf. 2022, 248, 114281. [Google Scholar] [CrossRef]
- Marcho, C.; Oluwayiose, O.A.; Pilsner, J.R. The preconception environment and sperm epigenetics. Andrology 2020, 8, 924–942. [Google Scholar] [CrossRef]
- Okutsu, J.; Noor, M.I.; Shelton, D.S. Swimming Into View: Zebrafish Uncover Targets, Mechanisms, and Therapies for Cadmium Toxicity. Curr. Environ. Health Rep. 2025, 12, 20. [Google Scholar] [CrossRef]
- Cai, C.; Chen, Q. Father’s diet influences son’s metabolic health through sperm RNA. Nature 2024, 630, 571–573. [Google Scholar] [CrossRef]
- Vaz, C.; Burton, M.; Kermack, A.J.; Tan, P.F.; Huan, J.; Yoo, T.P.X.; Donnelly, K.; Wellstead, S.J.; Wang, D.; Fisk, H.L.; et al. Short-term diet intervention comprising of olive oil, vitamin D, and omega-3 fatty acids alters the small non-coding RNA (sncRNA) landscape of human sperm. Sci. Rep. 2025, 15, 7790. [Google Scholar] [CrossRef] [PubMed]
- Perera, B.P.U.; Tsai, Z.T.; Colwell, M.L.; Jones, T.R.; Goodrich, J.M.; Wang, K.; Sartor, M.A.; Faulk, C.; Dolinoy, D.C. Somatic expression of piRNA and associated machinery in the mouse identifies short, tissue-specific piRNA. Epigenetics 2019, 14, 504–521. [Google Scholar] [CrossRef]
- Maxwell, D.L.; Oluwayiose, O.A.; Houle, E.; Roth, K.; Nowak, K.; Sawant, S.; Paskavitz, A.L.; Liu, W.; Gurdziel, K.; Petriello, M.C.; et al. Mixtures of per- and polyfluoroalkyl substances (PFAS) alter sperm methylation and long-term reprogramming of offspring liver and fat transcriptome. Environ. Int. 2024, 186, 108577. [Google Scholar] [CrossRef]
- Ashapkin, V.; Suvorov, A.; Pilsner, J.R.; Krawetz, S.A.; Sergeyev, O. Age-associated epigenetic changes in mammalian sperm: Implications for offspring health and development. Hum. Reprod. Update 2023, 29, 24–44. [Google Scholar] [CrossRef]
- Hamada, H.; Casciaro, C.; Moisiadis, V.G.; Constantinof, A.; Kostaki, A.; Matthews, S.G. Prenatal maternal glucocorticoid exposure modifies sperm miRNA profiles across multiple generations in the guinea-pig. J. Physiol. 2024, 602, 2127–2139. [Google Scholar] [CrossRef]
- Kropp, J.; Carrillo, J.A.; Namous, H.; Daniels, A.; Salih, S.M.; Song, J.; Khatib, H. Male fertility status is associated with DNA methylation signatures in sperm and transcriptomic profiles of bovine preimplantation embryos. BMC Genom. 2017, 18, 280. [Google Scholar] [CrossRef]
- Le Blevec, E.; Muronova, J.; Ray, P.F.; Arnoult, C. Paternal epigenetics: Mammalian sperm provide much more than DNA at fertilization. Mol. Cell Endocrinol. 2020, 518, 110964. [Google Scholar] [CrossRef]
- Nobles, C.; Canty, T.P.; Mendola, P.; Russo, L.M.; Rabeya, K.; Schliep, K.C.; Shaaban, M.; Singh, A.; Ring, A.M.; Hemmert, R.; et al. Heat exposure during susceptible windows of spermatogenesis and sperm epigenetic age. Hum. Reprod. 2026, in press. [Google Scholar] [CrossRef]
- Oluwayiose, O.A.; Houle, E.; Wu, H.; Whitcomb, B.W.; Mumford, S.L.; Schisterman, E.F.; Suvorov, A.; Balzer, L.B.; Pilsner, J.R. Urinary phthalate metabolites and their mixtures are associated with advanced sperm epigenetic aging in a general population. Environ. Res. 2022, 214, 114115. [Google Scholar] [CrossRef]
- Oluwayiose, O.A.; Wu, H.; Saddiki, H.; Whitcomb, B.W.; Balzer, L.B.; Brandon, N.; Suvorov, A.; Tayyab, R.; Sites, C.K.; Hill, L.; et al. Sperm DNA methylation mediates the association of male age on reproductive outcomes among couples undergoing infertility treatment. Sci. Rep. 2021, 11, 3216. [Google Scholar] [CrossRef]
- Pilsner, J.R.; Saddiki, H.; Whitcomb, B.W.; Suvorov, A.; Buck Louis, G.M.; Mumford, S.L.; Schisterman, E.F.; Oluwayiose, O.A.; Balzer, L.B. Sperm epigenetic clock associates with pregnancy outcomes in the general population. Hum. Reprod. 2022, 37, 1581–1593. [Google Scholar]
- Sawant, S.; Oluwayiose, O.A.; Nowak, K.; Maxwell, D.L.; Houle, E.; Paskavitz, A.L.; Saddiki, H.; Bertolla, R.P.; Pilsner, J.R. Associations between Sperm Epigenetic Age and Semen Parameters: An Evaluation of Clinical and Non-Clinical Cohorts. Curr. Issues Mol. Biol. 2024, 46, 1567–1578. [Google Scholar] [CrossRef]
- Suvorov, A.; Shershebnev, A.; Wu, H.; Medvedeva, Y.; Sergeyev, O.; Pilsner, J.R. Perinatal exposure to low dose 2,2′,4,4′-tetrabromodiphenyl ether (BDE-47) alters sperm DNA methylation in adult rats. Reprod. Toxicol. 2018, 75, 136–143. [Google Scholar] [CrossRef]
- Zhang, H.; Shan, L.; Aniagu, S.; Jiang, Y.; Chen, T. Paternal acrylamide exposure induces transgenerational effects on sperm parameters and learning capability in mice. Food Chem. Toxicol. 2022, 161, 112817. [Google Scholar] [CrossRef]
- Karam, J.A.; Parikh, R.Y.; Nayak, D.; Rosenkranz, D.; Gangaraju, V.K. Co-chaperone Hsp70/Hsp90-organizing protein (Hop) is required for transposon silencing and Piwi-interacting RNA (piRNA) biogenesis. J. Biol. Chem. 2017, 292, 6039–6046. [Google Scholar] [CrossRef]
- Negi, P.; Rai, A.N.; Suprasanna, P. Moving through the Stressed Genome: Emerging Regulatory Roles for Transposons in Plant Stress Response. Front. Plant Sci. 2016, 7, 1448. [Google Scholar] [CrossRef]
- Sienski, G.; Donertas, D.; Brennecke, J. Transcriptional silencing of transposons by Piwi and maelstrom and its impact on chromatin state and gene expression. Cell 2012, 151, 964–980. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, H.; Xiao, Y.; Liu, M.; Ren, A.; Liu, S.; Wu, L. Integrated small and long RNA sequencing reveals piRNA mediated transposon repression during human oogenesis. Nat. Commun. 2026, 17, 3804. [Google Scholar] [CrossRef]
- Wheeler, B.S. Small RNAs, big impact: Small RNA pathways in transposon control and their effect on the host stress response. Chromosome Res. 2013, 21, 587–600. [Google Scholar] [CrossRef]
- Wang, H.L.; Chekanova, J.A. Small RNAs: Essential regulators of gene expression and defenses against environmental stresses in plants. Wiley Interdiscip. Rev. RNA 2016, 7, 356–381. [Google Scholar] [CrossRef]
- Mandujano-Tinoco, E.A.; Sultan, E.; Ottolenghi, A.; Gershoni-Yahalom, O.; Rosental, B. Evolution of Cellular Immunity Effector Cells; Perspective on Cytotoxic and Phagocytic Cellular Lineages. Cells 2021, 10, 1853. [Google Scholar] [CrossRef]
- Bajgar, A.; Krejcova, G. On the origin of the functional versatility of macrophages. Front. Physiol. 2023, 14, 1128984. [Google Scholar] [CrossRef]
- Lim, J.J.; Grinstein, S.; Roth, Z. Diversity and Versatility of Phagocytosis: Roles in Innate Immunity, Tissue Remodeling, and Homeostasis. Front. Cell Infect. Microbiol. 2017, 7, 191. [Google Scholar] [CrossRef]
- Arefin, B.; Kucerova, L.; Krautz, R.; Kranenburg, H.; Parvin, F.; Theopold, U. Apoptosis in Hemocytes Induces a Shift in Effector Mechanisms in the Drosophila Immune System and Leads to a Pro-Inflammatory State. PLoS ONE 2015, 10, e0136593. [Google Scholar] [CrossRef]
- Balog, J.A.; Honti, V.; Kurucz, E.; Kari, B.; Puskas, L.G.; Ando, I.; Szebeni, G.J. Immunoprofiling of Drosophila Hemocytes by Single-cell Mass Cytometry. Genom. Proteom. Bioinform. 2021, 19, 243–252. [Google Scholar] [CrossRef]
- Braun, A.; Lemaitre, B.; Lanot, R.; Zachary, D.; Meister, M. Drosophila immunity: Analysis of larval hemocytes by P-element-mediated enhancer trap. Genetics 1997, 147, 623–634. [Google Scholar] [CrossRef]
- Myers, A.L.; Harris, C.M.; Choe, K.M.; Brennan, C.A. Inflammatory production of reactive oxygen species by Drosophila hemocytes activates cellular immune defenses. Biochem. Biophys. Res. Commun. 2018, 505, 726–732. [Google Scholar] [CrossRef]
- Tirouvanziam, R.; Davidson, C.J.; Lipsick, J.S.; Herzenberg, L.A. Fluorescence-activated cell sorting (FACS) of Drosophila hemocytes reveals important functional similarities to mammalian leukocytes. Proc. Natl. Acad. Sci. USA 2004, 101, 2912–2917. [Google Scholar] [CrossRef]
- Varga, V.; Szinyakovics, J.; Bebes, A.; Szikszai, F.; Kovacs, T. Role of Hemocytes in the Aging of Drosophila Male Germline. Cells 2025, 14, 315. [Google Scholar] [CrossRef]
- Yoon, S.H.; Cho, B.; Lee, D.; Kim, H.; Shim, J.; Nam, J.W. Molecular traces of Drosophila hemocytes reveal transcriptomic conservation with vertebrate myeloid cells. PLoS Genet. 2023, 19, e1011077. [Google Scholar] [CrossRef]
- Aigner, G.P.; Peer, V.; Fiechtner, B.; Piechnik, C.A.; Hockner, M. Wound healing and Cadmium detoxification in the earthworm Lumbricus terrestris-a potential case for coelomocytes? Front. Immunol. 2023, 14, 1272191. [Google Scholar] [CrossRef]
- Franco, C.F.; Santos, R.; Coelho, A.V. Proteome characterization of sea star coelomocytes--the innate immune effector cells of echinoderms. Proteomics 2011, 11, 3587–3592. [Google Scholar] [CrossRef]
- Matranga, V.; Toia, G.; Bonaventura, R.; Muller, W.E. Cellular and biochemical responses to environmental and experimentally induced stress in sea urchin coelomocytes. Cell Stress. Chaperones 2000, 5, 113–120. [Google Scholar] [CrossRef]
- Pagliara, P.; Mazzei, A.; Stasi, F.; Stetco, A.C.; Barca, A.; Vizioli, J.; Verri, T. Post traumatic response of coelomocytes in the common sea star Echinastersepositus. Fish Shellfish. Immunol. 2026, 174, 111365. [Google Scholar] [CrossRef]
- Amiss, A.S.; von Pein, J.B.; Webb, J.R.; Condon, N.D.; Harvey, P.J.; Phan, M.D.; Schembri, M.A.; Currie, B.J.; Sweet, M.J.; Craik, D.J.; et al. Modified horseshoe crab peptides target and kill bacteria inside host cells. Cell Mol. Life Sci. 2021, 79, 38. [Google Scholar] [CrossRef]
- Willenz, P.; Van de Vyver, G. Endocytosis of latex beads by the exopinacoderm in the fresh water sponge Ephydatia fluviatilis: An in vitro and in situ study in SEM and TEM. J. Ultrastruct. Res. 1982, 79, 294–306. [Google Scholar] [CrossRef]
- Pollard, J.W. Trophic macrophages in development and disease. Nat. Rev. Immunol. 2009, 9, 259–270. [Google Scholar] [CrossRef]
- Guan, F.; Wang, R.; Yi, Z.; Luo, P.; Liu, W.; Xie, Y.; Liu, Z.; Xia, Z.; Zhang, H.; Cheng, Q. Tissue macrophages: Origin, heterogenity, biological functions, diseases and therapeutic targets. Signal Transduct. Target. Ther. 2025, 10, 93. [Google Scholar] [CrossRef]
- Vuscan, P.; Kischkel, B.; Joosten, L.A.B.; Netea, M.G. Trained immunity: General and emerging concepts. Immunol. Rev. 2024, 323, 164–185. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, X.; Xu, M.; Yuan, J.; Liu, Y. Monocyte/macrophage-mediated trained immunity in disease prevention and immunotherapy. Bioact. Mater. 2026, 59, 745–762. [Google Scholar] [CrossRef]
- Yan, L.; Wang, J.; Cai, X.; Liou, Y.C.; Shen, H.M.; Hao, J.; Huang, C.; Luo, G.; He, W. Macrophage plasticity: Signaling pathways, tissue repair, and regeneration. MedComm 2024, 5, e658. [Google Scholar] [CrossRef]
- Chen, S.; Saeed, A.; Liu, Q.; Jiang, Q.; Xu, H.; Xiao, G.G.; Rao, L.; Duo, Y. Macrophages in immunoregulation and therapeutics. Signal Transduct. Target. Ther. 2023, 8, 207. [Google Scholar] [CrossRef]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef]
- Wang, L.; Yang, K.; Xie, X.; Wang, S.; Gan, H.; Wang, X.; Wei, H. Macrophages as Multifaceted Orchestrators of Tissue Repair: Bridging Inflammation, Regeneration, and Therapeutic Innovation. J. Inflamm. Res. 2025, 18, 8945–8959. [Google Scholar] [CrossRef]
- Chi, Y.; Jiang, H.; Yin, Y.; Zhou, X.; Shao, Y.; Li, Y.; Rao, J. Macrophage Signaling Pathways in Health and Disease: From Bench to Bedside Applications. MedComm 2025, 6, e70256. [Google Scholar] [CrossRef]
- Ji, Y.; Li, X.; Yao, X.; Sun, J.; Yi, J.; Shen, Y.; Chen, B.; Sun, H. Macrophage polarization: Molecular mechanisms, disease implications, and targeted therapeutic strategies. Front. Immunol. 2025, 16, 1732718. [Google Scholar] [CrossRef]
- Bai, X.; Guo, Y.R.; Zhao, Z.M.; Li, X.Y.; Dai, D.Q.; Zhang, J.K.; Li, Y.S.; Zhang, C.D. Macrophage polarization in cancer and beyond: From inflammatory signaling pathways to potential therapeutic strategies. Cancer Lett. 2025, 625, 217772. [Google Scholar] [CrossRef]
- Boutilier, A.J.; Elsawa, S.F. Macrophage Polarization States in the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 6995. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Zhong, S.; Chen, X.; Liu, H.Y.; Lu, C.; Zhu, H.; Chen, Y.; Wang, J.; Yu, J.; et al. Macrophage-derived exosome piR-50971 exacerbates sepsis-induced myocardial injury by inhibiting autophagy through the upregulation of N6-Methyladenosine modification of mTOR. Burn. Trauma. 2025, 13, tkaf045. [Google Scholar] [CrossRef]
- Sun, D.S.; Chang, H.H. Extracellular vesicles: Function, resilience, biomarker, bioengineering, and clinical implications. Tzu Chi Med. J. 2024, 36, 251–259. [Google Scholar] [CrossRef]
- Saviana, M.; Romano, G.; Del Valle-Morales, D.; Acunzo, M.; Nana-Sinkam, P. Look Who Is Talking: Extracellular Vesicles as Mediators of Intercellular and Interorgan Communication. Compr. Physiol. 2026, 16, e70128. [Google Scholar] [CrossRef]
- Salomon, C.; Das, S.; Erdbrugger, U.; Kalluri, R.; Kiang Lim, S.; Olefsky, J.M.; Rice, G.E.; Sahoo, S.; Andy Tao, W.; Vader, P.; et al. Extracellular Vesicles and Their Emerging Roles as Cellular Messengers in Endocrinology: An Endocrine Society Scientific Statement. Endocr. Rev. 2022, 43, 441–468. [Google Scholar] [CrossRef]
- Yarana, C.; Thompson, H.; Chaiswing, L.; Butterfield, D.A.; Weiss, H.; Bondada, S.; Alhakeem, S.; Sukati, S.; St Clair, D.K. Extracellular vesicle-mediated macrophage activation: An insight into the mechanism of thioredoxin-mediated immune activation. Redox Biol. 2019, 26, 101237. [Google Scholar] [CrossRef]
- Tian, J.W.; Fang, Y.H.; Zhang, H.J.; Yu, Z.L. The dual effects of macrophage-derived extracellular vesicles on tumor cell behavior: Mechanisms and clinical potential. Front. Oncol. 2025, 15, 1586083. [Google Scholar] [CrossRef]
- Cianciaruso, C.; Beltraminelli, T.; Duval, F.; Nassiri, S.; Hamelin, R.; Mozes, A.; Gallart-Ayala, H.; Ceada Torres, G.; Torchia, B.; Ries, C.H.; et al. Molecular Profiling and Functional Analysis of Macrophage-Derived Tumor Extracellular Vesicles. Cell Rep. 2019, 27, 3062–3080.e11. [Google Scholar] [CrossRef]
- Casara, A.; Conti, M.; Bernardinello, N.; Tine, M.; Baraldo, S.; Turato, G.; Semenzato, U.; Celi, A.; Spagnolo, P.; Saetta, M.; et al. Unveiling the Cutting-Edge Impact of Polarized Macrophage-Derived Extracellular Vesicles and MiRNA Signatures on TGF-beta Regulation within Lung Fibroblasts. Int. J. Mol. Sci. 2024, 25, 7490. [Google Scholar] [CrossRef]
- Chai, Z.; Dong, X.; Li, S.; Dai, M.; Li, H.; He, Z.; Gu, P. Macrophage-derived extracellular vesicles in the remodeling of the prostate cancer immune microenvironment and therapeutic resistance. J. Transl. Med. 2026, 24, 553. [Google Scholar] [CrossRef]
- Lv, K.; Zhang, Y.; Yin, G.; Li, X.; Zhong, M.; Zhu, X.; Pei, W. Extracellular vesicles derived from lung M2 macrophages enhance group 2 innate lymphoid cells function in allergic airway inflammation. Exp. Mol. Med. 2025, 57, 1202–1215. [Google Scholar] [CrossRef]
- Deville, S.; Garcia Romeu, H.; Oeyen, E.; Mertens, I.; Nelissen, I.; Salvati, A. Macrophages Release Extracellular Vesicles of Different Properties and Composition Following Exposure to Nanoparticles. Int. J. Mol. Sci. 2022, 24, 260. [Google Scholar] [CrossRef]
- Shi, S.; Ou, X.; Wang, Q.; Zhang, L. Macrophage-Derived Extracellular Vesicles: A Novel Therapeutic Alternative for Diabetic Wound. Int. J. Nanomed. 2025, 20, 5763–5777. [Google Scholar] [CrossRef]
- Cosentino, C.; Klein, R.; Menoud, V.; Guay, C.; Aiello, E.; Auddino, S.; Di Giuseppe, G.; Ciccarelli, G.; Galli, A.; Alabiso, F.; et al. 5′tRNA-derived fragments modulate beta-cell homeostasis and islet macrophage activation in type 2 diabetes. Nat. Commun. 2026, in press. [Google Scholar] [CrossRef]
- Dou, R.; Zhang, X.; Xu, X.; Wang, P.; Yan, B. Mesenchymal stem cell exosomal tsRNA-21109 alleviate systemic lupus erythematosus by inhibiting macrophage M1 polarization. Mol. Immunol. 2021, 139, 106–114. [Google Scholar] [CrossRef]
- Lu, L.; Li, J.; Wei, R.; Guidi, I.; Cozzuto, L.; Ponomarenko, J.; Prats-Ejarque, G.; Boix, E. Selective cleavage of ncRNA and antiviral activity by RNase2/EDN in THP1-induced macrophages. Cell Mol. Life Sci. 2022, 79, 209. [Google Scholar] [CrossRef]
- Lu, S.; Wei, X.; Tao, L.; Dong, D.; Hu, W.; Zhang, Q.; Tao, Y.; Yu, C.; Sun, D.; Cheng, H. A novel tRNA-derived fragment tRF-3022b modulates cell apoptosis and M2 macrophage polarization via binding to cytokines in colorectal cancer. J. Hematol. Oncol. 2022, 15, 176. [Google Scholar] [CrossRef]
- Wang, X.; Hu, Z. tRNA derived fragment tsRNA-14783 promotes M2 polarization of macrophages in keloid. Biochem. Biophys. Res. Commun. 2022, 636, 119–127. [Google Scholar] [CrossRef]
- Winek, K.; Lobentanzer, S.; Nadorp, B.; Dubnov, S.; Dames, C.; Jagdmann, S.; Moshitzky, G.; Hotter, B.; Meisel, C.; Greenberg, D.S.; et al. Transfer RNA fragments replace microRNA regulators of the cholinergic poststroke immune blockade. Proc. Natl. Acad. Sci. USA 2020, 117, 32606–32616. [Google Scholar] [CrossRef]
- Ye, J.; Liu, X. Macrophage-Derived Small Extracellular Vesicles in Multiple Diseases: Biogenesis, Function, and Therapeutic Applications. Front. Cell Dev. Biol. 2022, 10, 913110. [Google Scholar] [CrossRef]
- Lin, F.; Luo, H.; Wang, J.; Li, Q.; Zha, L. Macrophage-derived extracellular vesicles as new players in chronic non-communicable diseases. Front. Immunol. 2024, 15, 1479330. [Google Scholar] [CrossRef]
- Smith, T.I.; Russell, A.E. Extracellular vesicles in reproduction and pregnancy. Extracell. Vesicles Circ. Nucl. Acids 2022, 3, 292–317. [Google Scholar] [CrossRef]
- Tannetta, D.; Dragovic, R.; Alyahyaei, Z.; Southcombe, J. Extracellular vesicles and reproduction-promotion of successful pregnancy. Cell Mol. Immunol. 2014, 11, 548–563. [Google Scholar] [CrossRef]
- Chan, J.C.; Morgan, C.P.; Adrian Leu, N.; Shetty, A.; Cisse, Y.M.; Nugent, B.M.; Morrison, K.E.; Jasarevic, E.; Huang, W.; Kanyuch, N.; et al. Reproductive tract extracellular vesicles are sufficient to transmit intergenerational stress and program neurodevelopment. Nat. Commun. 2020, 11, 1499. [Google Scholar] [CrossRef]
- Simon, C.; Greening, D.W.; Bolumar, D.; Balaguer, N.; Salamonsen, L.A.; Vilella, F. Extracellular Vesicles in Human Reproduction in Health and Disease. Endocr. Rev. 2018, 39, 292–332. [Google Scholar] [CrossRef]
- Barrachina, F.; Battistone, M.A.; Castillo, J.; Mallofre, C.; Jodar, M.; Breton, S.; Oliva, R. Sperm acquire epididymis-derived proteins through epididymosomes. Hum. Reprod. 2022, 37, 651–668. [Google Scholar] [CrossRef]
- Trigg, N.A.; Lee, G.S.; Leach, A.G.; Conine, C.C. Epididymal extracellular vesicles harbor and convey mRNA to sperm for transfer to zygotes. Nucleic. Acids Res. 2026, 54, gkag330. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.; Talluri, T.R.; Nag, P.; Kumaresan, A. Epididymosomes: A potential male fertility influencer. Andrologia 2021, 53, e14155. [Google Scholar] [CrossRef]
- Pantano, L.; Jodar, M.; Bak, M.; Ballesca, J.L.; Tommerup, N.; Oliva, R.; Vavouri, T. The small RNA content of human sperm reveals pseudogene-derived piRNAs complementary to protein-coding genes. RNA 2015, 21, 1085–1095. [Google Scholar] [CrossRef]
- Du, L.; Chen, W.; Zhang, D.; Cui, Y.; He, Z. The functions and mechanisms of piRNAs in mediating mammalian spermatogenesis and their applications in reproductive medicine. Cell Mol. Life Sci. 2024, 81, 379. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, K.; Zhao, Z.; Feng, M.; Song, L.; Xu, Z. piRNA: Molecular Mechanisms from Germline Silencing to Somatic Regulation and Roles in Disease. Int. J. Mol. Sci. 2026, 27, 2685. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Gao, Q.; Peng, X.; Choi, S.Y.; Sarma, K.; Ren, H.; Morris, A.J.; Frohman, M.A. piRNA-associated germline nuage formation and spermatogenesis require MitoPLD profusogenic mitochondrial-surface lipid signaling. Dev. Cell 2011, 20, 376–387. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, M.; Liu, S.; Guo, J.; Lu, Y.; Cheng, J.; Liu, J. Macrophage-derived extracellular vesicles: Diverse mediators of pathology and therapeutics in multiple diseases. Cell Death Dis. 2020, 11, 924. [Google Scholar] [CrossRef]
- Wu, H.; Hauser, R.; Krawetz, S.A.; Pilsner, J.R. Environmental Susceptibility of the Sperm Epigenome During Windows of Male Germ Cell Development. Curr. Environ. Health Rep. 2015, 2, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Tomar, A.; Gomez-Velazquez, M.; Gerlini, R.; Comas-Armangue, G.; Makharadze, L.; Kolbe, T.; Boersma, A.; Dahlhoff, M.; Burgstaller, J.P.; Lassi, M.; et al. Epigenetic inheritance of diet-induced and sperm-borne mitochondrial RNAs. Nature 2024, 630, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Donkin, I.; Barres, R. Sperm epigenetics and influence of environmental factors. Mol. Metab. 2018, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Lu, D.; Reisinger, S.N.; Mehrabadi, M.R.; Gubert, C.; Hannan, A.J. Epigenetic effects of paternal environmental exposures and experiences on offspring phenotypes. Trends Genet. 2025, 41, 735–761. [Google Scholar] [CrossRef]
- McSwiggin, H.; Magalhaes, R.; Nilsson, E.E.; Yan, W.; Skinner, M.K. Epigenetic transgenerational inheritance of toxicant exposure-specific non-coding RNA in sperm. Environ. Epigenet. 2024, 10, dvae014. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Losada, M.; Blom-Dahl, D.; Cordoba, S.; Estella, C. Specification and Patterning of Drosophila Appendages. J. Dev. Biol. 2018, 6, 17. [Google Scholar] [CrossRef]
- Parikh, R.Y.; Gangaraju, V.K. Hexavalent chromium-induced epigenetic instability and transposon activation lead to phenotypic variations and tumors in Drosophila. Environ. Epigenet. 2023, 9, dvac030. [Google Scholar] [CrossRef]
- Wood, W.; Jacinto, A. Drosophila melanogaster embryonic haemocytes: Masters of multitasking. Nat. Rev. Mol. Cell Biol. 2007, 8, 542–551. [Google Scholar] [CrossRef]
- Boniakowski, A.E.; Kimball, A.S.; Joshi, A.; Schaller, M.; Davis, F.M.; den Dekker, A.; Obi, A.T.; Moore, B.B.; Kunkel, S.L.; Gallagher, K.A. Murine macrophage chemokine receptor CCR2 plays a crucial role in macrophage recruitment and regulated inflammation in wound healing. Eur. J. Immunol. 2018, 48, 1445–1455. [Google Scholar] [CrossRef]
- Koh, T.J.; DiPietro, L.A. Inflammation and wound healing: The role of the macrophage. Expert. Rev. Mol. Med. 2011, 13, e23. [Google Scholar] [CrossRef]
- Yu, Y.; Yue, Z.; Xu, M.; Zhang, M.; Shen, X.; Ma, Z.; Li, J.; Xie, X. Macrophages play a key role in tissue repair and regeneration. PeerJ 2022, 10, e14053. [Google Scholar] [CrossRef]
- Minutti, C.M.; Knipper, J.A.; Allen, J.E.; Zaiss, D.M. Tissue-specific contribution of macrophages to wound healing. Semin. Cell Dev. Biol. 2017, 61, 3–11. [Google Scholar] [CrossRef]
- Hersperger, F.; Meyring, T.; Weber, P.; Chhatbar, C.; Monaco, G.; Dionne, M.S.; Paeschke, K.; Prinz, M.; Gross, O.; Classen, A.K.; et al. DNA damage signaling in Drosophila macrophages modulates systemic cytokine levels in response to oxidative stress. eLife 2024, 12, RP86700. [Google Scholar] [CrossRef] [PubMed]
- Wood, W.; Martin, P. Macrophage Functions in Tissue Patterning and Disease: New Insights from the Fly. Dev. Cell 2017, 40, 221–233. [Google Scholar] [CrossRef]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef] [PubMed]
- Iraci, N.; Leonardi, T.; Gessler, F.; Vega, B.; Pluchino, S. Focus on Extracellular Vesicles: Physiological Role and Signalling Properties of Extracellular Membrane Vesicles. Int. J. Mol. Sci. 2016, 17, 171. [Google Scholar] [CrossRef]
- Liu, S.; Wu, X.; Chandra, S.; Lyon, C.; Ning, B.; Jiang, L.; Fan, J.; Hu, T.Y. Extracellular vesicles: Emerging tools as therapeutic agent carriers. Acta Pharm. Sin. B 2022, 12, 3822–3842. [Google Scholar] [CrossRef]
- Rimmer, M.P.; Gregory, C.D.; Mitchell, R.T. The transformative impact of extracellular vesicles on developing sperm. Reprod. Fertil. 2021, 2, R51–R66. [Google Scholar] [CrossRef]
- Qamar, A.Y.; Hussain, T.; Rafique, M.K.; Bang, S.; Tanga, B.M.; Seong, G.; Fang, X.; Saadeldin, I.M.; Cho, J. The Role of Stem Cells and Their Derived Extracellular Vesicles in Restoring Female and Male Fertility. Cells 2021, 10, 2460. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Liu, S.P.; Dai, X.F.; Lan, D.F.; Song, T.; Wang, X.Y.; Kong, Q.H.; Tan, J.; Zhang, J.D. The emerging role of exosomes in the development of testicular. Asian J. Androl. 2023, 25, 547–555. [Google Scholar] [CrossRef]
- Skinner, M.K. Environmental epigenetic transgenerational inheritance and somatic epigenetic mitotic stability. Epigenetics 2011, 6, 838–842. [Google Scholar] [CrossRef]
- Ewe, C.K.; Rechavi, O. The third barrier to transgenerational inheritance in animals: Somatic epigenetic resetting. EMBO Rep. 2023, 24, e56615. [Google Scholar] [CrossRef]
- Conine, C.C.; Rando, O.J. Soma-to-germline RNA communication. Nat. Rev. Genet. 2022, 23, 73–88. [Google Scholar] [CrossRef]
- Chennakesavan, K.; Haorah, J.; Samikkannu, T. piRNA/PIWI pathways and epigenetic crosstalk in human diseases: Molecular insights into HIV-1 infection and drugs of abuse. Mol. Ther. Nucleic Acids 2025, 36, 102473. [Google Scholar] [CrossRef] [PubMed]
- Atikukke, G.; Albosta, P.; Zhang, H.; Finley, R.L., Jr. A role for Drosophila Cyclin J in oogenesis revealed by genetic interactions with the piRNA pathway. Mech. Dev. 2014, 133, 64–76. [Google Scholar] [CrossRef]
- Wahba, L.; Hansen, L.; Fire, A.Z. An essential role for the piRNA pathway in regulating the ribosomal RNA pool in C. elegans. Dev. Cell 2021, 56, 2295–2312.e6. [Google Scholar] [CrossRef]
- Xu, H.; Guo, J.; Huang, Y.; Zhang, M.; Wang, Y.; Xia, L.; Cheng, X.; Meng, T.; Hao, R.; Wei, X.; et al. Insights into the role of hnRNPK in spermatogenesis via the piRNA pathway. Sci. Rep. 2025, 15, 6438. [Google Scholar] [CrossRef]
- Stefater, J.A., 3rd; Ren, S.; Lang, R.A.; Duffield, J.S. Metchnikoff’s policemen: Macrophages in development, homeostasis and regeneration. Trends Mol. Med. 2011, 17, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Theret, M.; Mounier, R.; Rossi, F. The origins and non-canonical functions of macrophages in development and regeneration. Development 2019, 146, dev156000. [Google Scholar] [CrossRef]
- Bohaud, C.; Johansen, M.D.; Jorgensen, C.; Kremer, L.; Ipseiz, N.; Djouad, F. The Role of Macrophages During Mammalian Tissue Remodeling and Regeneration Under Infectious and Non-Infectious Conditions. Front. Immunol. 2021, 12, 707856. [Google Scholar] [CrossRef]
- Kloosterman, D.J.; Akkari, L. Macrophages at the interface of the co-evolving cancer ecosystem. Cell 2023, 186, 1627–1651. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.; Siddiqui, H.; Riguene, E.; Nomikos, M. Zebrafish: A Versatile and Powerful Model for Biomedical Research. Bioessays 2025, 47, e70080. [Google Scholar] [CrossRef] [PubMed]
- Houwing, S.; Berezikov, E.; Ketting, R.F. Zili is required for germ cell differentiation and meiosis in zebrafish. EMBO J. 2008, 27, 2702–2711. [Google Scholar] [CrossRef]
- Li, L.; Yan, B.; Shi, Y.Q.; Zhang, W.Q.; Wen, Z.L. Live imaging reveals differing roles of macrophages and neutrophils during zebrafish tail fin regeneration. J. Biol. Chem. 2012, 287, 25353–25360. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, Y.; Hu, N.; Long, D.; Cao, Y. The uses of zebrafish (Danio rerio) as an in vivo model for toxicological studies: A review based on bibliometrics. Ecotoxicol. Environ. Saf. 2024, 272, 116023. [Google Scholar] [CrossRef] [PubMed]
- Garcia, G.R.; Noyes, P.D.; Tanguay, R.L. Advancements in zebrafish applications for 21st century toxicology. Pharmacol. Ther. 2016, 161, 11–21. [Google Scholar] [CrossRef]
- Beyret, E.; Liu, N.; Lin, H. piRNA biogenesis during adult spermatogenesis in mice is independent of the ping-pong mechanism. Cell Res. 2012, 22, 1429–1439. [Google Scholar] [CrossRef] [PubMed]
- Girardi, E.; Miesen, P.; Pennings, B.; Frangeul, L.; Saleh, M.C.; van Rij, R.P. Histone-derived piRNA biogenesis depends on the ping-pong partners Piwi5 and Ago3 in Aedes aegypti. Nucleic Acids Res. 2017, 45, 4881–4892. [Google Scholar]
- Morazzani, E.M.; Wiley, M.R.; Murreddu, M.G.; Adelman, Z.N.; Myles, K.M. Production of virus-derived ping-pong-dependent piRNA-like small RNAs in the mosquito soma. PLoS Pathog. 2012, 8, e1002470. [Google Scholar] [CrossRef]
- Xiong, M.; Yin, L.; Gui, Y.; Lv, C.; Ma, X.; Guo, S.; Wu, Y.; Feng, S.; Fan, X.; Zhou, S.; et al. ADAD2 interacts with RNF17 in P-bodies to repress the Ping-pong cycle in pachytene piRNA biogenesis. J. Cell Biol. 2023, 222, e202206067. [Google Scholar] [CrossRef]
- Yu, J.; Kawasaki, F.; Izumi, N.; Kiuchi, T.; Katsuma, S.; Tomari, Y.; Shoji, K. Autonomous shaping of the piRNA sequence repertoire by competition between adjacent ping-pong amplification sites. Mol. Cell 2025, 85, 1134–1146.e4. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, J.; Koppetsch, B.S.; Wang, J.; Tipping, C.; Ma, S.; Weng, Z.; Theurkauf, W.E.; Zamore, P.D. Heterotypic piRNA Ping-Pong requires qin, a protein with both E3 ligase and Tudor domains. Mol. Cell 2011, 44, 572–584. [Google Scholar] [CrossRef]
- Li, P.; Liu, M.; Liu, Y.; Guo, J.; Jiang, J.; Wang, G.; Cao, C.; Wang, X.; Qu, J.; Sun, Z. Recent Advances of Trained immunity in Macrophages. Int. J. Biol. Sci. 2025, 21, 5258–5283. [Google Scholar] [CrossRef]
- Arzola-Martinez, L.; Ptaschinski, C.; Lukacs, N.W. Trained innate immunity, epigenetics, and food allergy. Front. Allergy 2023, 4, 1105588. [Google Scholar] [CrossRef]
- Fanucchi, S.; Dominguez-Andres, J.; Joosten, L.A.B.; Netea, M.G.; Mhlanga, M.M. The Intersection of Epigenetics and Metabolism in Trained Immunity. Immunity 2021, 54, 32–43. [Google Scholar] [CrossRef]
- Feinberg, M.W. Editorial: The Role of Oxidative Stress, Epigenetics and Non-coding RNA in Regulating Trained Immunity. Front. Immunol. 2020, 11, 2114. [Google Scholar] [CrossRef]
- van der Heijden, C.; Noz, M.P.; Joosten, L.A.B.; Netea, M.G.; Riksen, N.P.; Keating, S.T. Epigenetics and Trained Immunity. Antioxid. Redox Signal. 2018, 29, 1023–1040. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Ruden, D.M. A Theoretical Hypothesis on How Immune Cells May Transmit Acquired Traits: A Macrophage–piRNA Pathway for Transgenerational Inheritance. Cells 2026, 15, 1030. https://doi.org/10.3390/cells15111030
Ruden DM. A Theoretical Hypothesis on How Immune Cells May Transmit Acquired Traits: A Macrophage–piRNA Pathway for Transgenerational Inheritance. Cells. 2026; 15(11):1030. https://doi.org/10.3390/cells15111030
Chicago/Turabian StyleRuden, Douglas M. 2026. "A Theoretical Hypothesis on How Immune Cells May Transmit Acquired Traits: A Macrophage–piRNA Pathway for Transgenerational Inheritance" Cells 15, no. 11: 1030. https://doi.org/10.3390/cells15111030
APA StyleRuden, D. M. (2026). A Theoretical Hypothesis on How Immune Cells May Transmit Acquired Traits: A Macrophage–piRNA Pathway for Transgenerational Inheritance. Cells, 15(11), 1030. https://doi.org/10.3390/cells15111030