
Isolation and Characterization of Mouse Choroidal Melanocytes and Their Proinflammatory Characteristics


Abstract

1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Animals
2.3. Tissue Preparation and Culture of ChMC
2.4. Isolation and Culture of Other Cell Types
2.5. Morphology Evaluations
2.6. Immunostaining of ChMC
2.7. Flow Cytometry Analysis
2.8. Reverse Transcription Quantitative PCR Analysis (RT-qPCR)
2.9. Choroid/RPE Tissue Staining of Melanocytes
2.10. Melanogenesis Studies
2.11. The Impact of Extracellular Stressors on Inflammatory Properties of ChMC
2.12. Statistical Analysis
3. Results
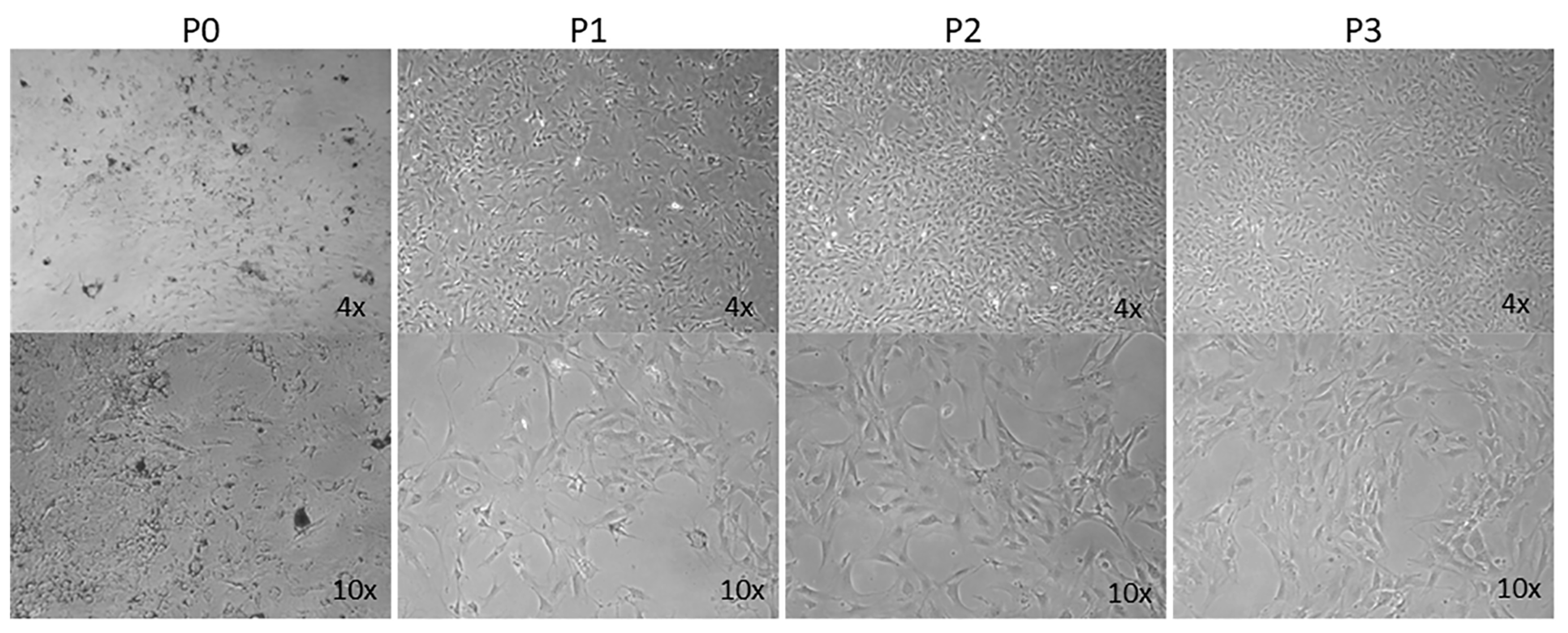
3.1. Isolation of Mouse ChMC
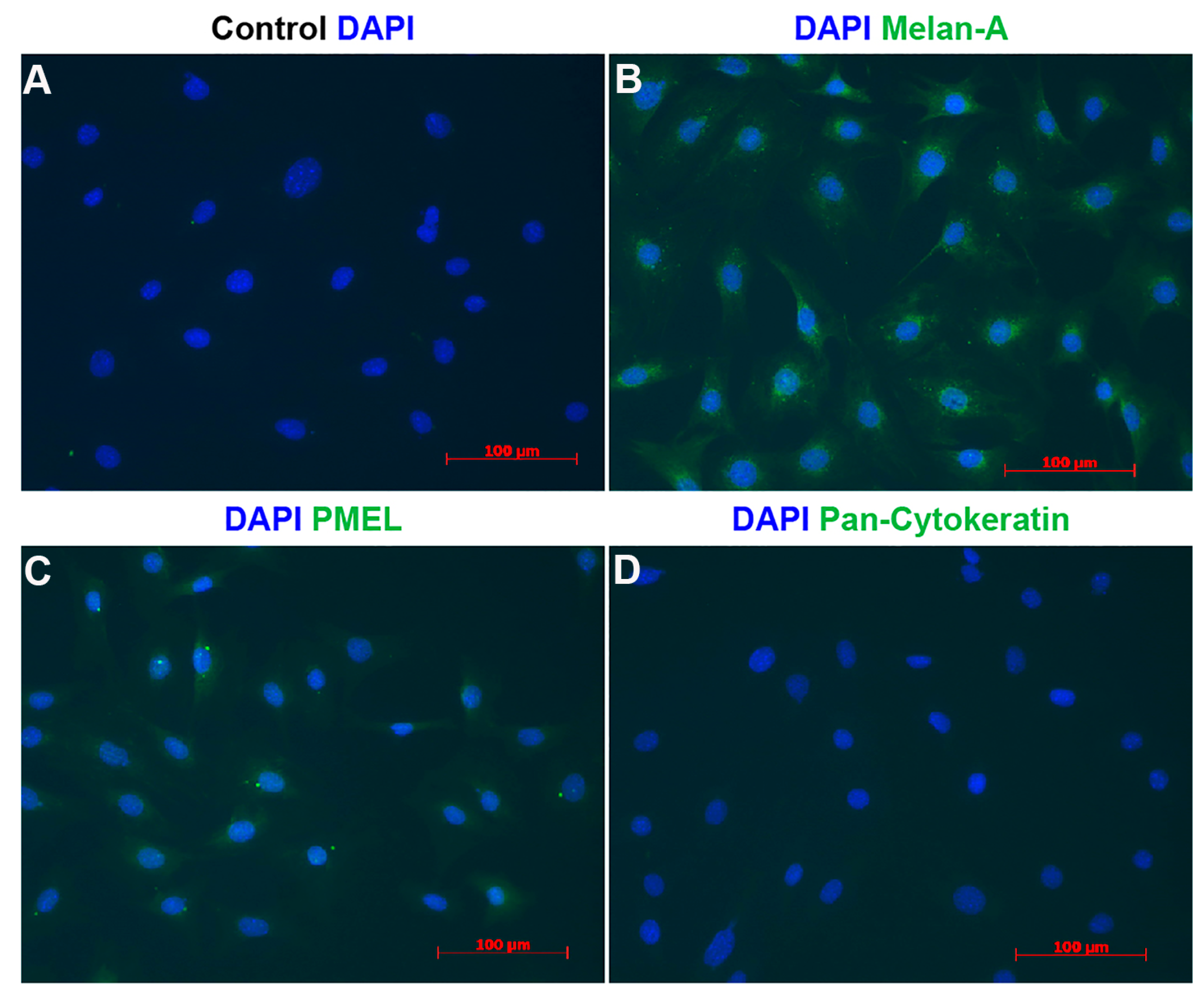
3.2. Expression of Cell Adhesion Proteins in ChMC
3.3. Localization of Melanocytes in the Choroid
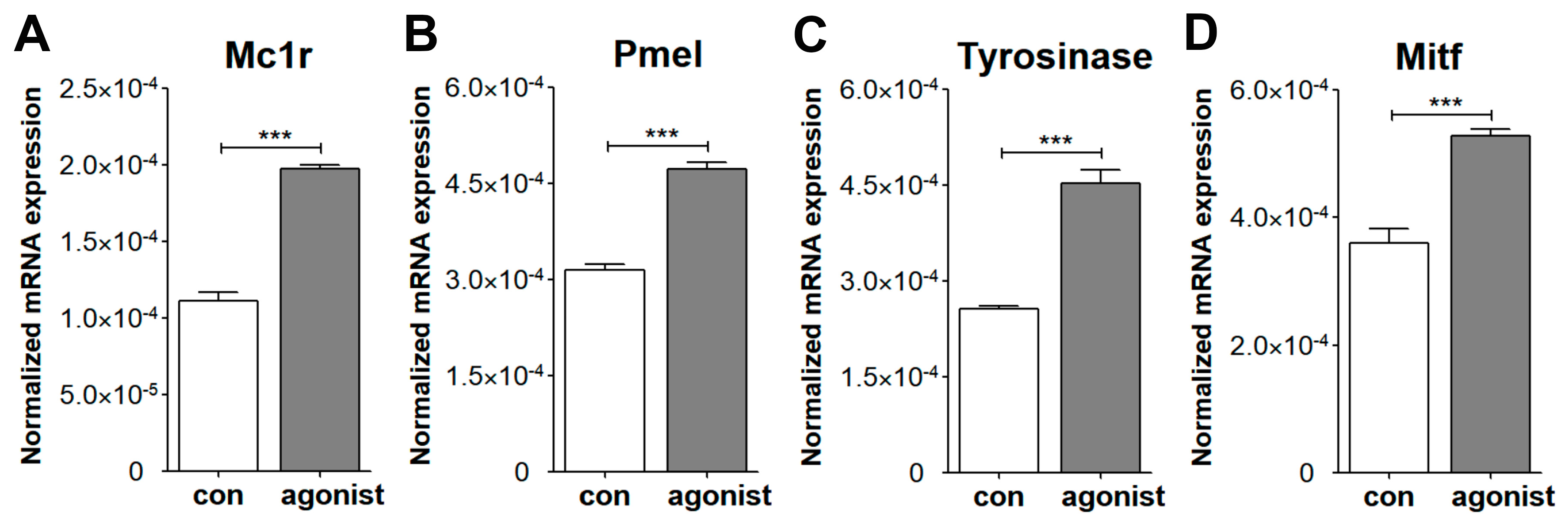
3.4. Melanogenic Responses of Mouse ChMC
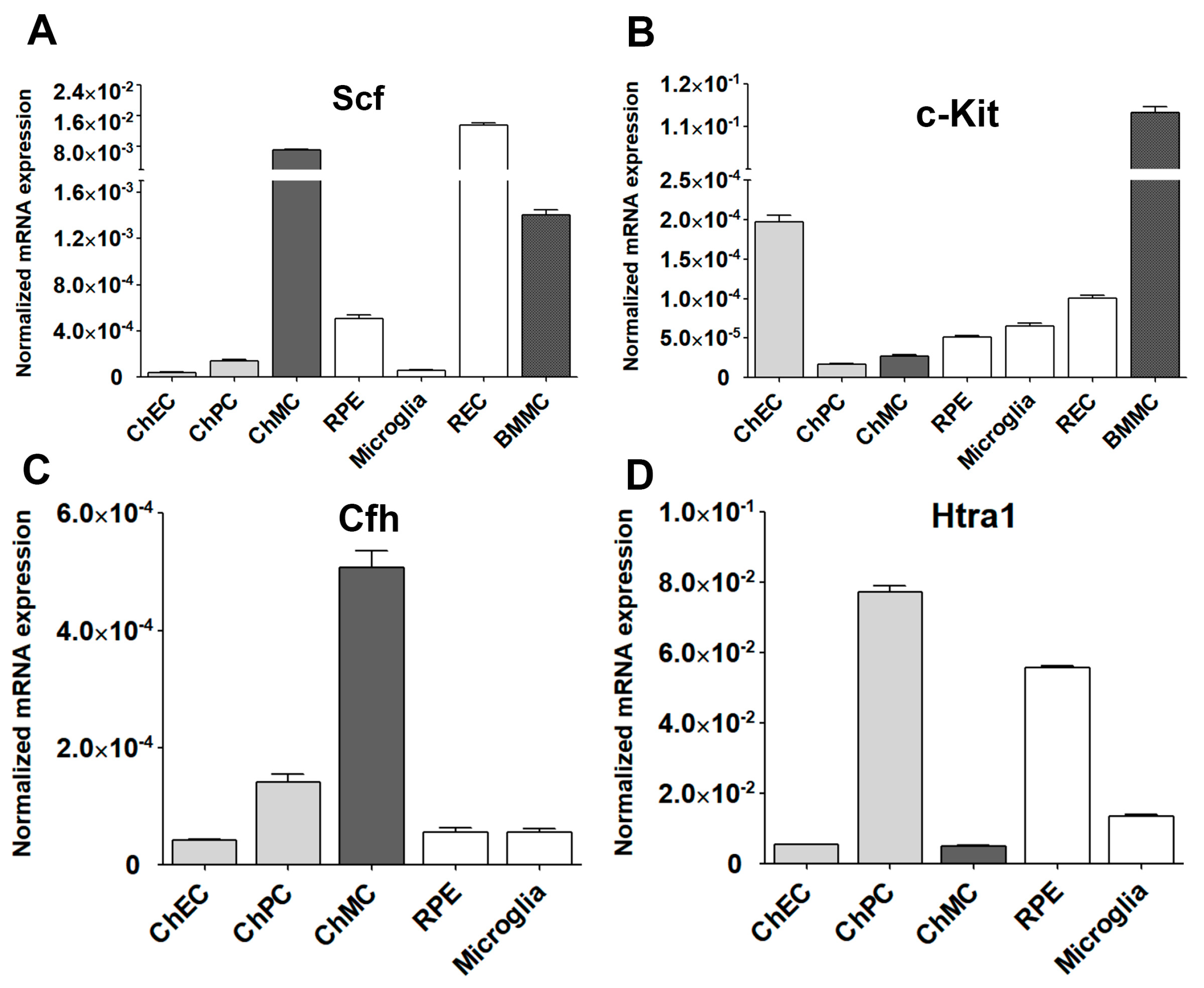
3.5. Expression of Stem Cell Factor (Scf) and Its Receptor (c-Kit) in ChMC
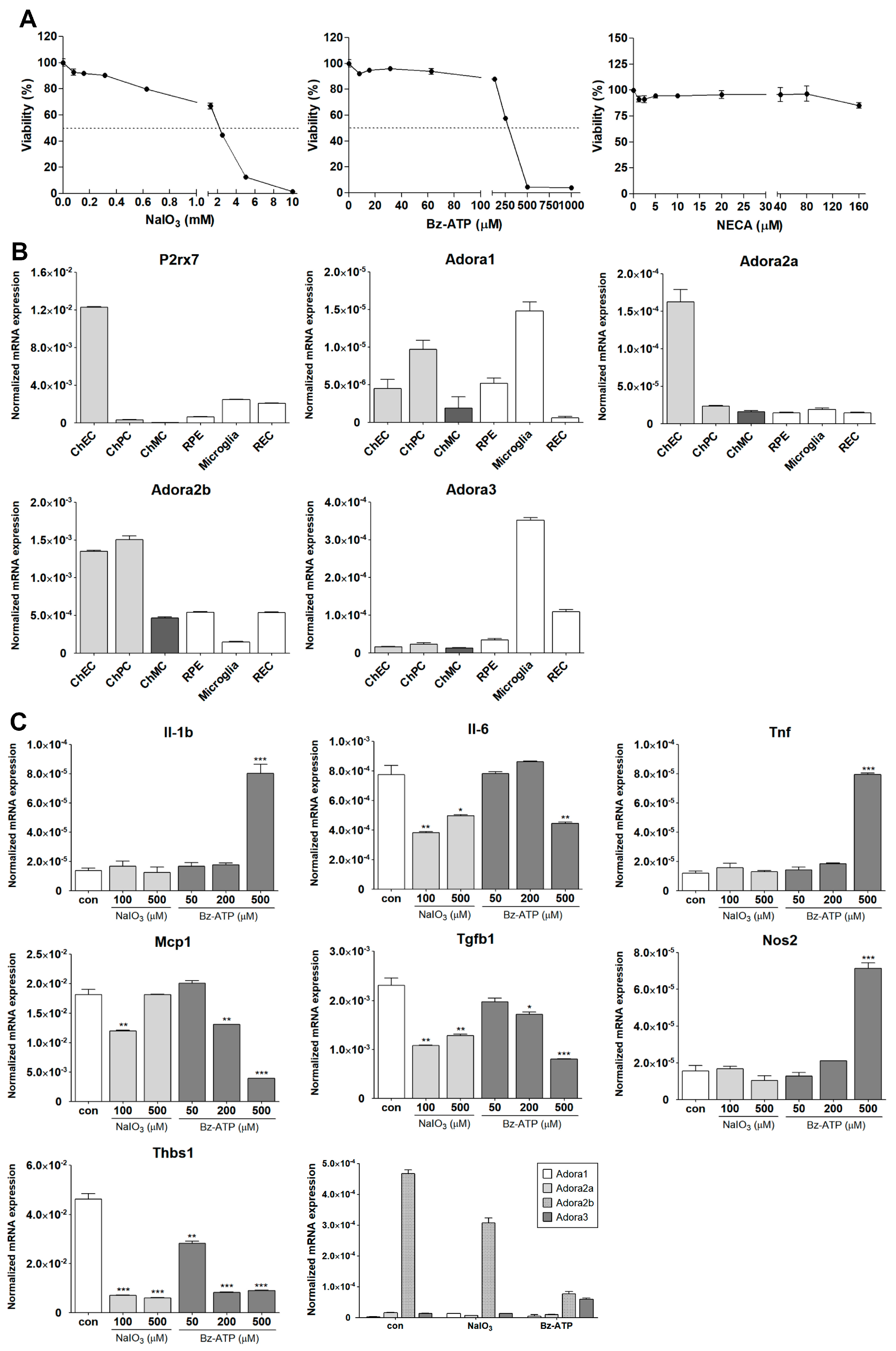
3.6. Inflammatory Responses of ChMC to Various Extracellular Stressors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shibuya, H.; Watanabe, R.; Maeno, A.; Ichimura, K.; Tamura, M.; Wakana, S.; Shiroishi, T.; Ohba, K.; Takeda, K.; Tomita, H.; et al. Melanocytes contribute to the vasculature of the choroid. Genes Genet. Syst. 2018, 93, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Adameyko, I.; Lallemend, F.; Furlan, A.; Zinin, N.; Aranda, S.; Kitambi, S.S.; Blanchart, A.; Favaro, R.; Nicolis, S.; Lübke, M.; et al. Sox2 and mitf cross-regulatory interactions consolidate progenitor and melanocyte lineages in the cranial neural crest. Development 2012, 139, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.N.; Savage, H.E.; Roberts, J.E. Uveal melanocytes, ocular pigment epithelium, and müller cells in culture: In vitro toxicology. Int. J. Toxicol. 2002, 21, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Jehs, T.; Faber, C.; Udsen, M.S.; Jager, M.J.; Clark, S.J.; Nissen, M.H. Induction of chemokine secretion and monocyte migration by human choroidal melanocytes in response to proinflammatory cytokines. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6568–6579. [Google Scholar] [CrossRef]
- Scott, G.; Leopardi, S. The camp signaling pathway has opposing effects on rac and rho in b16f10 cells: Implications for dendrite formation in melanocytic cells. Pigment Cell Res. 2003, 16, 139–148. [Google Scholar] [CrossRef]
- Hu, D.N. Methodology for evaluation of melanin content and production of pigment cells in vitro. Photochem. Photobiol. 2008, 84, 645–649. [Google Scholar] [CrossRef]
- Valtink, M.; Engelmann, K. Serum-free cultivation of adult normal human choroidal melanocytes. Graefes Arch. Clin. Exp. Ophthalmol. 2007, 245, 1487–1494. [Google Scholar] [CrossRef]
- Hu, D.N. Regulation of growth and melanogenesis of uveal melanocytes. Pigment Cell Res. 2000, 13 (Suppl. S8), 81–86. [Google Scholar] [CrossRef]
- McMenamin, P.G.; Shields, G.T.; Seyed-Razavi, Y.; Kalirai, H.; Insall, R.H.; Machesky, L.M.; Coupland, S.E. Melanoblasts populate the mouse choroid earlier in development than previously described. Investig. Ophthalmol. Vis. Sci. 2020, 61, 33. [Google Scholar] [CrossRef]
- Hu, D.N.; McCormick, S.A.; Ritch, R.; Pelton-Henrion, K. Studies of human uveal melanocytes in vitro: Isolation, purification and cultivation of human uveal melanocytes. Investig. Ophthalmol. Vis. Sci. 1993, 34, 2210–2219. [Google Scholar]
- Goodall, T.; Buffey, J.A.; Rennie, I.G.; Benson, M.; Parsons, M.A.; Faulkner, M.K.; MacNeil, S. Effect of melanocyte stimulating hormone on human cultured choroidal melanocytes, uveal melanoma cells, and retinal epithelial cells. Investig. Ophthalmol. Vis. Sci. 1994, 35, 826–837. [Google Scholar]
- Elshaw, S.R.; Sisley, K.; Cross, N.; Murray, A.K.; MacNeil, S.M.; Wagner, M.; Nichols, C.E.; Rennie, I.G. A comparison of ocular melanocyte and uveal melanoma cell invasion and the implication of alpha1beta1, alpha4beta1 and alpha6beta1 integrins. Br. J. Ophthalmol. 2001, 85, 732–738. [Google Scholar] [CrossRef]
- Harbour, J.W.; Brantley, M.A., Jr.; Hollingsworth, H.; Gordon, M. Association between choroidal pigmentation and posterior uveal melanoma in a white population. Br. J. Ophthalmol. 2004, 88, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.N.; McCormick, S.A.; Orlow, S.J.; Rosemblat, S.; Lin, A.Y.; Wo, K. Melanogenesis by human uveal melanocytes in vitro. Investig. Ophthalmol. Vis. Sci. 1995, 36, 931–938. [Google Scholar]
- Prota, G.; Hu, D.N.; Vincensi, M.R.; McCormick, S.A.; Napolitano, A. Characterization of melanins in human irides and cultured uveal melanocytes from eyes of different colors. Exp. Eye Res. 1998, 67, 293–299. [Google Scholar] [CrossRef]
- Dawson-Baglien, E.M.; Winkler, P.A.; Bruewer, A.R.; Petersen-Jones, S.M.; Bartoe, J.T. Isolation and cultivation of canine uveal melanocytes. Vet. Ophthalmol. 2015, 18, 285–290. [Google Scholar] [CrossRef]
- Hu, F.; Teramura, D.J.; Mah, K. Normal uveal melanocytes in culture. Pigment Cell Res. 1987, 1, 94–103. [Google Scholar] [CrossRef]
- Hu, F.; Mah, K. Choroidal melanocytes—A model for studying the aging process in nonreplicative differentiated cells. Mech. Ageing Dev. 1979, 11, 227–235. [Google Scholar] [CrossRef]
- Weidmann, C.; Pomerleau, J.; Trudel-Vandal, L.; Landreville, S. Differential responses of choroidal melanocytes and uveal melanoma cells to low oxygen conditions. Mol. Vis. 2017, 23, 103–115. [Google Scholar]
- Sharif, N.A.; Crider, J.Y. Human choroidal melanocyte signal transduction responses to various pharmacological agents: Focus on endothelin receptors. Curr. Eye Res. 2011, 36, 462–468. [Google Scholar] [CrossRef]
- Mouriaux, F.; Chahud, F.; Maurage, C.A.; Malecaze, F.; Labalette, P. Implication of stem cell factor in the proliferation of choroidal melanocytes. Exp. Eye Res. 2001, 73, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Cioanca, A.V.; McCluskey, P.J.; Eamegdool, S.S.; Madigan, M.C. Human choroidal melanocytes express functional toll-like receptors (tlrs). Exp. Eye Res. 2018, 173, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Sorenson, C.M.; Sheibani, N. Isolation and characterization of murine retinal endothelial cells. Mol. Vis. 2003, 9, 171–178. [Google Scholar]
- Scheef, E.; Wang, S.; Sorenson, C.M.; Sheibani, N. Isolation and characterization of murine retinal astrocytes. Mol. Vis. 2005, 11, 613–624. [Google Scholar]
- Scheef, E.A.; Sorenson, C.M.; Sheibani, N. Attenuation of proliferation and migration of retinal pericytes in the absence of thrombospondin-1. Am. J. Physiol. Cell Physiol. 2009, 296, C724–C734. [Google Scholar] [CrossRef]
- Fei, P.; Zaitoun, I.; Farnoodian, M.; Fisk, D.L.; Wang, S.; Sorenson, C.M.; Sheibani, N. Expression of thrombospondin-1 modulates the angioinflammatory phenotype of choroidal endothelial cells. PLoS ONE 2014, 9, e116423. [Google Scholar] [CrossRef]
- Farnoodian, M.; Kinter, J.B.; Yadranji Aghdam, S.; Zaitoun, I.; Sorenson, C.M.; Sheibani, N. Expression of pigment epithelium-derived factor and thrombospondin-1 regulate proliferation and migration of retinal pigment epithelial cells. Physiol. Rep. 2015, 3, e12266. [Google Scholar] [CrossRef]
- Falero-Perez, J.; Park, S.; Sorenson, C.M.; Sheibani, N. Pedf expression affects retinal endothelial cell proangiogenic properties through alterations in cell adhesive mechanisms. Am. J. Physiol. Cell Physiol. 2017, 313, C405–c420. [Google Scholar] [CrossRef]
- Lehmann, G.L.; Hanke-Gogokhia, C.; Hu, Y.; Bareja, R.; Salfati, Z.; Ginsberg, M.; Nolan, D.J.; Mendez-Huergo, S.P.; Dalotto-Moreno, T.; Wojcinski, A.; et al. Single-cell profiling reveals an endothelium-mediated immunomodulatory pathway in the eye choroid. J. Exp. Med. 2020, 217, e20190730. [Google Scholar] [CrossRef]
- Cioanca, A.V.; Wu, C.S.; Natoli, R.; Conway, R.M.; McCluskey, P.J.; Jager, M.J.; Sitiwin, E.I.; Eamegdool, S.S.; Madigan, M.C. The role of melanocytes in the human choroidal microenvironment and inflammation: Insights from the transcriptome. Pigment Cell Melanoma Res. 2021, 34, 928–945. [Google Scholar] [CrossRef]
- Farnoodian, M.; Wang, S.; Dietz, J.; Nickells, R.W.; Sorenson, C.M.; Sheibani, N. Negative regulators of angiogenesis: Important targets for treatment of exudative amd. Clin. Sci. 2017, 131, 1763–1780. [Google Scholar] [CrossRef] [PubMed]
- Toomey, C.B.; Johnson, L.V.; Bowes Rickman, C.; Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. Complement factor h in amd: Bridging genetic associations and pathobiology the impact of oxidative stress and inflammation on rpe degeneration in non-neovascular amd. Prog. Retin. Eye Res. 2018, 62, 38–57. [Google Scholar] [CrossRef]
- Hu, D.N.; Bi, M.; Zhang, D.Y.; Ye, F.; McCormick, S.A.; Chan, C.C. Constitutive and lps-induced expression of mcp-1 and il-8 by human uveal melanocytes in vitro and relevant signal pathways. Investig. Ophthalmol. Vis. Sci. 2014, 55, 5760–5769. [Google Scholar] [CrossRef]
- Zaitoun, I.S.; Song, Y.S.; Zaitoun, H.B.; Sorenson, C.M.; Sheibani, N. Assessment of choroidal vasculature and innate immune cells in the eyes of albino and pigmented mice. Cells 2022, 11, 3329. [Google Scholar] [CrossRef]
- Abdel-Malek, Z.; Swope, V.B.; Suzuki, I.; Akcali, C.; Harriger, M.D.; Boyce, S.T.; Urabe, K.; Hearing, V.J. Mitogenic and melanogenic stimulation of normal human melanocytes by melanotropic peptides. Proc. Natl. Acad. Sci. USA 1995, 92, 1789–1793. [Google Scholar] [CrossRef]
- Suzuki, I.; Cone, R.D.; Im, S.; Nordlund, J.; Abdel-Malek, Z.A. Binding of melanotropic hormones to the melanocortin receptor mc1r on human melanocytes stimulates proliferation and melanogenesis. Endocrinology 1996, 137, 1627–1633. [Google Scholar] [CrossRef]
- Coutant, K.; Magne, B.; Ferland, K.; Fuentes-Rodriguez, A.; Chancy, O.; Mitchell, A.; Germain, L.; Landreville, S. Melanocytes in regenerative medicine applications and disease modeling. J. Transl. Med. 2024, 22, 336. [Google Scholar] [CrossRef]
- Giebel, L.B.; Spritz, R.A. Mutation of the kit (mast/stem cell growth factor receptor) protooncogene in human piebaldism. Proc. Natl. Acad. Sci. USA 1991, 88, 8696–8699. [Google Scholar] [CrossRef]
- Geissler, E.N.; Ryan, M.A.; Housman, D.E. The dominant-white spotting (w) locus of the mouse encodes the c-kit proto-oncogene. Cell 1988, 55, 185–192. [Google Scholar] [CrossRef]
- Zsebo, K.M.; Williams, D.A.; Geissler, E.N.; Broudy, V.C.; Martin, F.H.; Atkins, H.L.; Hsu, R.Y.; Birkett, N.C.; Okino, K.H.; Murdock, D.C.; et al. Stem cell factor is encoded at the sl locus of the mouse and is the ligand for the c-kit tyrosine kinase receptor. Cell 1990, 63, 213–224. [Google Scholar] [CrossRef]
- Yang, Z.; Camp, N.J.; Sun, H.; Tong, Z.; Gibbs, D.; Cameron, D.J.; Chen, H.; Zhao, Y.; Pearson, E.; Li, X.; et al. A variant of the htra1 gene increases susceptibility to age-related macular degeneration. Science 2006, 314, 992–993. [Google Scholar] [CrossRef] [PubMed]
- Hageman, G.S.; Anderson, D.H.; Johnson, L.V.; Hancox, L.S.; Taiber, A.J.; Hardisty, L.I.; Hageman, J.L.; Stockman, H.A.; Borchardt, J.D.; Gehrs, K.M.; et al. A common haplotype in the complement regulatory gene factor h (hf1/cfh) predisposes individuals to age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 7227–7232. [Google Scholar] [CrossRef] [PubMed]
- Aredo, B.; Li, T.; Chen, X.; Zhang, K.; Wang, C.X.; Gou, D.; Zhao, B.; He, Y.; Ufret-Vincenty, R.L. A chimeric cfh transgene leads to increased retinal oxidative stress, inflammation, and accumulation of activated subretinal microglia in mice. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3427–3440. [Google Scholar] [CrossRef]
- Notomi, S.; Hisatomi, T.; Kanemaru, T.; Takeda, A.; Ikeda, Y.; Enaida, H.; Kroemer, G.; Ishibashi, T. Critical involvement of extracellular atp acting on p2rx7 purinergic receptors in photoreceptor cell death. Am. J. Pathol. 2011, 179, 2798–2809. [Google Scholar] [CrossRef]
- Antonioli, L.; Blandizzi, C.; Pacher, P.; Haskó, G. The purinergic system as a pharmacological target for the treatment of immune-mediated inflammatory diseases. Pharmacol. Rev. 2019, 71, 345–382. [Google Scholar] [CrossRef]
- Malih, S.; Song, Y.S.; Sorenson, C.M.; Sheibani, N. Choroidal mast cells and pathophysiology of age-related macular degeneration. Cells 2023, 13, 50. [Google Scholar] [CrossRef]
- Nasti, T.H.; Timares, L. Mc1r, eumelanin and pheomelanin: Their role in determining the susceptibility to skin cancer. Photochem. Photobiol. 2015, 91, 188–200. [Google Scholar] [CrossRef]
- Li, L.; Hu, D.N.; Zhao, H.; McCormick, S.A.; Nordlund, J.J.; Boissy, R.E. Uveal melanocytes do not respond to or express receptors for alpha-melanocyte-stimulating hormone. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4507–4512. [Google Scholar] [CrossRef]
- Smith-Thomas, L.; Richardson, P.; Thody, A.J.; Graham, A.; Palmer, I.; Flemming, L.; Parsons, M.A.; Rennie, I.G.; MacNeil, S. Human ocular melanocytes and retinal pigment epithelial cells differ in their melanogenic properties in vivo and in vitro. Curr. Eye Res. 1996, 15, 1079–1091. [Google Scholar] [CrossRef]
- Kiuchi, K.; Yoshizawa, K.; Shikata, N.; Moriguchi, K.; Tsubura, A. Morphologic characteristics of retinal degeneration induced by sodium iodate in mice. Curr. Eye Res. 2002, 25, 373–379. [Google Scholar] [CrossRef]
- Hariri, S.; Tam, M.C.; Lee, D.; Hileeto, D.; Moayed, A.A.; Bizheva, K. Noninvasive imaging of the early effect of sodium iodate toxicity in a rat model of outer retina degeneration with spectral domain optical coherence tomography. J. Biomed. Opt. 2013, 18, 26017. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Wang, C.; Zhang, Y.; Su, G. Morphologic and histopathologic change of sodium iodate-induced retinal degeneration in adult rats. Int. J. Clin. Exp. Pathol. 2019, 12, 443–454. [Google Scholar] [PubMed]
- Zhao, S.; Overbeek, P.A. Regulation of choroid development by the retinal pigment epithelium. Mol. Vis. 2001, 7, 277–282. [Google Scholar] [PubMed]
- Notomi, S.; Hisatomi, T.; Murakami, Y.; Terasaki, H.; Sonoda, S.; Asato, R.; Takeda, A.; Ikeda, Y.; Enaida, H.; Sakamoto, T.; et al. Dynamic increase in extracellular atp accelerates photoreceptor cell apoptosis via ligation of p2rx7 in subretinal hemorrhage. PLoS ONE 2013, 8, e53338. [Google Scholar] [CrossRef]
- Auchampach, J.A. Adenosine receptors and angiogenesis. Circ. Res. 2007, 101, 1075–1077. [Google Scholar] [CrossRef]
- Sorenson, C.M.; Song, Y.-S.; Zaitoun, I.S.; Wang, S.; Hanna, B.A.; Darjatmoko, S.R.; Gurel, Z.; Fisk, D.L.; McDowell, C.M.; McAdams, R.M.; et al. Caffeine inhibits choroidal neovascularization through mitigation of inflammatory and angiogenesis activities. Front. Cell Dev. Biol. 2021, 9, 737426. [Google Scholar] [CrossRef]
- Wang, S.; Sorenson, C.M.; Sheibani, N. Lack of thrombospondin 1 and exacerbation of choroidal neovascularization. Arch. Ophthalmol. 2012, 130, 615–620. [Google Scholar] [CrossRef]
- Rani, R.; Smulian, A.G.; Greaves, D.R.; Hogan, S.P.; Herbert, D.R. Tgf-β limits il-33 production and promotes the resolution of colitis through regulation of macrophage function. Eur. J. Immunol. 2011, 41, 2000–2009. [Google Scholar] [CrossRef]
- Berkowitz, B.A.; Podolsky, R.H.; Lenning, J.; Khetarpal, N.; Tran, C.; Wu, J.Y.; Berri, A.M.; Dernay, K.; Shafie-Khorassani, F.; Roberts, R. Sodium iodate produces a strain-dependent retinal oxidative stress response measured in vivo using quest mri. Investig. Ophthalmol. Vis. Sci. 2017, 58, 3286–3293. [Google Scholar] [CrossRef]
- Chowers, G.; Cohen, M.; Marks-Ohana, D.; Stika, S.; Eijzenberg, A.; Banin, E.; Obolensky, A. Course of sodium iodate-induced retinal degeneration in albino and pigmented mice. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2239–2249. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward 5′ to 3′ | Reverse 5′ to 3′ |
|---|---|---|
| Adora1 | GTCAAGATCCCTCTCCGGTA | CAAGGGAGAGAATCCAGCAG |
| Adora2a | GGTCCTCACGCAGAGTTCC | TCACCAAGCCATTGTACCG |
| Adora2b | CCGATATCTGGCCATTCG | AGTCAATCCAATGCCAAAGG |
| Adora3 | CTCTTTGCTAGGATTGCTTGG | AGAAGGAATGCCAAGAGCAG |
| Cd31 | GTGTGGAAGCCAACAGCCA | TCCATTAAGGGAGCCTTCCG |
| Cfh | GGGCAAGTGGAAGTGATGTG | TGTCAATAGGTGGAGGAGGC |
| c-kit | GAAGGACTCCTCCTGCTTTAGA | CAAATACACTCAGGGGAGCAC |
| Htra1 | GGATGTGGATGAAAAGGCGG | AATTCTCCAGGTCTCAGCTCTG |
| Il-1b | GTTCCCATTAGACAACTGCACT | CCGACAGCACGAGGCTTTT |
| Il-6 | CAACCACGGCCTTCCCTACT | TTGGGAGTGGTATCCTCTGTGA |
| Mc1r | CTACAAGCACACAGCCGTTC | AGTGCCAGCATGGCTAGAAA |
| Mcp1 | GTCTGTGCTGACCCCAAGAAG | TGGTTCCGATCCAGGTTTTTA |
| Mitf | GGTGACAACATAGGGAATGGTT | CCAGTCCCTGAAGAATCCA |
| Nos2 | GGCAGCCTGTGAGACCTTTG | CATTGGAAGTGAAGCGTTTCG |
| P2rx7 | CGTCTTTTCCTACATTAGCTTTGC | ATGCCTTTGACCTTGGTGTG |
| Pdgfrb | ATCGCGCCACCTTAATCAAC | GCTAAGAAGTCCATGCCGTT |
| Pmel | CTCTTGTTTCCTGTGGTTCCT | GTAGTGGTTCCTTGCCTAGATG |
| Rpe65 | CCTGGTTCTGAATGCCAAAG | CATGGAAGGTCACAGGGATATT |
| SCF | AATGAATGGAAAAATCTGTTGTGTAA | TGCGTACAGAATAGCTAAGATTTCA |
| Tgfb1 | GCAGTGGCTGAACCAAGGA | AGCAGTGAGCGCTGAATCG |
| Thbs1 | TGGCCAGCGTTGCCA | TCTGCAGCACCCCCTGAA |
| Tnf | ACCGTCAGCCGATTTGCTAT | TTGACGGCAGAGAGGAGGTT |
| Tyrosinase | GGGATTGGAGAGATGCAGAAA | TCTGCCAGGAGGAGAAGAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.-S.; Park, S.; Fisk, D.; Sorenson, C.M.; Sheibani, N. Isolation and Characterization of Mouse Choroidal Melanocytes and Their Proinflammatory Characteristics. Cells 2025, 14, 646. https://doi.org/10.3390/cells14090646
Song Y-S, Park S, Fisk D, Sorenson CM, Sheibani N. Isolation and Characterization of Mouse Choroidal Melanocytes and Their Proinflammatory Characteristics. Cells. 2025; 14(9):646. https://doi.org/10.3390/cells14090646
Chicago/Turabian StyleSong, Yong-Seok, SunYoung Park, Debra Fisk, Christine M. Sorenson, and Nader Sheibani. 2025. "Isolation and Characterization of Mouse Choroidal Melanocytes and Their Proinflammatory Characteristics" Cells 14, no. 9: 646. https://doi.org/10.3390/cells14090646
APA StyleSong, Y.-S., Park, S., Fisk, D., Sorenson, C. M., & Sheibani, N. (2025). Isolation and Characterization of Mouse Choroidal Melanocytes and Their Proinflammatory Characteristics. Cells, 14(9), 646. https://doi.org/10.3390/cells14090646