

Rejuvenated Autologous Adult Stem Cells: Emerging Front Runners in the Fight Against Aging and Associated Diseases

Abstract
1. Introduction
2. Aging and Currently Prevailing Anti-Aging Measures
3. Very Small Embryonic-Like (VSEL) Stem Cells
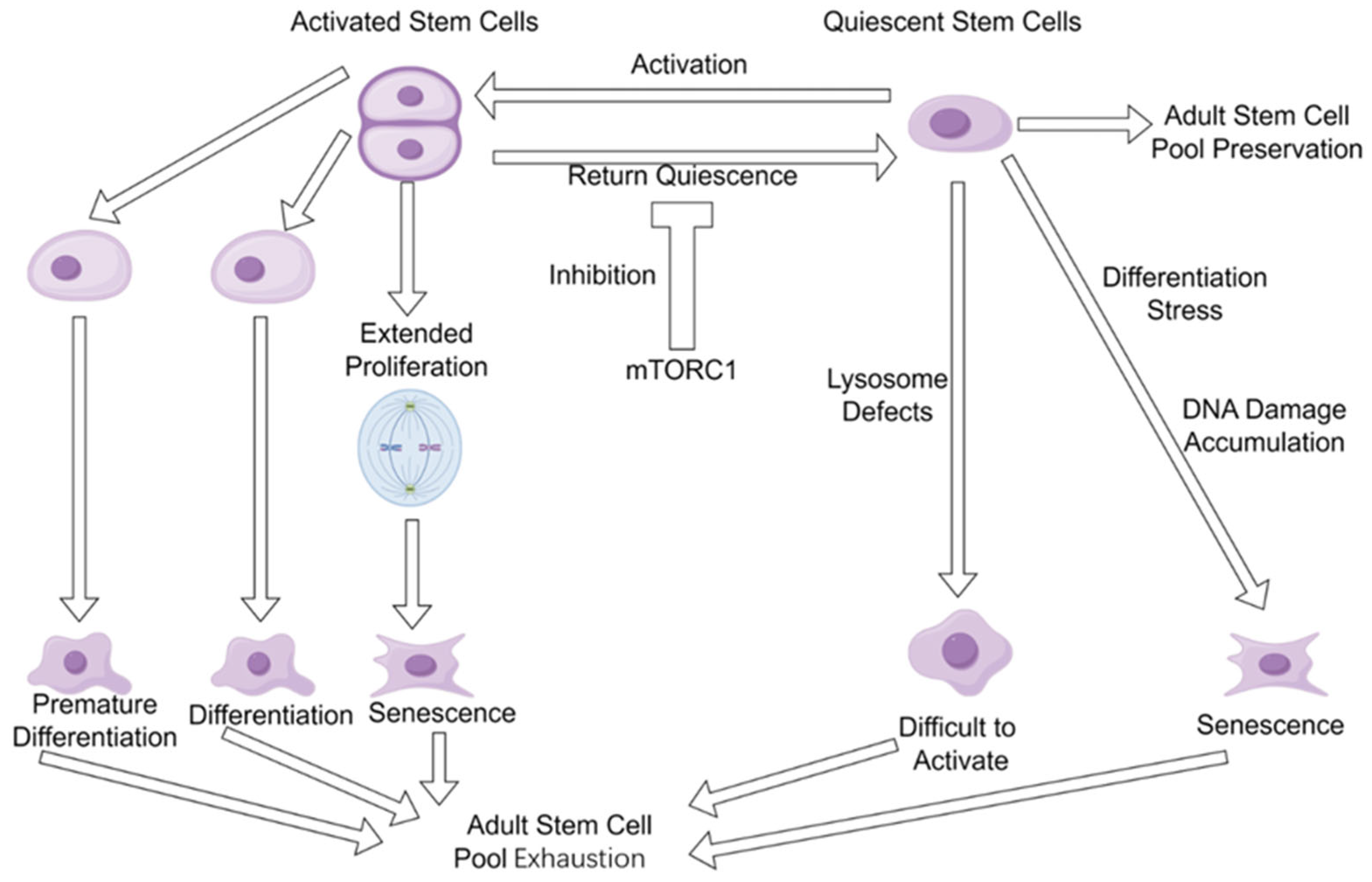
4. Adult Stem Cell Pool Exhaustion Is Inevitable During Natural Aging
5. Adult Stem Cell Transplantation
6. Dedifferentiation and Rejuvenation
7. Challenges of Gaining and Using Rejuvenated Adult Stem Cells
8. Summary
Author Contributions
Funding
Conflicts of Interest
References
- Boehm, A.M.; Khalturin, K.; Anton-Erxleben, F.; Hemmrich, G.; Klostermeier, U.C.; Lopez-Quintero, J.A.; Oberg, H.H.; Puchert, M.; Rosenstiel, P.; Wittlieb, J.; et al. FoxO is a critical regulator of stem cell maintenance in immortal Hydra. Proc. Natl. Acad. Sci. USA 2012, 109, 19697–19702. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Shen, J.; Tsai, Y.T.; Dimarco, N.M.; Long, M.A.; Sun, X.; Tang, L. Transplantation of mesenchymal stem cells from young donors delays aging in mice. Sci. Rep. 2011, 1, 67. [Google Scholar] [CrossRef]
- Guderyon, M.J.; Chen, C.; Bhattacharjee, A.; Ge, G.; Fernandez, R.A.; Gelfond, J.A.L.; Gorena, K.M.; Cheng, C.J.; Li, Y.; Nelson, J.F.; et al. Mobilization-based transplantation of young-donor hematopoietic stem cells extends lifespan in mice. Aging Cell 2020, 19, e13110. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Wu, S.; Li, R.; Huang, K.; Zeng, T.; Zhou, Z.; Zhong, X.; Songyang, Z.; Liu, F. mTORC1-Induced Bone Marrow-Derived Mesenchymal Stem Cell Exhaustion Contributes to the Bone Abnormalities in Klotho-Deficient Mice of Premature Aging. Stem Cells Dev. 2023, 32, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Haller, S.; Kapuria, S.; Riley, R.R.; O’Leary, M.N.; Schreiber, K.H.; Andersen, J.K.; Melov, S.; Que, J.; Rando, T.A.; Rock, J.; et al. mTORC1 Activation during Repeated Regeneration Impairs Somatic Stem Cell Maintenance. Cell Stem Cell 2017, 21, 806–818.e5. [Google Scholar] [CrossRef]
- Bose, G.; Atkins, H.L.; Bowman, M.; Freedman, M.S. Autologous hematopoietic stem cell transplantation improves fatigue in multiple sclerosis. Mult. Scler. 2019, 25, 1764–1772. [Google Scholar] [CrossRef] [PubMed]
- Genchi, A.; Brambilla, E.; Sangalli, F.; Radaelli, M.; Bacigaluppi, M.; Furlan, R.; Andolfo, A.; Drago, D.; Magagnotti, C.; Scotti, G.M.; et al. Neural stem cell transplantation in patients with progressive multiple sclerosis: An open-label, phase 1 study. Nat. Med. 2023, 29, 75–85. [Google Scholar] [CrossRef]
- Ghobrial, G.M.; Anderson, K.D.; Dididze, M.; Martinez-Barrizonte, J.; Sunn, G.H.; Gant, K.L.; Levi, A.D. Human Neural Stem Cell Transplantation in Chronic Cervical Spinal Cord Injury: Functional Outcomes at 12 Months in a Phase II Clinical Trial. Neurosurgery 2017, 64, 87–91. [Google Scholar] [CrossRef]
- Izadi, M.; Sadr Hashemi Nejad, A.; Moazenchi, M.; Masoumi, S.; Rabbani, A.; Kompani, F.; Hedayati Asl, A.A.; Abbasi Kakroodi, F.; Jaroughi, N.; Mohseni Meybodi, M.A.; et al. Mesenchymal stem cell transplantation in newly diagnosed type-1 diabetes patients: A phase I/II randomized placebo-controlled clinical trial. Stem Cell Res. Ther. 2022, 13, 264. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, M.; Wang, F.; Heng, B.C.; Zhou, J.; Cai, Z.; Liu, H. Understanding the Immunological Mechanisms of Mesenchymal Stem Cells in Allogeneic Transplantation: From the Aspect of Major Histocompatibility Complex Class I. Stem Cells Dev. 2019, 28, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Ziada, A.S.; Smith, M.R.; Côté, H.C.F. Updating the Free Radical Theory of Aging. Front. Cell Dev. Biol. 2020, 8, 575645. [Google Scholar] [CrossRef] [PubMed]
- Blagosklonny, M.V. Rapamycin treatment early in life reprograms aging: Hyperfunction theory and clinical practice. Aging 2022, 14, 8140–8149. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.R.; Tian, X.; Sinclair, D.A. The Information Theory of Aging. Nat. Aging 2023, 3, 1486–1499. [Google Scholar] [CrossRef]
- Yang, J.H.; Hayano, M.; Griffin, P.T.; Amorim, J.A.; Bonkowski, M.S.; Apostolides, J.K.; Salfati, E.L.; Blanchette, M.; Munding, E.M.; Bhakta, M.; et al. Loss of epigenetic information as a cause of mammalian aging. Cell 2023, 186, 305–326.e27. [Google Scholar] [CrossRef]
- Amor, C.; Feucht, J.; Leibold, J.; Ho, Y.J.; Zhu, C.; Alonso-Curbelo, D.; Mansilla-Soto, J.; Boyer, J.A.; Li, X.; Giavridis, T.; et al. Senolytic CAR T cells reverse senescence-associated pathologies. Nature 2020, 583, 127–132. [Google Scholar] [CrossRef]
- Bitto, A.; Ito, T.K.; Pineda, V.V.; LeTexier, N.J.; Huang, H.Z.; Sutlief, E.; Tung, H.; Vizzini, N.; Chen, B.; Smith, K.; et al. Transient rapamycin treatment can increase lifespan and healthspan in middle-aged mice. Elife 2016, 5, e16351. [Google Scholar] [CrossRef]
- Harrison, D.E.; Strong, R.; Sharp, Z.D.; Nelson, J.F.; Astle, C.M.; Flurkey, K.; Nadon, N.L.; Wilkinson, J.E.; Frenkel, K.; Carter, C.S.; et al. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature 2009, 460, 392–395. [Google Scholar] [CrossRef]
- Miller, R.A.; Harrison, D.E.; Astle, C.M.; Baur, J.A.; Boyd, A.R.; de Cabo, R.; Fernandez, E.; Flurkey, K.; Javors, M.A.; Nelson, J.F.; et al. Rapamycin, but not resveratrol or simvastatin, extends life span of genetically heterogeneous mice. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66, 191–201. [Google Scholar] [CrossRef]
- Miller, R.A.; Harrison, D.E.; Astle, C.M.; Fernandez, E.; Flurkey, K.; Han, M.; Javors, M.A.; Li, X.; Nadon, N.L.; Nelson, J.F.; et al. Rapamycin-mediated lifespan increase in mice is dose and sex dependent and metabolically distinct from dietary restriction. Aging Cell 2014, 13, 468–477. [Google Scholar] [CrossRef]
- Neff, F.; Flores-Dominguez, D.; Ryan, D.P.; Horsch, M.; Schröder, S.; Adler, T.; Afonso, L.C.; Aguilar-Pimentel, J.A.; Becker, L.; Garrett, L.; et al. Rapamycin extends murine lifespan but has limited effects on aging. J. Clin. Investig. 2013, 123, 3272–3291. [Google Scholar] [CrossRef]
- Chaveroux, C.; Eichner, L.J.; Dufour, C.R.; Shatnawi, A.; Khoutorsky, A.; Bourque, G.; Sonenberg, N.; Giguère, V. Molecular and genetic crosstalks between mTOR and ERRα are key determinants of rapamycin-induced nonalcoholic fatty liver. Cell Metab. 2013, 17, 586–598. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.; Ma, C.; Cui, J.; Dong, X.; Sun, L.; Li, Y.; Yu, A. Rapamycin suppresses inflammation and increases the interaction between p65 and IκBα in rapamycin-induced fatty livers. PLoS ONE 2023, 18, e0281888. [Google Scholar] [CrossRef]
- Houde, V.P.; Brûlé, S.; Festuccia, W.T.; Blanchard, P.G.; Bellmann, K.; Deshaies, Y.; Marette, A. Chronic rapamycin treatment causes glucose intolerance and hyperlipidemia by upregulating hepatic gluconeogenesis and impairing lipid deposition in adipose tissue. Diabetes 2010, 59, 1338–1348. [Google Scholar] [CrossRef]
- Shi, G.; Ozog, S.; Torbett, B.E.; Compton, A.A. mTOR inhibitors lower an intrinsic barrier to virus infection mediated by IFITM3. Proc. Natl. Acad. Sci. USA 2018, 115, E10069–E10078. [Google Scholar] [CrossRef]
- Martin-Montalvo, A.; Mercken, E.M.; Mitchell, S.J.; Palacios, H.H.; Mote, P.L.; Scheibye-Knudsen, M.; Gomes, A.P.; Ward, T.M.; Minor, R.K.; Blouin, M.J.; et al. Metformin improves healthspan and lifespan in mice. Nat. Commun. 2013, 4, 2192. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.; Fleming, G.A.; Chen, K.; Bicsak, T.A. Metformin-associated lactic acidosis: Current perspectives on causes and risk. Metabolism 2016, 65, 20–29. [Google Scholar] [CrossRef]
- Lalau, J.D. Lactic acidosis induced by metformin: Incidence, management and prevention. Drug Saf. 2010, 33, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Palliyaguru, D.L.; Minor, R.K.; Mitchell, S.J.; Palacios, H.H.; Licata, J.J.; Ward, T.M.; Abulwerdi, G.; Elliott, P.; Westphal, C.; Ellis, J.L.; et al. Combining a High Dose of Metformin With the SIRT1 Activator, SRT1720, Reduces Life Span in Aged Mice Fed a High-Fat Diet. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 2037–2041. [Google Scholar] [CrossRef]
- Loffredo, F.S.; Steinhauser, M.L.; Jay, S.M.; Gannon, J.; Pancoast, J.R.; Yalamanchi, P.; Sinha, M.; Dall’Osso, C.; Khong, D.; Shadrach, J.L.; et al. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Cell 2013, 153, 828–839. [Google Scholar] [CrossRef] [PubMed]
- Sinha, M.; Jang, Y.C.; Oh, J.; Khong, D.; Wu, E.Y.; Manohar, R.; Miller, C.; Regalado, S.G.; Loffredo, F.S.; Pancoast, J.R.; et al. Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle. Science 2014, 344, 649–652. [Google Scholar] [CrossRef]
- Egerman, M.A.; Cadena, S.M.; Gilbert, J.A.; Meyer, A.; Nelson, H.N.; Swalley, S.E.; Mallozzi, C.; Jacobi, C.; Jennings, L.L.; Clay, I.; et al. GDF11 Increases with Age and Inhibits Skeletal Muscle Regeneration. Cell Metab. 2015, 22, 164–174. [Google Scholar] [CrossRef]
- Smith, S.C.; Zhang, X.; Zhang, X.; Gross, P.; Starosta, T.; Mohsin, S.; Franti, M.; Gupta, P.; Hayes, D.; Myzithras, M.; et al. GDF11 does not rescue aging-related pathological hypertrophy. Circ. Res. 2015, 117, 926–932. [Google Scholar] [CrossRef]
- Kucia, M.; Wysoczynski, M.; Ratajczak, J.; Ratajczak, M.Z. Identification of very small embryonic like (VSEL) stem cells in bone marrow. Cell Tissue Res. 2008, 331, 125–134. [Google Scholar] [CrossRef]
- Ratajczak, M.Z.; Zuba-Surma, E.K.; Shin, D.M.; Ratajczak, J.; Kucia, M. Very small embryonic-like (VSEL) stem cells in adult organs and their potential role in rejuvenation of tissues and longevity. Exp. Gerontol. 2008, 43, 1009–1017. [Google Scholar] [CrossRef]
- Kim, Y.; Jeong, J.; Kang, H.; Lim, J.; Heo, J.; Ratajczak, J.; Ratajczak, M.Z.; Shin, D.M. The molecular nature of very small embryonic-like stem cells in adult tissues. Int. J. Stem Cells 2014, 7, 55–62. [Google Scholar] [CrossRef]
- Havens, A.M.; Shiozawa, Y.; Jung, Y.; Sun, H.; Wang, J.; McGee, S.; Mishra, A.; Taichman, L.S.; Danciu, T.; Jiang, Y.; et al. Human very small embryonic-like cells generate skeletal structures, in vivo. Stem Cells Dev. 2013, 22, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Havens, A.M.; Sun, H.; Shiozawa, Y.; Jung, Y.; Wang, J.; Mishra, A.; Jiang, Y.; O’Neill, D.W.; Krebsbach, P.H.; Rodgerson, D.O.; et al. Human and murine very small embryonic-like cells represent multipotent tissue progenitors, in vitro and in vivo. Stem Cells Dev. 2014, 23, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Zuba-Surma, E.K.; Guo, Y.; Taher, H.; Sanganalmath, S.K.; Hunt, G.; Vincent, R.J.; Kucia, M.; Abdel-Latif, A.; Tang, X.L.; Ratajczak, M.Z.; et al. Transplantation of expanded bone marrow-derived very small embryonic-like stem cells (VSEL-SCs) improves left ventricular function and remodelling after myocardial infarction. J. Cell. Mol. Med. 2011, 15, 1319–1328. [Google Scholar] [CrossRef]
- de Morree, A.; Rando, T.A. Regulation of adult stem cell quiescence and its functions in the maintenance of tissue integrity. Nat. Rev. Mol. Cell Biol. 2023, 24, 334–354. [Google Scholar] [CrossRef]
- Urbán, N.; Cheung, T.H. Stem cell quiescence: The challenging path to activation. Development 2021, 148, dev165084. [Google Scholar] [CrossRef]
- Kim, J.H.; Han, G.C.; Seo, J.Y.; Park, I.; Park, W.; Jeong, H.W.; Lee, S.H.; Bae, S.H.; Seong, J.; Yum, M.K.; et al. Sex hormones establish a reserve pool of adult muscle stem cells. Nat. Cell Biol. 2016, 18, 930–940. [Google Scholar] [CrossRef]
- Kalamakis, G.; Brüne, D.; Ravichandran, S.; Bolz, J.; Fan, W.; Ziebell, F.; Stiehl, T.; Catalá-Martinez, F.; Kupke, J.; Zhao, S.; et al. Quiescence Modulates Stem Cell Maintenance and Regenerative Capacity in the Aging Brain. Cell 2019, 176, 1407–1419.e14. [Google Scholar] [CrossRef] [PubMed]
- Leeman, D.S.; Hebestreit, K.; Ruetz, T.; Webb, A.E.; McKay, A.; Pollina, E.A.; Dulken, B.W.; Zhao, X.; Yeo, R.W.; Ho, T.T.; et al. Lysosome activation clears aggregates and enhances quiescent neural stem cell activation during aging. Science 2018, 359, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Beerman, I.; Seita, J.; Inlay, M.A.; Weissman, I.L.; Rossi, D.J. Quiescent hematopoietic stem cells accumulate DNA damage during aging that is repaired upon entry into cell cycle. Cell Stem Cell 2014, 15, 37–50. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, Y.; Min, Y.; Guo, K.; Chen, Y.; Huang, Z.; Long, C. Effects and Mechanisms of Bone Marrow Mesenchymal Stem Cell Transplantation for Treatment of Ischemic Stroke in Hypertensive Rats. Int. J. Stem Cells 2022, 15, 217–226. [Google Scholar] [CrossRef]
- Klein, B.; Ciesielska, A.; Losada, P.M.; Sato, A.; Shah-Morales, S.; Ford, J.B.; Higashikubo, B.; Tager, D.; Urry, A.; Bombosch, J.; et al. Modified human mesenchymal stromal/stem cells restore cortical excitability after focal ischemic stroke in rats. Mol. Ther. 2025, 33, 375–400. [Google Scholar] [CrossRef] [PubMed]
- Ercelen, N.; Karasu, N.; Kahyaoglu, B.; Cerezci, O.; Akduman, R.C.; Ercelen, D.; Erturk, G.; Gulay, G.; Alpaydin, N.; Boyraz, G.; et al. Clinical experience: Outcomes of mesenchymal stem cell transplantation in five stroke patients. Front. Med. 2023, 10, 1051831. [Google Scholar] [CrossRef]
- Wang, P.; Liu, P.; Ding, Y.; Zhang, G.; Wang, N.; Sun, X.; Li, M.; Li, M.; Bao, X.; Chen, X. Transplantation of human neural stem cells repairs neural circuits and restores neurological function in the stroke-injured brain. Neural Regen. Res. 2024, 21, 1162–1171. [Google Scholar] [CrossRef]
- Chen, K.S.; Noureldein, M.H.; McGinley, L.M.; Hayes, J.M.; Rigan, D.M.; Kwentus, J.F.; Mason, S.N.; Mendelson, F.E.; Savelieff, M.G.; Feldman, E.L. Human neural stem cells restore spatial memory in a transgenic Alzheimer’s disease mouse model by an immunomodulating mechanism. Front. Aging Neurosci. 2023, 15, 1306004. [Google Scholar] [CrossRef]
- Ji, Q.; Lv, Y.; Hu, B.; Su, Y.; Shaikh, I.I.; Zhu, X. Study on the therapeutic potential of induced neural stem cells for Alzheimer’s disease in mice. Biol. Res. 2024, 57, 89. [Google Scholar] [CrossRef]
- Santana-Gonçalves, M.; De Santis, P.B.; Malmegrim, K.C.R.; Oliveira, M.C. T-cell Recovery After Autologous Hematopoietic Stem Cell Transplantation in Autoimmune Diseases. Adv. Exp. Med. Biol. 2025, 1471, 301–323. [Google Scholar] [CrossRef]
- Tompkins, B.A.; DiFede, D.L.; Khan, A.; Landin, A.M.; Schulman, I.H.; Pujol, M.V.; Heldman, A.W.; Miki, R.; Goldschmidt-Clermont, P.J.; Goldstein, B.J.; et al. Allogeneic Mesenchymal Stem Cells Ameliorate Aging Frailty: A Phase II Randomized, Double-Blind, Placebo-Controlled Clinical Trial. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2017, 72, 1513–1522. [Google Scholar] [CrossRef]
- Myers, M.I.; Hines, K.J.; Gray, A.; Spagnuolo, G.; Rosenwasser, R.; Iacovitti, L. Intracerebral Transplantation of Autologous Mesenchymal Stem Cells Improves Functional Recovery in a Rat Model of Chronic Ischemic Stroke. Transl. Stroke Res. 2025, 16, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Cheng, J.H.; Wang, C.J.; Ko, J.Y.; Hsu, S.L.; Hsu, T.C. Shockwave Therapy Combined with Autologous Adipose-Derived Mesenchymal Stem Cells Is Better than with Human Umbilical Cord Wharton’s Jelly-Derived Mesenchymal Stem Cells on Knee Osteoarthritis. Int. J. Mol. Sci. 2020, 21, 1217. [Google Scholar] [CrossRef]
- Ma, Z.X.; Wu, X.F.; Cao, L.; Jiao, C.Y.; Ma, D.P.; Zhao, Y.H.; Yang, Z.X.; Hu, M. Regenerative fibroblasts derived from autologous skin tissue for the treatment of Sjögren’s syndrome: A case report. Front. Immunol. 2025, 16, 1529883. [Google Scholar] [CrossRef] [PubMed]
- Doi, H.; Kitajima, Y.; Luo, L.; Yan, C.; Tateishi, S.; Ono, Y.; Urata, Y.; Goto, S.; Mori, R.; Masuzaki, H.; et al. Potency of umbilical cord blood- and Wharton’s jelly-derived mesenchymal stem cells for scarless wound healing. Sci. Rep. 2016, 6, 18844. [Google Scholar] [CrossRef] [PubMed]
- Warwick, R.; Armitage, S. Cord blood banking. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 995–1011. [Google Scholar] [CrossRef]
- Liang, H.; Suo, H.; Wang, Z.; Feng, W. Progress in the treatment of osteoarthritis with umbilical cord stem cells. Hum. Cell 2020, 33, 470–475. [Google Scholar] [CrossRef]
- Attar, A.; Farjoud Kouhanjani, M.; Hessami, K.; Vosough, M.; Kojuri, J.; Ramzi, M.; Hosseini, S.A.; Faghih, M.; Monabati, A. Effect of once versus twice intracoronary injection of allogeneic-derived mesenchymal stromal cells after acute myocardial infarction: BOOSTER-TAHA7 randomized clinical trial. Stem Cell Res. Ther. 2023, 14, 264. [Google Scholar] [CrossRef]
- Wu, K.J.; Yu, S.J.; Chiang, C.W.; Lee, Y.W.; Yen, B.L.; Tseng, P.C.; Hsu, C.S.; Kuo, L.W.; Wang, Y. Neuroprotective Action of Human Wharton’s Jelly-Derived Mesenchymal Stromal Cell Transplants in a Rodent Model of Stroke. Cell Transplant. 2018, 27, 1603–1612. [Google Scholar] [CrossRef]
- Jahan, S.; Kaushal, R.; Pasha, R.; Pineault, N. Current and Future Perspectives for the Cryopreservation of Cord Blood Stem Cells. Transfus. Med. Rev. 2021, 35, 95–102. [Google Scholar] [CrossRef]
- Lombardo, G.; Lechanteur, C.; Briquet, A.; Seidel, L.; Willems, E.; Servais, S.; Baudoux, E.; Kerre, T.; Zachee, P.; Herman, J.; et al. Co-infusion of mesenchymal stromal cells to prevent GVHD after allogeneic hematopoietic cell transplantation from HLA-mismatched unrelated donors after reduced-intensity conditioning: A double-blind randomized study and literature review. Stem Cell Res. Ther. 2024, 15, 461. [Google Scholar] [CrossRef]
- Wang, M.; Lemos, B. Ribosomal DNA harbors an evolutionarily conserved clock of biological aging. Genome Res. 2019, 29, 325–333. [Google Scholar] [CrossRef]
- Abad, M.; Mosteiro, L.; Pantoja, C.; Cañamero, M.; Rayon, T.; Ors, I.; Graña, O.; Megías, D.; Domínguez, O.; Martínez, D.; et al. Reprogramming in vivo produces teratomas and iPS cells with totipotency features. Nature 2013, 502, 340–345. [Google Scholar] [CrossRef]
- Ocampo, A.; Reddy, P.; Martinez-Redondo, P.; Platero-Luengo, A.; Hatanaka, F.; Hishida, T.; Li, M.; Lam, D.; Kurita, M.; Beyret, E.; et al. In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming. Cell 2016, 167, 1719–1733.e12. [Google Scholar] [CrossRef]
- Sarkar, T.J.; Quarta, M.; Mukherjee, S.; Colville, A.; Paine, P.; Doan, L.; Tran, C.M.; Chu, C.R.; Horvath, S.; Qi, L.S.; et al. Transient non-integrative expression of nuclear reprogramming factors promotes multifaceted amelioration of aging in human cells. Nat. Commun. 2020, 11, 1545. [Google Scholar] [CrossRef] [PubMed]
- Olova, N.; Simpson, D.J.; Marioni, R.E.; Chandra, T. Partial reprogramming induces a steady decline in epigenetic age before loss of somatic identity. Aging Cell 2019, 18, e12877. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Wang, G.; Wang, J.; Zhang, Z.; Fu, Y.; Cheng, L.; Meng, G.; Lyu, Y.; Zhu, J.; Li, Y.; et al. Chemical reprogramming of human somatic cells to pluripotent stem cells. Nature 2022, 605, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Petty, C.A.; Dixon-McDougall, T.; Lopez, M.V.; Tyshkovskiy, A.; Maybury-Lewis, S.; Tian, X.; Ibrahim, N.; Chen, Z.; Griffin, P.T.; et al. Chemically induced reprogramming to reverse cellular aging. Aging 2023, 15, 5966–5989. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhong, X.; He, H.; Cao, J.; Zhou, Z.; Dong, J.; Li, H.; Zhang, A.; Lyu, Y.; Li, C.; et al. Generation of human expandable limb-bud-like progenitors via chemically induced dedifferentiation. Cell Stem Cell 2024, 31, 1732–1740.e6. [Google Scholar] [CrossRef]
- Roux, A.E.; Zhang, C.; Paw, J.; Zavala-Solorio, J.; Malahias, E.; Vijay, T.; Kolumam, G.; Kenyon, C.; Kimmel, J.C. Diverse partial reprogramming strategies restore youthful gene expression and transiently suppress cell identity. Cell Syst. 2022, 13, 574–587.e13. [Google Scholar] [CrossRef] [PubMed]
- Spitzhorn, L.S.; Megges, M.; Wruck, W.; Rahman, M.S.; Otte, J.; Degistirici, Ö.; Meisel, R.; Sorg, R.V.; Oreffo, R.O.C.; Adjaye, J. Human iPSC-derived MSCs (iMSCs) from aged individuals acquire a rejuvenation signature. Stem Cell Res. Ther. 2019, 10, 100. [Google Scholar] [CrossRef]
- Lai, P.L.; Lin, H.; Chen, S.F.; Yang, S.C.; Hung, K.H.; Chang, C.F.; Chang, H.Y.; Lu, F.L.; Lee, Y.H.; Liu, Y.C.; et al. Efficient Generation of Chemically Induced Mesenchymal Stem Cells from Human Dermal Fibroblasts. Sci. Rep. 2017, 7, 44534. [Google Scholar] [CrossRef]
- Kaise, T.; Fukui, M.; Sueda, R.; Piao, W.; Yamada, M.; Kobayashi, T.; Imayoshi, I.; Kageyama, R. Functional rejuvenation of aged neural stem cells by Plagl2 and anti-Dyrk1a activity. Genes Dev. 2022, 36, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Yi, W.W.; Liu, B.; Zhang, C.E.; Yang, C.C.; Zeng, L.; Li, L.; Luo, G.; Zhang, L.; Ju, Z.Y.; et al. Tetrahydroxy stilbene glucoside rejuvenates aging hematopoietic stem cells with predilection for lymphoid differentiation via AMPK and Tet2. J. Adv. Res. 2025, 70, 515–529. [Google Scholar] [CrossRef]
- Zeng, X.; Shi, C.; Han, Y.; Hu, K.; Li, X.; Wei, C.; Ding, L.; Cui, J.; Huang, S.; Xu, Y.; et al. A metabolic atlas of blood cells in young and aged mice identifies uridine as a metabolite to rejuvenate aged hematopoietic stem cells. Nat. Aging 2024, 4, 1477–1492. [Google Scholar] [CrossRef]
- Montserrat-Vazquez, S.; Ali, N.J.; Matteini, F.; Lozano, J.; Zhaowei, T.; Mejia-Ramirez, E.; Marka, G.; Vollmer, A.; Soller, K.; Sacma, M.; et al. Transplanting rejuvenated blood stem cells extends lifespan of aged immunocompromised mice. NPJ Regen. Med. 2022, 7, 78. [Google Scholar] [CrossRef]
- Lei, J.; Xin, Z.; Liu, N.; Ning, T.; Jing, Y.; Qiao, Y.; He, Z.; Jiang, M.; Yang, Y.; Zhang, Z.; et al. Senescence-resistant human mesenchymal progenitor cells counter aging in primates. Cell 2025. [Google Scholar] [CrossRef]
- Bi, Y.; Qiao, X.; Cai, Z.; Zhao, H.; Ye, R.; Liu, Q.; Gao, L.; Liu, Y.; Liang, B.; Liu, Y.; et al. Exosomal miR-302b rejuvenates aging mice by reversing the proliferative arrest of senescent cells. Cell Metab. 2025, 37, 527–541.e6. [Google Scholar] [CrossRef]
- Chantachotikul, P.; Liu, S.; Furukawa, K.; Deguchi, S. AP2A1 modulates cell states between senescence and rejuvenation. Cell. Signal. 2025, 127, 111616. [Google Scholar] [CrossRef]
- Kerepesi, C.; Gladyshev, V.N. Intersection clock reveals a rejuvenation event during human embryogenesis. Aging Cell 2023, 22, e13922. [Google Scholar] [CrossRef] [PubMed]
- Kerepesi, C.; Zhang, B.; Lee, S.G.; Trapp, A.; Gladyshev, V.N. Epigenetic clocks reveal a rejuvenation event during embryogenesis followed by aging. Sci. Adv. 2021, 7, eabg6082. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, R115. [Google Scholar] [CrossRef]
- Haghani, A.; Li, C.Z.; Robeck, T.R.; Zhang, J.; Lu, A.T.; Ablaeva, J.; Acosta-Rodríguez, V.A.; Adams, D.M.; Alagaili, A.N.; Almunia, J.; et al. DNA methylation networks underlying mammalian traits. Science 2023, 381, eabq5693. [Google Scholar] [CrossRef] [PubMed]
- Daios, S.; Anogeianaki, A.; Kaiafa, G.; Kontana, A.; Veneti, S.; Gogou, C.; Karlafti, E.; Pilalas, D.; Kanellos, I.; Savopoulos, C. Telomere Length as a Marker of Biological Aging: A Critical Review of Recent Literature. Curr. Med. Chem. 2022, 29, 5478–5495. [Google Scholar] [CrossRef]
- Demanelis, K.; Jasmine, F.; Chen, L.S.; Chernoff, M.; Tong, L.; Delgado, D.; Zhang, C.; Shinkle, J.; Sabarinathan, M.; Lin, H.; et al. Determinants of telomere length across human tissues. Science 2020, 369, eaaz6876. [Google Scholar] [CrossRef]
- Wikgren, M.; Karlsson, T.; Nilbrink, T.; Nordfjäll, K.; Hultdin, J.; Sleegers, K.; Van Broeckhoven, C.; Nyberg, L.; Roos, G.; Nilsson, L.G.; et al. APOE ε4 is associated with longer telomeres, and longer telomeres among ε4 carriers predicts worse episodic memory. Neurobiol. Aging 2012, 33, 335–344. [Google Scholar] [CrossRef]
- Liao, L.; Shi, B.; Chang, H.; Su, X.; Zhang, L.; Bi, C.; Shuai, Y.; Du, X.; Deng, Z.; Jin, Y. Heparin improves BMSC cell therapy: Anticoagulant treatment by heparin improves the safety and therapeutic effect of bone marrow-derived mesenchymal stem cell cytotherapy. Theranostics 2017, 7, 106–116. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Young/Rejuvenated Stem Cells | Experimental Animals | Effects of Transplantation | Reference |
|---|---|---|---|
| Young Mesenchymal Stem Cells | mouse | slow the loss of bone density, and prolong the lifespan of old mice. | [4] |
| Young Hematopoietic Stem Cells | mouse | reductions in frailty measures and increases in food intake and body weight of aged recipients, and extend the lifespan of old mice. | [5] |
| Rejuvenated Hematopoietic Stem Cells | mouse | Enhance lymphoid and regenerative capacity in serial transplantation assays, and extend the healthspan and lifespan of aged mice. | [79] |
| FOXO3 double-mutated senescence-resistant human mesenchymal progenitor cells | macaque | a systemic reduction in aging indicators, such as cellular senescence, chronic inflammation, and tissue degeneration, without any detected adverse effects. Enhanced brain architecture and cognitive function, and alleviated the reproductive system decline. | [80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, A.; Ma, C.; Hu, M. Rejuvenated Autologous Adult Stem Cells: Emerging Front Runners in the Fight Against Aging and Associated Diseases. Cells 2025, 14, 1153. https://doi.org/10.3390/cells14151153
Yu A, Ma C, Hu M. Rejuvenated Autologous Adult Stem Cells: Emerging Front Runners in the Fight Against Aging and Associated Diseases. Cells. 2025; 14(15):1153. https://doi.org/10.3390/cells14151153
Chicago/Turabian StyleYu, An, Changguo Ma, and Min Hu. 2025. "Rejuvenated Autologous Adult Stem Cells: Emerging Front Runners in the Fight Against Aging and Associated Diseases" Cells 14, no. 15: 1153. https://doi.org/10.3390/cells14151153
APA StyleYu, A., Ma, C., & Hu, M. (2025). Rejuvenated Autologous Adult Stem Cells: Emerging Front Runners in the Fight Against Aging and Associated Diseases. Cells, 14(15), 1153. https://doi.org/10.3390/cells14151153