
Molecular Insights into the Superiority of Platelet Lysate over FBS for hASC Expansion and Wound Healing

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of hASCs
2.2. Cell Proliferation Assay
2.3. RNA Sequencing
2.4. Quantitative Reverse Transcription‒Polymerase Chain Reaction (RT‒qPCR)
2.5. Stress Resistance Assay
2.6. Immunofluorescence Assay
2.7. Apoptosis Assay Using Annexin V
2.8. Senescence β-Galactosidase (SA-β-gal) Assay
2.9. Murine Wound Repair
2.10. Histological and Immunochemical Analysis of Wound Healing
2.11. Statistical Analysis
3. Results
3.1. Proliferative and Stress Resistance Effects of PL on hASCs Cultured with PL
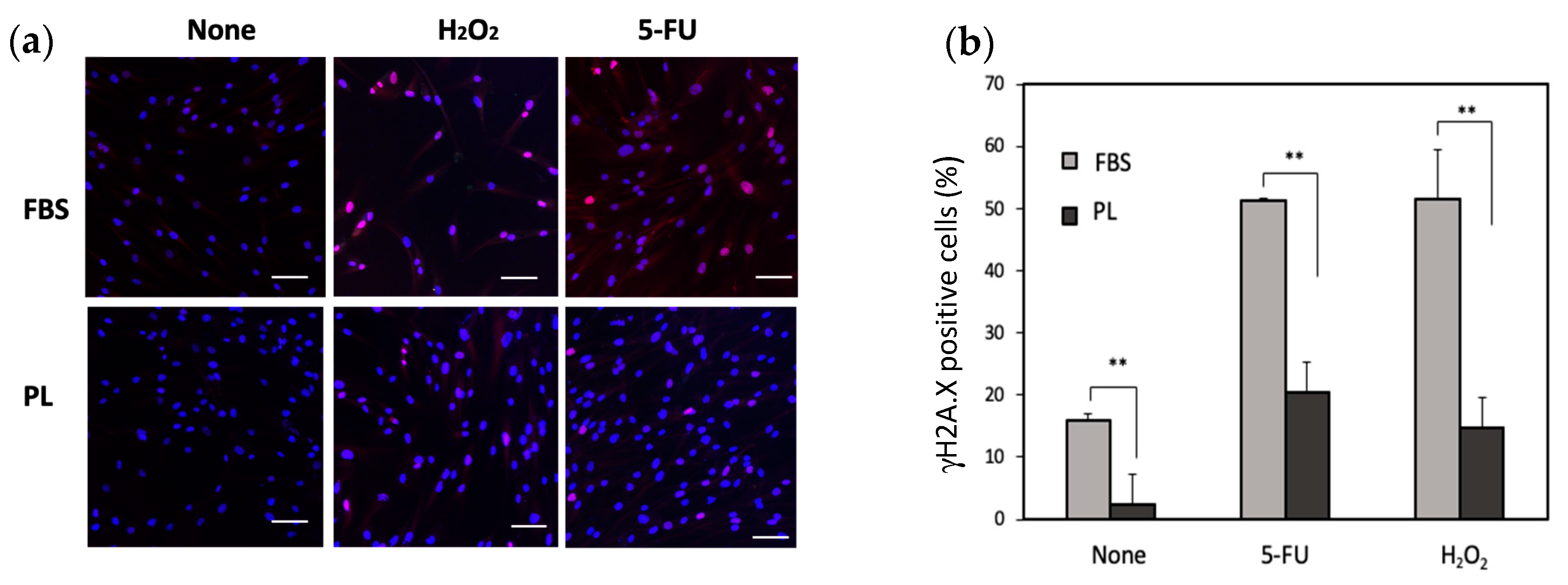
3.2. Effect of PL on the Survival of hASCs Treated with Stress-Inducing Agents
3.3. Effect of PL on the Apoptosis of hASCs Treated with Stress-Inducing Agents
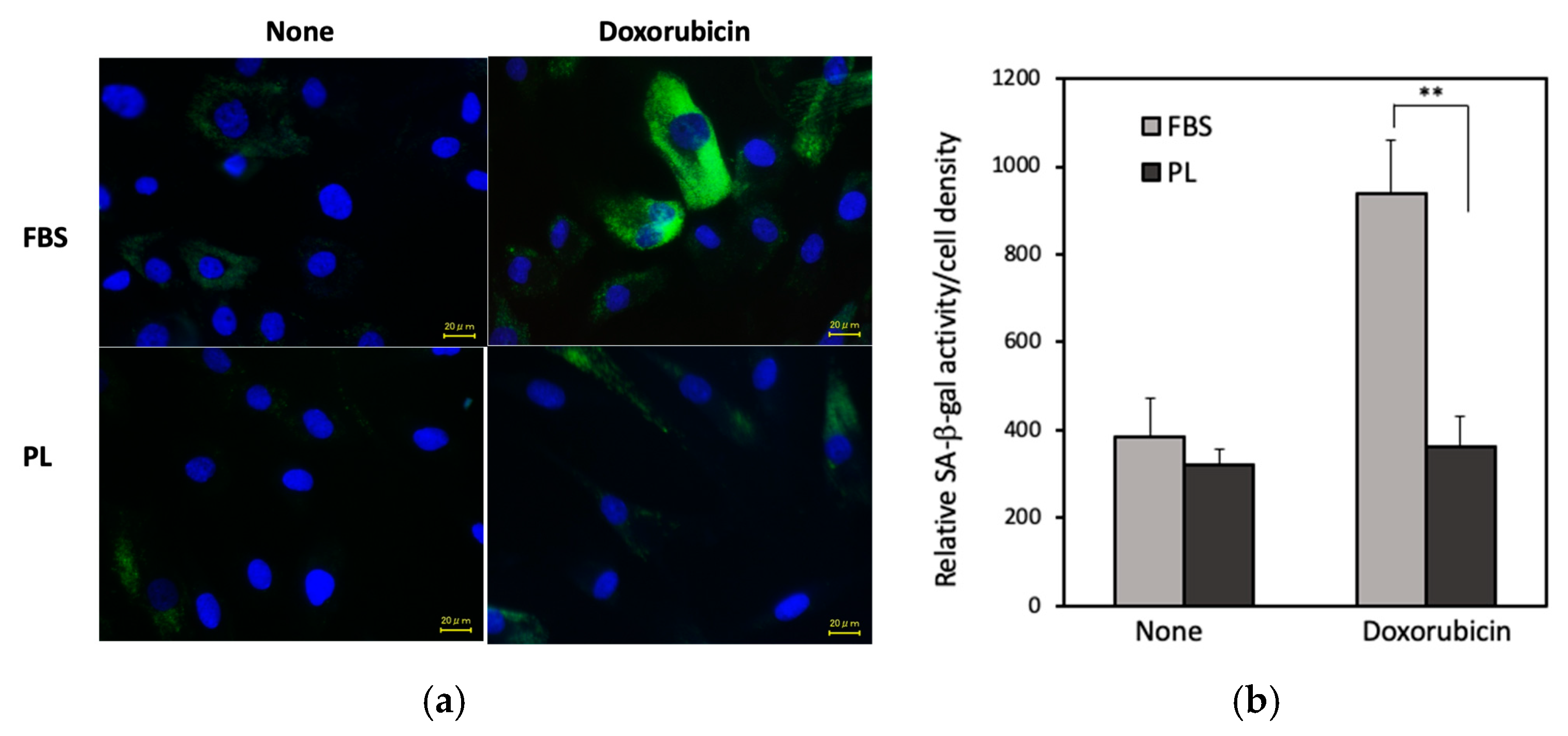
3.4. Effect of PL on Doxorubicin-Induced hASC Aging
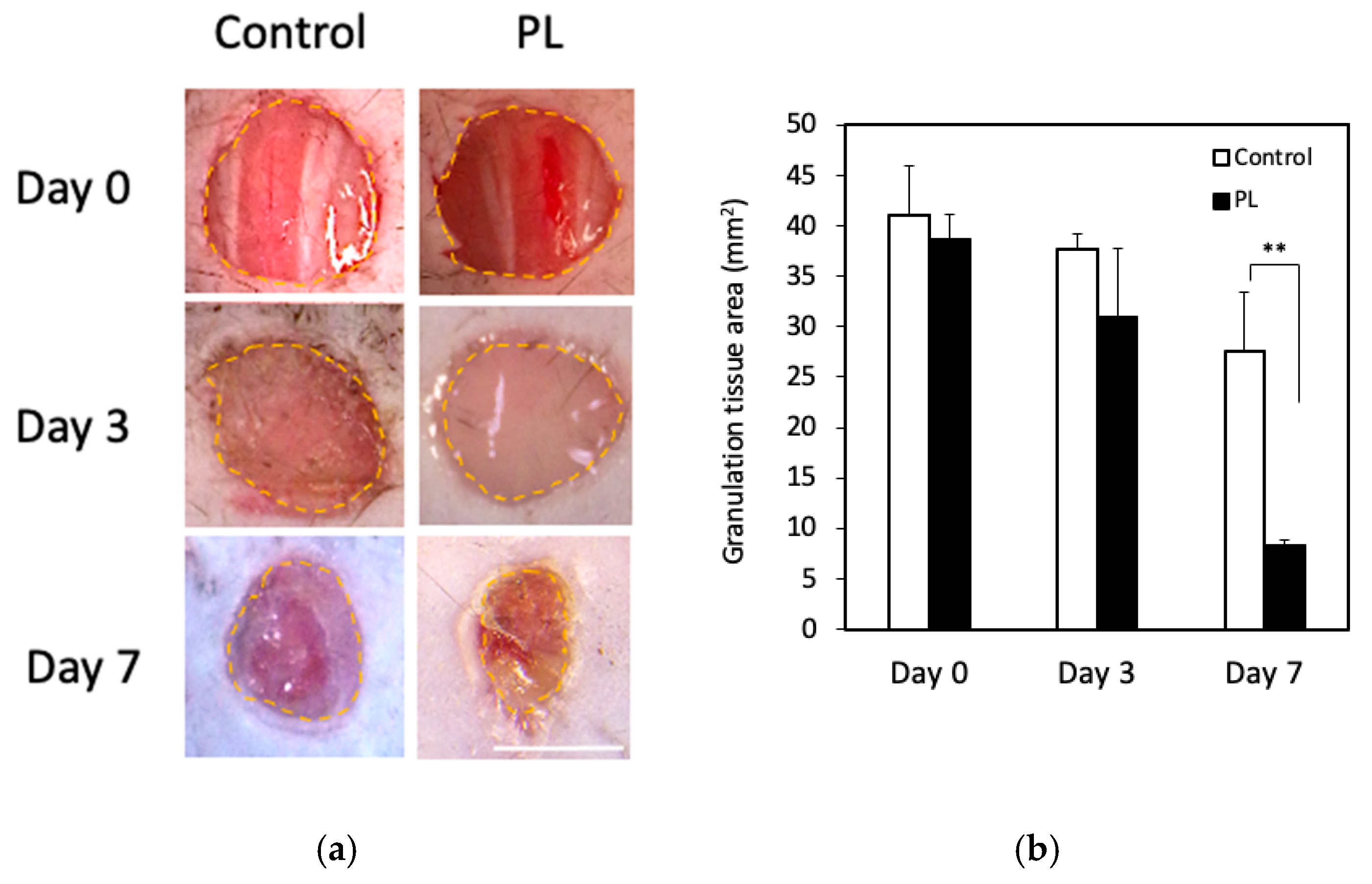
3.5. Effect of PL on Wound Healing In Vivo
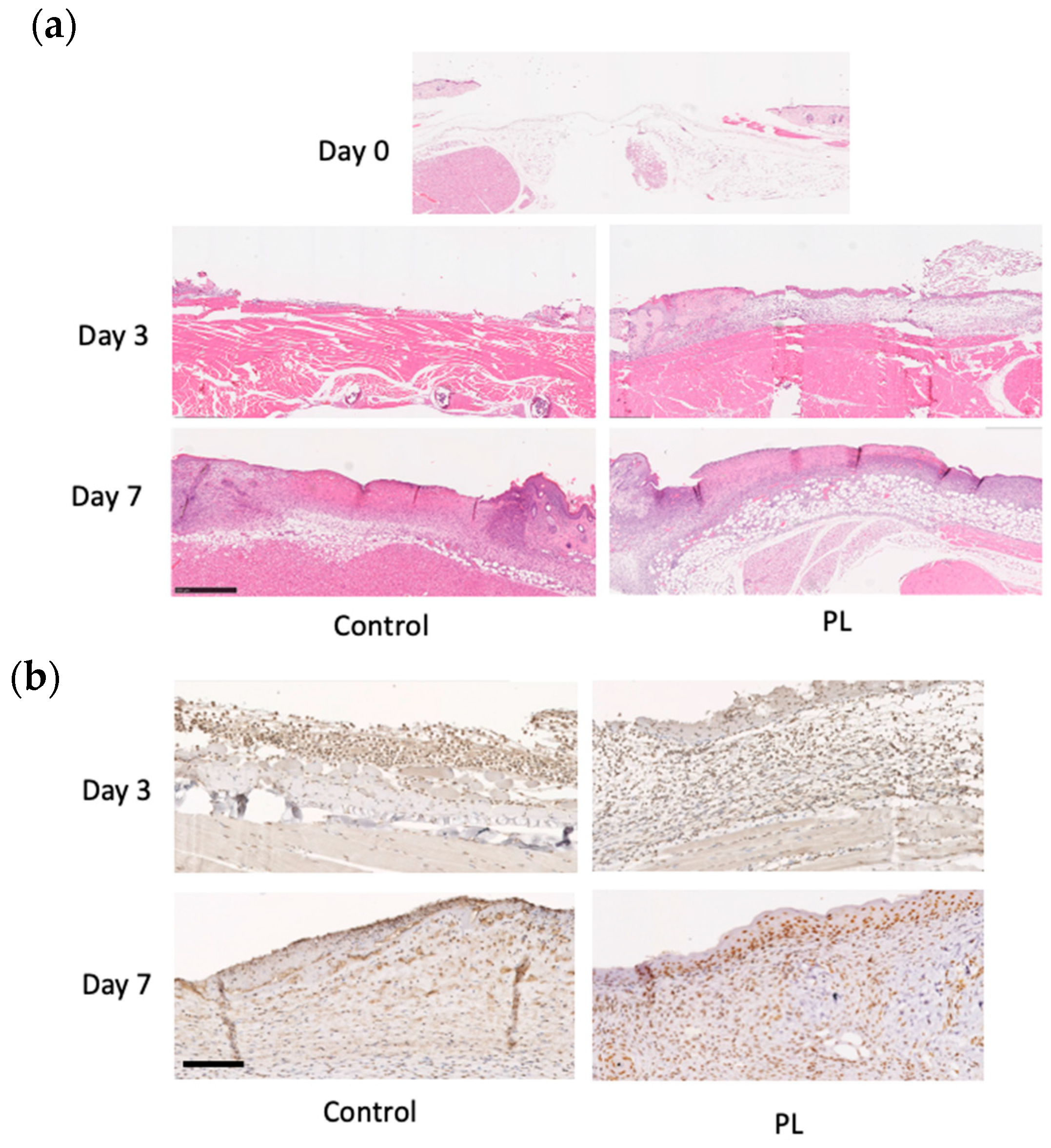
3.6. Histological and Immunohistochemical Analysis of the Wound Healing Effect of PL
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| hASCs | human adipose-derived stem cells |
| MSCs | mesenchymal stem cells |
| VEGF | vascular endothelial growth factor |
| HGF | hepatocyte growth factor |
| FGF | fibroblast growth factor |
| IGF-1 | insulin-like growth factor-1 |
| BDNF | brain-derived neurotrophic factor |
| IL-1Ra | interleukin-1 receptor antagonist |
| IL-6 | interleukin-6 |
| IL-8 | interleukin-8 |
| FBS | fetal bovine serum |
| PL | human platelet lysate; in all experimental sections of this study, PL refers to the commercial pooled human platelet lysate PLTMax® (Merck Millipore, Germany) |
| PRP | platelet-rich plasma |
| PDGF | platelet-derived growth factor |
| EGF | epidermal growth factor |
| GMPs | Good Manufacturing Practices |
| HUVECs | human umbilical vein endothelial cells |
| RNA-seq | RNA sequencing |
| PCA | principal-component analysis |
| ORA | over-representation analysis |
| DMEM | Dulbecco’s modified Eagle’s medium |
| CCK-8 | Cell Counting Kit-8 |
| chk1 | checkpoint kinase 1 |
| RT‒qPCR | reverse transcription‒quantitative polymerase chain reaction |
| 5-FU | 5-fluorouracil |
| SA-β-gal | senescence-associated β-galactosidase |
| CGSs | Collagen/Gelatin Sponges |
| HE | hematoxylin and eosin |
| SD | standard deviation |
| TGF-β | transforming growth factor-β |
References
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Miana, V.V.; Prieto González, E.A. Adipose tissue stem cells in regenerative medicine. Ecancer Med. Sci. 2018, 12, 822. [Google Scholar] [CrossRef] [PubMed]
- Koźlik, M.; Wójcicki, P. The use of stem cells in plastic and reconstructive surgery. Adv. Clin. Exp. Med. 2014, 23, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghadban, S.; Bunnell, B.A. Adipose tissue-derived stem cells: Immunomodulatory effects and therapeutic potential. Physiology 2020, 35, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Mazini, L.; Rochette, L.; Admou, B.; Amal, S.; Malka, G. Hopes and limits of adipose-derived stem cells (ADSCs) and mesenchymal stem cells (MSCs) in wound healing. Int. J. Mol. Sci. 2020, 21, 1306. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.; Brito, S.; Kwak, B.M.; Park, S.; Lee, M.G.; Bin, B.H. Applications of mesenchymal stem cells in skin regeneration and rejuvenation. Int. J. Mol. Sci. 2021, 22, 2410. [Google Scholar] [CrossRef] [PubMed]
- Schneider, I.; Calcagni, M.; Buschmann, J. Adipose-derived stem cells applied in skin diseases, wound healing and skin defects: A review. Cytotherapy 2023, 25, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Epstein, G.K.; Epstein, J.S. Mesenchymal stem cells and stromal vascular fraction for hair loss: Current status. Facial Plast. Surg. Clin. N. Am. 2018, 26, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, K.; Sato, K.; Aoi, N.; Kurita, M.; Hirohi, T.; Harii, K. Cell-assisted lipotransfer for cosmetic breast augmentation: Supportive use of adipose-derived stem/stromal cells. Aesthet. Plast. Surg. 2008, 32, 48–55. [Google Scholar] [CrossRef] [PubMed]
- da Fonseca, L.; Santos, G.S.; Huber, S.C.; Setti, T.M.; Setti, T.; Lana, J.F. Human platelet lysate—A potent (and overlooked) orthobiologic. J. Clin. Orthop. Trauma 2021, 21, 101534. [Google Scholar] [CrossRef] [PubMed]
- Burnouf, T.; Strunk, D.; Koh, M.B.C.; Schallmoser, K. Human platelet lysate: Replacing fetal bovine serum as a gold standard for human cell propagation? Biomaterials 2016, 76, 371–387. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.J.; Sun, Y.C.; Christopher, K.; Pai, A.S.I.; Lu, C.J.; Hu, F.R.; Lin, S.-Y.; Chen, W.-L.; Ljubimov, A.V. Comparison of corneal epitheliotrophic capacities among human platelet lysates and other blood derivatives. PLoS ONE 2017, 12, e0171008. [Google Scholar] [CrossRef] [PubMed]
- Valtetsiotis, K.; Di Martino, A.; Brunello, M.; D’Agostino, C.; Poluzzi, R.; Ferri, R.; Mora, P.; Traina, F.; Faldini, C. Platelet lysate for the treatment of osteoarthritis: A systematic review of preclinical and clinical studies. Musculoskelet. Surg. 2024, 108, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Altaie, A.; Owston, H.; Jones, E. Use of platelet lysate for bone regeneration—Are we ready for clinical translation? World J. Stem Cells 2016, 8, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Griffith, G.L.; Holt, A.W.; Eriksson, E.; Johnson, A.J.; McDaniel, J.S. Human platelet lysate delivered via an ocular wound chamber for the treatment of corneal epithelial injuries. Exp. Eye Res. 2021, 206, 108493. [Google Scholar] [CrossRef] [PubMed]
- Kakudo, N.; Minakata, T.; Mitsui, T.; Kushida, S.; Notodihardjo, F.Z.; Kusumoto, K. Proliferation-promoting effect of platelet-rich plasma on human adipose–derived stem cells and human dermal fibroblasts. Plast. Reconstr. Surg. 2008, 122, 1352. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Ku, N.J.; Sung, T.C.; Higuchi, A.; Hung, C.S.; Lee, H.H.C.; Ling, Q.-D.; Cheng, N.-C.; Umezawa, A.; Barro, L.; et al. The effect of human platelet lysate on the differentiation ability of human adipose-derived stem cells cultured on ECM-coated surfaces. J. Mater. Chem. B 2019, 7, 7110–7119. [Google Scholar] [CrossRef] [PubMed]
- Peters, K.; Helmert, T.; Gebhard, S.; Mailänder, V.; Unger, R.E.; Nezi-Cahn, S.; Hasenburg, A.; Heller, M.; Schwab, R.; Brenner, W. Standardized human platelet lysates as adequate substitute to fetal calf serum in endothelial cell culture for tissue engineering. Biomed. Res. Int. 2022, 2022, 3807314. [Google Scholar] [CrossRef] [PubMed]
- Wakamoto, S.; Furukawa, T.; Kawabori, M.; Akino, M.; Kato, S.; Fuse, H.; Ohtsuki, S.; Torimoto, Y.; Fujimura, M.; Kino, S. Human platelet lysate produced from leukoreduction filter contents enables sufficient MSC growth. Stem Cell Res. Ther. 2025, 16, 205. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.; Kakudo, N.; Morimoto, N.; Taketani, S.; Hara, T.; Ogawa, T.; Kusumoto, K. Platelet-rich plasma enhances the proliferation of human adipose stem cells through multiple signaling pathways. Stem Cell Res. Ther. 2018, 9, 107. [Google Scholar] [CrossRef] [PubMed]
- Fukui, M.; Lai, F.; Hihara, M.; Mitsui, T.; Matsuoka, Y.; Sun, Z.; Kunieda, S.; Taketani, S.; Odaka, T.; Okuma, K.; et al. Activation of cell adhesion and migration is an early event of platelet-rich plasma (PRP)-dependent stimulation of human adipose-derived stem/stromal cells. Hum. Cell 2024, 37, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Fukui, M.; Taketani, S.; Kako, A.; Kunieda, S.; Kakudo, N. Predominant control of PDGF/PDGF receptor signaling in the migration and proliferation of human adipose derived stem cells under culture conditions with a combination of growth factors. Exp. Ther. Med. 2024, 27, 156. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Zhang, C.; Xu, L.; Chen, H.; Fan, X.; Sun, B.; Tang, Q.; Zhan, Y.; Chen, T.; Wang, X. BAK plays a key role in A-1331852-induced apoptosis in senescent chondrocytes. Biochem. Biophys. Res. Commun. 2022, 609, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Notodihardjo, S.C.; Morimoto, N.; Kakudo, N.; Mitsui, T.; Le, T.M.; Tabata, Y.; Kusumoto, K. Efficacy of gelatin hydrogel impregnated with concentrated platelet lysate in murine wound healing. J. Surg. Res. 2019, 234, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Kakudo, N.; Morimoto, N.; Ma, Y.; Kusumoto, K. Differences between the proliferative effects of human platelet lysate and fetal bovine serum on human adipose-derived stem cells. Cells 2019, 8, 1218. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.T.; Nardini, M.; Ruggiu, A.; Cancedda, R.; Descalzi, F.; Mastrogiacomo, M. Platelet lysate induces in human osteoblasts resumption of cell proliferation and activation of pathways relevant for revascularization and regeneration of damaged bone. Int. J. Mol. Sci. 2020, 21, 5123. [Google Scholar] [CrossRef] [PubMed]
- Losi, P.; Al Kayal, T.; Buscemi, M.; Foffa, I.; Cavallo, A.; Soldani, G. Bilayered fibrin-based electrospun-sprayed scaffold loaded with platelet lysate enhances wound healing in a diabetic mouse model. Nanomaterials 2020, 10, 2128. [Google Scholar] [CrossRef] [PubMed]
- Widyaningrum, R.; Burnouf, T.; Nebie, O.; Delila, L.; Wang, T.J. A purified human platelet pellet lysate rich in neurotrophic factors and antioxidants repairs and protects corneal endothelial cells from oxidative stress. Biomed. Pharmacother. 2021, 142, 112046. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.S.; Luo, H.L.; Chuang, Y.C.; Lee, W.C.; Wang, H.J.; Chancellor, M.B. Platelet lysate therapy attenuates hypoxia-induced apoptosis in human uroepithelial SV-HUC-1 cells through regulating the oxidative stress and mitochondrial-mediated intrinsic apoptotic pathway. Biomedicines 2023, 11, 935. [Google Scholar] [CrossRef] [PubMed]
- Wirohadidjojo, Y.W.; Budiyanto, A.; Soebono, H. Platelet-rich fibrin lysate can ameliorate dysfunction of chronically UVA-irradiated human dermal fibroblasts. Yonsei Med. J. 2016, 57, 1282–1285. [Google Scholar] [CrossRef] [PubMed]
- Hudgens, J.L.; Sugg, K.B.; Grekin, J.A.; Gumucio, J.P.; Bedi, A.; Mendias, C.L. Platelet-rich plasma activates proinflammatory signaling pathways and induces oxidative stress in tendon fibroblasts. Am. J. Sports Med. 2016, 44, 1931–1940. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liu, C.; Zhang, Y.; Bi, L. Effect of super activated platelet lysate on cell proliferation, repair and osteogenesis. Biomed. Mater. Eng. 2023, 34, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Coryell, P.R.; Diekman, B.O.; Loeser, R.F. Mechanisms and therapeutic implications of cellular senescence in osteoarthritis. Nat. Rev. Rheumatol. 2021, 17, 47–57. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Sharpless, N.E. Senescence in health and disease. Cell 2017, 169, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wei, D.; Xiao, H. Methods of cellular senescence induction using oxidative stress. Methods Mol. Biol. 2013, 1048, 135–144. [Google Scholar] [PubMed]
- Huang, W.; Hickson, L.J.; Eirin, A.; Kirkland, J.L.; Lerman, L.O. Cellular senescence: The good, the bad and the unknown. Nat. Rev. Nephrol. 2022, 18, 611–627. [Google Scholar] [CrossRef] [PubMed]
- Chitchongyingcharoen, N.; Tawonsawatruk, T.; Phetfong, J.; Aroontanee, W.; Supokawej, A. Application of human platelet lysate in chondrocyte expansion promotes chondrogenic phenotype and slows senescence progression via BMP–TAK1–p38 pathway. Sci. Rep. 2023, 13, 21106. [Google Scholar] [CrossRef] [PubMed]
- Weng, H.P.; Cheng, Y.Y.; Lee, H.L.; Hsu, T.Y.; Chang, Y.T.; Shen, Y.A. Enhanced platelet-rich plasma (ePRP) stimulates wound healing through effects on metabolic reprogramming in fibroblasts. Int. J. Mol. Sci. 2021, 22, 12623. [Google Scholar] [CrossRef] [PubMed]
- Paterson, K.L.; Hunter, D.J.; Metcalf, B.R.; Eyles, J.; Duong, V.; Kazsa, J.; Wang, Y.; Buchbinder, R.; Cicuttini, F.; Forbes, A.; et al. Efficacy of intra-articular injections of platelet-rich plasma as a symptom- and disease-modifying treatment for knee osteoarthritis—The RESTORE trial protocol. BMC Musculoskelet. Disord. 2018, 19, 272. [Google Scholar] [CrossRef] [PubMed]
- Alhawari, H.; Jafar, H.; Al Soudi, M.; Ameereh, L.A.; Fawaris, M.; Saleh, M.; Aladwan, S.; Younes, N.; Awidi, A. Perilesional injections of human platelet lysate versus platelet poor plasma for the treatment of diabetic foot ulcers: A double-blinded prospective clinical trial. Int. Wound J. 2023, 20, 3116–3122. [Google Scholar] [CrossRef] [PubMed]
- Pakhomova, E.E.; Smirnova, I.O. Comparative evaluation of the clinical efficacy of PRP-therapy, minoxidil, and their combination with immunohistochemical study of the dynamics of cell proliferation in the treatment of men with androgenetic alopecia. Int. J. Mol. Sci. 2020, 21, 6516. [Google Scholar] [CrossRef] [PubMed]
- Scopelliti, F.; Cattani, C.; Dimartino, V.; Mirisola, C.; Cavani, A. Platelet derivatives and the immunomodulation of wound healing. Int. J. Mol. Sci. 2022, 23, 8370. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Kumar, S.; Garg, P.; Verma, Y.K. Platelet-rich plasma: A comparative and economical therapy for wound healing and tissue regeneration. Cell Tissue Bank. 2023, 24, 285–306. [Google Scholar] [CrossRef] [PubMed]
- Smits, V.A.J.; Gillespie, D.A. DNA damage control: Regulation and functions of checkpoint kinase 1. FEBS J. 2015, 282, 3681–3692. [Google Scholar] [CrossRef] [PubMed]
- Peddibhotla, S.; Rosen, J.M. Checking and executing cell division to prevent genomic instability. Cell Cycle 2009, 8, 2339–2342. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunieda, S.; Fukui, M.; Kuro, A.; Mitsui, T.; Li, H.; Sun, Z.; Ueda, T.; Taketani, S.; Higasa, K.; Kakudo, N. Molecular Insights into the Superiority of Platelet Lysate over FBS for hASC Expansion and Wound Healing. Cells 2025, 14, 1154. https://doi.org/10.3390/cells14151154
Kunieda S, Fukui M, Kuro A, Mitsui T, Li H, Sun Z, Ueda T, Taketani S, Higasa K, Kakudo N. Molecular Insights into the Superiority of Platelet Lysate over FBS for hASC Expansion and Wound Healing. Cells. 2025; 14(15):1154. https://doi.org/10.3390/cells14151154
Chicago/Turabian StyleKunieda, Sakurako, Michika Fukui, Atsuyuki Kuro, Toshihito Mitsui, Huan Li, Zhongxin Sun, Takayuki Ueda, Shigeru Taketani, Koichiro Higasa, and Natsuko Kakudo. 2025. "Molecular Insights into the Superiority of Platelet Lysate over FBS for hASC Expansion and Wound Healing" Cells 14, no. 15: 1154. https://doi.org/10.3390/cells14151154
APA StyleKunieda, S., Fukui, M., Kuro, A., Mitsui, T., Li, H., Sun, Z., Ueda, T., Taketani, S., Higasa, K., & Kakudo, N. (2025). Molecular Insights into the Superiority of Platelet Lysate over FBS for hASC Expansion and Wound Healing. Cells, 14(15), 1154. https://doi.org/10.3390/cells14151154