
The Influence of Circadian Rhythms on Transcranial Direct Current Stimulation (tDCS) Effects: Theoretical and Practical Considerations


Abstract
1. Introduction
1.1. Borbély’s Two-Process Model of Sleep Regulation
1.2. Synaptic Homeostasis Hypothesis
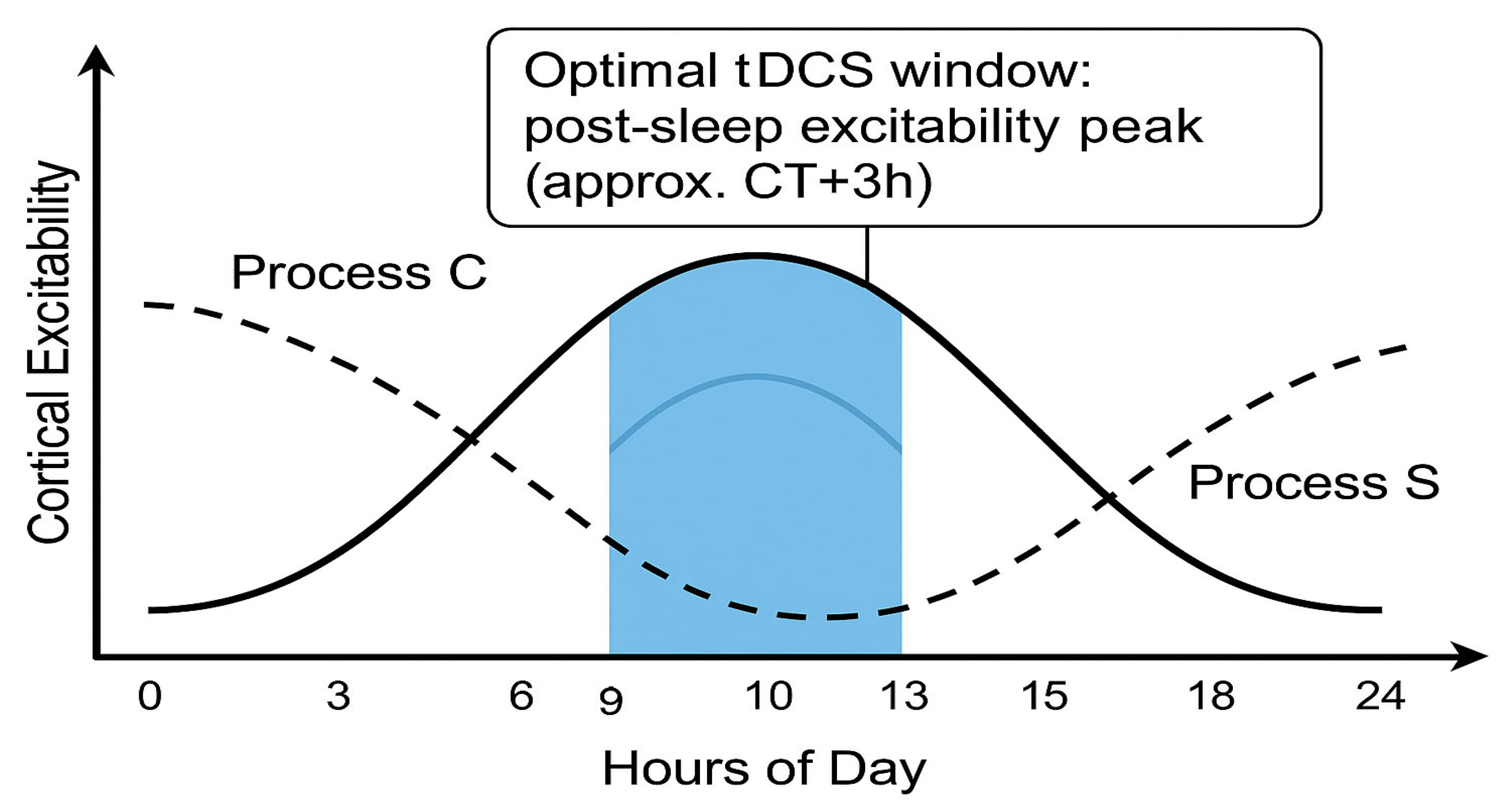
- On the circadian axis (Process C), the window opens during the late biological morning and early evening, when intrinsic alertness and membrane depolarisation peak.
- On the homeostatic axis (Process S), the window is widest after consolidated sleep, when synaptic weights have been renormalised.
- Outside this overlap—i.e., under high sleep pressure or during the circadian trough—baseline excitability is either saturated or too low, glucocorticoid tone is unfavourable, and tDCS effects are diminished or even reversed.
1.3. Search Strategy, Inclusion Criteria, and Synthesis Approach
2. Circadian Rhythms and Brain Function, Metabolism, Body Temperature, Cortisol, and Neurotransmitter Systems
3. Circadian Modulation of Cognitive Functions
4. The Influence of the Circadian Rhythm on Cortical Excitability and Synaptic Plasticity in Animal and Human Studies
4.1. Animal Studies
4.2. Human Studies
4.3. Meta-Analytic Evidence for Sleep-Pressure-Dependent Shifts in Cortical Excitability
4.4. Borbély’s Two-Process Model and Cortical Excitability
4.5. Where Transcranial Direct Current Stimulation Sits in the Two-Process Theory?
5. The Influence of Neurotransmitter Levels
Translating Neurotransmitter Rhythms into tDCS Protocol Design
6. Glucocorticoids and the HPA Axis: A Circadian Gate on Plasticity
7. Circadian Rhythms, Body Temperature, and Possible tDCS Outcomes
8. Cortisol and Temperature as Modulators in a tDCS “Window of Efficacy”
9. Individual Variability and Personalised Approaches
10. The Impact of tDCS on the Circadian Rhythm—A Bidirectional Relationship?
11. Practical Implications and Future Directions
Operational Details of the Chronobiological tDCS Framework
- Actigraphy (≥7 consecutive days; wrist-worn; 30 s epoch resolution) to extract sleep onset/offset, midsleep, and social jet-lag.
- Sleep diary (Karolinska or Consensus diary) to corroborate actigraphy.
- Morningness–Eveningness Questionnaire (MEQ) to classify chronotype but not to dictate stimulation timing.
- Optional EEG (home dry-electrode headband): Nightly frontal θ/α ratio and daytime resting-state α peak to index circadian sleep pressure and cortical excitability.
- Compute the individual’s average mid-sleep on free days corrected for sleep debt (MSF) as a phase marker.
- Define CT 0 (circadian time zero) as MSF—3 h; morning cortical-excitability peaks typically occur at CT +2 to +4 h.
- Default rule: Schedule tDCS at CT +3 h (≈2–3 h after habitual wake time).
- If objective EEG shows a delayed α-peak (>CT +4 h), shift session earlier by 30 min blocks until α-peak minus 30 min is reached.
- Montage: Target-specific (e.g., left DLPFC anode/right supra-orbital cathode for cognitive tasks).
- Intensity/duration: 1.5–2 mA × 20 min (ramp 30 s).
- Contra-indicator: If the participant slept < 6 h the previous night, postpone by 24 h to preserve the “well-rested” criterion.
- Neurophysiology: TMS-MEP I/O curve slope; resting-state EEG β-band power.
- Cognition: Three-back accuracy (working memory), Psychomotor Vigilance Task median RT.
- Sleep and circadian: Actigraphy-derived sleep efficiency, phase-angle difference (habitual wake time—tDCS start).
- Subjective: Karolinska Sleepiness Scale, tDCS side-effect questionnaire.
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kronfeld-Schor, N.; Bloch, G.; Schwartz, W.J. Animal clocks: When science meets nature. Proc. Biol. Sci. 2013, 280, 20131354. [Google Scholar] [CrossRef]
- Honma, S. The mammalian circadian system: A hierarchical multi-oscillator structure for generating circadian rhythm. J. Physiol. Sci. 2018, 68, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Merrow, M. The Circadian Clock and Human Health. Curr. Biol. 2016, 26, R432–R443. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef] [PubMed]
- Morf, J.; Schibler, U. Body temperature cycles: Gatekeepers of circadian clocks. Cell Cycle 2013, 12, 539–540. [Google Scholar] [CrossRef]
- Marcheva, B.; Ramsey, K.M.; Peek, C.B.; Affinati, A.; Maury, E.; Bass, J. Circadian clocks and metabolism. Handb. Exp. Pharmacol. 2013, 217, 127–155. [Google Scholar]
- Schmidt, C.; Collette, F.; Cajochen, C.; Peigneux, P. A time to think: Circadian rhythms in human cognition. Cogn. Neuropsychol. 2007, 24, 755–789. [Google Scholar] [CrossRef]
- Fishbein, A.B.; Knutson, K.L.; Zee, P.C. Circadian disruption and human health. J. Clin. Investig. 2021, 131, e148286. [Google Scholar] [CrossRef]
- Pinato, L.; Galina Spilla, C.S.; Markus, R.P.; da Silveira Cruz-Machado, S. Dysregulation of Circadian Rhythms in Autism Spectrum Disorders. Curr. Pharm. Des. 2019, 25, 4379–4393. [Google Scholar] [CrossRef]
- Alloy, L.B.; Ng, T.H.; Titone, M.K.; Boland, E.M. Circadian Rhythm Dysregulation in Bipolar Spectrum Disorders. Curr. Psychiatry Rep. 2017, 19, 21. [Google Scholar] [CrossRef]
- Canever, J.B.; Queiroz, L.Y.; Soares, E.S.; de Avelar, N.C.P.; Cimarosti, H.I. Circadian rhythm alterations affecting the pathology of neurodegenerative diseases. J. Neurochem. 2024, 168, 1475–1489. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Zhou, H.; Yan, R.; Yao, Z.; Lu, Q. Chronotype, circadian rhythm, and psychiatric disorders: Recent evidence and potential mechanisms. Front. Neurosci. 2022, 16, 811771. [Google Scholar] [CrossRef] [PubMed]
- Au, J.; Reece, J. The relationship between chronotype and depressive symptoms: A meta-analysis. J. Affect. Disord. 2017, 218, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Godos, J.; Currenti, W.; Ferri, R.; Lanza, G.; Caraci, F.; Frias-Toral, E.; Guglielmetti, M.; Ferraris, C.; Lipari, V.; Carvajal Altamiranda, S.; et al. Chronotype and Cancer: Emerging Relation Between Chrononutrition and Oncology from Human Studies. Nutrients 2025, 17, 529. [Google Scholar] [CrossRef]
- Heikkala, E.; Oura, P.; Korpela, T.; Karppinen, J.; Paananen, M. Chronotypes and disabling musculoskeletal pain: A Finnish birth cohort study. Eur. J. Pain. 2022, 26, 1069–1078. [Google Scholar] [CrossRef]
- James, S.M.; Honn, K.A.; Gaddameedhi, S.; Van Dongen, H.P.A. Shift Work: Disrupted Circadian Rhythms and Sleep-Implications for Health and Well-Being. Curr. Sleep Med. Rep. 2017, 3, 104–112. [Google Scholar] [CrossRef]
- Ashok Kumar, P.V.; Dakup, P.P.; Sarkar, S.; Modasia, J.B.; Motzner, M.S.; Gaddameedhi, S. It’s About Time: Advances in Understanding the Circadian Regulation of DNA Damage and Repair in Carcinogenesis and Cancer Treatment Outcomes. Yale J. Biol. Med. 2019, 92, 305–316. [Google Scholar]
- Bumgarner, J.R.; Walker, W.H., 2nd; Nelson, R.J. Circadian rhythms and pain. Neurosci. Biobehav. Rev. 2021, 129, 296–306. [Google Scholar] [CrossRef]
- Borbély, A.A. A two-process model of sleep regulation. Hum. Neurobiol. 1982, 1, 195–204. [Google Scholar]
- Daan, S.; Beersma, D.G.M.; Borbély, A.A. Timing of human sleep: Recovery process gated by a circadian pacemaker. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1984, 246, R161–R183. [Google Scholar] [CrossRef]
- Borbély, A.A.; Neuhaus, H.U. Sleep deprivation: Effects on sleep and EEG in the rat. J. Comp. Physiol. 1979, 133, 71–87. [Google Scholar] [CrossRef]
- Edgar, D.M.; Dement, W.C.; Fuller, C.A. Effect of SCN lesions on sleep in squirrel monkeys: Evidence for opponent processes in sleep–wake regulation. J. Neurosci. 1993, 13, 1065–1079. [Google Scholar] [CrossRef] [PubMed]
- Achermann, P.; Dijk, D.J.; Brunner, D.P.; Borbély, A.A. A model of human sleep homeostasis based on EEG slow-wave activity: Quantitative comparison of data and simulations. Brain Res. Bull. 1993, 31, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, J.K.; Ritz-De Cecco, A.; Czeisler, C.A.; Dijk, D.J. Circadian temperature and melatonin rhythms, sleep, and neurobehavioral function in humans living on a 20-h day. Am. J. Physiol. 1999, 277 Pt 2, R1152–R1163. [Google Scholar] [CrossRef] [PubMed]
- Dijk, D.J.; Czeisler, C.A. Contribution of the circadian pacemaker and the sleep homeostat to sleep propensity, sleep structure, electroencephalographic slow waves, and sleep spindle activity in humans. J. Neurosci. 1995, 15 Pt 1, 3526–3538. [Google Scholar] [CrossRef]
- Van Dongen, H.P.; Dinges, D.F. Investigating the interaction between the homeostatic and circadian processes of sleep-wake regulation for the prediction of waking neurobehavioural performance. J. Sleep Res. 2003, 12, 181–187. [Google Scholar] [CrossRef]
- Landolt, H.P. Sleep homeostasis: A role for adenosine in humans? Biochem. Pharmacol. 2008, 75, 2070–2079. [Google Scholar] [CrossRef]
- Lewy, A.J.; Ahmed, S.; Jackson, J.M.; Sack, R.L. Melatonin shifts human circadian rhythms according to a phase-response curve. Chronobiol. Int. 1992, 9, 380–392. [Google Scholar] [CrossRef]
- Refinetti, R. Circadian Physiology, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Trachsel, L.; Edgar, D.M.; Seidel, W.F.; Heller, H.C.; Dement, W.C. Sleep homeostasis in suprachiasmatic nuclei-lesioned rats: Effects of sleep deprivation and triazolam administration. Brain Res. 1992, 589, 253–261. [Google Scholar] [CrossRef]
- Tononi, G.; Cirelli, C. Sleep function and synaptic homeostasis. Sleep Med. Rev. 2006, 10, 49–62. [Google Scholar] [CrossRef]
- Kuhn, M.; Wolf, E.; Maier, J.G.; Mainberger, F.; Feige, B.; Schmid, H.; Bürklin, J.; Maywald, S.; Mall, V.; Jung, N.H.; et al. Sleep recalibrates homeostatic and associative synaptic plasticity in the human cortex. Nat. Commun. 2016, 7, 12455. [Google Scholar] [CrossRef]
- Tononi, G.; Cirelli, C. Sleep and the price of plasticity: From synaptic and cellular homeostasis to memory consolidation and integration. Neuron 2014, 81, 12–34. [Google Scholar] [CrossRef] [PubMed]
- Vyazovskiy, V.V.; Cirelli, C.; Pfister-Genskow, M.; Faraguna, U.; Tononi, G. Molecular and electrophysiological evidence for net synaptic potentiation in wake and depression in sleep. Nat. Neurosci. 2008, 11, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Maret, S.; Faraguna, U.; Nelson, A.B.; Cirelli, C.; Tononi, G. Sleep and waking modulate spine turnover in the adolescent mouse cortex. Nat. Neurosci. 2011, 14, 1418–1420. [Google Scholar] [CrossRef]
- Bushey, D.; Tononi, G.; Cirelli, C. Sleep and synaptic homeostasis: Structural evidence in Drosophila. Science 2011, 332, 1576–1581. [Google Scholar] [CrossRef]
- Donlea, J.M.; Ramanan, N.; Shaw, P.J. Use-dependent plasticity in clock neurons regulates sleep need in Drosophila. Science 2009, 324, 105–108. [Google Scholar] [CrossRef]
- Suppermpool, A.; Lyons, D.G.; Broom, E.; Rihel, J. Sleep pressure modulates single-neuron synapse number in zebrafish. Nature 2024, 629, 639–645. [Google Scholar] [CrossRef]
- Huber, R.; Mäki, H.; Rosanova, M.; Casarotto, S.; Canali, P.; Casali, A.G.; Tononi, G.; Massimini, M. Human cortical excitability increases with time awake. Cereb. Cortex 2013, 23, 332–338. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Paulus, W. Excitability Changes Induced in the Human Motor Cortex by Weak Transcranial Direct Current Stimulation. J. Psychol. 2000, 527, 633–639. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Z.; Chen, Q.; Fu, Y.; Zheng, K. Transcranial Direct Current Stimulation Enhances Cognitive Function in Patients with Mild Cognitive Impairment and Early/Mid Alzheimer’s Disease: A Systematic Review and Meta-Analysis. Brain Sci. 2022, 12, 562. [Google Scholar] [CrossRef]
- Mohammadi, A. Induction of Neuroplasticity by Transcranial Direct Current Stimulation. J. Biomed. Phys. Eng. 2016, 6, 205–208. [Google Scholar] [PubMed]
- Salehinejad, M.A.; Siniatchkin, M. Safety of noninvasive brain stimulation in children. Curr. Opin. Psychiatry 2024, 37, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.J.; Antal, A.; Bikson, M.; Boggio, P.S.; Brunoni, A.R.; Celnik, P.; Cohen, L.G.; Fregni, F.; Herrmann, C.S.; Kappenman, E.S. A technical guide to tDCS, and related non-invasive brain stimulation tools. Clin. Neurophysiol. 2016, 127, 1031–1048. [Google Scholar] [CrossRef]
- Lang, N.; Siebner, H.R.; Ward, N.S.; Lee, L.; Nitsche, M.A.; Paulus, W.; Rothwell, J.C.; Lemon, R.N.; Frackowiak, R.S. How does transcranial DC stimulation of the primary motor cortex alter regional neuronal activity in the human brain? Eur. J. Neurosci. 2005, 22, 495–504. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Fricke, K.; Henschke, U.; Schlitterlau, A.; Liebetanz, D.; Lang, N.; Henning, S.; Tergau, F.; Paulus, W. Pharmacological Modulation of Cortical Excitability Shifts Induced by Transcranial Direct Current Stimulation in Humans. J. Psychol. 2003, 553, 293–301. [Google Scholar] [CrossRef]
- Yavari, F.; Jamil, A.; Mosayebi Samani, M.; Vidor, L.P.; Nitsche, M.A. Basic and functional effects of transcranial Electrical Stimulation (tES)—An introduction. Neurosci. Biobehav. Rev. 2018, 85, 81–92. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Paulus, W. Sustained excitability elevations induced by transcranial DC motor cortex stimulation in humans. Neurology 2001, 57, 1899–1901. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Liebetanz, D.; Schlitterlau, A.; Henschke, U.; Fricke, K.; Frommann, K.; Lang, N.; Henning, S.; Paulus, W.; Tergau, F. GABAergic modulation of DC stimulation-induced motor cortex excitability shifts in humans. Eur. J. Neurosci. 2004, 19, 2720–2726. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Seeber, A.; Frommann, K.; Klein, C.C.; Rochford, C.; Nitsche, M.S.; Fricke, K.; Liebetanz, D.; Lang, N.; Antal, A.; et al. Modulating parameters of excitability during and after transcranial direct current stimulationof the human motor cortex. J. Physiol. 2005, 568 Pt 1, 291–303. [Google Scholar] [CrossRef]
- Lisman, J.E. Three Ca2+ levels affect plasticity differently: The LTP zone, the LTD zone and no man’s land. J. Physiol. 2001, 532 Pt 2, 28. [Google Scholar] [CrossRef]
- Boggio, P.S.; Rêgo, G.G.; Marques, L.M.; Costa, T.L. Transcranial Direct Current Stimulation in Social and Emotion Research. In Transcranial Direct Current Stimulation in Neuropsychiatric Disorders: Clinical Principles and Management; Brunoni, A., Nitsche, M., Loo, C., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 143–152. [Google Scholar]
- Greenwood, P.M.; Blumberg, E.J.; Scheldrup, M.R. Hypothesis for cognitive effects of transcranial direct current stimulation: Externally- and internally-directed cognition. Neurosci. Biobehav. Rev. 2018, 86, 226–238. [Google Scholar] [CrossRef]
- Fregni, F.; El-Hagrassy, M.M.; Pacheco-Barrios, K.; Carvalho, S.; Leite, J.; Simis, M.; Brunelin, J.; Nakamura-Palacios, E.M.; Marangolo, P.; Venkatasubramanian, G.; et al. Evidence-Based Guidelines and Secondary Meta-Analysis for the Use of Transcranial Direct Current Stimulation in Neurological and Psychiatric Disorders. Int. J. Neuropsychopharmacol. 2020, 24, 256–313. [Google Scholar] [CrossRef]
- Salehinejad, M.A.; Ghanavati, E.; Glinski, B.; Hallajian, A.H.; Azarkolah, A. A systematic review of randomized controlled trials on efficacy and safety of transcranial direct current stimulation in major neurodevelopmental disorders: ADHD, autism, and dyslexia. Brain Behav. 2022, 12, e2724. [Google Scholar] [CrossRef]
- Alizadehgoradel, J.; Molaei, B.; Barzegar Jalali, K.; Pouresmali, A.; Sharifi, K.; Hallajian, A.-H.; Nejati, V.; Glinski, B.; Vicario, C.M.; Nitsche, M.A.; et al. Targeting the prefrontal-supplementary motor network in obsessive-compulsive disorder with intensified electrical stimulation in two dosages: A randomized, controlled trial. Transl. Psychiatry 2024, 14, 78. [Google Scholar] [CrossRef]
- Salehinejad, M.A.; Wischnewski, M.; Ghanavati, E.; Mosayebi-Samani, M.; Kuo, M.-F.; Nitsche, M.A. Cognitive functions and underlying parameters of human brain physiology are associated with chronotype. Nat. Commun. 2021, 12, 4672. [Google Scholar] [CrossRef]
- Dondé, C.; Brunelin, J.; Micoulaud-Franchi, J.-A.; Maruani, J.; Lejoyeux, M.; Polosan, M.; Geoffroy, P.A. The effects of transcranial electrical stimulation of the brain on sleep: A systematic review. Front. Psychiatry 2021, 12, 646569. [Google Scholar] [CrossRef]
- Herrero Babiloni, A.; Bellemare, A.; Beetz, G.; Vinet, S.-A.; Martel, M.O.; Lavigne, G.J.; De Beaumont, L. The effects of non-invasive brain stimulation on sleep disturbances among different neurological and neuropsychiatric conditions: A systematic review. Sleep Med. Rev. 2021, 55, 101381. [Google Scholar] [CrossRef]
- Alfonsi, V.; D’Atri, A.; Scarpelli, S.; Gorgoni, M.; Giacinti, F.; Annarumma, L.; Salfi, F.; Amicucci, G.; Corigliano, D.; De Gennaro, L. The effects of bifrontal anodal transcranial direct current stimulation (tDCS) on sleepiness and vigilance in partially sleep-deprived subjects: A multidimensional study. J. Sleep Res. 2023, 32, e13869. [Google Scholar] [CrossRef]
- Reddy, A.B.; O’Neill, J.S. Healthy clocks, healthy body, healthy mind. Trends Cell Biol. 2010, 20, 36–44. [Google Scholar] [CrossRef]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat. Rev. Neurosci. 2018, 19, 453–469. [Google Scholar] [CrossRef]
- Göz, D.; Studholme, K.; Lappi, D.A.; Rollag, M.D.; Provencio, I.; Morin, L.P. Targeted destruction of photosensitive retinal ganglion cells with a saporin conjugate alters the effects of light on mouse circadian rhythms. PLoS ONE 2008, 3, e3153. [Google Scholar] [CrossRef]
- Dibner, C.; Schibler, U.; Albrecht, U. The mammalian circadian timing system: Organization and coordination of central and peripheral clocks. Annu. Rev. Physiol. 2010, 72, 517–549. [Google Scholar] [CrossRef]
- Rosenwasser, A.M. Functional neuroanatomy of sleep and circadian rhythms. Brain Res. Rev. 2009, 61, 281–306. [Google Scholar] [CrossRef]
- Swanson, L.W.; Cowan, W.M.; Jones, E.G. An autoradiographic study of the efferent projections of the ventral lateral geniculate nucleus in the albino rat and cat. J. Comp. Neurol. 1974, 156, 143–164. [Google Scholar] [CrossRef]
- Morin, L.P. Neuroanatomy of the extended circadian rhythm system. Exp. Neurol. 2013, 243, 4–20. [Google Scholar] [CrossRef]
- Albers, H.E.; Ferris, C.F. Neuropeptide Y: Role in light-dark entrainment of hamster circadian rhythms. Neurosci. Lett. 1984, 50, 163–168. [Google Scholar] [CrossRef]
- Zeitzer, J.M.; Dijk, D.J.; Kronauer, R.; Brown, E.; Czeisler, C. Sensitivity of the human circadian pacemaker to nocturnal light: Melatonin phase resetting and suppression. J. Physiol. 2000, 526 Pt 3, 695–702. [Google Scholar] [CrossRef]
- Mul Fedele, M.L.; Galiana, M.D.; Golombek, D.A.; Muñoz, E.M.; Plano, S.A. Alterations in Metabolism and Diurnal Rhythms following Bilateral Surgical Removal of the Superior Cervical Ganglia in Rats. Front. Endocrinol. 2018, 8, 370. [Google Scholar] [CrossRef]
- Starnes, A.N.; Jones, J.R. Inputs and Outputs of the Mammalian Circadian Clock. Biology 2023, 12, 508. [Google Scholar] [CrossRef]
- Van Drunen, R.; Eckel-Mahan, K. Circadian Rhythms of the Hypothalamus: From Function to Physiology. Clocks Sleep 2021, 3, 189–226. [Google Scholar] [CrossRef]
- Andreadi, A.; Andreadi, S.; Todaro, F.; Ippoliti, L.; Bellia, A.; Magrini, A.; Chrousos, G.P.; Lauro, D. Modified Cortisol Circadian Rhythm: The Hidden Toll of Night-Shift Work. Int. J. Mol. Sci. 2025, 26, 2090. [Google Scholar] [CrossRef]
- Bowles, N.P.; Thosar, S.S.; Butler, M.P.; Clemons, N.A.; Robinson, L.D.; Ordaz, O.H.; Herzig, M.X.; McHill, A.W.; Rice, S.P.M.; Emens, J.; et al. The circadian system modulates the cortisol awakening response in humans. Front. Neurosci. 2022, 16, 995452. [Google Scholar] [CrossRef] [PubMed]
- Oster, H.; Challet, E.; Ott, V.; Arvat, E.; de Kloet, E.R.; Dijk, D.J.; Lightman, S.; Vgontzas, A.; Van Cauter, E. The Functional and Clinical Significance of the 24-Hour Rhythm of Circulating Glucocorticoids. Endocr. Rev. 2017, 38, 3–45. [Google Scholar] [CrossRef] [PubMed]
- Joëls, M.; Karst, H.; Tasker, J.G. The emerging role of rapid corticosteroid actions on excitatory and inhibitory synaptic signaling in the brain. Front. Neuroendocrinol. 2024, 74, 101146. [Google Scholar] [CrossRef] [PubMed]
- Hannibal, K.E.; Bishop, M.D. Chronic stress, cortisol dysfunction, and pain: A psychoneuroendocrine rationale for stress management in pain rehabilitation. Phys. Ther. 2014, 94, 1816–1825. [Google Scholar] [CrossRef]
- Ly, J.Q.; Gaggioni, G.; Chellappa, S.L.; Papachilleos, S.; Brzozowski, A.; Borsu, C.; Rosanova, M.; Sarasso, S.; Middleton, B.; Luxen, A.; et al. Circadian regulation of human cortical excitability. Nat. Commun. 2016, 7, 11828. [Google Scholar] [CrossRef]
- Henry, M.; Thomas, K.G.F.; Ross, I.L. Sleep, Cognition and Cortisol in Addison’s Disease: A Mechanistic Relationship. Front. Endocrinol. 2021, 12, 694046. [Google Scholar] [CrossRef]
- Gobbo, M.; Falciati, L. Neuromodulation and cognitive rehabilitation: Addressing the methodological issue of circadian rhythms. Front. Psychiatry 2014, 5, 150. [Google Scholar] [CrossRef]
- Chan, H.K.C. Circadian Rhythms and Memory: Exploring the Mechanisms of Synaptic Plasticity, Hormonal Modulation, and Cognitive Function. Theor. Nat. Sci. 2025, 77, 98–106. [Google Scholar] [CrossRef]
- Sandi, C. Memory Impairments Associated with Stress and Aging. In Neural Plasticity and Memory: From Genes to Brain Imaging; Bermúdez-Rattoni, F., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2007; Chapter 12. Available online: https://www.ncbi.nlm.nih.gov/books/NBK3914/ (accessed on 27 May 2025).
- Finsterwald, C.; Alberini, C.M. Stress and glucocorticoid receptor-dependent mechanisms in long-term memory: From adaptive responses to psychopathologies. Neurobiol. Learn. Mem. 2014, 112, 17–29. [Google Scholar] [CrossRef]
- Munnilari, M.; Bommasamudram, T.; Easow, J.; Tod, D.; Varamenti, E.; Edwards, B.J.; Ravindrakumar, A.; Gallagher, C.; Pullinger, S.A. Diurnal variation in variables related to cognitive performance: A systematic review. Sleep Breath. 2024, 28, 495–510. [Google Scholar] [CrossRef]
- Johnson, M.P.; Duffy, J.F.; Dijk, D.J.; Ronda, J.M.; Dyal, C.M.; Czeisler, C.A. Short-term memory, alertness and performance: A reappraisal of their relationship to body temperature. J. Sleep Res. 1992, 1, 24–29. [Google Scholar] [CrossRef]
- Hidalgo, V.; Almela, M.; Pulopulos, M.M.; Salvador, A. Memory performance is related to the cortisol awakening response in older people, but not to the diurnal cortisol slope. Psychoneuroendocrinology 2016, 71, 136–146. [Google Scholar] [CrossRef]
- Xiong, B.; Chen, C.; Tian, Y.; Zhang, S.; Liu, C.; Evans, T.M.; Fernández, G.; Wu, J.; Qin, S. Brain preparedness: The proactive role of the cortisol awakening response in hippocampal-prefrontal functional interactions. Prog. Neurobiol. 2021, 205, 102127. [Google Scholar] [CrossRef]
- Reser, J.E. Chronic stress, cortical plasticity and neuroecology. Behav. Processes 2016, 129, 105–115. [Google Scholar] [CrossRef]
- Sapolsky, R.M. Glucocorticoids, stress, and their adverse neurological effects: Relevance to aging. Exp. Gerontol. 1999, 34, 721–732. [Google Scholar] [CrossRef]
- Sale, M.V.; Ridding, M.C.; Nordstrom, M.A. Cortisol inhibits neuroplasticity induction in human motor cortex. J. Neurosci. 2008, 28, 8285–8293. [Google Scholar] [CrossRef]
- Clow, A.; Law, R.; Evans, P.; Vallence, A.M.; Hodyl, N.A.; Goldsworthy, M.R.; Rothwell, J.R.; Ridding, M.C. Day differences in the cortisol awakening response predict day differences in synaptic plasticity in the brain. Stress 2014, 17, 219–223. [Google Scholar] [CrossRef]
- Zhu, L.; Zee, P.C. Circadian rhythm sleep disorders. Neurol. Clin. 2012, 30, 1167–1191. [Google Scholar] [CrossRef]
- Geneva, I. Human Body Temperature Circadian Rhythm in Health and Disease; IntechOpen: Rijeka, Croatia, 2024. [Google Scholar] [CrossRef]
- Frank, M.G. Circadian Regulation of Synaptic Plasticity. Biology 2016, 5, 31. [Google Scholar] [CrossRef]
- Molzof, H.E.; Prapanjaroensin, A.; Patel, V.H.; Mokashi, M.V.; Gamble, K.L.; Patrician, P.A. Misaligned core body temperature rhythms impact cognitive performance of hospital shift work nurses. Neurobiol. Learn. Mem. 2019, 160, 151–159. [Google Scholar] [CrossRef]
- Christoforou, R.; Pallubinsky, H.; Maria, T.; Rewitz, K.; El-Mokadem, M.; Bardey, J.; Müller, D.; Schweiker, M. O-369 The effects of constant ambient temperature on alertness: Potential benefits of dynamic conditions in offices. Occup. Med. 2024, 74 (Suppl. 1), 0. [Google Scholar] [CrossRef]
- Lodovichi, C.; Ratto, G.M. Control of circadian rhythm on cortical excitability and synaptic plasticity. Front. Neural Circuits 2023, 17, 1099598. [Google Scholar] [CrossRef] [PubMed]
- Refinetti, R. Circadian rhythmicity of body temperature and metabolism. Temperature 2020, 7, 321–362. [Google Scholar] [CrossRef] [PubMed]
- Panda, S. Circadian physiology of metabolism. Science 2016, 354, 1008–1015. [Google Scholar] [CrossRef]
- Kim, K.J. The Role of Circadian Clocks in Metabolism. Chronobiol. Med. 2019, 1, 107–110. [Google Scholar] [CrossRef]
- Kiehn, J.T.; Faltraco, F.; Palm, D.; Thome, J.; Oster, H. Circadian Clocks in the Regulation of Neurotransmitter Systems. Pharmacopsychiatry 2023, 56, 108–117. [Google Scholar] [CrossRef]
- Ciarleglio, C.M.; Resuehr, H.E.; McMahon, D.G. Interactions of the serotonin and circadian systems: Nature and nurture in rhythms and blues. Neuroscience 2011, 197, 8–16. [Google Scholar] [CrossRef]
- Pontes, A.L.B.; Engelberth, R.C.G.J.; Nascimento, E.D.S., Jr.; Cavalcante, J.C.; Costa, M.S.M.D.O.; Pinato, L.; Toledo, C.A.B.D.; Cavalcante, J.D.S. Serotonin and circadian rhythms. Psychol. Neurosci. 2010, 3, 217–228. [Google Scholar] [CrossRef]
- Estrada-Rojo, F.; Escobar, C.; Navarro, L. Circadian variations of neurotransmitters in the brain—Its importance for neuroprotection. Rev. Mex. Neurocienc. 2020, 21, 31–38. [Google Scholar] [CrossRef]
- Reghunandana, V. Functional interactions between neurotransmitters and neuropeptides in regulating suprachiasmatic nucleus function and circadian rhythms. Explor. Neurosci. 2024, 3, 434–477. [Google Scholar] [CrossRef]
- Valdez, P. Circadian Rhythms in Attention. Yale J. Biol. Med. 2019, 92, 81–92. [Google Scholar] [PubMed]
- Smarr, B.L.; Jennings, K.J.; Driscoll, J.R.; Kriegsfeld, L.J. A time to remember: The role of circadian clocks in learning and memory. Behav. Neurosci. 2014, 128, 283–303. [Google Scholar] [CrossRef] [PubMed]
- Hahn, C.; Cowell, J.M.; Wiprzycka, U.J.; Goldstein, D.; Ralph, M.; Hasher, L.; Zelazo, P.D. Circadian rhythms in executive function during the transition to adolescence: The effect of synchrony between chronotype and time of day. Dev. Sci. 2012, 15, 408–416. [Google Scholar] [CrossRef]
- Correa, Á.; Alguacil, S.; Ciria, L.F.; Jiménez, A.; Ruz, M. Circadian rhythms and decision-making: A review and new evidence from electroencephalography. Chronobiol. Int. 2020, 37, 520–541. [Google Scholar] [CrossRef]
- Wright, K.P.; Lowry, C.A.; Lebourgeois, M.K. Circadian and wakefulness-sleep modulation of cognition in humans. Front. Mol. Neurosci. 2012, 5, 50. [Google Scholar] [CrossRef]
- Xu, S.; Akioma, M.; Yuan, Z. Relationship between circadian rhythm and brain cognitive functions. Front. Optoelectron. 2021, 14, 278–287. [Google Scholar] [CrossRef]
- Adan, A.; Archer, S.N.; Hidalgo, M.P.; Di Milia, L.; Natale, V.; Randler, C. Circadian typology: A comprehensive review. Chronobiol. Int. 2012, 29, 1153–1175. [Google Scholar] [CrossRef]
- Sylvester, C.M.; Krout, K.E.; Loewy, A.D. Suprachiasmatic nucleus projection to the medial prefrontal cortex: A viral transneuronal tracing study. Neuroscience 2002, 114, 1071–1080. [Google Scholar] [CrossRef]
- Snider, K.H.; Sullivan, K.A.; Obrietan, K. Circadian Regulation of Hippocampal-Dependent Memory: Circuits, Synapses, and Molecular Mechanisms. Neural Plast. 2018, 2018, 7292540. [Google Scholar] [CrossRef]
- Harding, C.; Bechtold, D.A.; Brown, T.M. Suprachiasmatic nucleus-dependent and independent outputs driving rhythmic activity in hypothalamic and thalamic neurons. BMC Biol. 2020, 18, 134. [Google Scholar] [CrossRef]
- Anand, K.S.; Dhikav, V. Hippocampus in health and disease: An overview. Ann. Indian Acad. Neurol. 2012, 15, 239–246. [Google Scholar] [CrossRef]
- Lim, J.; Dinges, D.F. A meta-analysis of the impact of short-term sleep deprivation on cognitive variables. Psychol. Bull. 2010, 136, 375–389. [Google Scholar] [CrossRef]
- Salehinejad, M.A.; Ghanavati, E.; Reinders, J.; Hengstler, J.G.; Kuo, M.-F.; Nitsche, M.A. Sleep-dependent upscaled excitability, saturated neuroplasticity, and modulated cognition in the human brain. eLife 2022, 11, e69308. [Google Scholar] [CrossRef]
- Wong, L.Y.X.; Gray, S.J.; Gallo, D.A. Does tDCS over prefrontal cortex improve episodic memory retrieval? Potential importance of time of day. Cogn. Neurosci. 2018, 9, 167–180. [Google Scholar] [CrossRef]
- Martini, B.; Buffington, A.L.; Welsh-Bohmer, K.A.; Brandt, J.; ADAPT Research Group. Time of day affects episodic memory in older adults. Neuropsychol. Dev. Cogn. B Aging Neuropsychol. Cogn. 2008, 15, 146–164. [Google Scholar] [CrossRef]
- Goel, N.; Basner, M.; Rao, H.; Dinges, D.F. Circadian rhythms, sleep deprivation, and human performance. Prog. Mol. Biol. Transl. Sci. 2013, 119, 155–190. [Google Scholar] [PubMed]
- Blundell, J.; Kouser, M.; Powell, C.M. Systemic inhibition of mammalian target of rapamycin inhibits fear memory reconsolidation. Neurobiol. Learn. Mem. 2008, 90, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.; Anderson, F.E.; Jung, Y.J.; Dziema, H.; Obrietan, K. Circadian regulation of mammalian target of rapamycin signaling in the mouse suprachiasmatic nucleus. Neuroscience 2011, 181, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Buffington, S.A.; Huang, W.; Costa-Mattioli, M. Translational control in synaptic plasticity and cognitive dysfunction. Annu. Rev. Neurosci. 2014, 37, 17–38. [Google Scholar] [CrossRef]
- Nguyen, L.H.; Bordey, A. Convergent and divergent mechanisms of epileptogenesis in mTORopathies. Front. Neuroanat. 2021, 15, 664695. [Google Scholar] [CrossRef] [PubMed]
- McCauley, J.P.; Petroccione, M.A.; D’Brant, L.Y.; Todd, G.C.; Affinnih, N.; Wisnoski, J.J.; Zahid, S.; Shree, S.; Sousa, A.A.; De Guzman, R.M.; et al. Circadian modulation of neurons and astrocytes controls synaptic plasticity in hippocampal area CA. Cell Rep. 2020, 33, 108255. [Google Scholar] [CrossRef] [PubMed]
- Goode, L.K.; Fusilier, A.R.; Remiszewski, N.; Reeves, J.M.; Abiraman, K.; Defenderfer, M.; Paul, J.R.; McMahon, L.L.; Gamble, K.L. Examination of diurnal variation and sex differences in hippocampal neurophysiology and spatial memory. ENeuro 2022, 9, ENEURO.0124-22.2022. [Google Scholar] [CrossRef]
- Romei, V.; Rihs, T.; Brodbeck, V.; Thut, G. Resting electroencephalogram alpha-power over posterior sites indexes baseline visual cortex excitability. Neuroreport 2008, 19, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Zrenner, C.; Desideri, D.; Belardinelli, P.; Ziemann, U. Real-time EEG-defined excitability states determine efficacy of TMS-induced plasticity in human motor cortex. Brain Stimul. 2018, 11, 374–389. [Google Scholar] [CrossRef]
- McIntire, L.K.; McKinley, R.A.; Goodyear, C.; Nelson, J. A comparison of the effects of transcranial direct current stimulation and caffeine on vigilance and cognitive performance during extended wakefulness. Brain Stimul. 2014, 7, 499–507. [Google Scholar] [CrossRef]
- McIntire, L.K.; McKinley, R.A.; Nelson, J.M.; Goodyear, C. Transcranial direct current stimulation versus caffeine as a fatigue countermeasure. Brain Stimul. 2017, 10, 1070–1078. [Google Scholar] [CrossRef]
- Beldzik, E.; Yang, Z.; Williams, S.; Lewis, L. 0182 Distinct Spectral Pattern of Cognitive, Drowsiness, and Fatigue-related Theta/alpha EEG Activity During Wakefulness. Sleep 2024, 47 (Suppl. 1), A78–A79. [Google Scholar] [CrossRef]
- Vyazovskiy, V.V.; Tobler, I. Theta activity in the waking EEG is a marker of sleep propensity in the rat. Brain Res. 2005, 1050, 64–71. [Google Scholar] [CrossRef]
- Chatburn, A.; Lushington, K.; Cross, Z.R. Considerations towards a neurobiologically-informed EEG measurement of sleepiness. Brain Res. 2024, 1841, 149088. [Google Scholar] [CrossRef]
- Snipes, S.; Krugliakova, E.; Meier, E.; Huber, R. The Theta Paradox: 4-8 Hz EEG Oscillations Reflect Both Sleep Pressure and Cognitive Control. J. Neurosci. 2022, 42, 8569–8586. [Google Scholar] [CrossRef] [PubMed]
- Fattinger, S.; Kurth, S.; Ringli, M.; Jenni, O.G.; Huber, R. Theta waves in children’s waking electroencephalogram resemble local aspects of sleep during wakefulness. Sci. Rep. 2017, 7, 11187. [Google Scholar] [CrossRef] [PubMed]
- Cajochen, C.; Brunner, D.P.; Krauchi, K.; Graw, P.; Wirz-Justice, A. Power density in theta/alpha frequencies of the waking EEG progressively increases during sustained wakefulness. Sleep 1995, 18, 890–894. [Google Scholar] [CrossRef] [PubMed]
- Aeschbach, D.; Matthews, J.R.; Postolache, T.T.; Jackson, M.A.; Giesen, H.A.; Wehr, T.A. Dynamics of the human EEG during prolonged wakefulness: Evidence for frequency-specific circadian and homeostatic influences. Neurosci. Lett. 1997, 239, 121–124. [Google Scholar] [CrossRef]
- Finelli, L.A.; Baumann, H.; Borbély, A.A.; Achermann, P. Dual electroencephalogram markers of human sleep homeostasis: Correlation between theta activity in waking and slow-wave activity in sleep. Neuroscience 2000, 101, 523–529. [Google Scholar] [CrossRef]
- Hung, C.S.; Sarasso, S.; Ferrarelli, F.; Riedner, B.; Ghilardi, M.F.; Cirelli, C.; Tononi, G. Local experience-dependent changes in the wake EEG after prolonged wakefulness. Sleep 2013, 36, 59–72. [Google Scholar] [CrossRef]
- Vyazovskiy, V.V.; Olcese, U.; Lazimy, Y.M.; Faraguna, U.; Esser, S.K.; Williams, J.C.; Cirelli, C.; Tononi, G. Cortical firing and sleep homeostasis. Neuron 2009, 63, 865–878. [Google Scholar] [CrossRef]
- Fehér, K.D.; Omlin, X.; Tarokh, L.; Schneider, C.L.; Morishima, Y.; Züst, M.A.; Wunderlin, M.; Koenig, T.; Hertenstein, E.; Ellenberger, B.; et al. Feasibility, efficacy, and functional relevance of automated auditory closed-loop suppression of slow-wave sleep in humans. J. Sleep Res. 2023, 32, e13846. [Google Scholar] [CrossRef]
- Yamada, Y.; Sumiyoshi, T. Neurobiological Mechanisms of Transcranial Direct Current Stimulation for Psychiatric Disorders; Neurophysiological, Chemical, and Anatomical Considerations. Front. Hum. Neurosci. 2021, 15, 631838. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, Y.; Zhang, Y.; Jiao, J.; Tang, X. Cortical excitability on sleep deprivation measured by transcranial magnetic stimulation: A systematic review and meta-analysis. Brain Res. Bull. 2025, 221, 111190. [Google Scholar] [CrossRef]
- Mroczek, M.; de Grado, A.; Pia, H.; Nochi, Z.; Tankisi, H. Effects of sleep deprivation on cortical excitability: A threshold-tracking TMS study and review of the literature. Clin. Neurophysiol. Pract. 2023, 9, 13–20. [Google Scholar] [CrossRef]
- Civardi, C.; Boccagni, C.; Vicentini, R.; Bolamperti, L.; Tarletti, R.; Varrasi, C.; Monaco, F.; Cantello, R. Cortical excitability and sleep deprivation: A transcranial magnetic stimulation study. J. Neurol. Neurosurg. Psychiatry 2001, 71, 809–812. [Google Scholar] [CrossRef] [PubMed]
- De Gennaro, L.; Marzano, C.; Veniero, D.; Moroni, F.; Fratello, F.; Curcio, G.; Ferrara, M.; Ferlazzo, F.; Novelli, L.; Concetta Pellicciari, M.; et al. Neurophysiological correlates of sleepiness: A combined TMS and EEG study. Neuroimage 2007, 36, 1277–1287. [Google Scholar] [CrossRef] [PubMed]
- Chia, C.H.; Tang, X.W.; Cao, Y.; Cao, H.T.; Zhang, W.; Wu, J.F.; Zhu, Y.L.; Chen, Y.; Lin, Y.; Wu, Y.; et al. Cortical excitability signatures for the degree of sleepiness in human. Elife 2021, 10, e65099. [Google Scholar] [CrossRef] [PubMed]
- Marshall, L.; Helgadóttir, H.; Mölle, M.; Born, J. Boosting slow oscillations during sleep potentiates memory. Nature 2006, 444, 610–613. [Google Scholar] [CrossRef]
- Salehinejad, M.A.; Ghanavati, E.; Kuo, M.F.; Nitsche, M.A. The role of circadian preferred time of day and sleep pressure in tDCS-induced neuroplasticity and associated cognition. Brain Stimul. Basic Transl. Clin. Res. Neuromodul. 2023, 16, 203–204. [Google Scholar] [CrossRef]
- Meldrum, B.S. Glutamate as a neurotransmitter in the brain: Review of physiology and pathology. J. Nutr. 2000, 130 (Suppl. S4), 1007S–1015S. [Google Scholar] [CrossRef]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate receptor ion channels: Structure, regulation, and function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef]
- Stagg, C.J.; Nitsche, M.A. Physiological basis of transcranial direct current stimulation. Neuroscientist 2011, 17, 37–53. [Google Scholar] [CrossRef]
- Alvarez-Alvarado, S.; Boutzoukas, E.M.; Kraft, J.N.; O’Shea, A.; Indahlastari, A.; Albizu, A.; Nissim, N.R.; Evangelista, N.D.; Cohen, R.; Porges, E.C.; et al. Impact of Transcranial Direct Current Stimulation and Cognitive Training on Frontal Lobe Neurotransmitter Concentrations. Front. Aging Neurosci. 2021, 13, 761348. [Google Scholar] [CrossRef]
- Farrant, M.; Nusser, Z. Variations on an inhibitory theme: Phasic and tonic activation of GABA(A) receptors. Nat. Rev. Neurosci. 2005, 6, 215–229. [Google Scholar] [CrossRef]
- Bunai, T.; Hirosawa, T.; Kikuchi, M.; Fukai, M.; Yokokura, M.; Ito, S.; Takata, Y.; Terada, T.; Ouchi, Y. tDCS-induced modulation of GABA concentration and dopamine release in the human brain: A combination study of magnetic resonance spectroscopy and positron emission tomography. Brain Stimul. 2021, 14, 154–160. [Google Scholar] [CrossRef]
- Ono, D.; Honma, K.I.; Yanagawa, Y.; Yamanaka, A.; Honma, S. Role of GABA in the regulation of the central circadian clock of the suprachiasmatic nucleus. J. Physiol. Sci. 2018, 68, 333–343. [Google Scholar] [CrossRef]
- Sandhu, M.R.S.; Dhaher, R.; Gruenbaum, S.E.; Raaisa, R.; Spencer, D.D.; Pavlova, M.K.; Zaveri, H.P.; Eid, T. Circadian-Like Rhythmicity of Extracellular Brain Glutamate in Epilepsy. Front. Neurol. 2020, 11, 398. [Google Scholar] [CrossRef]
- Dickmeis, T. Glucocorticoids and the circadian clock. J. Endocrinol. 2009, 200, 3–22. [Google Scholar] [CrossRef]
- Kirschbaum, C.; Hellhammer, D.H. Salivary cortisol in psychoneuroendocrine research: Recent developments and applications. Psychoneuroendocrinology 1994, 19, 313–333. [Google Scholar] [CrossRef]
- Clow, A.; Thorn, L.; Evans, P.; Hucklebridge, F. The awakening cortisol response: Methodological issues and significance. Stress 2004, 7, 29–37. [Google Scholar] [CrossRef]
- Milani, P.; Piu, P.; Popa, T.; della Volpe, R.; Bonifazi, M.; Rossi, A.; Mazzocchio, R. Cortisol-induced effects on human cortical excitability. Brain Stimul. 2010, 3, 131–139. [Google Scholar] [CrossRef]
- Li, L.M.; Uehara, K.; Hanakawa, T. The contribution of interindividual factors to variability of response in transcranial direct current stimulation studies. Front. Cell Neurosci. 2015, 9, 181. [Google Scholar] [CrossRef]
- Lang, N.; Rothkegel, H.; Reiber, H.; Hasan, A.; Sueske, E.; Tergau, F.; Ehrenreich, H.; Wuttke, W.; Paulus, W. Circadian modulation of GABA-mediated cortical inhibition. Cereb. Cortex 2011, 21, 2299–2306. [Google Scholar] [CrossRef]
- Roy, S.; Fan, Y.; Mosayebi-Samani, M.; Claus, M.; Yavari, F.; Kleinsorge, T.; Nitsche, M.A. Modulating prefrontal cortex activity to alleviate stress-induced working memory deficits: A transcranial direct current (tDCS) study. Int. J. Clin. Health Psychol. 2025, 25, 100569. [Google Scholar] [CrossRef]
- Ankri, Y.L.; Braw, Y.; Luboshits, G.; Meiron, O. The effects of stress and transcranial direct current stimulation (tDCS) on working memory: A randomized controlled trial. Cogn. Affect. Behav. Neurosci. 2020, 20, 103–114. [Google Scholar] [CrossRef]
- Brunoni, A.R.; Vanderhasselt, M.A.; Boggio, P.S.; Fregni, F.; Dantas, E.M.; Mill, J.G.; Lotufo, P.A.; Benseñor, I.M. Polarity- and valence-dependent effects of prefrontal transcranial direct current stimulation on heart rate variability and salivary cortisol. Psychoneuroendocrinology 2013, 38, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Vignaud, P.; Adam, O.; Palm, U.; Baeken, C.; Prieto, N.; Poulet, E.; Brunelin, J. Can a single session of noninvasive brain stimulation applied over the prefrontal cortex prevent stress-induced cortisol release? Prog. Neuropsychopharmacol. Biol. Psychiatry 2023, 121, 110667. [Google Scholar] [CrossRef]
- Littmann, A.E.; Shields, R.K. Whole body heat stress increases motor cortical excitability and skill acquisition in humans. Clin. Neurophysiol. 2016, 127, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Jafari, E.; Alizadehgoradel, J.; Pourmohseni Koluri, F.; Nikoozadehkordmirza, E.; Refahi, M.; Taherifard, M.; Nejati, V.; Hallajian, A.-H.; Ghanavati, E.; Vicario, C.M.; et al. Intensified electrical stimulation targeting lateral and medial prefrontal cortices for the treatment of social anxiety disorder: A randomized, double-blind, parallel-group, dose-comparison study. Brain Stimul. 2021, 14, 974–986. [Google Scholar] [CrossRef]
- Glinski, B.; Salehinejad, M.A.; Takahashi, K.; Kuo, M.-F.; Nitsche, M. Induction and stabilization of gamma oscillations in the human brain. Brain Stimul. Basic Transl. Clin. Res. Neuromodul. 2023, 16, 347–348. [Google Scholar] [CrossRef]
- Salehinejad, M.A.; Abdi, M.; Dadashi, M.; Zolghadriha, A.; Salvador, R.; Ruffini, G.; Nitsche, M.A. Optimized Multichannel tDCS Protocol for Clinical Use in Patients with Major Depressive Disorder: A Randomized, Controlled Trial. medRxiv 2025. [Google Scholar] [CrossRef]
- Saleh, O.; Assaf, M.; Alzoubi, A.; Anshase, A.; Tarkhan, H.; Ayoub, M.; Abuelazm, M. The effects of transcranial direct current stimulation on cognitive function for mild cognitive impairment: A systematic review and meta-analysis of randomized controlled trials. Aging Clin. Exp. Res. 2023, 35, 2293–2306. [Google Scholar] [CrossRef]
- Mentzelou, M.; Papadopoulou, S.K.; Papandreou, D.; Spanoudaki, M.; Dakanalis, A.; Vasios, G.K.; Voulgaridou, G.; Pavlidou, E.; Mantzorou, M.; Giaginis, C. Evaluating the Relationship between Circadian Rhythms and Sleep, Metabolic and Cardiovascular Disorders: Current Clinical Evidence in Human Studies. Metabolites 2023, 13, 370. [Google Scholar] [CrossRef]
- Frase, L.; Piosczyk, H.; Zittel, S.; Jahn, F.; Selhausen, P.; Krone, L.; Feige, B.; Mainberger, F.; Maier, J.G.; Kuhn, M.; et al. Modulation of Total Sleep Time by Transcranial Direct Current Stimulation (tDCS). Neuropsychopharmacology 2016, 41, 2577–2586. [Google Scholar] [CrossRef]
- Jung, K.; Jun, J. Efficacy of transcranial direct-current stimulation on chronic insomnia. Brain Stimul. 2019, 12, 557. [Google Scholar] [CrossRef]
- McIntire, L.K.; McKinley, R.A.; Goodyear, C.; McIntire, J.P. The Effects of Anodal Transcranial Direct Current Stimulation on Sleep Time and Efficiency. Front. Hum. Neurosci. 2020, 14, 357. [Google Scholar] [CrossRef] [PubMed]
- San-Juan, D.; Mas, R.N.M.; Gutiérrez, C.; Morales, J.; Díaz, A.; Quiñones, G.; Galindo, A.K.; Baigts, L.A.; Ximenez-Camilli, C.; Anschel, D. Effect of the anodal transcranial direct current electrical stimulation on cognition of medical residents with acute sleep deprivation. Sleep Sci. 2022, 15, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.X.; Zhao, X.; Qiu, J.; Jiang, Y.; Ren, J.; Sun, S.; Wang, R.; Su, C. Effects of transcranial direct current stimulation on performance and recovery sleep during acute sleep deprivation: A pilot study. Sleep Med. 2021, 79, 124–133. [Google Scholar] [CrossRef]
- Dalong, G.; Jiyuan, L.; Ying, Z.; Lei, Z.; Yanhong, H.; Yongcong, S. Transcranial direct current stimulation reconstructs diminished thalamocortical connectivity during prolonged resting wakefulness: A resting-state fMRI pilot study. Brain Imaging Behav. 2020, 14, 278–288. [Google Scholar] [CrossRef]

{kind=link}
| tDCS Implication | Circadian Influence | Peak Performance Time | Cognitive Function | |
|---|---|---|---|---|
| Morning tDCS may enhance attention-biased tasks | Higher cortical excitability and fast reaction times | Morning | Attention and Alertness | 1 |
| Midday tDCS could optimise working memory improvements | Glutamate levels peak, enhancing short-term information processing | Late Morning to Afternoon | Working Memory | 2 |
| Afternoon tDCS may maximise executive function gains | Enhanced prefrontal cortex function for problem-solving | Afternoon | Executive Function | 3 |
| Late afternoon stimulation may enhance motor skill learning | Increased plasticity in motor regions, optimal for skill learning | Late Afternoon to Evening | Motor Learning | 4 |
| Evening tDCS could facilitate memory consolidation | Higher hippocampal activity, improved memory retention | Evening to Night | Memory Consolidation | 5 |
| Factor | Consideration | Practical Recommendation |
|---|---|---|
| Chronotype | Stimulation during morning circadian phases enhances efficacy for most individuals post-sleep recovery | Schedule tDCS ~2–3 h after habitual wake time (CT + 3 h) to align with the morning excitability peak |
| Time of Day | Aligning tDCS with cognitive peaks enhances effectiveness | Time sessions when the targeted cognitive domain naturally peaks |
| Sleep Deprivation | Sleep deprivation alters cortical excitability and can reverse tDCS effects | Avoid tDCS after < 6 h of sleep; if so, postpone until after a full night’s recovery sleep |
| Neurotransmitter Levels | tDCS interacts with glutamate/GABA balance; optimal timing depends on current neurochemical milieu | Leverage EEG or neuroimaging to identify when glutamate/GABA ratios favour LTP-like plasticity |
| Task-Specific Stimulation | Stimulation should target the cognitive function naturally peaking at that circadian phase | Combine tDCS with task-specific training during the corresponding peak window for maximal transfer |
| Misaligned tDCS | Circadian-Aligned tDCS | Outcome Measure | Benefit Domain |
|---|---|---|---|
| Blunted or reversed plasticity; anodal fails to induce LTP, cathodal may paradoxically induce facilitation | Robust polarity-specific LTP/LTD, prolonged after-effects (up to ≥90 min) | Magnitude and duration of LTP-like after-effects | Neuroplasticity |
| Impaired learning, memory, and attentional performance after misaligned/timed poorly relative to chronotype | Faster motor skill acquisition; ↑ accuracy and speed in WM and attention tasks | Motor learning (SRTT), working memory (3-back), attention (Stroop) | Cognitive Performance |
| No change or further phase delays; fragmented sleep persists | Phase-angle misalignment (∼–45 min earlier melatonin onset), ↑ nightly sleep efficiency | Melatonin onset, sleep efficiency | Sleep and Circadian Health |
| No improvement or potential worsening in mood/inflammation | Self-rated depressive/anxiety scores; ↓ systemic inflammation markers (e.g., IL-6) | Depressive/anxiety symptoms, inflammatory markers | Mood and Disease Risk |
| High inter-subject and inter-session variability; poor reproducibility | Variability across sessions; more predictable polarity-specific effects | Inter-session variability and reproducibility of tDCS outcomes | Therapeutic Consistency |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chmiel, J.; Malinowska, A. The Influence of Circadian Rhythms on Transcranial Direct Current Stimulation (tDCS) Effects: Theoretical and Practical Considerations. Cells 2025, 14, 1152. https://doi.org/10.3390/cells14151152
Chmiel J, Malinowska A. The Influence of Circadian Rhythms on Transcranial Direct Current Stimulation (tDCS) Effects: Theoretical and Practical Considerations. Cells. 2025; 14(15):1152. https://doi.org/10.3390/cells14151152
Chicago/Turabian StyleChmiel, James, and Agnieszka Malinowska. 2025. "The Influence of Circadian Rhythms on Transcranial Direct Current Stimulation (tDCS) Effects: Theoretical and Practical Considerations" Cells 14, no. 15: 1152. https://doi.org/10.3390/cells14151152
APA StyleChmiel, J., & Malinowska, A. (2025). The Influence of Circadian Rhythms on Transcranial Direct Current Stimulation (tDCS) Effects: Theoretical and Practical Considerations. Cells, 14(15), 1152. https://doi.org/10.3390/cells14151152