
Hyperalgesia in the Psychological Stress-Induced Fibromyalgia Model Shows Sexual Dimorphism Mediated by LPA1 and LPA3



Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experiments
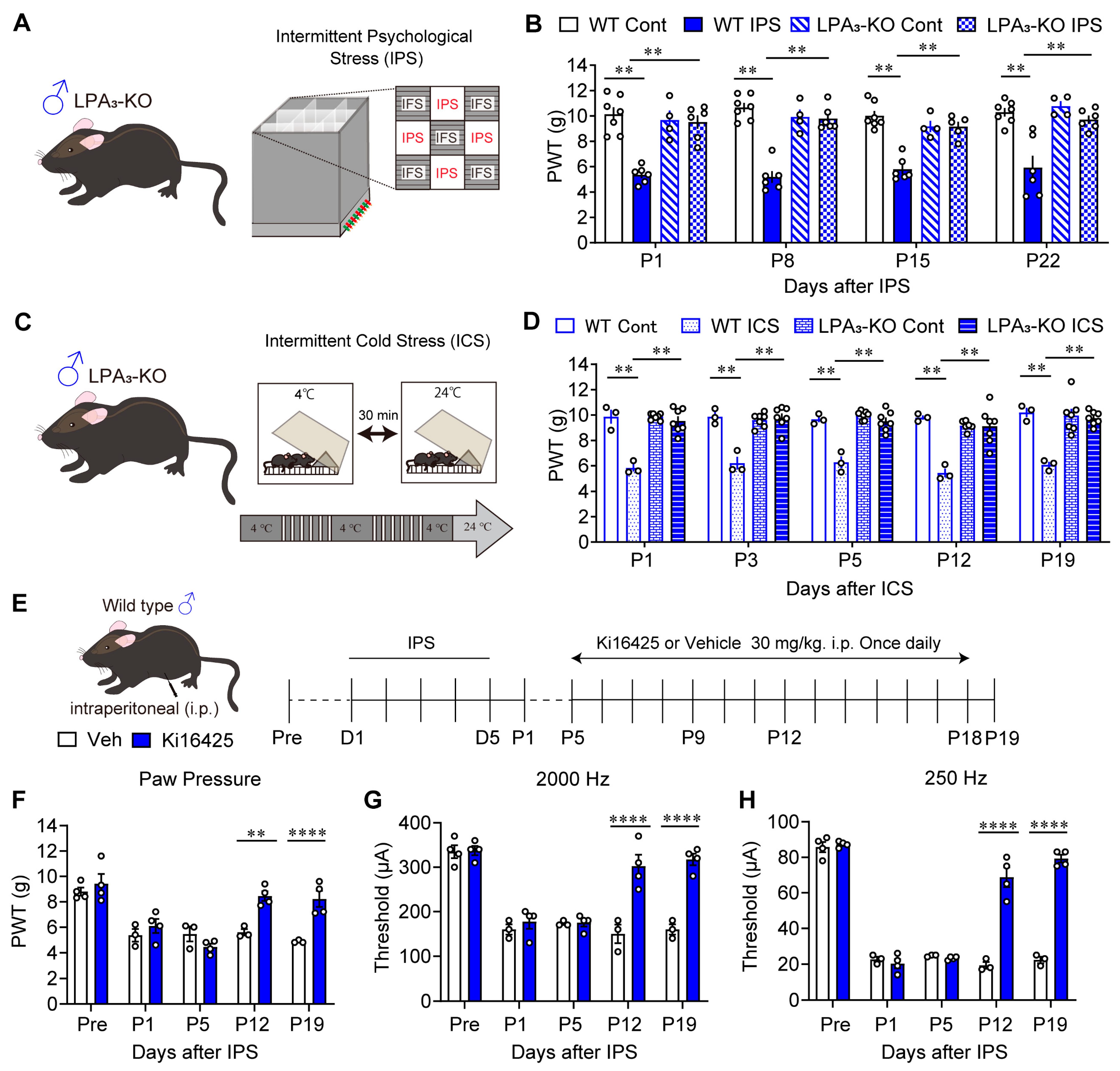
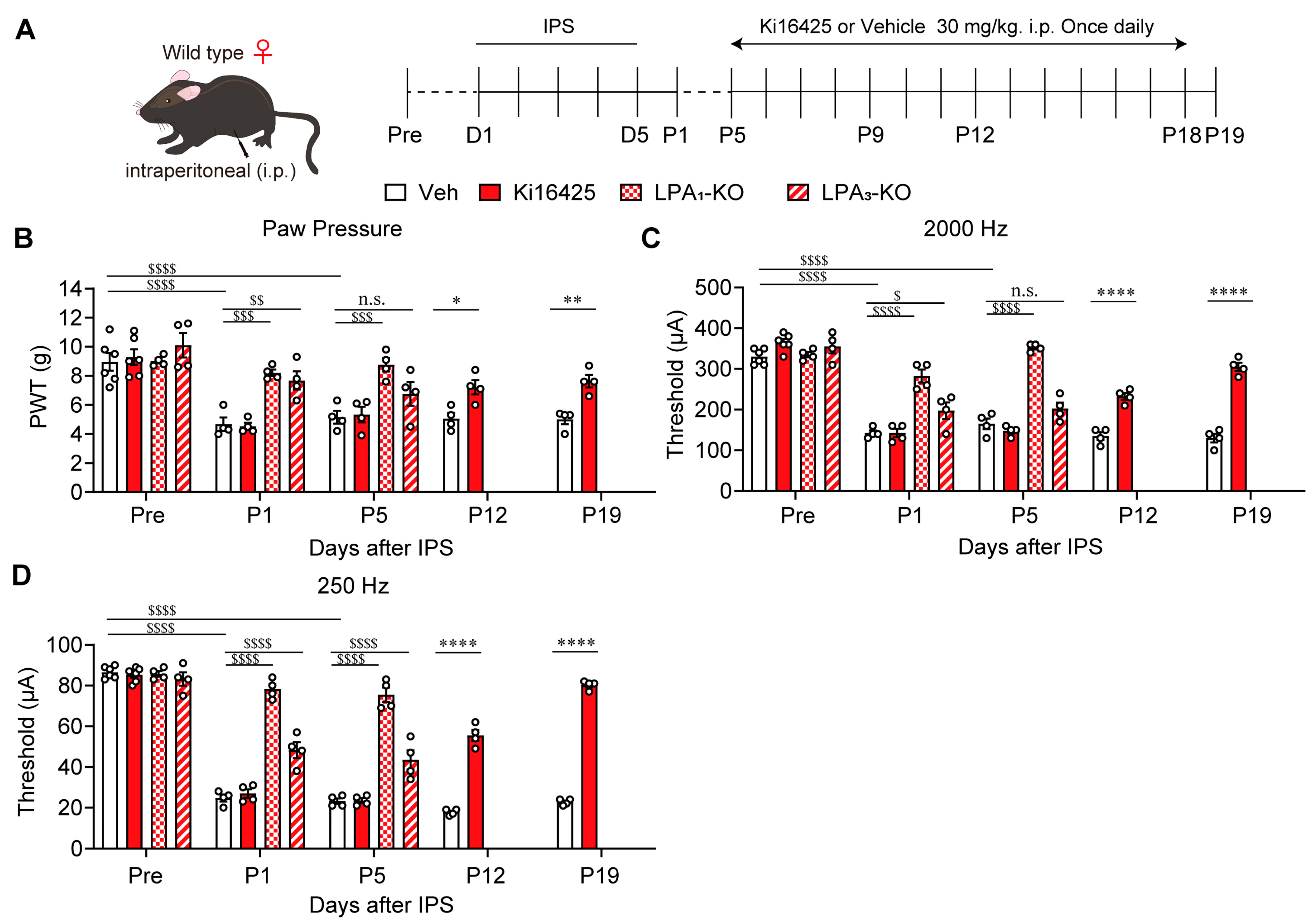
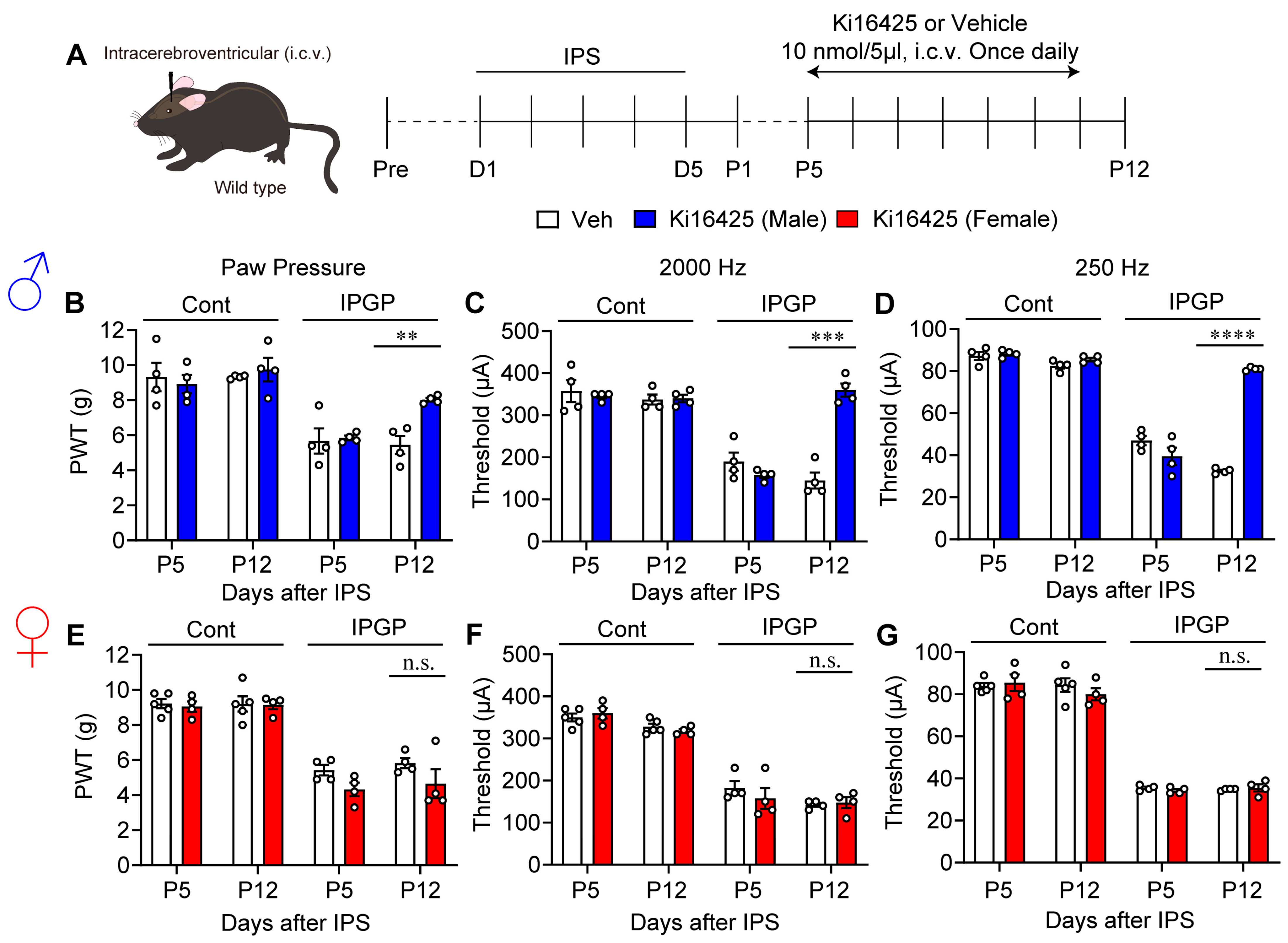
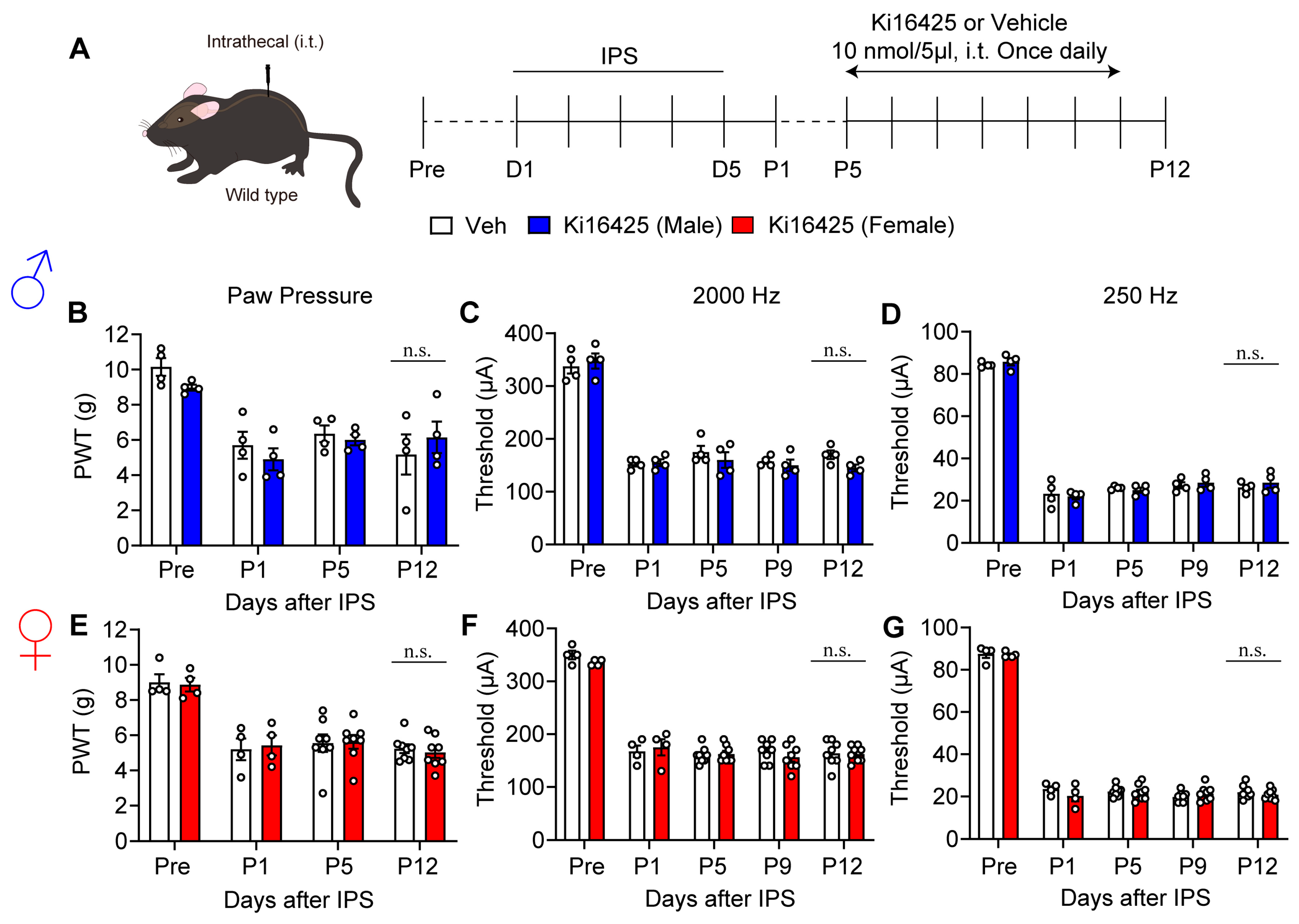
2.2. IPGP and ICGP Model
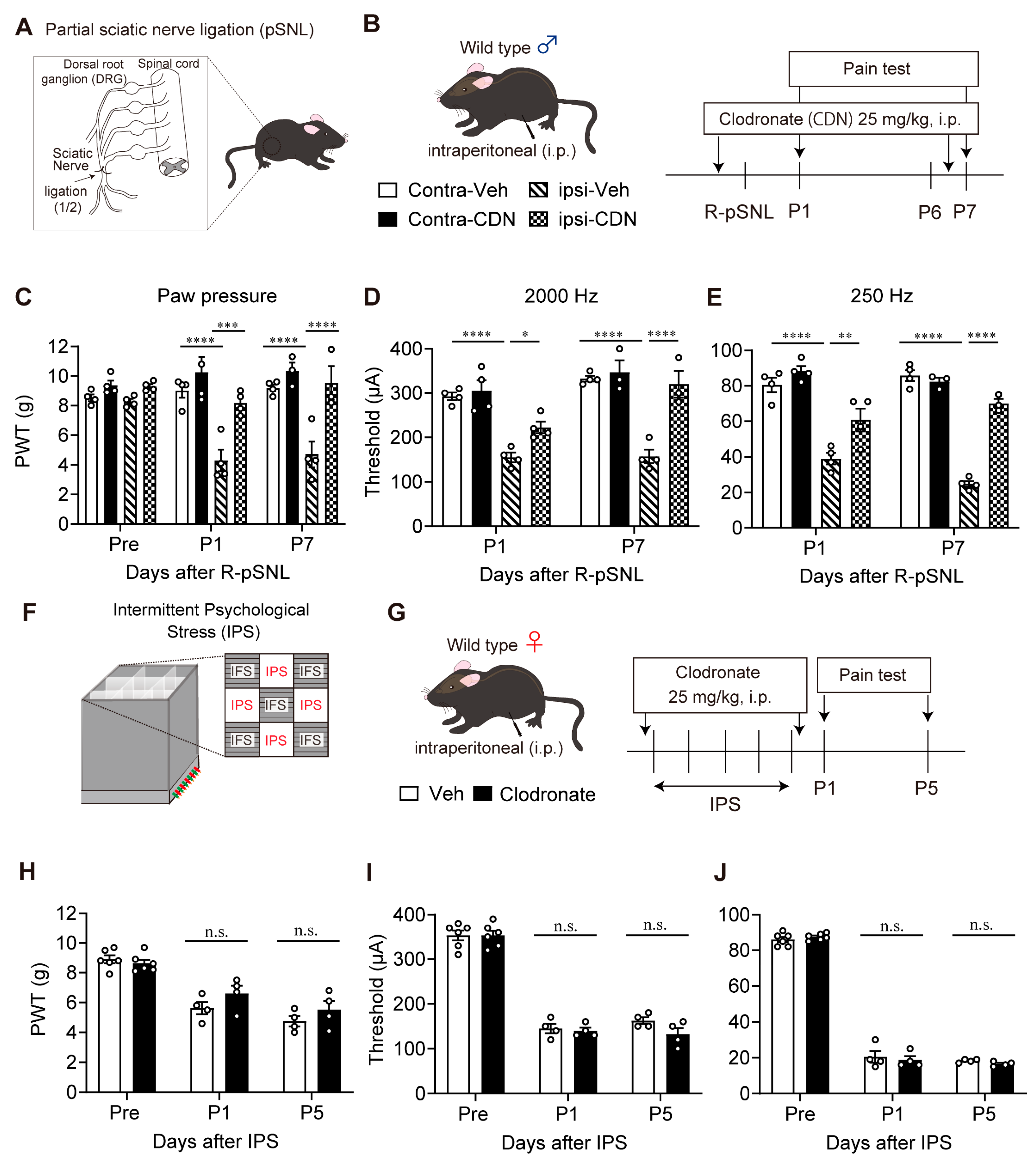
2.3. Partial Sciatic Nerve Ligation (pSNL) Model
2.4. Pain Tests
2.5. Clodronate Liposome Treatments
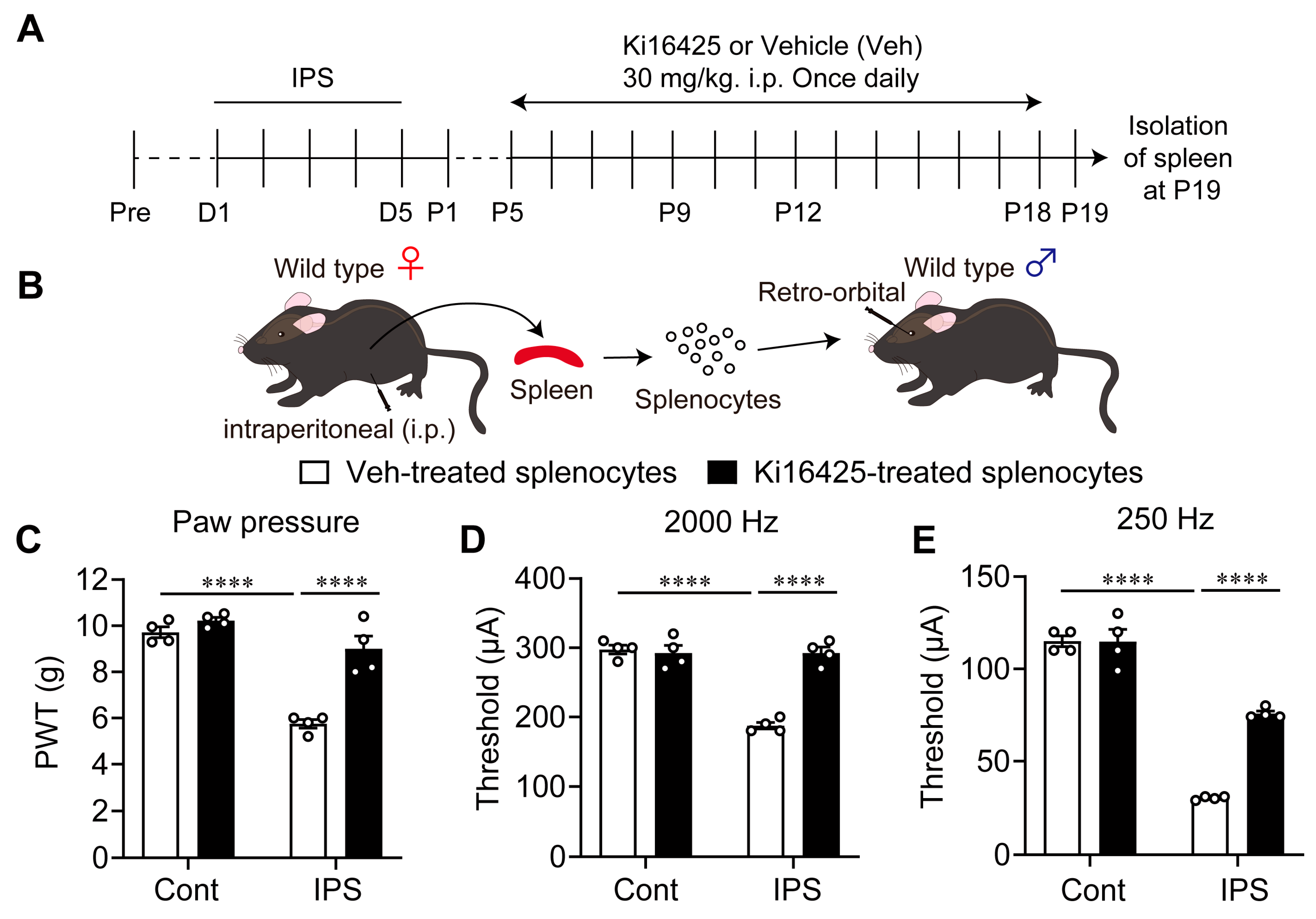
2.6. Isolation and Adoptive Transfer of Splenocytes
2.7. Drug Treatments
2.8. Statistical Analysis
3. Results
3.1. Involvement of LPA3 Signaling in IPGP and ICGP Models in Male Mice
3.2. Involvement of LPA1 and LPA3 Signaling in the IPGP Model in Female Mice
3.3. Sex Differences in the Effect of Intracerebroventricular Administration of Ki16425
3.4. Lack of Blocking Effects on Hyperalgesia and Hypersensitivity by Intrathecal Administration of Ki16425
3.5. Lack of Blocking Effect of Clodronate Liposome on Female IPGP Model
3.6. Blocking Effect of Ki16425 on the Induction of Pain Hypersensitivity by Splenocytes Derived from Female IPGP Model Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FM | Fibromyalgia |
| ICGP | Intermittent Cold Stress-Induced Generalized Pain |
| IPGP | Intermittent Psychological Stress-Induced Generalized Pain |
| LPA | Lysophosphatidic acid |
| NeuP | Neuropathic Pain |
| pSNL | Partial Sciatic Nerve ligation |
| IGGs | Immunoglobulin G |
References
- Fillingim, R.B.; King, C.D.; Ribeiro-Dasilva, M.C.; Rahim-Williams, B.; Riley, J.L., III. Sex, gender, and pain: A review of recent clinical and experimental findings. J. Pain 2009, 10, 447–485. [Google Scholar] [CrossRef]
- Todd, A.; McNamara, C.L.; Balaj, M.; Huijts, T.; Akhter, N.; Thomson, K.; Kasim, A.; Eikemo, T.A.; Bambra, C. The European epidemic: Pain prevalence and socioeconomic inequalities in pain across 19 European countries. Eur. J. Pain 2019, 23, 1425–1436. [Google Scholar] [CrossRef]
- Tsang, A.; Von Korff, M.; Lee, S.; Alonso, J.; Karam, E.; Angermeyer, M.C.; Borges, G.L.; Bromet, E.J.; Demytteneare, K.; de Girolamo, G.; et al. Common chronic pain conditions in developed and developing countries: Gender and age differences and comorbidity with depression-anxiety disorders. J. Pain 2008, 9, 883–891. [Google Scholar] [CrossRef]
- Zimmer, Z.; Fraser, K.; Grol-Prokopczyk, H.; Zajacova, A. A global study of pain prevalence across 52 countries: Examining the role of country-level contextual factors. Pain 2022, 163, 1740–1750. [Google Scholar] [CrossRef]
- Inoue, M.; Rashid, M.H.; Fujita, R.; Contos, J.J.; Chun, J.; Ueda, H. Initiation of neuropathic pain requires lysophosphatidic acid receptor signaling. Nat. Med. 2004, 10, 712–718. [Google Scholar] [CrossRef]
- Ueda, H. Lysophosphatidic acid signaling is the definitive mechanism underlying neuropathic pain. Pain 2017, 158 (Suppl. S1), S55–S65. [Google Scholar] [CrossRef]
- Ueda, H. Pathogenic mechanisms of lipid mediator lysophosphatidic acid in chronic pain. Prog. Lipid Res. 2021, 81, 101079. [Google Scholar] [CrossRef]
- Uranbileg, B.; Ito, N.; Kurano, M.; Saigusa, D.; Saito, R.; Uruno, A.; Kano, K.; Ikeda, H.; Yamada, Y.; Sumitani, M.; et al. Alteration of the lysophosphatidic acid and its precursor lysophosphatidylcholine levels in spinal cord stenosis: A study using a rat cauda equina compression model. Sci. Rep. 2019, 9, 16578. [Google Scholar] [CrossRef]
- Ahn, D.K.; Lee, S.Y.; Han, S.R.; Ju, J.S.; Yang, G.Y.; Lee, M.K.; Youn, D.H.; Bae, Y.C. Intratrigeminal ganglionic injection of LPA causes neuropathic pain-like behavior and demyelination in rats. Pain 2009, 146, 114–120. [Google Scholar] [CrossRef]
- McDougall, J.J.; Albacete, S.; Schuelert, N.; Mitchell, P.G.; Lin, C.; Oskins, J.L.; Bui, H.H.; Chambers, M.G. Lysophosphatidic acid provides a missing link between osteoarthritis and joint neuropathic pain. Osteoarthr. Cartil. 2017, 25, 926–934. [Google Scholar] [CrossRef]
- O’Brien, M.S.; Philpott, H.T.A.; McDougall, J.J. Targeting the Nav1.8 ion channel engenders sex-specific responses in lysophosphatidic acid-induced joint neuropathy. Pain 2019, 160, 269–278. [Google Scholar] [CrossRef]
- Ueda, H.; Neyama, H.; Nagai, J.; Matsushita, Y.; Tsukahara, T.; Tsukahara, R. Involvement of lysophosphatidic acid-induced astrocyte activation underlying the maintenance of partial sciatic nerve injury-induced neuropathic pain. Pain 2018, 159, 2170–2178. [Google Scholar] [CrossRef]
- Edamura, T.; Sumitani, M.; Hayakawa, K.; Inoue, R.; Abe, H.; Tsuchida, R.; Chikuda, H.; Ogata, T.; Kurano, M.; Aoki, J.; et al. Different Profiles of the Triad of Lysophosphatidylcholine, Lysophosphatidic Acid, and Autotaxin in Patients with Neuropathic Pain Diseases: A Preliminary Observational Study. Pain Ther. 2022, 11, 1439–1449. [Google Scholar] [CrossRef]
- Velasco, M.; O’Sullivan, C.; Sheridan, G.K. Lysophosphatidic acid receptors (LPARs): Potential targets for the treatment of neuropathic pain. Neuropharmacology 2017, 113, 608–617. [Google Scholar] [CrossRef]
- Khasar, S.G.; Miao, F.J.; Janig, W.; Levine, J.D. Vagotomy-induced enhancement of mechanical hyperalgesia in the rat is sympathoadrenal-mediated. J. Neurosci. 1998, 18, 3043–3049. [Google Scholar] [CrossRef]
- Sluka, K.A.; Kalra, A.; Moore, S.A. Unilateral intramuscular injections of acidic saline produce a bilateral, long-lasting hyperalgesia. Muscle Nerve 2001, 24, 37–46. [Google Scholar] [CrossRef]
- Nishiyori, M.; Ueda, H. Prolonged gabapentin analgesia in an experimental mouse model of fibromyalgia. Mol. Pain 2008, 4, 52. [Google Scholar] [CrossRef]
- Khasar, S.G.; Dina, O.A.; Green, P.G.; Levine, J.D. Sound stress-induced long-term enhancement of mechanical hyperalgesia in rats is maintained by sympathoadrenal catecholamines. J. Pain 2009, 10, 1073–1077. [Google Scholar] [CrossRef]
- Nagakura, Y.; Oe, T.; Aoki, T.; Matsuoka, N. Biogenic amine depletion causes chronic muscular pain and tactile allodynia accompanied by depression: A putative animal model of fibromyalgia. Pain 2009, 146, 26–33. [Google Scholar] [CrossRef]
- Ueda, H.; Neyama, H. LPA1 receptor involvement in fibromyalgia-like pain induced by intermittent psychological stress, empathy. Neurobiol. Pain 2017, 1, 16–25. [Google Scholar] [CrossRef]
- Ueda, H.; Neyama, H. Fibromyalgia Animal Models Using Intermittent Cold and Psychological Stress. Biomedicines 2023, 12, 56. [Google Scholar] [CrossRef]
- Amaral, R.F.; Geraldo, L.H.M.; Einicker-Lamas, M.; TCLS, E.S.; Mendes, F.; Lima, F.R.S. Microglial lysophosphatidic acid promotes glioblastoma proliferation and migration via LPA1 receptor. J. Neurochem. 2021, 156, 499–512. [Google Scholar] [CrossRef]
- Gaire, B.P.; Choi, J.W. Critical Roles of Lysophospholipid Receptors in Activation of Neuroglia and Their Neuroinflammatory Responses. Int. J. Mol. Sci. 2021, 22, 7864. [Google Scholar] [CrossRef]
- Gaire, B.P.; Sapkota, A.; Song, M.R.; Choi, J.W. Lysophosphatidic acid receptor 1 (LPA1) plays critical roles in microglial activation and brain damage after transient focal cerebral ischemia. J. Neuroinflamm. 2019, 16, 170. [Google Scholar] [CrossRef]
- Kwon, J.H.; Gaire, B.P.; Park, S.J.; Shin, D.Y.; Choi, J.W. Identifying lysophosphatidic acid receptor subtype 1 (LPA1) as a novel factor to modulate microglial activation and their TNF-α production by activating ERK1/2. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2018, 1863, 1237–1245. [Google Scholar] [CrossRef]
- Plastira, I.; Bernhart, E.; Joshi, L.; Koyani, C.N.; Strohmaier, H.; Reicher, H.; Malle, E.; Sattler, W. MAPK signaling determines lysophosphatidic acid (LPA)-induced inflammation in microglia. J. Neuroinflamm. 2020, 17, 127. [Google Scholar] [CrossRef]
- Ueda, H.; Dozono, N.; Tanaka, K.; Kaneko, S.; Neyama, H.; Uchida, H. Allodynia by Splenocytes from Mice with Acid-Induced Fibromyalgia-Like Generalized Pain and Its Sexual Dimorphic Regulation by Brain Microglia. Front. Neurosci. 2020, 14, 600166. [Google Scholar] [CrossRef]
- Boerner, K.E.; Chambers, C.T.; Gahagan, J.; Keogh, E.; Fillingim, R.B.; Mogil, J.S. Conceptual complexity of gender and its relevance to pain. Pain 2018, 159, 2137–2141. [Google Scholar] [CrossRef]
- Mapplebeck, J.C.; Beggs, S.; Salter, M.W. Molecules in pain and sex: A developing story. Mol. Brain 2017, 10, 9. [Google Scholar] [CrossRef]
- Mapplebeck, J.C.S.; Beggs, S.; Salter, M.W. Sex differences in pain: A tale of two immune cells. Pain 2016, 157 (Suppl. S1), S2–S6. [Google Scholar] [CrossRef]
- Mogil, J.S. Qualitative sex differences in pain processing: Emerging evidence of a biased literature. Nat. Rev. Neurosci. 2020, 21, 353–365. [Google Scholar] [CrossRef]
- Sorge, R.E.; LaCroix-Fralish, M.L.; Tuttle, A.H.; Sotocinal, S.G.; Austin, J.S.; Ritchie, J.; Chanda, M.L.; Graham, A.C.; Topham, L.; Beggs, S.; et al. Spinal cord Toll-like receptor 4 mediates inflammatory and neuropathic hypersensitivity in male but not female mice. J. Neurosci. 2011, 31, 15450–15454. [Google Scholar] [CrossRef]
- Sorge, R.E.; Mapplebeck, J.C.; Rosen, S.; Beggs, S.; Taves, S.; Alexander, J.K.; Martin, L.J.; Austin, J.S.; Sotocinal, S.G.; Chen, D.; et al. Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Nat. Neurosci. 2015, 18, 1081–1083. [Google Scholar] [CrossRef]
- Contos, J.J.; Fukushima, N.; Weiner, J.A.; Kaushal, D.; Chun, J. Requirement for the lpA1 lysophosphatidic acid receptor gene in normal suckling behavior. Proc. Natl. Acad. Sci. USA 2000, 97, 13384–13389. [Google Scholar] [CrossRef]
- Ye, X.; Hama, K.; Contos, J.J.; Anliker, B.; Inoue, A.; Skinner, M.K.; Suzuki, H.; Amano, T.; Kennedy, G.; Arai, H.; et al. LPA3-mediated lysophosphatidic acid signalling in embryo implantation and spacing. Nature 2005, 435, 104–108. [Google Scholar] [CrossRef]
- du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. Br. J. Pharmacol. 2020, 177, 3617–3624. [Google Scholar] [CrossRef]
- Koga, K.; Furue, H.; Rashid, M.H.; Takaki, A.; Katafuchi, T.; Yoshimura, M. Selective activation of primary afferent fibers evaluated by sine-wave electrical stimulation. Mol. Pain 2005, 1, 13. [Google Scholar] [CrossRef]
- Nguyen, T.; Du, J.; Li, Y.C. A protocol for macrophage depletion and reconstitution in a mouse model of sepsis. STAR Protoc. 2021, 2, 101004. [Google Scholar] [CrossRef]
- Yamashita, S.; Dozono, N.; Tobori, S.; Nagayasu, K.; Kaneko, S.; Shirakawa, H.; Ueda, H. Peripheral Beta-2 Adrenergic Receptors Mediate the Sympathetic Efferent Activation from Central Nervous System to Splenocytes in a Mouse Model of Fibromyalgia. Int. J. Mol. Sci. 2023, 24, 3465. [Google Scholar] [CrossRef]
- Yardeni, T.; Eckhaus, M.; Morris, H.D.; Huizing, M.; Hoogstraten-Miller, S. Retro-orbital injections in mice. Lab Anim. 2011, 40, 155–160. [Google Scholar] [CrossRef]
- University Animal Care Committee Standard Operating Procedure. Available online: https://www.queensu.ca/animals-in-science/policies-procedures/sop/mice/7-11 (accessed on 3 July 2025).
- Haley, T.J.; McCormick, W.G. Pharmacological effects produced by intracerebral injection of drugs in the conscious mouse. Br. J. Pharmacol. Chemother. 1957, 12, 12–15. [Google Scholar] [CrossRef]
- Hylden, J.L.; Wilcox, G.L. Intrathecal morphine in mice: A new technique. Eur. J. Pharmacol. 1980, 67, 313–316. [Google Scholar] [CrossRef]
- Matsumoto, M.; Xie, W.; Ma, L.; Ueda, H. Pharmacological switch in Aβ-fiber stimulation-induced spinal transmission in mice with partial sciatic nerve injury. Mol. Pain 2008, 4, 25. [Google Scholar] [CrossRef]
- Ma, L.; Nagai, J.; Ueda, H. Microglial activation mediates de novo lysophosphatidic acid production in a model of neuropathic pain. J. Neurochem. 2010, 115, 643–653. [Google Scholar] [CrossRef]
- Kiguchi, N.; Kobayashi, D.; Saika, F.; Matsuzaki, S.; Kishioka, S. Pharmacological Regulation of Neuropathic Pain Driven by Inflammatory Macrophages. Int. J. Mol. Sci. 2017, 18, 2296. [Google Scholar] [CrossRef]
- Liu, T.; van Rooijen, N.; Tracey, D.J. Depletion of macrophages reduces axonal degeneration and hyperalgesia following nerve injury. Pain 2000, 86, 25–32. [Google Scholar] [CrossRef]
- Ellergezen, P.; Alp, A.; Cavun, S.; Celebi, M.; Macunluoglu, A.C. Pregabalin inhibits proinflammatory cytokine release in patients with fibromyalgia syndrome. Arch. Rheumatol. 2023, 38, 307–314. [Google Scholar] [CrossRef]
- Gerdle, B.; Leinhard, O.D.; Lund, E.; Lundberg, P.; Forsgren, M.F.; Ghafouri, B. Pain and the biochemistry of fibromyalgia: Patterns of peripheral cytokines and chemokines contribute to the differentiation between fibromyalgia and controls and are associated with pain, fat infiltration and content. Front. Pain Res. 2024, 5, 1288024. [Google Scholar] [CrossRef]
- Peck, M.M.; Maram, R.; Mohamed, A.; Crespo, D.O.; Kaur, G.; Ashraf, I.; Malik, B.H. The Influence of Pro-inflammatory Cytokines and Genetic Variants in the Development of Fibromyalgia: A Traditional Review. Cureus 2020, 12, e10276. [Google Scholar] [CrossRef]
- Goebel, A.; Krock, E.; Gentry, C.; Israel, M.R.; Jurczak, A.; Urbina, C.M.; Sandor, K.; Vastani, N.; Maurer, M.; Cuhadar, U.; et al. Passive transfer of fibromyalgia symptoms from patients to mice. J. Clin. Investig. 2021, 131, e144201. [Google Scholar] [CrossRef]
- Fanton, S.; Menezes, J.; Krock, E.; Sandstrom, A.; Tour, J.; Sandor, K.; Jurczak, A.; Hunt, M.; Baharpoor, A.; Kadetoff, D.; et al. Anti-satellite glia cell IgG antibodies in fibromyalgia patients are related to symptom severity and to metabolite concentrations in thalamus and rostral anterior cingulate cortex. Brain Behav. Immun. 2023, 114, 371–382. [Google Scholar] [CrossRef]
- Jakobsson, J.E.; Menezes, J.; Krock, E.; Hunt, M.A.; Carlsson, H.; Vaivade, A.; Khoonsari, P.E.; Agalave, N.M.; Sandstrom, A.; Kadetoff, D.; et al. Fibromyalgia patients have altered lipid concentrations associated with disease symptom severity and anti-satellite glial cell IgG antibodies. J. Pain 2025, 29, 105331. [Google Scholar] [CrossRef]
- Krock, E.; Morado-Urbina, C.E.; Menezes, J.; Hunt, M.A.; Sandstrom, A.; Kadetoff, D.; Tour, J.; Verma, V.; Kultima, K.; Haglund, L.; et al. Fibromyalgia patients with elevated levels of anti-satellite glia cell immunoglobulin G antibodies present with more severe symptoms. Pain 2023, 164, 1828–1840. [Google Scholar] [CrossRef]
- Fan, C.Y.; McAllister, B.B.; Stokes-Heck, S.; Harding, E.K.; de Vasconcelos, A.P.; Mah, L.K.; Lima, L.V.; van den Hoogen, N.J.; Rosen, S.F.; Ham, B.; et al. Divergent sex-specific pannexin-1 mechanisms in microglia and T cells underlie neuropathic pain. Neuron 2025, 113, 896–911.e899. [Google Scholar] [CrossRef]
- Lister, K.C.; Wong, C.; Cai, W.; Uttam, S.; Stecum, P.; Rodrigues, R.; Hooshmandi, M.; Brown, N.; Fan, J.; Francois-Saint-Cyr, N.; et al. 4E-BP1-dependent translation in microglia controls mechanical hypersensitivity in male and female mice. J. Clin. Investig. 2025, 135, e180190. [Google Scholar] [CrossRef]
- Saika, F.; Sato, T.; Nakabayashi, T.; Fukazawa, Y.; Hino, S.; Suzuki, K.; Kiguchi, N. Male-Dominant Spinal Microglia Contribute to Neuropathic Pain by Producing CC-Chemokine Ligand 4 Following Peripheral Nerve Injury. Cells 2025, 14, 484. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IPS Model | Sex | Hyperalgesia (Paw Pressure) | Hypersensitivity (2000 Hz) | Hypersensitivity (250 Hz) |
|---|---|---|---|---|
| LPA1-KO | Male | − (1) | No data | No data |
| Female | − | − | − | |
| LPA3-KO | Male | − (2) | No data | No data |
| Female | − | − | − | |
| WT Ki 16425, i.p. | Male | − | − | − |
| Female | − | − | − | |
| WT Ki 1642, i.c.v. | Male | − | − | − |
| Female | + | + | + | |
| WT Ki 16425, i.t. | Male | + | + | + |
| Female | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ueda, H.; Neyama, H.; Dozono, N.; Aoki, J.; Chun, J. Hyperalgesia in the Psychological Stress-Induced Fibromyalgia Model Shows Sexual Dimorphism Mediated by LPA1 and LPA3. Cells 2025, 14, 1022. https://doi.org/10.3390/cells14131022
Ueda H, Neyama H, Dozono N, Aoki J, Chun J. Hyperalgesia in the Psychological Stress-Induced Fibromyalgia Model Shows Sexual Dimorphism Mediated by LPA1 and LPA3. Cells. 2025; 14(13):1022. https://doi.org/10.3390/cells14131022
Chicago/Turabian StyleUeda, Hiroshi, Hiroyuki Neyama, Naoki Dozono, Junken Aoki, and Jerold Chun. 2025. "Hyperalgesia in the Psychological Stress-Induced Fibromyalgia Model Shows Sexual Dimorphism Mediated by LPA1 and LPA3" Cells 14, no. 13: 1022. https://doi.org/10.3390/cells14131022
APA StyleUeda, H., Neyama, H., Dozono, N., Aoki, J., & Chun, J. (2025). Hyperalgesia in the Psychological Stress-Induced Fibromyalgia Model Shows Sexual Dimorphism Mediated by LPA1 and LPA3. Cells, 14(13), 1022. https://doi.org/10.3390/cells14131022